
Phylogenetic and Functional Diversity of Saprolegniales and Fungi Isolated from Temperate Lakes in Northeast Germany

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Oomycetes and Fungal Strains
2.2. Molecular Characterization of the Strains
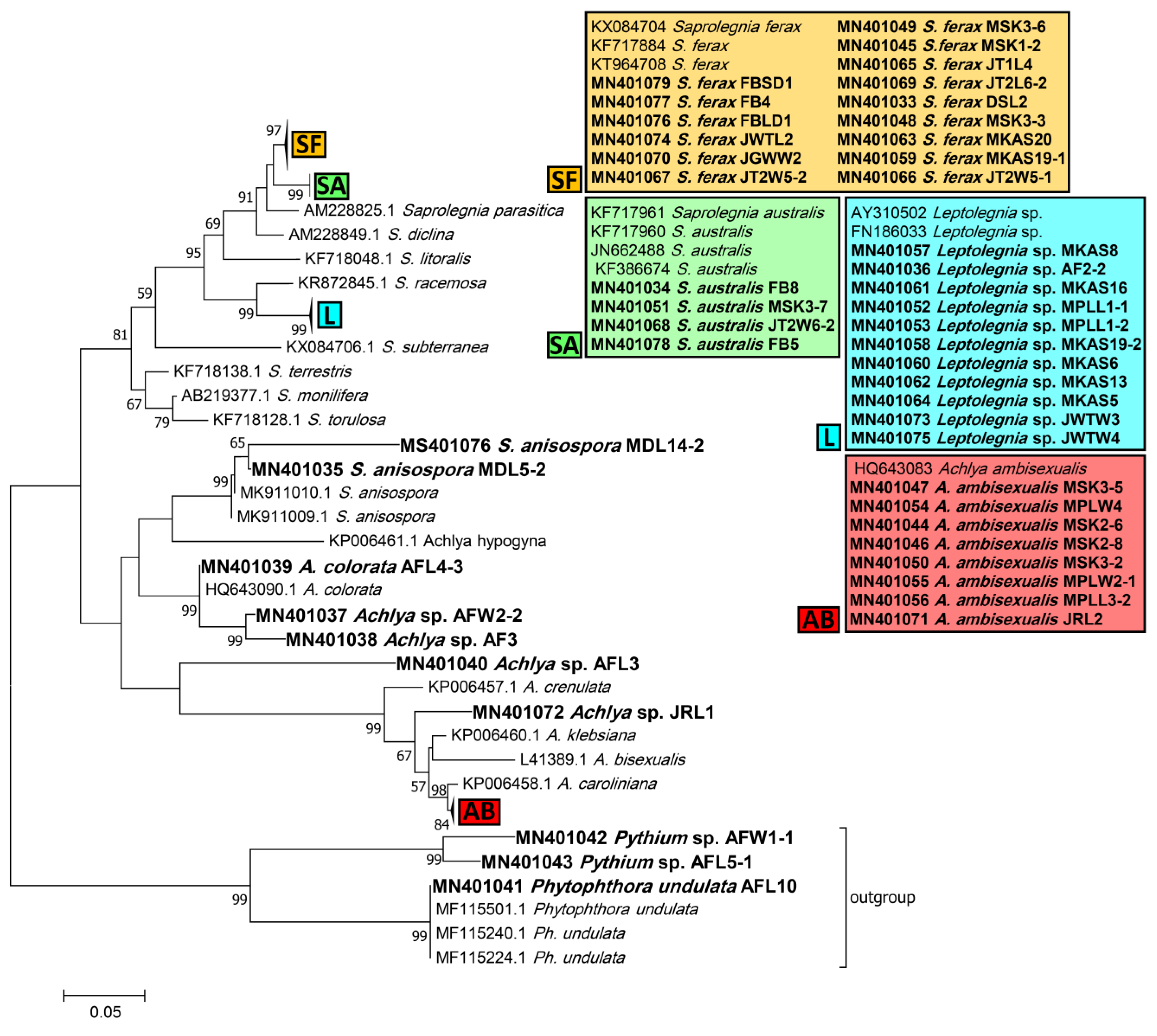
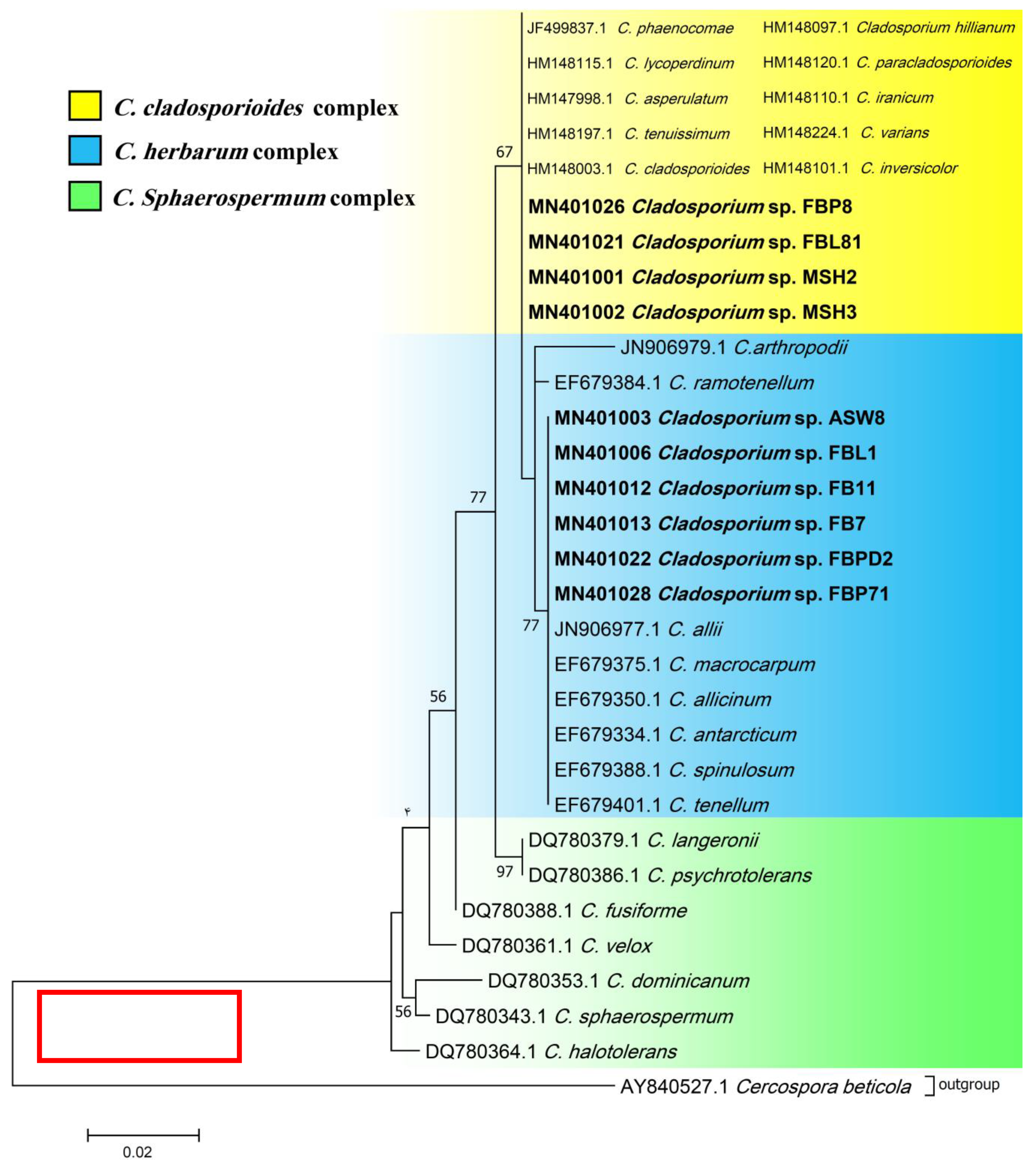
2.3. Phylogenetic Analysis
2.4. Screening for Lignino-, Cellulo-, and Chitinolytic Activity
2.4.1. Ligninolytic Activity
2.4.2. Cellulolytic Activity
2.4.3. Chitinolytic Activity
2.5. Physiological Profiling Based on 95 Labile Carbon Sources
2.6. DOM Utilization and HS Degradation and Production
2.7. Statistical Analysis
3. Results
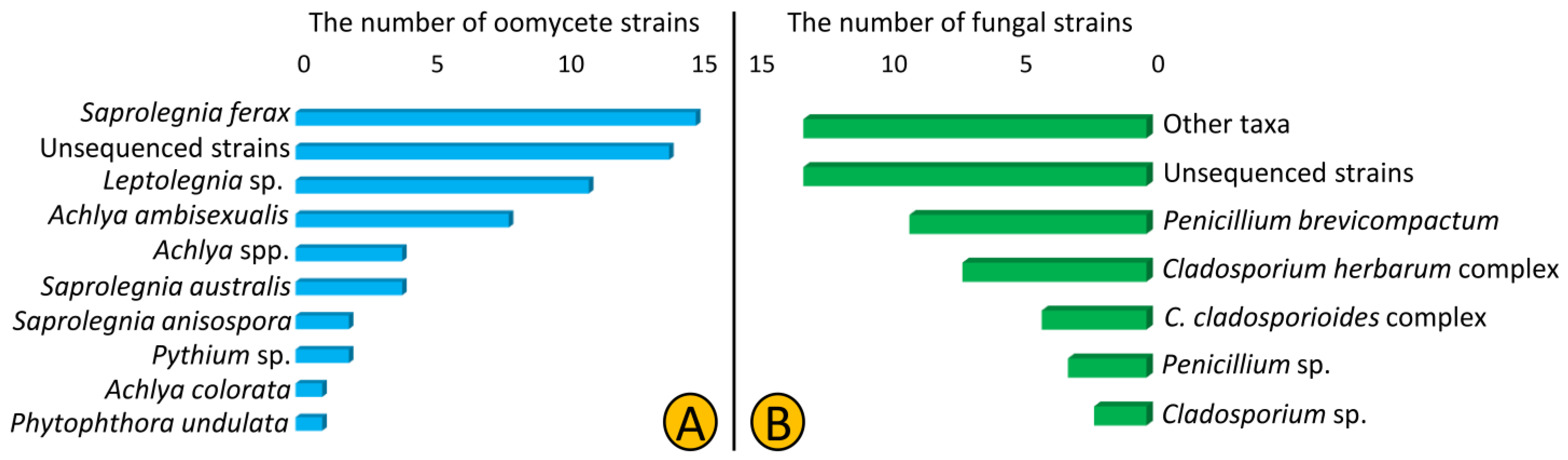
3.1. Isolation and Identification
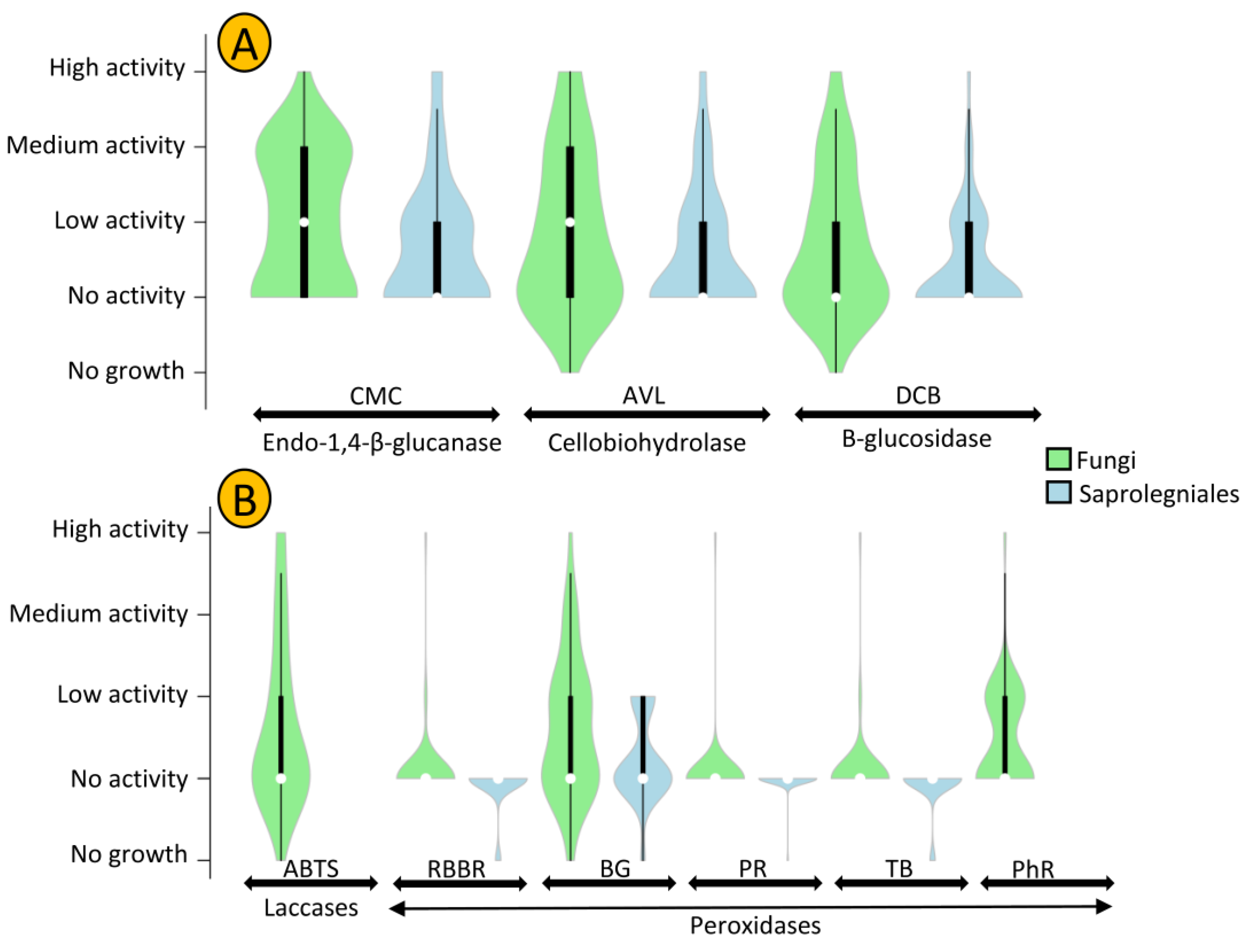
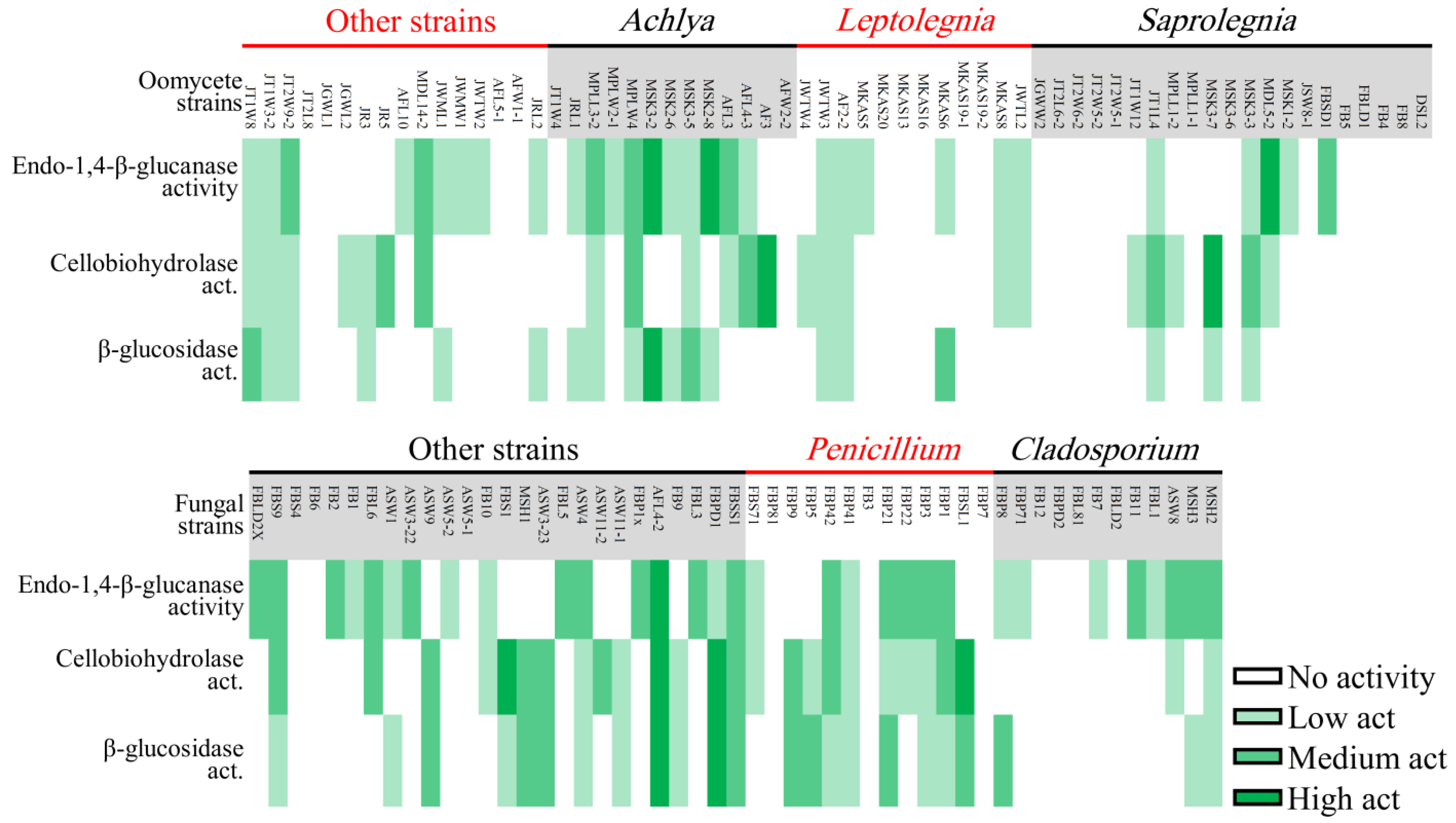
3.2. Ligninolytic, Cellulolytic, and Chitinolytic Activity
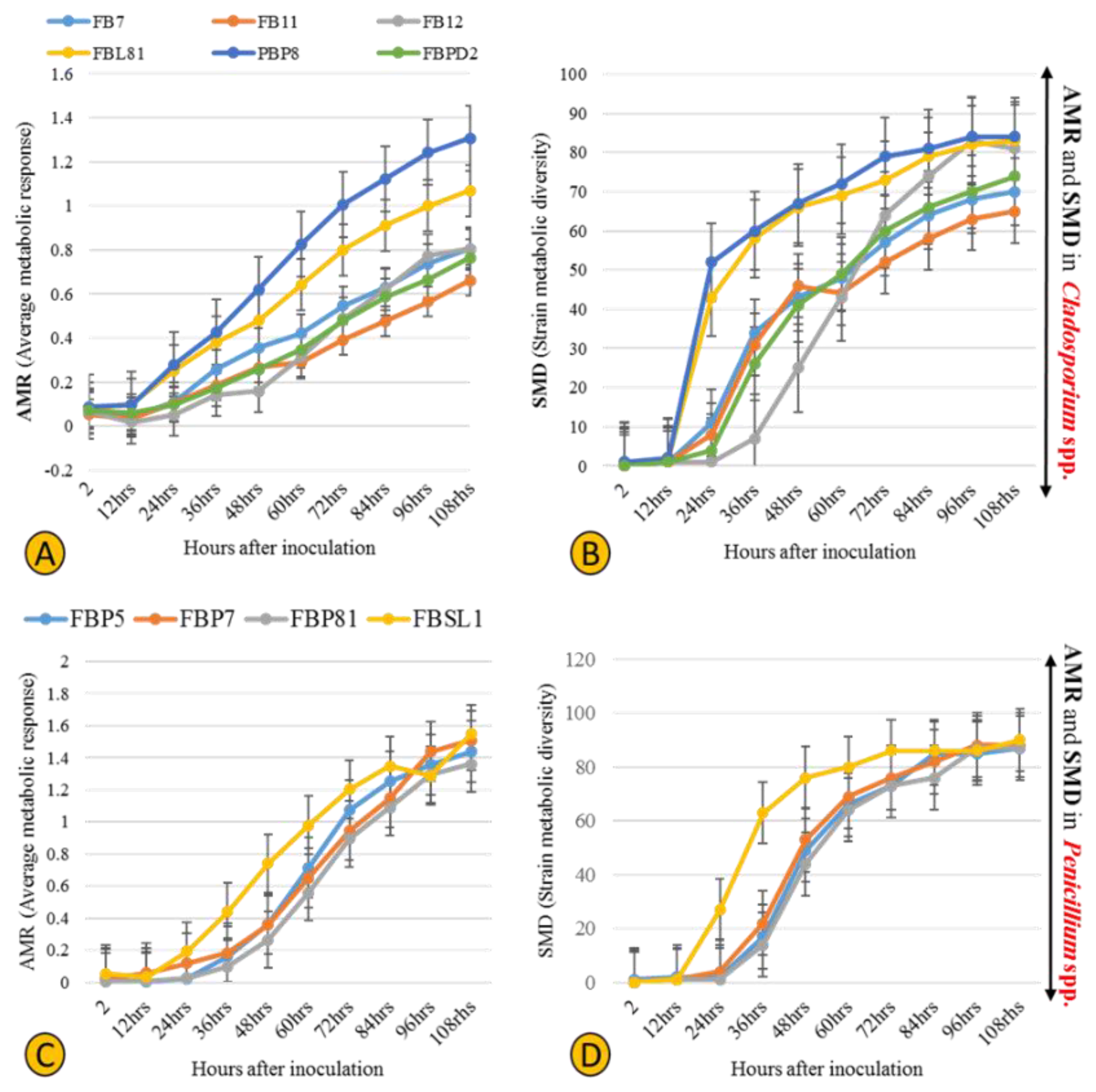
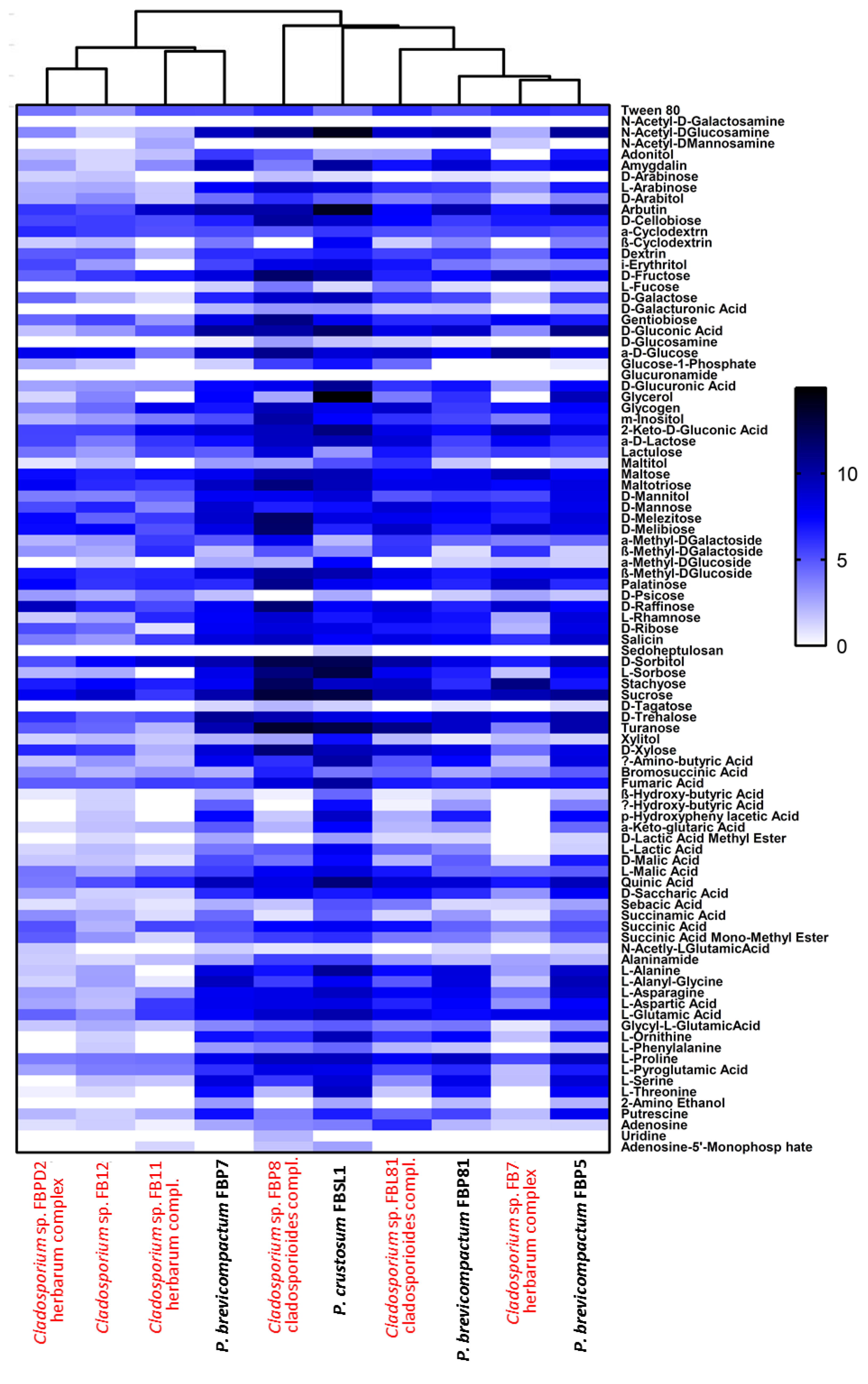
3.3. Physiological Profiling of Strains Based on Utilization of 95 Labile Carbon Sources
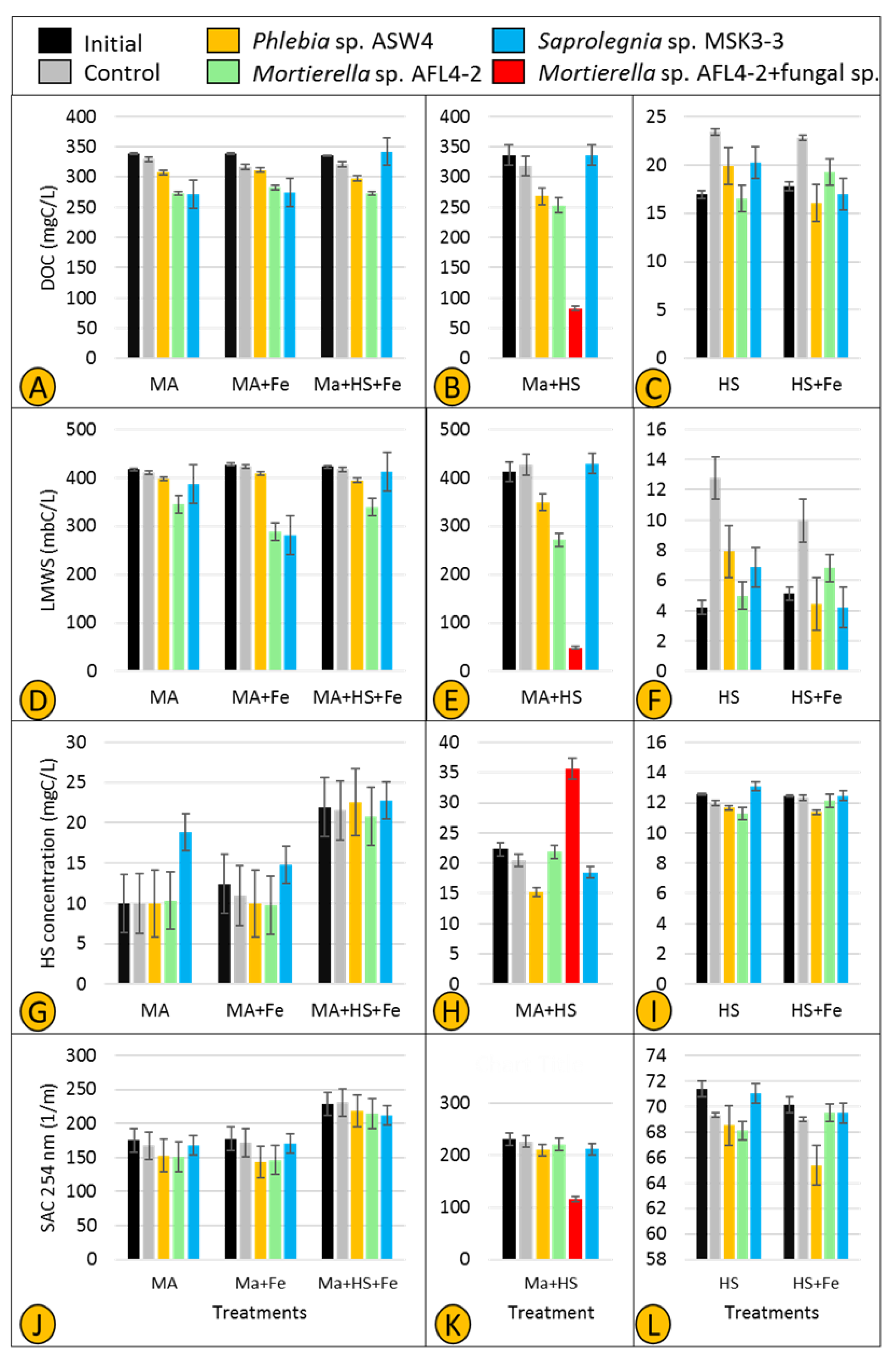
3.4. DOM Utilization and HS Degradation/Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stedmon, C.A.; Markager, S.; Bro, R. Tracing dissolved organic matter in aquatic environments using a new approach to fluorescence spectroscopy. Mar. Chem. 2003, 82, 239–254. [Google Scholar] [CrossRef]
- Mattsson, T.; Kortelainen, P.; Räike, A. Export of DOM from Boreal Catchments: Impacts of Land Use Cover and Climate. Biogeochemistry 2005, 76, 373–394. [Google Scholar] [CrossRef]
- McDonald, S.; Bishop, A.G.; Prenzler, P.D.; Robards, K. Analytical chemistry of freshwater humic substances. Anal. Chim. Acta 2004, 527, 105–124. [Google Scholar] [CrossRef]
- Sime-Ngando, T.; Lefèvre, E.; Gleason, F.H. Hidden diversity among aquatic heterotrophic flagellates: Ecological potentials of zoosporic fungi. Hydrobiologia 2011, 659, 5–22. [Google Scholar] [CrossRef]
- Romani, A.M.; Fischer, H.; Mille-Lindblom, C.; Tranvik, L.J. Interactions of bacteria and fungi on decomposing litter: Differential extracellular enzyme activities. Ecology 2006, 87, 2559–2569. [Google Scholar] [CrossRef] [Green Version]
- Klein, O.I.; Isakova, E.P.; Deryabina, Y.I.; Kulikova, N.A.; Badun, G.A.; Chernysheva, M.G.; Stepanova, E.V.; Koroleva, O.V. Humic Substances Enhance Growth and Respiration in the Basidiomycetes Trametes Maxima Under Carbon Limited Conditions. J. Chem. Ecol. 2014, 40, 643–652. [Google Scholar] [CrossRef]
- Jobard, M.; Rasconi, S.; Sime-Ngando, T. Diversity and functions of microscopic fungi: A missing component in pelagic food webs. Aquat. Sci. 2010, 72, 255–268. [Google Scholar] [CrossRef]
- Claus, H.; Filip, Z. Degradation and Transformation of Aquatic Humic Substances by Laccase-producing Fungi Cladosporium cladosporioides and Polyporus versicolor. Acta Hydrochim. Hydrobiol. 1998, 26, 180–185. [Google Scholar] [CrossRef]
- Rojas-Jimenez, K.; Fonvielle, J.A.; Ma, H.; Grossart, H.-P. Transformation of humic substances by the freshwater Ascomycete Cladosporium sp. Limnol. Oceanogr. 2017, 62, 1955–1962. [Google Scholar] [CrossRef]
- Collado, S.; Oulego, P.; Suárez-Iglesias, O.; Díaz, M. Biodegradation of dissolved humic substances by fungi. Appl. Microbiol. Biotechnol. 2018, 102, 3497–3511. [Google Scholar] [CrossRef]
- van West, P. Saprolegnia parasitica, an oomycete pathogen with a fishy appetite: New challenges for an old problem. Mycologist 2006, 20, 99–104. [Google Scholar] [CrossRef]
- Svoboda, J.; Mrugała, A.; Kozubíková-Balcarová, E.; Petrusek, A. Hosts and transmission of the crayfish plague pathogen Aphanomyces astaci: A review. J. Fish Dis. 2016, 40, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Molloy, D.P.; Glockling, S.L.; Siegfried, C.A.; Beakes, G.W.; James, T.Y.; Mastitsky, S.E.; Wurdak, E.; Giamberini, L.; Gaylo, M.J.; Nemeth, M.J. Aquastella gen. nov.: A new genus of saprolegniaceous oomycete rotifer parasites related to Aphanomyces, with unique sporangial outgrowths. Fungal Biol. 2014, 118, 544–558. [Google Scholar] [CrossRef]
- Czeczuga, B.; Mazalska, B.; Godlewska, A.; Muszynska, E. Aquatic fungi growing on dead fragments of submerged plants. Limnologica 2005, 35, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Unestam, T. Chitinolytic, Cellulolytic, and Pectinolytic Activity in vitro of Some Parasitic and Saprophytic Oomycctes. Physiol. Plant. 1966, 19, 15–30. [Google Scholar] [CrossRef]
- Berner, K.E.; Chapman, E.S. The cellulolytic activity of six Oomycetes. Mycologia 1977, 69, 1232–1236. [Google Scholar] [CrossRef]
- Masigol, H.; Khodaparast, S.A.; Woodhouse, J.N.; Rojas-Jimenez, K.; Fonvielle, J.; Rezakhani, F.; Mostowfizadeh-Ghalamfarsa, R.; Neubauer, D.; Goldhammer, T.; Grossart, H. The contrasting roles of aquatic fungi and oomycetes in the degradation and transformation of polymeric organic matter. Limnol. Oceanogr. 2019, 64, 2662–2678. [Google Scholar] [CrossRef] [Green Version]
- Seymour, R.L. The genus Saprolegnia. Nova Hedwigia 1970, 19, 1–124. [Google Scholar]
- Siepmann, R.; Johnson, T.W. Isolation and culture of fungi from wood submerged in saline and fresh waters. J. Elisha Mitchell Sci. Soc. 1960, 76, 150–154. [Google Scholar]
- Montero-Pau, J.; Gomez, A.; Munoz, J. Application of an inexpensive and high-throughput genomic DNA extraction method for the molecular ecology of zooplanktonic diapausing eggs. Limnol. Oceanogr. Methods 2008, 6, 218–222. [Google Scholar] [CrossRef] [Green Version]
- Borneman, J.; Hartin, R.J. PCR Primers That Amplify Fungal rRNA Genes from Environmental Samples. Appl. Environ. Microbiol. 2000, 66, 4356–4360. [Google Scholar] [CrossRef] [Green Version]
- White, T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Innis, M.A., Gelfand, D.H., Sininsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Stielow, J.B.; Lévesque, C.A.; Seifert, K.A.; Meyer, W.; Irinyi, L.; Smits, D.; Renfurm, R.; Verkley, G.J.M.; Groenewald, M.; Chaduli, D.; et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Pers. Mol. Phylogeny Evol. Fungi 2015, 35, 242–263. [Google Scholar] [CrossRef] [Green Version]
- Robideau, G.P.; De Cock, A.W.A.M.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pointing, S.B. Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Fungal Divers. 1999, 2, 17–33. [Google Scholar]
- Agrawal, T.; Kotasthane, A.S. Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India. SpringerPlus 2012, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huber, S.A.; Balz, A.; Abert, M.; Pronk, W. Characterisation of aquatic humic and non-humic matter with size-exclusion chromatography—organic carbon detection—organic nitrogen detection (LC-OCD-OND). Water Res. 2011, 45, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Dziallas, C.; Zwirnmann, E.; Gjessing, E.T.; Grossart, H.-P. UV irradiation of natural organic matter (NOM): Impact on organic carbon and bacteria. Aquat. Sci. 2011, 74, 443–454. [Google Scholar] [CrossRef]
- Miki, T.; Yokokawa, T.; Ke, P.-J.; Hsieh, I.-F.; Hsieh, C.-H.; Kume, T.; Yoneya, K.; Matsui, K. Statistical recipe for quantifying microbial functional diversity from EcoPlate metabolic profiling. Ecol. Res. 2017, 33, 249–260. [Google Scholar] [CrossRef]
- Liu, Y.; Kurtán, T.; Wang, C.Y.; Lin, W.H.; Orfali, R.; Mueller, W.E.; Daletos, G. Proksch, P. Cladosporinone, a new viridi-toxin derivative from the hypersaline lake derived fungus Cladosporium cladosporioides. J. Antibiot. Res. 2016, 69, 702–706. [Google Scholar] [CrossRef]
- Zalar, P.D.; de Hoog, G.S.; Schroers, H.-J.; Crous, P.W.; Groenewald, J.Z.; Gunde-Cimerman, N. Phylogeny and ecology of the ubiquitous saprobe Cladosporium sphaerospermum, with descriptions of seven new species from hypersaline environments. Stud. Mycol. 2007, 58, 157–183. [Google Scholar] [CrossRef] [PubMed]
- Shearer, C.A. The freshwater ascomycetes. Nova Hedwig. 1993, 56, 1–33. [Google Scholar]
- Rocha, S.C.; Lopez-Lastra, C.C.; Marano, A.V.; de Souza, J.I.; Rueda-Páramo, M.E.; Pires-Zottarelli, C.L. New phylogenetic insights into Saprolegniales (Oomycota, Straminipila) based upon studies of specimens isolated from Brazil and Argentina. Mycol. Prog. 2018, 17, 691–700. [Google Scholar] [CrossRef]
- Roberts, R.E. A study of the distribution of certain members of the saprolegniales. Trans. Br. Mycol. Soc. 1963, 46, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.B.; Goodell, B. Mechanisms of wood degradation by brown-rot fungi: Chelator-mediated cellulose degradation and binding of iron by cellulose. J. Biotechnol. 2001, 87, 43–57. [Google Scholar] [CrossRef]
- Varnaitė, R.; Paškevičius, A.; Raudonienė, V. Cellulose degradation in rye straw by micromycetes and their complexes. Ekologija 2008, 54, 29–31. [Google Scholar] [CrossRef]
- Koechli, C.; Campbell, A.N.; Pepe-Ranney, C.; Buckley, D.H. Assessing fungal contributions to cellulose degradation in soil by using high-throughput stable isotope probing. Soil Biol. Biochem. 2019, 130, 150–158. [Google Scholar] [CrossRef]
- Dong, Y.-C.; Wang, W.; Hu, Z.-C.; Fu, M.-L.; Chen, Q.-H. The synergistic effect on production of lignin-modifying enzymes through submerged co-cultivation of Phlebia radiata, Dichomitus squalens and Ceriporiopsis subvermispora using agricultural residues. Bioprocess Biosyst. Eng. 2011, 35, 751–760. [Google Scholar] [CrossRef]
- Thompstone, A.; Dix, N. Cellulase activity in the Saprolegniaceae. Trans. Br. Mycol. Soc. 1985, 85, 361–366. [Google Scholar] [CrossRef]
- Masigol, H.; Khodaparast, S.A.; Mostowfizadeh-Ghalamfarsa, R.; Rojas-Jimenez, K.; Woodhouse, J.N.; Neubauer, D.; Grossart, H.-P. Taxonomical and functional diversity of Saprolegniales in Anzali lagoon, Iran. Aquat. Ecol. 2020, 54, 323–336. [Google Scholar] [CrossRef] [Green Version]
- Järvinen, J.; Taskila, S.; Isomäki, R.; Ojamo, H. Screening of white-rot fungi manganese peroxidases: A comparison between the specific activities of the enzyme from different native producers. AMB Express 2012, 2, 62. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Olson, Å.; Wu, M.; Broberg, A.; Sandgren, M. Biochemical studies of two lytic polysaccharide monooxygenases from the white-rot fungus Heterobasidion irregulare and their roles in lignocellulose degradation. PLoS ONE 2017, 12, e0189479. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Mori, T.; Kamei, I.; Kondo, R. A novel metabolic pathway for biodegradation of DDT by the white rot fungi, Phlebia lindtneri and Phlebia brevispora. Biogeochemistry 2010, 22, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Faraco, V. Induction and Transcriptional Regulation of Laccases in Fungi. Curr. Genom. 2011, 12, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.Z.; Li, P.Q. Genome-wide identification of laccase gene family in three Phytophthora species. Genetica 2012, 140, 477–484. [Google Scholar] [CrossRef]
- Feng, B.Z.; Li, P.Q. Cloning and expression of a novel laccase gene from Phytophthora capsici. J. Plant Pathol. 2013, 95, 417–421. [Google Scholar]
- Feng, B.Z.; Li, P.Q. Cloning, characterization and expression of a novel laccase gene Pclac2 from Phytophthora capsici. Braz. J. Microbiol. 2014, 45, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.Z.; Li, P.Q.; Fu, L.; Yu, X.M. Exploring laccase genes from plant pathogen genomes: A bioinformatic approach. Genet. Mol. Res. 2015, 14, 14019–14036. [Google Scholar] [CrossRef]
- Luis, P.; Gauthier, A.; Trouvelot, S.; Poinssot, B.; Frettinger, P. Identification of Plasmopara viticola Genes Potentially Involved in Pathogenesis on Grapevine Suggests New Similarities Between Oomycetes and True Fungi. Phytopathology 2013, 103, 1035–1044. [Google Scholar] [CrossRef] [Green Version]
- Savory, F.; Leonard, G.; Richards, T.A. The Role of Horizontal Gene Transfer in the Evolution of the Oomycetes. PLoS Pathog. 2015, 11, e1004805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.H.Y.; de Bruijn, I.; Haas, B.J.; Belmonte, R.; Löbach, L.; Christie, J.; Ackerveken, G.V.D.; Bottin, A.; Bulone, V.; Díaz-Moreno, S.M.; et al. Distinctive Expansion of Potential Virulence Genes in the Genome of the Oomycete Fish Pathogen Saprolegnia parasitica. PLoS Genet. 2013, 9, e1003272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masigol, H.; Khodaparast, S.; Mostowfizadeh-Ghalamfarsa, R.; Mousanejad, S.; Rojas-Jimenez, K.; Grossart, H.P. Notes on Dictyuchus species (Stramenopila, Oomycetes) from Anzali lagoon, Iran. Mycol. Iran. 2018, 5, 79–89. [Google Scholar] [CrossRef]
- Shi, Z.; Han, C.; Zhang, X.; Tian, L.; Wang, L. Novel Synergistic Mechanism for Lignocellulose Degradation by a Thermophilic Filamentous Fungus and a Thermophilic Actinobacterium Based on Functional Proteomics. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Taxonomy | Sampling Site | GenBank Accession Number | |||
|---|---|---|---|---|---|---|
| SSU | ITS | LSU | coxI | |||
| Oomycetes | ||||||
| DSL2 | Saprolegnia ferax | Großer Stechlinsee | - | MN401033 | - | - |
| FB8 | Saprolegnia australis | Großer Stechlinsee | - | MN401034 | - | - |
| FB4 | Saprolegnia ferax | Großer Stechlinsee | - | MN401077 | - | - |
| FBLD1 | Saprolegnia ferax | Großer Stechlinsee | - | MN401076 | - | - |
| FB5 | Saprolegnia australis | Großer Stechlinsee | - | MN401078 | - | - |
| FBSD1 | Saprolegnia ferax | Großer Stechlinsee | - | MN401079 | - | - |
| JSW8-1 | Unknown Saprolegniales | Großer Stechlinsee | - | - | - | - |
| MDL14-2 | Saprolegnia anisospora | Dagowsee | - | MS401076 | - | MN398165 |
| MDL5-2 | Saprolegnia anisospora | Dagowsee | - | MN401035 | - | MN398166 |
| AF2-2 | Leptolegnia sp. | Große Fuchskuhle | - | MN401036 | - | MN398167 |
| AFW2-2 | Achlya sp. | Große Fuchskuhle | - | MN401037 | - | MN398168 |
| AF3 | Achlya sp. | Große Fuchskuhle | - | MN401038 | - | - |
| AFL4-3 | Achlya colorata | Große Fuchskuhle | - | MN401039 | - | - |
| AFL3 | Achlya sp. | Große Fuchskuhle | - | MN401040 | - | - |
| AFL10 | Phytophthora undulata | Große Fuchskuhle | - | MN401041 | - | MN398169 |
| AFW1-1 | Pythium sp. | Große Fuchskuhle | - | MN401042 | - | MN398170 |
| AFL5-1 | Pythium sp. | Große Fuchskuhle | - | MN401043 | - | MN398171 |
| MSK2-6 | Achlya ambisexualis | Breiter Luzin A | - | MN401044 | - | - |
| MSK1-2 | Saprolegnia ferax | Breiter Luzin A | - | MN401045 | - | - |
| MSK2-8 | Achlya ambisexualis | Breiter Luzin A | - | MN401046 | - | MN398172 |
| MSK3-5 | Achlya ambisexualis | Breiter Luzin B | - | MN401047 | - | - |
| MSK3-3 | Saprolegnia ferax | Breiter Luzin B | - | MN401048 | - | MN398173 |
| MSK3-6 | Saprolegnia ferax | Breiter Luzin B | - | MN401049 | - | - |
| MSK3-2 | Achlya ambisexualis | Breiter Luzin B | - | MN401050 | - | - |
| MSK3-7 | Saprolegnia australis | Breiter Luzin B | - | MN401051 | - | - |
| MPLL1-1 | Leptolegnia sp. | Peetschsee | - | MN401052 | - | - |
| MPLL1-2 | Leptolegnia sp. | Peetschsee | - | MN401053 | - | - |
| MPLW4 | Achlya ambisexualis | Peetschsee | - | MN401054 | - | - |
| MPLW2-1 | Achlya ambisexualis | Peetschsee | - | MN401055 | - | - |
| MPLL3-2 | Achlya ambisexualis | Peetschsee | - | MN401056 | - | - |
| MKAS8 | Leptolegnia sp. | Kochsee | - | MN401057 | - | - |
| MKAS19-2 | Leptolegnia sp. | Kochsee | - | MN401058 | - | - |
| MKAS19-1 | Saprolegnia ferax | Kochsee | - | MN401059 | - | - |
| MKAS6 | Leptolegnia sp. | Kochsee | - | MN401060 | - | MN398174 |
| MKAS16 | Leptolegnia sp. | Kochsee | - | MN401061 | - | - |
| MKAS13 | Leptolegnia sp. | Kochsee | - | MN401062 | - | - |
| MKAS20 | Saprolegnia ferax | Kochsee | - | MN401063 | - | MN398175 |
| MKAS5 | Leptolegnia sp. | Kochsee | - | MN401064 | - | - |
| JT1W3-2 | Unknown Saprolegniales | Tiefwarensee | - | - | - | - |
| JT1W8 | Unknown Saprolegniales | Tiefwarensee | - | - | - | - |
| JT1L4 | Saprolegnia ferax | Tiefwarensee | - | MN401065 | - | - |
| JT1W12 | Unknown Saprolegniales | Tiefwarensee | - | - | - | MN398163 |
| JT1W4 | Unknown Saprolegniales | Tiefwarensee | - | - | - | MN398164 |
| JT2L8 | Unknown Saprolegniales | Melzer See | - | - | - | - |
| JT2W9-2 | Unknown Saprolegniales | Melzer See | - | - | - | - |
| JT2W5-1 | Saprolegnia ferax | Melzer See | - | MN401066 | - | - |
| JT2W5-2 | Saprolegnia ferax | Melzer See | - | MN401067 | - | - |
| JT2W6-2 | Saprolegnia australis | Melzer See | - | MN401068 | - | - |
| JT2L6-2 | Saprolegnia ferax | Melzer See | - | MN401069 | - | MN398176 |
| JGWL2 | Unknown Saprolegniales | Gerlinsee | - | - | - | - |
| JGWL1 | Unknown Saprolegniales | Gerlinsee | - | - | - | - |
| JGWW2 | Saprolegnia ferax | Gerlinsee | - | MN401070 | MN401080 | - |
| JRL2 | Achlya ambisexualis | Roofensee | - | MN401071 | MN401081 | - |
| JR5 | Unknown Saprolegniales | Roofensee | - | - | - | - |
| JR3 | Unknown Saprolegniales | Roofensee | - | - | - | - |
| JRL1 | Achlya sp. | Roofensee | - | MN401072 | - | - |
| JWTW3 | Leptolegnia sp. | Wittwesee | - | MN401073 | MN401082 | - |
| JWTL2 | Saprolegnia ferax | Wittwesee | - | MN401074 | MN401083 | - |
| JWTW4 | Leptolegnia sp. | Wittwesee | - | MN401075 | MN401084 | - |
| JWTW2 | Unknown Saprolegniales | Wittwesee | - | - | - | - |
| JWMW1 | Unknown Saprolegniales | Großer Wummsee | - | - | - | - |
| JWML1 | Unknown Saprolegniales | Großer Wummsee | - | - | - | - |
| Fungi | ||||||
| ASW11-2 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| ASW3-23 | Cordycipitaceae | Großer Stechlinsee | MN396371 | - | MN396192 | - |
| ASW5-1 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| ASW11-1 | Neonectria sp. | Großer Stechlinsee | - | - | MN396194 | - |
| ASW5-2 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| MSH1 | Cordycipitaceae | Großer Stechlinsee | MN396372 | - | MN396193 | - |
| ASW9 | Unknown fungus | Großer Stechlinsee | - | - | - | |
| MSH2 | Cladosporium cladosporioides complex | Großer Stechlinsee | MN396373 | MN401001 | MN396195 | - |
| MSH3 | Cladosporium cladosporioides complex | Großer Stechlinsee | MN396374 | MN401002 | MN396196 | - |
| ASW8 | Cladosporium herbarum complex | Großer Stechlinsee | MN396375 | MN401003 | MN396197 | - |
| ASW3-22 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| ASW1 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| ASW4 | Phlebia radiata | Großer Stechlinsee | MN396376 | MN401004 | MN396198 | - |
| FBP1 | Penicillium brevicompactum | Großer Stechlinsee | MN396377 | MN401005 | MN396199 | - |
| FBL1 | Cladosporium herbarum complex | Großer Stechlinsee | - | MN401006 | MN396200 | - |
| FBP3 | Penicillium brevicompactum | Großer Stechlinsee | MN396378 | MN401007 | MN396201 | - |
| FBP22 | Penicillium sp. | Großer Stechlinsee | MN396379 | - | MN396202 | - |
| FBS1 | Didymosphaeriaceae | Großer Stechlinsee | - | MN401008 | MN396203 | - |
| FBL3 | Heterobasidion sp. | Großer Stechlinsee | - | MN401009 | MN396204 | - |
| FBP21 | Penicillium brevicompactum | Großer Stechlinsee | MN396380 | MN401010 | MN396205 | - |
| FBL6 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| FBPD1 | Fusarium sp. | Großer Stechlinsee | MN396381 | MN401011 | MN396206 | - |
| FB10 | Nectriaceae | Großer Stechlinsee | - | - | MN396207 | - |
| FB11 | Cladosporium herbarum complex | Großer Stechlinsee | - | MN401012 | MN396208 | - |
| FBLD2 | Cladosporium sp. | Großer Stechlinsee | - | - | MN396209 | - |
| FB3 | Penicillium levitum | Großer Stechlinsee | - | - | MN396210 | - |
| FB1 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| FB2 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| FB6 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| FB7 | Cladosporium herbarum complex | Großer Stechlinsee | - | MN401013 | MN396211 | - |
| FBP1x | Nectria sp. | Großer Stechlinsee | - | MN401014 | MN396212 | - |
| FBS4 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| FBP41 | Penicillium brevicompactum | Großer Stechlinsee | MN396382 | MN401015 | MN396213 | - |
| FBP42 | Penicillium brevicompactum | Großer Stechlinsee | - | MN401016 | MN396214 | - |
| FBP5 | Penicillium brevicompactum | Großer Stechlinsee | - | - | MN396215 | - |
| FBL5 | Phoma sp. | Großer Stechlinsee | - | - | MN396216.1 | - |
| FB12 | Cladosporium sp. | Großer Stechlinsee | - | - | MN396217 | - |
| FB9 | Massarina sp. | Großer Stechlinsee | - | - | MN396218 | - |
| FBL81 | Cladosporium cladosporioides complex | Großer Stechlinsee | - | - | MN396219 | - |
| FBPD2 | Cladosporium herbarum complex | Großer Stechlinsee | - | - | MN396220 | - |
| FBP9 | Penicillium brevicompactum | Großer Stechlinsee | - | - | MN396221 | - |
| FBS9 | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| FBP7 | Penicillium brevicompactum | Großer Stechlinsee | - | - | MN396222 | - |
| FBSL1 | Penicillium crustosum | Großer Stechlinsee | MN396383 | - | - | - |
| FBP8 | Cladosporium cladosporioides complex | Großer Stechlinsee | - | MN401026 | - | - |
| FBP81 | Penicillium brevicompactum | Großer Stechlinsee | - | - | MN396223 | - |
| FBP71 | Cladosporium herbarum complex | Großer Stechlinsee | - | - | MN396224 | - |
| FBS71 | Penicillium brevicompactum | Großer Stechlinsee | - | - | MN396225 | - |
| FBSS1 | Fusarium sp. | Großer Stechlinsee | - | - | MN396226 | - |
| FBLD2X | Unknown fungus | Großer Stechlinsee | - | - | - | - |
| AFL4-2 | Mortierella sp. | Große Fuchskuhle | - | MN401032.1 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masigol, H.; Woodhouse, J.N.; van West, P.; Mostowfizadeh-Ghalamfarsa, R.; Rojas-Jimenez, K.; Goldhammer, T.; Khodaparast, S.A.; Grossart, H.-P. Phylogenetic and Functional Diversity of Saprolegniales and Fungi Isolated from Temperate Lakes in Northeast Germany. J. Fungi 2021, 7, 968. https://doi.org/10.3390/jof7110968
Masigol H, Woodhouse JN, van West P, Mostowfizadeh-Ghalamfarsa R, Rojas-Jimenez K, Goldhammer T, Khodaparast SA, Grossart H-P. Phylogenetic and Functional Diversity of Saprolegniales and Fungi Isolated from Temperate Lakes in Northeast Germany. Journal of Fungi. 2021; 7(11):968. https://doi.org/10.3390/jof7110968
Chicago/Turabian StyleMasigol, Hossein, Jason Nicholas Woodhouse, Pieter van West, Reza Mostowfizadeh-Ghalamfarsa, Keilor Rojas-Jimenez, Tobias Goldhammer, Seyed Akbar Khodaparast, and Hans-Peter Grossart. 2021. "Phylogenetic and Functional Diversity of Saprolegniales and Fungi Isolated from Temperate Lakes in Northeast Germany" Journal of Fungi 7, no. 11: 968. https://doi.org/10.3390/jof7110968