
Phytophthora heterospora sp. nov., a New Pseudoconidia-Producing Sister Species of P. palmivora

 , , ,
, , ,  ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phytophthora Isolation and Culture Maintenance
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Phylogenetic Analyses
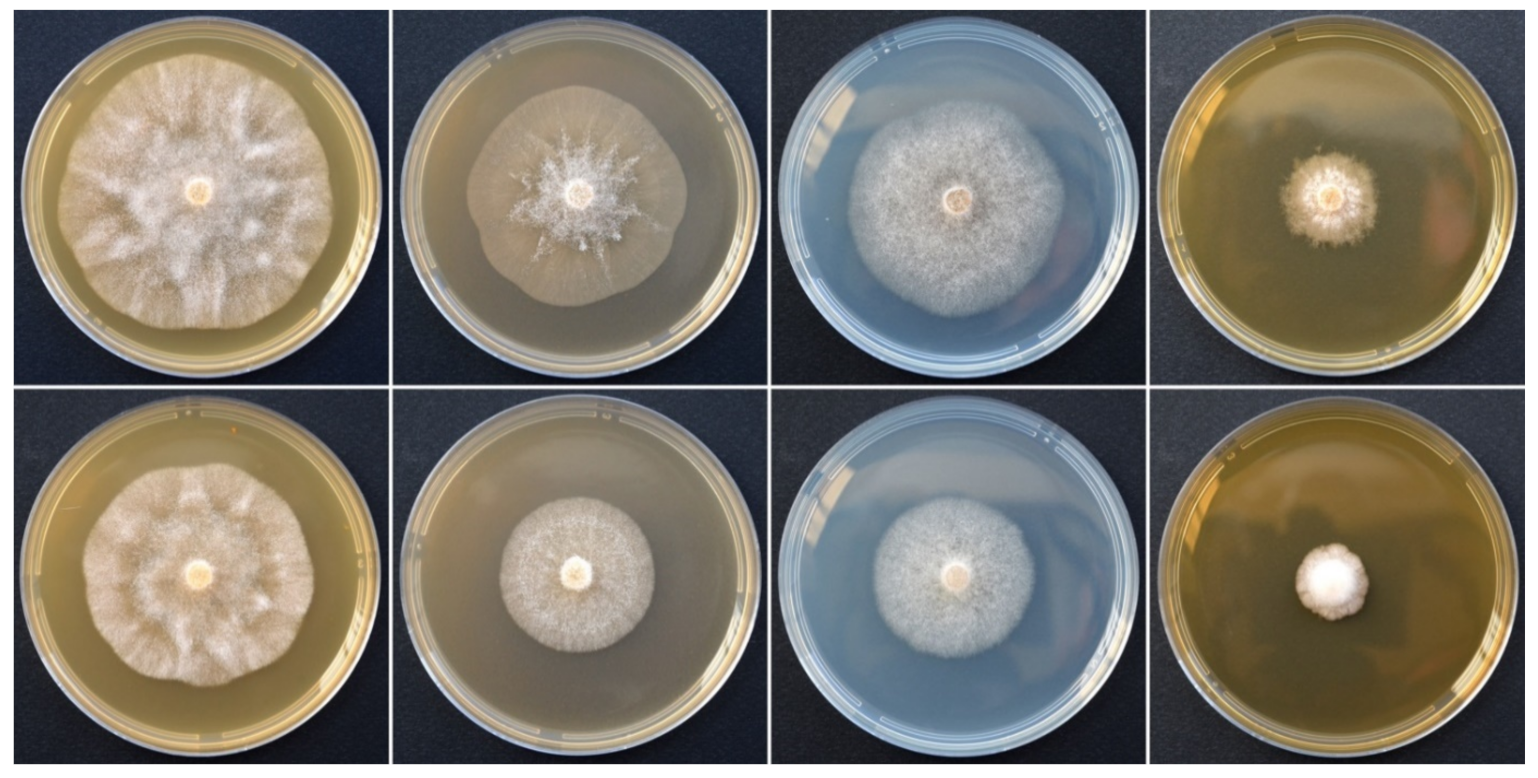
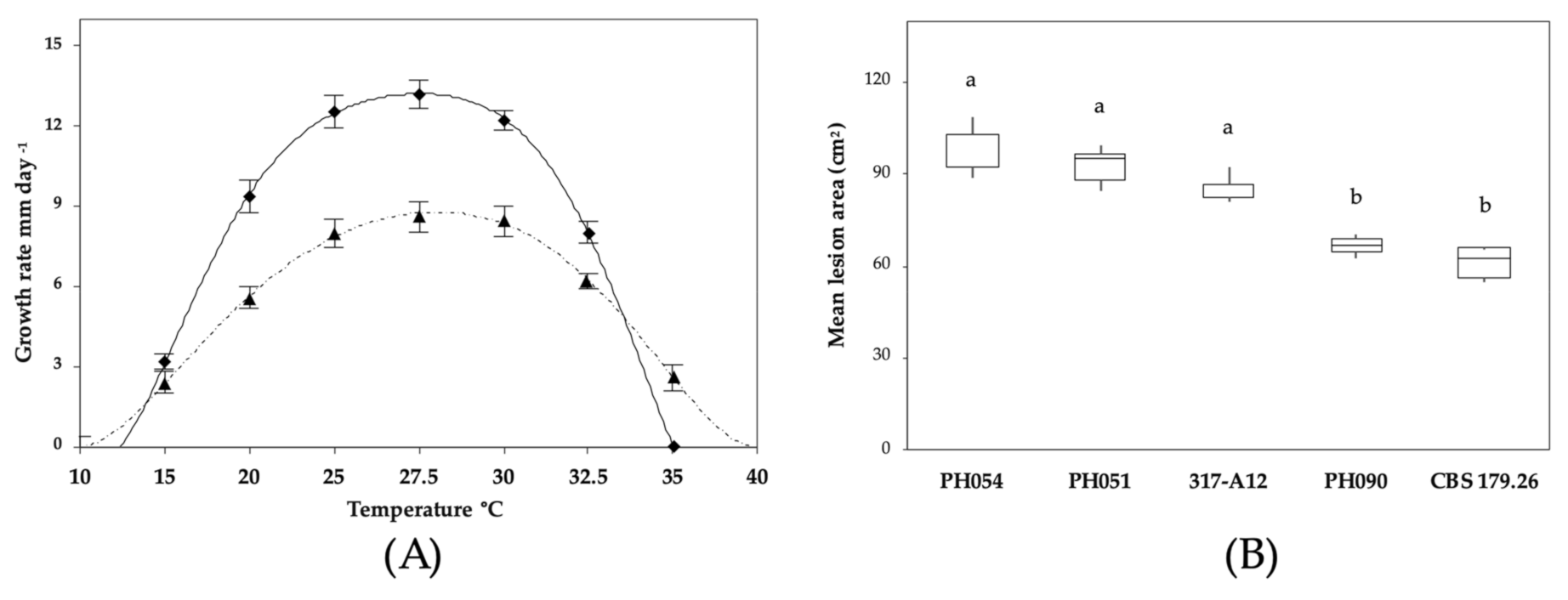
2.4. Morphological Characterization and Cardinal Temperature for Growth
2.5. Pathogenicity Test
3. Results
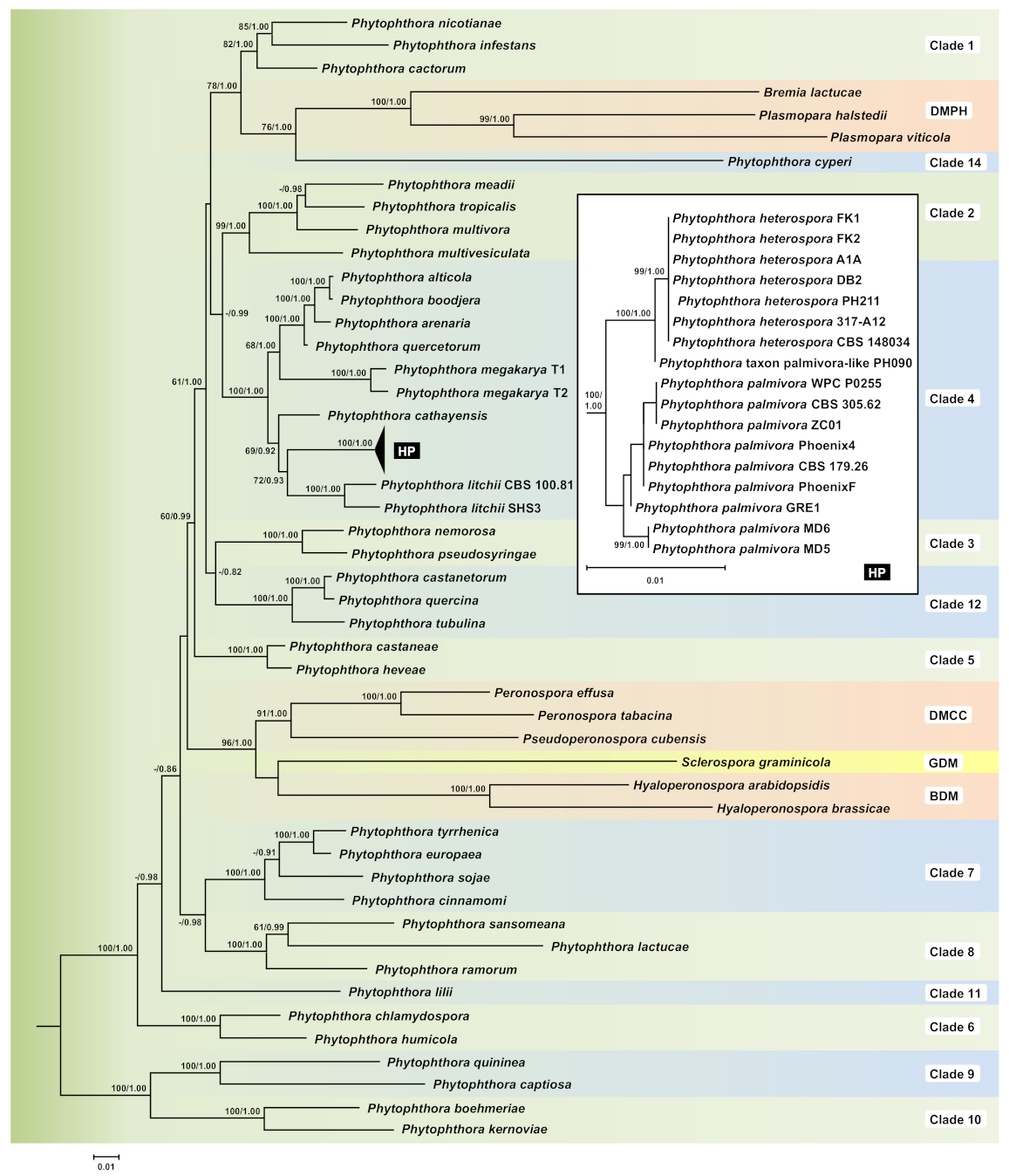
3.1. Phylogeny
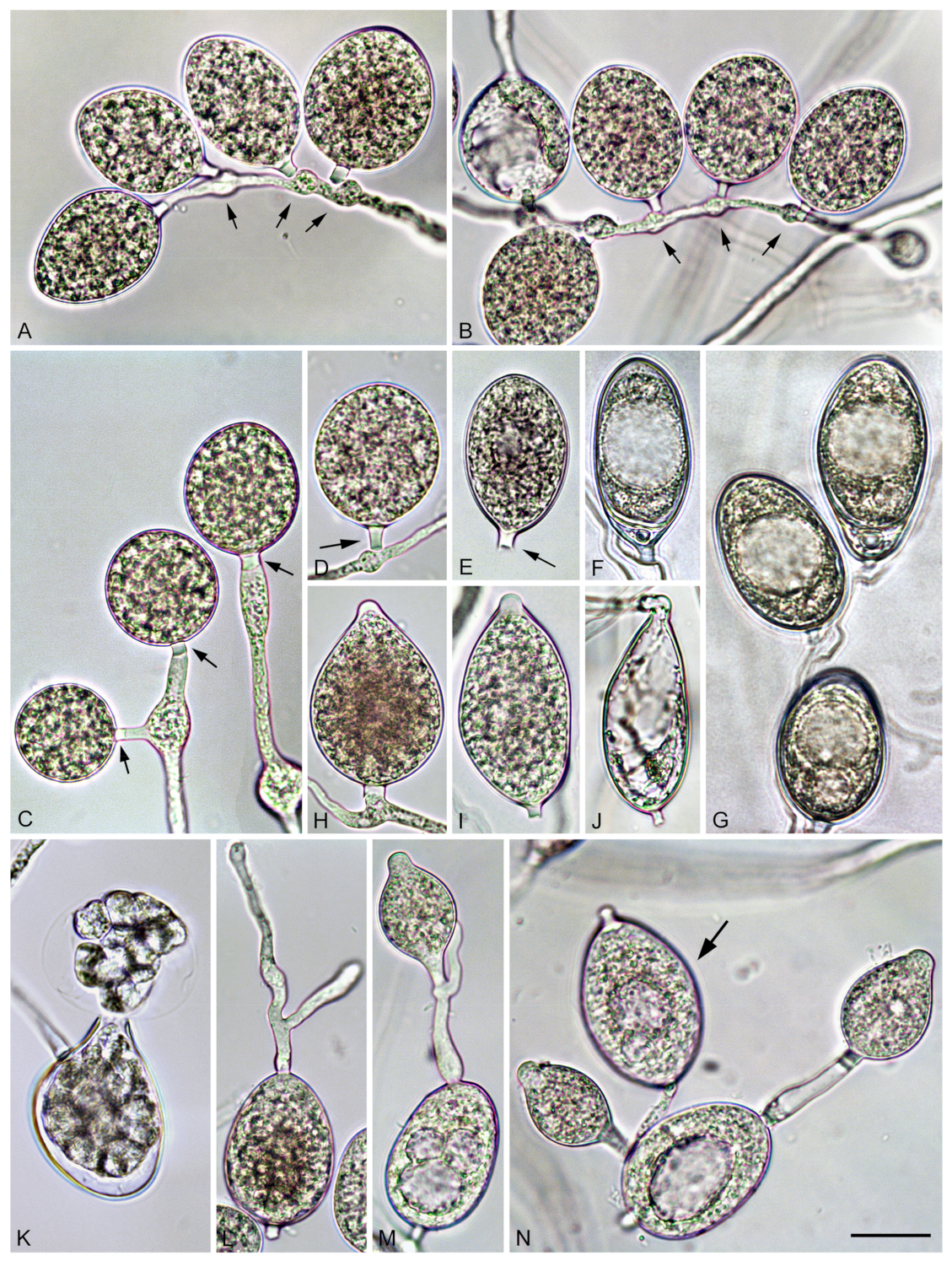
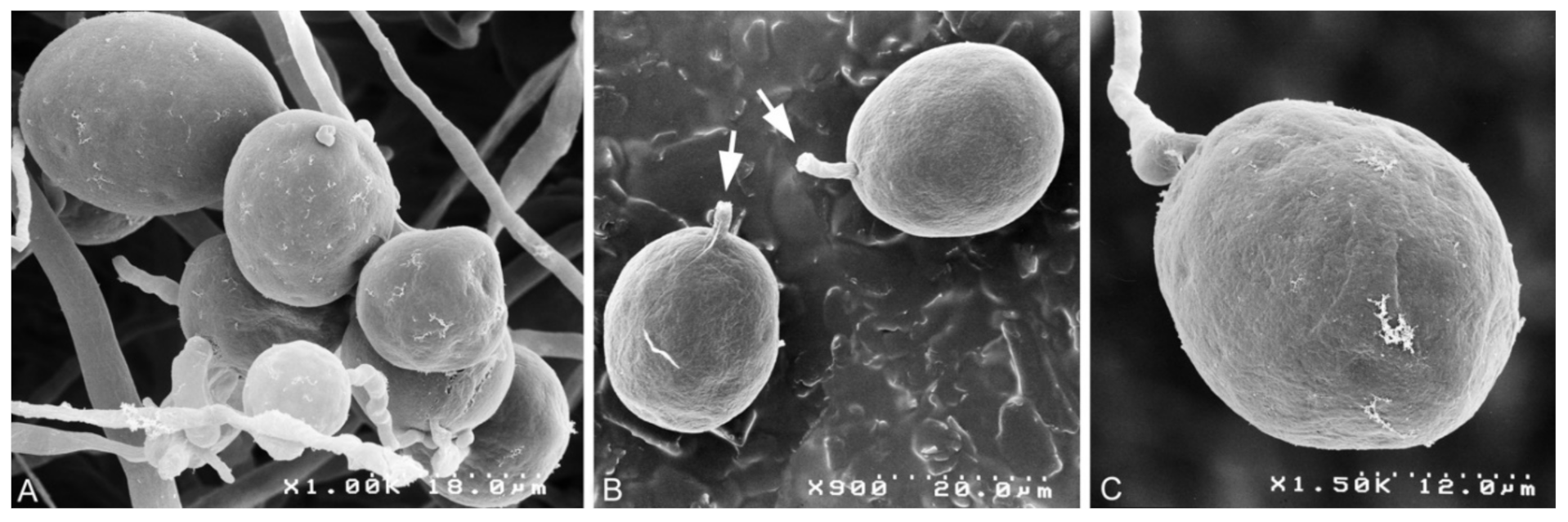
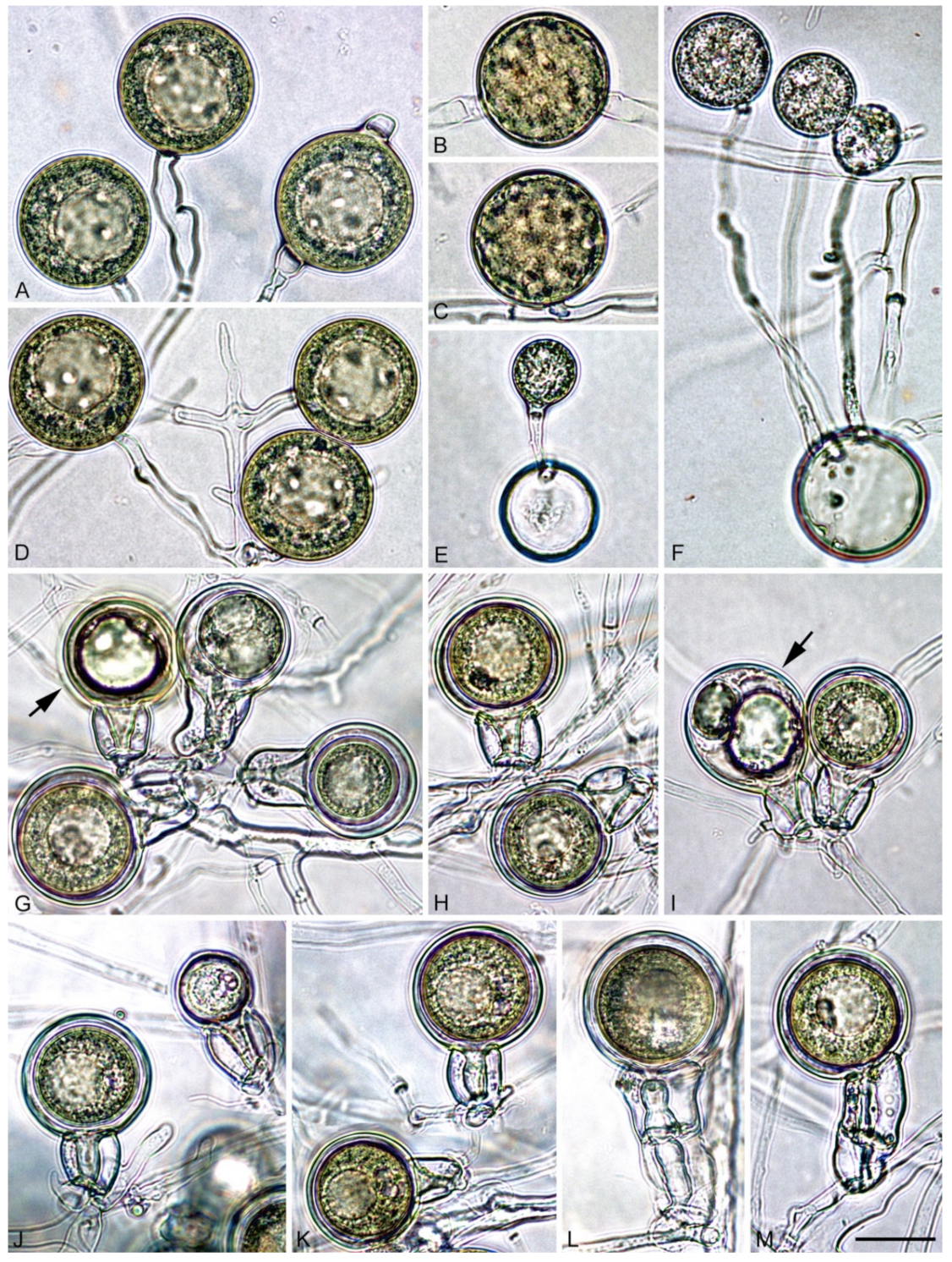
3.2. Taxonomy
3.3. Pathogenicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press American Phytopathological Society: St. Paul, MN, USA, 1996; p. 562. [Google Scholar]
- Hansen, E.M.; Reeser, P.W.; Sutton, W. Phytophthora beyond agriculture. Ann. Rev. Phytopathol. 2012, 50, 359–378. [Google Scholar] [CrossRef]
- Jung, T.; Sierra-Perez, A.; Duran, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [Green Version]
- Bourret, T.B.; Choudhury, R.A.; Mehl, H.K.; Blomquist, C.L.; McRoberts, N.; Rizzo, D.M. Multiple origins of downy mildews and mito-nuclear discordance within the paraphyletic genus Phytophthora. PLoS ONE 2018, 13, e0192502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbelotto, M.; Frankel, S.; Scanu, B. Soil-and waterborne Phytophthora species linked to recent outbreaks in Northern California restoration sites. Calif. Agric. 2018, 72, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Horta Jung, M.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oßwald, W.; Fleischmann, F.; Rigling, D.; Coelho, A.C.; Cravador, A.; Diez, J.; Dalio, R.J.; Horta Jung, M.; Pfanz, H.; Robin, C.; et al. Strategies of attack and defence in woody plant-Phytophthora interactions. For. Pathol. 2014, 44, 169–190. [Google Scholar] [CrossRef]
- Bassani, I.; Larousse, M.; Tran, Q.D.; Attard, A.; Galiana, E. Phytophthora zoospores: From perception of environmental signals to inoculum formation on the host-root surface. Comput. Struct. Biotechnol. J. 2020, 18, 3766–3773. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.R.; Webber, J.F. Sporulation potential, symptom expression and detection of Phytophthora ramorum on larch needles and other foliar hosts. Plant Pathol. 2016, 65, 1441–1451. [Google Scholar] [CrossRef] [Green Version]
- Scanu, B.; Webber, J.F. Dieback and mortality of Nothofagus in Britain: Ecology, pathogenicity and sporulation potential of the causal agent Phytophthora pseudosyringae. Plant Pathol. 2016, 65, 26–36. [Google Scholar] [CrossRef]
- Scanu, B.; Cacciola, S.O.; Linaldeddu, B.; Pane, A.; Franceschini, A.; Magnano di San Lio, G. Characterization and evolutionary significance of a new Phytophthora species producing conidia. J. Plant Pathol. 2015, 97, 38. [Google Scholar]
- Cacciola, S.O.; Agosteo, G.E.; Pane, A. First report of Phytophthora palmivora as a pathogen of olive in Italy. Plant Dis. 2000, 84, 1153. [Google Scholar] [CrossRef] [PubMed]
- Agosteo, G.E.; Magnano di San Lio, G.; Cacciola, S.O. Root rot of young olive trees caused by Phytophthora palmivora in southern Italy. Acta Hortic. 2002, 586, 709–712. [Google Scholar] [CrossRef]
- Babacauh, K.D. Structure des populations de Phytophthora palmivora (Butl.) Butl. Emend. Bras. Et Griff. Parasite du cacaoyer (Theobroma cacao L.) (Population structure Phytophthora palmivora from cacao [Theobroma cacao L.]). Bul. Soc. Bot. Fr. Lett. Bot. 1983, 130, 15–25. [Google Scholar]
- Scanu, B.; Hunter, G.C.; Linaldeddu, B.T.; Franceschini, A.; Maddau, L.; Jung, T.; Denman, S. A taxonomic re-evaluation reveals that Phytophthora cinnamomi and Pcinnamomi var. parvispora are separate species. For. Pathol. 2014, 44, 1–20. [Google Scholar]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. Eur. J. For. Path. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Jeffers, S.N.; Martin, S.B. Comparison of two media selective for Phytophthora and Pythium spp. Plant Dis. 1986, 70, 1038–1043. [Google Scholar] [CrossRef]
- Brasier, C.M.; Griffin, M.J. Taxonomy of Phytophthora palmivora on cocoa. T. Brit. Mycol. Soc. 1979, 71, 111–143. [Google Scholar] [CrossRef]
- Pane, A.; Allatta, C.; Sammarco, G.; Cacciola, S.O. First report of bud rot of Canary Island date palm caused by Phytophthora palmivora in Italy. Plant Dis. 2007, 91, 1059. [Google Scholar] [CrossRef]
- Cacciola, S.O.; Pennisi, A.M.; Agosteo, G.E.; Magnano di San Lio, G. First Report of Phytophthora palmivora on Grevillea spp. in Italy. Plant Dis. 2003, 87, 1006. [Google Scholar] [CrossRef]
- Van Tri, M.; Van Hoa, N.; Minh Chau, N.; Pane, A.; Faedda, R.; De Patrizio, A.; Schena, L.; Olsson, C.H.B.; Wright, S.A.I.; Ramstedt, M.; et al. Decline of jackfruit (Artocarpus heterophyllus) incited by Phytophthora palmivora in Vietnam. Phytopathol. Mediterr. 2015, 54, 275–280. [Google Scholar]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal DNA for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Blair, J.E.; Coffey, M.D.; Park, S.-Y.; Greiser, D.M.; Kang, S. A multi-locus phylogeny for Phytophthora utilizing markers derived from complete genome sequences. Fungal Genet. Biol. 2008, 45, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.N.; Tooley, P.W. Phylogenetic relationships among Phytophthora species inferred from sequence analysis of mitochondrially encoded cytochrome oxidase I and II genes. Mycologia 2003, 95, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Kroon, L.P.N.M.; Bakker, F.T.; van den Bosch, G.B.M.; Bonants, P.J.M.; Flier, W.G. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNA sequences. Fungal Genet. Biol. 2004, 41, 766–782. [Google Scholar] [CrossRef]
- Hall, T. BioEdit Version 5.0.6. Department of Microbiology, North Carolina State University. Available online: http://www.mbio.ncsu.edu/BioEdit/bioedit.html (accessed on 8 June 2021).
- O’Hanlon, R.; Destefanis, M.; Milenković, I.; Tomšovský, M.; Janoušek, J.; Bellgard, S.E.; Weir, B.S.; Kudláček, T.; Horta Jung, M.; Jung, T. Two new Nothophytophthora species from streams in Ireland and Northern Ireland: Nothophytophthora irlandica and N. lirii sp. nov. PLoS ONE 2021, 16, e0250527. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Klosterman, S.J.; Kummer, V.; Voglmayr, H.; Shin, H.D.; Thines, M. Multi-locus tree and species tree approaches toward resolving a complex clade of downy mildews (Straminipila, Oomycota), including pathogens of beet and spinach. Mol. Phylogenet. Evol. 2015, 86, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Heuelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol Evol. 2020, 12, 373–377. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. 2019 Version 3.61. Available online: http://www.mesquiteproject.org (accessed on 1 June 2021).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Scanu, B.; Linaldeddu, B.T.; Deidda, A.; Jung, T. Diversity of Phytophthora species from declining Mediterranean maquis vegetation, including two new species, Phytophthora crassamura and P. ornamentata sp. nov. PLoS ONE 2015, 10, e0143234. [Google Scholar] [CrossRef] [Green Version]
- Dick, M.W. Keys to Pythium; University of Reading Press: London, UK, 1990; p. 63. [Google Scholar]
- Drenth, A.; Guest, D.I. Diversity and Management of Phytophthora in Southeast Asia; ACIAR Monograph 114; Australian Centre for International Agricultural Research: Canberra, Australia, 2004; p. 238. [Google Scholar]
- Göker, M.; Voglmayr, H.; Riethmüller, A.; Oberwinkler, F. How do obligate parasites evolve? A multi-gene phylogenetic analysis of downy mildews. Fungal Genet. Biol. 2007, 44, 105–122. [Google Scholar] [PubMed]
- Thines, M.; Choi, Y.J. Evolution, diversity, and taxonomy of the Peronosporaceae, with focus on the genus Peronospora. Phytopathology 2016, 106, 6–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.G.; Zheng, X.B.; Wang, Y.C.; Ko, W.H. Evaluation of the rearrangement of taxonomic position of Peronophythora litchii based on partial DNA sequences. Bot. Stud. 2007, 48, 79–89. [Google Scholar]
- Ye, W.; Wang, Y.; Shen, D.; Li, D.; Pu, T.; Jiang, Z.; Zhang, Z.; Zheng, X.; Tyler, B.M.; Wang, Y. Sequencing of the litchi downy blight pathogen reveals it is a Phytophthora species with downy mildew-like characteristics. Mol. Plant-Microbe Interact. 2016, 29, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.H.; Zheng, F.C.; Zheng, H.C. Phytophthora cyperi on Digitaria ciliaris in Hainan Province of China. Mycotaxon 2004, 90, 431–435. [Google Scholar]
- Zheng, X.B.; Ho, H.H. The sexual stage of Phytophthora polygoni Saw. Bot Bull Acad Sin. 2000, 41, 57–60. [Google Scholar]
- Göker, M.; Voglmayr, H.; Riethmüeller, A.; Weiß, M.; Oberwinkler, F. Taxonomic aspects of Peronosporaceae inferred from Bayesian molecular phylogenetics. Can. J. Bot. 2003, 81, 672–683. [Google Scholar] [CrossRef]
- Thines, M.; Göker, M.; Oberwinkler, F.; Spring, O. A revision of Plasmopara penniseti, with implications for the host range of the downy mildews with pyriform haustoria. Mycol. Res. 2007, 111, 1377–1385. [Google Scholar] [CrossRef]
- Thines, M.; Göker, M.; Telle, S.; Ryley, M.; Mathur, K.; Narayana, Y.D.; Spring, O.; Thakur, R.P. Phylogenetic relationships in graminicolous downy mildews based on cox2 sequence data. Mycol. Res. 2008, 112, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Gaümann, E.A. The Fungi. A Description of Their Morphological Features and Evolutionary Development; Hafner Publishing: New York, NY, USA; London, UK, 1952. [Google Scholar]
- Waterhouse, G.M. Peronosporales. In The Fungi—An Advanced Treatise; Ainsworth, G.C., Sparrow, F.K., Sussman, A.S., Eds.; Academic Press: New York, NY, USA, 1973; pp. 165–183. [Google Scholar]
- Dick, M.W.; Wong, P.T.W.; Clark, G. The identity of the oomycete causing kikuyu yellows with a reclassification of the downy mildews. Bot. J. Linn. Soc. 1984, 89, 171–198. [Google Scholar] [CrossRef]
- Riethmüller, A.; Voglmayr, H.; Goker, M.; Weiss, M.; Oberwinkler, F. Phylogenetic relationships of the downy mildews (Per-onosporales) and related groups based on nuclear large subunit ribosomal DNA sequences. Mycologia 2002, 94, 834. [Google Scholar] [CrossRef]
- Runge, F.; Telle, S.; Ploch, S.; Savory, E.; Day, B.; Sharma, R. Thines M. The inclusion of downy mildews in a multi- locus-dataset and its reanalysis reveals a high degree of paraphyly in Phytophthora. IMA Fungus 2011, 2, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Martin, F.N.; Blair, J.E.; Coffey, M.D. A combined mitochondrial and nuclear multilocus phylogeny of the genus Phytophthora. Fungal Genet. Biol. 2014, 66, 19–32. [Google Scholar] [CrossRef]
- Fletcher, K.; Gil, J.; Bertier, L.D.; Kenefick, A.; Wood, K.J.; Zhang, L.; Reyes-Chin-Wo, S.; Cavanaugh, K.; Tsuchida, C.; Wong, J.; et al. Genomic signatures of heterokaryosis in the oomycete pathogen Bremia lactucae. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Butler, E.J. Report of the Imperial Mycologist, 1918–1919; Scientific Reports of the Agricultural Research Institute, Pusa: Calcutta, India, 1919; p. 82. [Google Scholar]
- IDphy: Molecular and Morphological Identification of Phytophthora. Available online: https://idtools.org/id/phytophthora/molecular.php (accessed on 8 September 2021).
- Linaldeddu, B.T.; Bregant, C.; Ruzzon, B.; Montecchio, L. Coniella granati and Phytophthora palmivora the main pathogens involved in pomegranate dieback and mortality in north-eastern Italy. Ital. J. Mycol. 2020, 49, 92–100. [Google Scholar]
- Kurbetli, İ.; Karaca, G.; Aydoğdu, M. Phytophthora species causing root and collar rot of pomegranate in Turkey. Eur. J. Plant. Pathol. 2020, 157, 485–496. [Google Scholar] [CrossRef]
- Chliyeh, M.; Ouazzani Touhami, A.; Filali-Maltouf, A.; El Modafar, C.; Moukhli, A.; Oukabli, A.; Benkirane, R.; Douira, A. Phytophthora palmivora: A new pathogen of olive trees in Morocco. Atlas J. Biol. 2013, 2, 130–135. [Google Scholar] [CrossRef]
- Cacciola, S.O.; Faedda, R.; Pane, A.; Scarito, G. Root and crown rot of olive caused by Phytophthora spp. In Olive Diseases and Disorders; Schena, L., Agosteo, G.E., Cacciola, S.O., Magnano di San Lio, G., Eds.; Transworld Research Network: Trivandrum, Kerala, India, 2011; pp. 305–327. [Google Scholar]
- Cacciola, S.O.; Gullino, M.L. Emerging and re-emerging fungus and oomycete soil-borne plant diseases in Italy. Phytopathol. Mediterr. 2019, 58, 451–472. [Google Scholar]
- Brasier, C.M. The biosecurity threat to the UK and global environment from international trade in plants. Plant Pathol. 2008, 57, 792–808. [Google Scholar] [CrossRef]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguin Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Migliorini, D.; Ghelardini, L.; Tondini, E.; Luchi, N.; Santini, A. The potential of symptomless potted plants for carrying invasive soil-borne plant pathogens. Divers. Distrib. 2015, 21, 1218–1229. [Google Scholar] [CrossRef]
- Drenth, A.; Guest, D. Phytophthora palmivora in Tropical Tree Crops. In Phytophthora—A Global Perspective; Lamour, K., Ed.; CABI: Oxfordshire, UK, 2013; pp. 187–196. [Google Scholar]
- Mchau, G.R.A.; Coffey, M.D. Isozyme diversity in Phytophthora palmivora: Evidence for a Southeast Asian centre of origin. Mycol. Res. 1994, 98, 1035–1043. [Google Scholar] [CrossRef]
- Zentmyer, G. Origin and distribution of four species of Phytophthora. Trans. Br. Mycol. Soc. 1988, 91, 367–378. [Google Scholar] [CrossRef]
- Brasier, C.M.; Vettraino, A.M.; Chang, T.T.; Vannini, A. Phytophthora lateralis discovered in an old growth Chamaecyparis forest in Taiwan. Plant Pathol. 2010, 59, 595–603. [Google Scholar] [CrossRef]
- Jung, T.; Chang, T.T.; Bakonyi, J.; Seress, D.; Pérez-Sierra, A.; Yang, X.; Hong, C.; Scanu, B.; Fu, C.H.; Hsueh, K.L.; et al. Diversity of Phytophthora species in natural ecosystems of Taiwan and association with disease symptoms. Plant Pathol. 2017, 66, 194–211. [Google Scholar] [CrossRef]
- Jung, T.; Horta Jung, M.; Webber, J.F.; Kageyama, K.; Hieno, A.; Masuya, H.; Uematsu, S.; Pérez-Sierra, A.; Harris, A.R.; Forster, J.; et al. The Destructive Tree Pathogen Phytophthora ramorum Originates from the Laurosilva Forests of East Asia. J. Fungi 2021, 7, 226. [Google Scholar] [CrossRef]
- Shakya, S.K.; Grünwald, N.J.; Fieland, V.J.; Knaus, B.J.; Weiland, J.E.; Maia, C.; Drenth, A.; Guest, D.I.; Liew, E.C.Y.; Crane, C.; et al. Phylogeography of the wide-host range panglobal plant pathogen Phytophthora cinnamomi. Mol Ecol. 2021, 30, 5164–5178. [Google Scholar] [CrossRef]
- Grünwald, N.J.; McDonald, B.A.; Milgroom, M.G. Population genomics of fungal and oomycete pathogens. Ann. Rev. Phytopat. 2016, 54, 323–346. [Google Scholar] [CrossRef]
- Ali, S.; Shao, J.; Lary, D.; Strem, M.; Meinhardt, L.; Bailey, B. Phytophthora megakarya and P. palmivora, Causal Agents of Black Pod Rot, Induce Similar Plant Defense Responses Late during Infection of Susceptible Cacao Pods. Front. Plant Sci. 2017, 8, 169. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Horta Jung, M.; Scanu, B.; Seress, D.; Kovács, G.M.; Maia, C.; Pérez-Sierra, A.; Chang, T.T.; Chandelier, A.; Heungens, K.; et al. Six new Phytophthora species from ITS Clade 7a including two sexually functional heterothallic hybrid species detected in natural ecosystems in Taiwan. Persoonia 2017, 38, 100–135. [Google Scholar] [CrossRef] [Green Version]
- Burgess, T.I. Molecular characterization of natural hybrids formed between five related indigenous Clade 6 Phytophthora species. PLoS ONE 2015, 10, e0134225. [Google Scholar] [CrossRef] [Green Version]
- Van Poucke, K.; Haegeman, A.; Goedefroit, T.; Focquet, F.; Leus, L.; Horta Jung, M.; Nave, C.; Redondo, M.A.; Husson, C.; Kostov, K.; et al. Unravelling hybridization in Phytophthora using phylogenomics and genome size estimation. IMA Fungus 2021, 12, 16. [Google Scholar] [CrossRef]
- Guo, Y.; Sakalidis, M.L.; Torres-Londoño, G.A.; Hausbeck, M. Population structure of a worldwide Phytophthora palmivora collection suggests lack of host specificity and reduced genetic diversity in South American and Caribbean. Plant Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Crops and Livestock Products. 2021. Available online: http://www.fao.org/faostat/en/#data/QCL (accessed on 8 September 2021).
- Kurbetli, I.; Sulu, G.; TaŞtekİn, E.; Polat, I. First report of Phytophthora inundata causing olive tree decline in Turkey. Can. J. Plant Pathol. 2016, 38, 254–257. [Google Scholar] [CrossRef]
- González, M.; Pérez-Sierra, A.; Serrano, M.S.; Sanchez, M.E. Two Phytophthora species causing decline of wild olive (Olea europaea subsp. europaea var. sylvestris). Plant Pathol. 2017, 66, 941–948. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Schena, L.; Agosteo, G.E.; Magnano di San Lio, G.; Cacciola, S.O. Phytophthora oleae sp. nov. causing fruit rot of olive in southern Italy. Plant Pathol. 2018, 67, 1362–1373. [Google Scholar] [CrossRef]
- Linaldeddu, B.T.; Bregant, C.; Montecchio, L.; Favaron, F.; Sella, L. First report of Phytophthora acerina, P. pini and P. plurivora causing root rot and sudden death on olive trees in Italy. Plant Dis. 2020, 104, 996. [Google Scholar] [CrossRef]
- Santilli, E.; Riolo, M.; La Spada, F.; Pane, A.; Cacciola, S.O. First report of root rot caused by Phytophthora bilorbang on Olea europaea in Italy. Plants 2020, 9, 826. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytophthora Taxa | Isolate Codes 1 | Country, Region | Year | Host |
|---|---|---|---|---|
| P. heterospora | PH047 | Italy, Sardinia | 2010 | Olea europaea |
| P. heterospora | PH051 (CBS 148035) | Italy, Sardinia | 2010 | Olea europaea |
| P. heterospora | PH052 | Italy, Sardinia | 2010 | Olea europaea |
| P. heterospora | PH054 (CBS 148034) (T) | Italy, Sardinia | 2010 | Olea europaea |
| P. heterospora | PH057 | Italy, Sardinia | 2010 | Olea europaea |
| P. heterospora | PH211 | Italy, Sardinia | 2013 | Juniperus oxycedrus |
| P. heterospora | 317-A12 (CBS 148036) | Italy, Sicily | 2014 | Capparis spinosa |
| P. heterospora | Palm2 2 | Italy, Calabria | 1999 | Olea europaea |
| P. heterospora | Campobello 2b | Italy, Sicily | 2005 | Olea europaea |
| P. heterospora | DB2 | Vietnam, Mekong Delta | 2013 | Durio zibethinus |
| P. heterospora | A1A | Vietnam, Mekong Delta | 2013 | Durio zibethinus |
| P. heterospora | A1B1 | Vietnam, Mekong Delta | 2013 | Durio zibethinus |
| P. heterospora | C2B1 | Vietnam, Mekong Delta | 2013 | Durio zibethinus |
| P. palmivora | CBS 179.26 3 | Sri Lanka, n.a. | 1979 | Theobroma cacao |
| P. palmivora | Phoenix4 4 | Italy, Sicily | 2005 | Phoenix canariensis |
| P. palmivora | PhoenixF 4 | Italy, Sicily | 2005 | Phoenix canariensis |
| P. palmivora | GRE1 (IMI 390579) 5 | Italy, Sicily | 2002 | Grevillea rosmarinifolia |
| P. palmivora | MD5 (IMI 503890) 6 | Vietnam, Mekong Delta | 2013 | Artocarpus heterophyllus |
| P. palmivora | MD6 (IMI 503891) 6 | Vietnam, Mekong Delta | 2013 | Artocarpus heterophyllus |
| P. taxon palmivora-like | PH083 | Italy, Sardinia | 2011 | Arbutus unedo |
| P. taxon palmivora-like | PH090 | Italy, Sardinia | 2011 | Arbutus unedo |
| Phytophthora spp. | Pseudoconidia/Sporangia/Chlamydospores | |
|---|---|---|
| CA | Water | |
| P. heterospora1 | 86/8/6 | 74/24/2 |
| P. heterospora2 | 34/56/10 | 26/74/8 |
| P. taxon palmivora-like | -/82/18 | -/85/15 |
| P. palmivora | -/78/22 | -/80/20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scanu, B.; Jung, T.; Masigol, H.; Linaldeddu, B.T.; Jung, M.H.; Brandano, A.; Mostowfizadeh-Ghalamfarsa, R.; Janoušek, J.; Riolo, M.; Cacciola, S.O. Phytophthora heterospora sp. nov., a New Pseudoconidia-Producing Sister Species of P. palmivora. J. Fungi 2021, 7, 870. https://doi.org/10.3390/jof7100870
Scanu B, Jung T, Masigol H, Linaldeddu BT, Jung MH, Brandano A, Mostowfizadeh-Ghalamfarsa R, Janoušek J, Riolo M, Cacciola SO. Phytophthora heterospora sp. nov., a New Pseudoconidia-Producing Sister Species of P. palmivora. Journal of Fungi. 2021; 7(10):870. https://doi.org/10.3390/jof7100870
Chicago/Turabian StyleScanu, Bruno, Thomas Jung, Hossein Masigol, Benedetto Teodoro Linaldeddu, Marília Horta Jung, Andrea Brandano, Reza Mostowfizadeh-Ghalamfarsa, Josef Janoušek, Mario Riolo, and Santa Olga Cacciola. 2021. "Phytophthora heterospora sp. nov., a New Pseudoconidia-Producing Sister Species of P. palmivora" Journal of Fungi 7, no. 10: 870. https://doi.org/10.3390/jof7100870