Yeasts in Liquid Swine Diets: Identification Methods, Growth Temperatures and Gas-Formation Potential


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Origin
2.2. Detection Techniques
2.2.1. Biochemical Differentiation
2.2.2. MALDI-TOF
2.2.3. Method Comparison
2.3. Temperature Comparison
2.4. PH-Value
2.5. Gas Pressure Measurement
2.6. Statistics
3. Results
3.1. Identification
3.1.1. Method Comparison
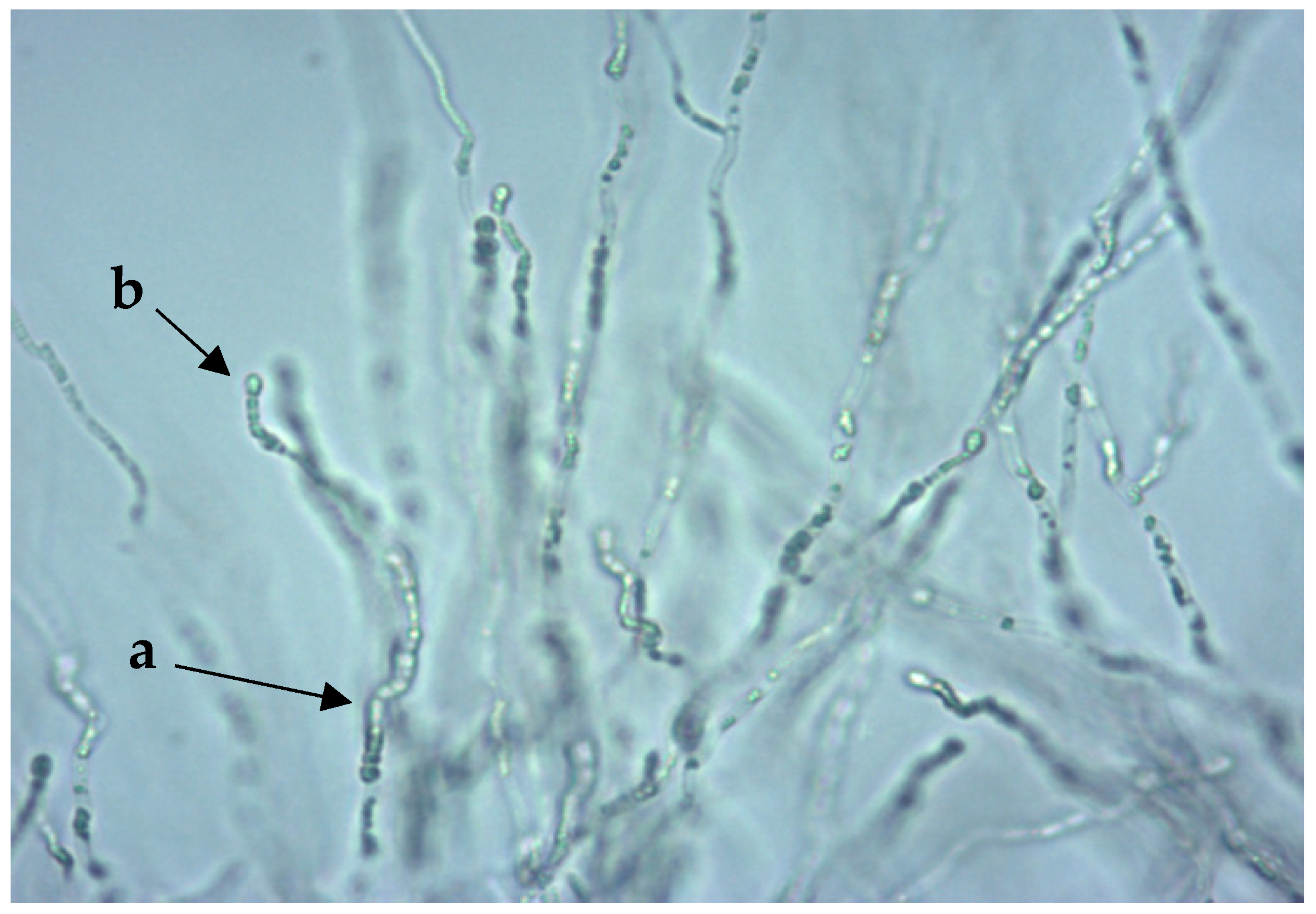
3.1.2. Biochemical Reactions
Metabolization of N-Acetylglucosamine (NAG)
Metabolization of Lactic Acid (LAT)
3.2. Temperature Comparison
3.3. pH-Value in Liquid Swine Diets
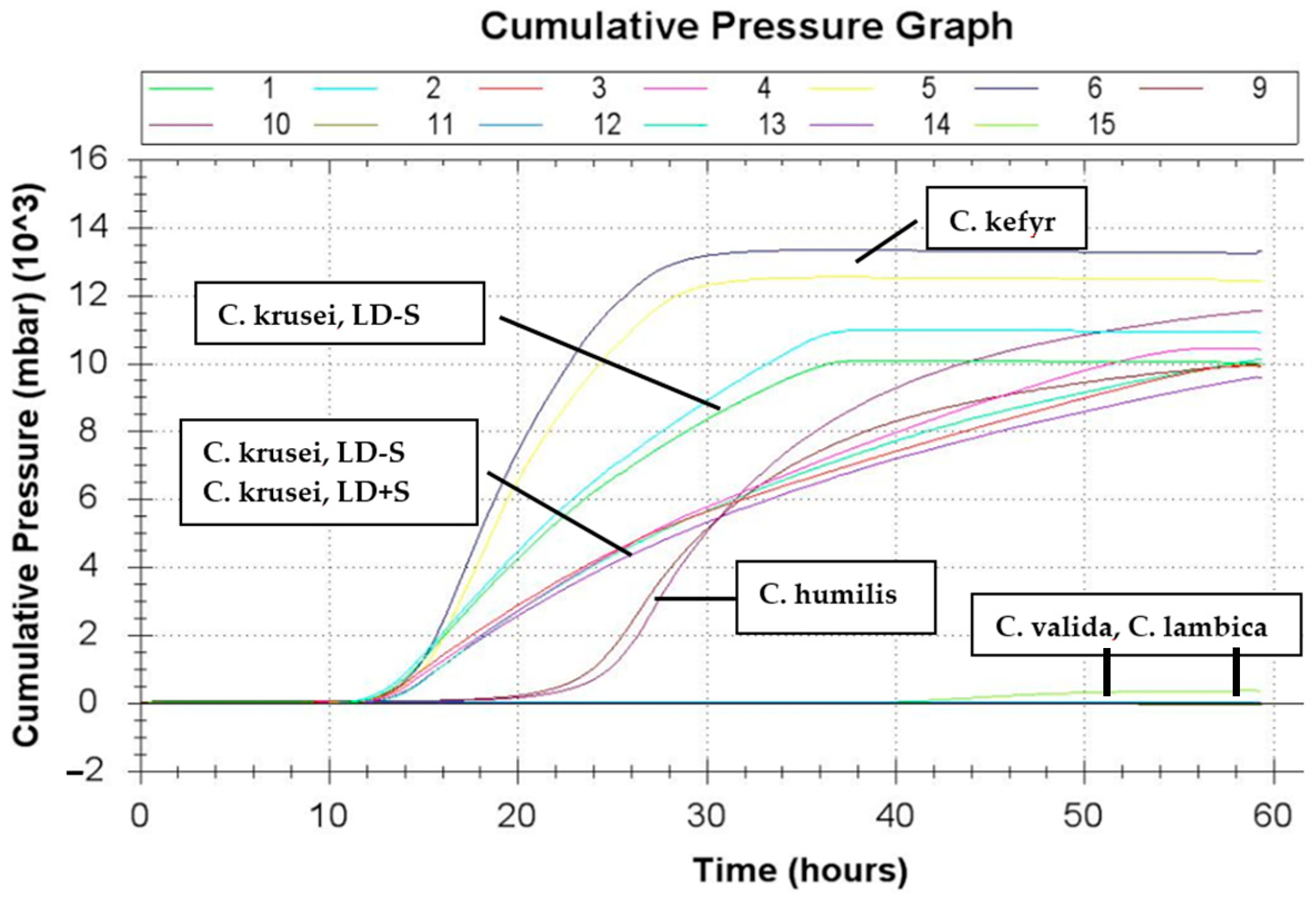
3.4. Gas Production
4. Discussion
4.1. Identification
4.1.1. C. krusei
4.1.2. C. lambica
4.1.3. Yeasts from Liquid Diets for Pigs
4.1.4. Method Comparison
ID32C
Selected Biochemical Reactions of the ID32C-Test
4.2. Temperature
4.3. pH-Value
4.4. Gas Production
4.5. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fiedler, B. Hefen: Lebensmittelassoziierte Mikroorganismen Fermentationsleistung und Verderbspotential, 2nd ed.; Behr’s Verlag GmbH: Hamburg, Germany, 2017; pp. 7, 12, 56. [Google Scholar]
- Büchl, N.R. Identifizierung von Hefen durch Fourier-transform Infrarotspektroskopie und künstlich neuronale Netzen. Ph.D. Thesis, Faculty Sciene Center Weihenstephan, Weihenstephan, Germany, 19 June 2009. Available online: http://mediatum.ub.tum.de/?id=684162 (accessed on 2 December 2020).
- Stalljohann, G. Tiergesundheitsmanagement auf betrieblicher Ebene. In Tiergesundheit Schwein; Brede, W., Blaha, T., Hoy, S., Eds.; DLG-Verlags-GmbH: Frankfurt am Main, Germany, 2010; pp. 236–241. [Google Scholar]
- Santos, M.C.; Golt, C.; Joerger, R.D.; Mechor, G.D.; Murao, G.B.; Kung, L., Jr. Identification of the major yeasts isolated from high moisture corn and corn silages in the United States using genetic and biochemical methods. J. Dairy Sci. 2017, 100, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, M. Hygiene in Flüssigfütterungsanlagen. In Themen zu Tierernährung, Fachtagung Deutsche Vilomix Tierernährung GmbH 2004/2005. 2005. Available online: http://docplayer.org/32990535-Hygiene-in-fluessigfuetterungsanlagen.html (accessed on 2 December 2020).
- Olstorpe, M.; Lyberg, K.; Lindberg, J.E.; Schnürer, J.; Passoth, V. Population diversity of yeasts and lactic acid bacteria in pig feed fermented with whey, wet wheat distillers’ grains or water at different temperatures. Appl. Environ. Microbiol. 2008, 74, 1696–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, P.H. Fermented liquid feed for pigs. CAB Rev. Perspect. Agric. Veter- Sci. Nutr. Nat. Resour. 2008, 3. [Google Scholar] [CrossRef]
- Jörling, U. Untersuchung zum Ansatzverhalten und zur Mikroflora im Kot von Mastschweinen unter den Bedingungen einer Konditionsbasierten Energie- und Nährstoffversorgung und Ad-libitum-fütterungsbedingungen. Master Thesis, University of Veterinary Medicine Hannover, Hanover, Germany, 2017. [Google Scholar]
- Jensen, B.B. The impact of feed additives on the microbial ecology of the gut in young pigs. J. Anim. Feed Sci. 1998, 7, 45–64. [Google Scholar] [CrossRef]
- Canibe, B.; Jensen, B.B. Fermented liquid feed—Microbial and nutritional aspects and impact on enteric diseases in pigs. Anim. Feed Sci. Technol. 2012, 173, 17–40. [Google Scholar] [CrossRef]
- Missotten, J.A.; Michiels, J.; Ovyn, A.; De Smet, S.; Dierick, N.A. Fermented liquid feed for pigs. Arch. Anim. Nutr. 2010, 64, 437–466. [Google Scholar] [CrossRef] [Green Version]
- Canibe, N.; Jensen, B.B. Fermented and nonfermented liquid feed to growing pigs: Effect on aspects of gastrointestinal ecology and growth performance. J. Anim. Sci. 2003, 81, 2019–2031. [Google Scholar] [CrossRef]
- Johannsen, U.; Strijkstra, G.; Klarmann, D.; Janthur, I. Untersuchungen zum Enterohämorrhagischen Syndrom (EHS) der Schweine. Prakt. Tierarzt. 2000, 12, 440–451. [Google Scholar]
- Reiner, G. Enterohaemorrhagisches Syndrom. In Krankes Schwein—Kranker Bestand; Eugen Ulmer Verlag: Stuttgart, Germany, 2015; pp. 132–134. [Google Scholar]
- Straw, B.; Dewey, C.; Kober, J.S.; Henry, C. Factors associated with death due to hemorrhagic bowel syndrome in two large commercial swine farms. J. Swine Health Prod. 2002, 10, 75–79. [Google Scholar]
- Missotten, J.; Michiels, A.; Degroote, J.; De Smet, S. Fermented liquid feed for pigs: An ancient technique for the future. J. Anim. Sci. Biotechnol. 2015, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Kamphues, J.; Wolf, P.; Coenen, M.; Eder, K.; Iben, C.; Kienzle, E.; Liesegang, A.; Männer, K.; Zebeli, Q.; Zentek, J. Beurteilung der Mikrobiologischen Qualität von FM. In Supplemente zur Tierernährung für Studium und Praxis, 12th ed.; Schaper Verlag: Hannover, Germany, 2014; p. 206. [Google Scholar]
- Nagel, M. Mikrobiologische Vorgänge in Flüssigfutter für Schweine. In Handbuch der Tierischen Veredlung; Kamlage-Verlag: Osnabrück, Germany, 1998; Volume 23, pp. 189–200. [Google Scholar]
- Grüne Broschüre 2020 Das geltende Futtermittelrecht. (Band FZ: Futtermittelzusatzstoffe), 2nd ed.; Allround Media Service e. K: Rheinbach, Germany, 2019; pp. 310–315, 322–323, 326–327, 338–339, 352–353.
- Rajkowska, K.; Kunicka-Styczyńska, A. Typing and virulence factors of food-borne Candida spp. isolates. Int. J. Food Microbiol. 2018, 279, 57–63. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherrington, S.L.; Sorsby, E.; Mahtey, N.; Kumwenda, P.M.; Lenardon, D.; Brown, I.E.; Ballou, R.D.; MacCallum, M.; Hall, R.A. Adaptation of Candida albicans to environmental pH induces cell wall remodelling and enhances innate immune recognition. PLoS Pathog. 2017, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puligundla, P.; Mok, C. Potential applications of nonthermal plasmas againstbiofilm-associated micro-organisms in vitro. J. Appl. Microbiol. 2017, 122, 1134–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Jugsharan, S.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Park, J.H.; Oh, J.; Cho, S.; Koo, J.; Park, I.C.; Kim, J.; Park, S.; Choi, J.S.; Shin, S.Y.; et al. Evaluation of a new matrix-assisted laser desorption/ionization time-of-flight mass spectrometry system for the identification of yeast isolation. J. Clin. Lab. Anal. 2018, 33, e22685. [Google Scholar] [CrossRef] [Green Version]
- Olstorpe, M.; Axelsson, L.; Schnürer, J.; Passoth, V. Effect of starter culture inoculation on feed hygiene and microbial population development in fermented pig feed composed of a cereal grain mix with wet wheat distillers’ grain. J. Appl. Microbiol. 2010, 108, 129–138. [Google Scholar] [CrossRef]
- Gori, K.; Kryger Bjørklund, M.; Canibe, N.; Petersen, A.O.; Jespersen, L. Occurrence and Identification of Yeast Species in Fermented Liquid Feed for Piglets. Microbial. Ecol. 2011, 61, 146–153. [Google Scholar] [CrossRef]
- Urubschurow, V.; Janczyk, P.; Pieper, R.; Souffrant, W.B. Biological diversity of yeasts in the gastrointestinal tract of weaned piglets under different farm conditions. FEMS Yeast Res. 2009, 8, 1349–1356. [Google Scholar] [CrossRef] [Green Version]
- Halm, M.; Hornbaek, T.; Arneborg, N.; Sefa-Dedeh, S.; Jespersen, L. Lactic acid tolerance determined by measurement of intracellular pH of single cells of Candida krusei and Saccharomyces cerevisiae isolated from fermented maize dough. Int. J. Food Microbiol. 2004, 94, 97–103. [Google Scholar] [CrossRef]
- Spicher, G.; Schröder, R. Die Mikroflora des Sauerteiges. Z. Lebensm. Unters. Forsch. 1980, 170, 119–123. [Google Scholar] [CrossRef]
- Silva, S.; Rodrigues, C.F.; Araùjo, D.; Rodrigues, M.E.; Henriques, M. Candida species biofilms’ antifungal resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Douglass, A.P.; Offei, B.; Braun-Galleani, S.; Coughlan, A.Y.; Martos, A.A.R. Population genomics shows no distinction between pathogenic Candida krusei and environmental Pichia kudriavzevii: One species, four names. PLoS Pathog. 2018, 14, e1007138. [Google Scholar] [CrossRef] [PubMed]
- Middelhoven, W.J.; de Jong, I.M.; de Winter, M. Yeasts and fungi occurring in ensiled whole-crop maize a7d other ensiled vegetable crops. Antonie Van Leeuwenhoek 1990, 57, 153–158. [Google Scholar] [CrossRef]
- Vervaeke, S.; Vandamme, K.; Boone, E.; De Laere, E.; Swinne, D.; Surmont, I. A case of Candida lambica fungemia misidentified as Candida krusei in an intravenous drug abuser. Med. Mycol. 2008, 46, 853–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plumed-Ferrer, C.; von Wright, A. Antimicrobial activity of weak acids in liquid feed fermentations, and its effects on yeasts and lactic acid bacteria. J. Sci. Food Agric. 2011, 91, 1032–1040. [Google Scholar] [CrossRef]
- Kuthan, M.; Devaux, F.; Janderová, B.; Jacq, C.I.; Palkov, Z. Domestication of wild Saccharomyces cerevisiae is accompanied by changes in gene expression and colony morphology. Mol. Microbiol. 2003, 47, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, S.G.; Shafik, A.; Hakim, S.T.; Anjum, Y.; Kazm, S.U. Effect of growth media, pH and temperature on yeast to hyphal transition in Candida albicans. Open J. Med. Microbiol. 2013, 3, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.; Schwaiger, K. Spezielle Mykologie und Prototheken. In Tiermedizinische Mikrobiologie, Infektions- und Seuchenlehre; Selbitz, H.-J., Truyen, U., Valentin-Weigand, P., Eds.; Enke Verlag: Stuttgart, Germany, 2015; p. 365. [Google Scholar]
- Pohlmann, L.M.; Chengappa, M.M. Yeasts—Cryptococcus, Malassezia and Candida. In Veterinary Microbiology, 3rd ed.; McVey, S.D., Kennedy, M., Chengappa, M.M., Eds.; Willey-Blackwell: Ames, IA, USA, 2013; pp. 319–320. [Google Scholar]
- mycobank.org. Available online: www.mycobank.org/BioloMICS.aspx?TableKey=14682616000000067&Rec=39099 (accessed on 14 August 2020).
- mycobank.org. Available online: www.mycobank.org/name/Geotrichumfragrans (accessed on 14 August 2020).
- Canibe, N.; Pedersen, A.O.; Jensen, B.B.; Jespersen, L. Microbiological and biochemical characterization of fermented liquid feed samples from 40 Danish farms. Livestock Sci. 2010, 134, 158–161. [Google Scholar] [CrossRef]
- Stefaniuk, E.; Baraniak, A.; Fortuna, M.; Hryniewicz, W. Usefulness of CHROMagar Candida medium, biochemical methods—API ID32C and VITEK 2 Compact and two MALDI-TOF MS Systems for Candida spp. Identification. Polish J. Microbiol. 2016, 65, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Ellis, C.; Lee, R.; Stamper, P.D.; Zhang, S.X.; Carroll, K.C. Prospective Evaluation of a Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry System in a Hospital Clinical Microbiology Laboratory for Identification of Bacteria and Yeasts: A Bench-by-Bench Study for Assessing the Impact on Time to Identification and Cost-Effectiveness. J. Clin. Microbiol. 2012, 50, 3301–3308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustini, B.; Silva, L.P.; Bloch, C. Evaluation of MALDI-TOF mass spectrometry for identification of environmental yeasts and development of supplementary database. Appl. Microbiol. Biotechnol. 2014, 98, 5645–5654. [Google Scholar] [CrossRef] [PubMed]
- Vlek, A.; Koecka, A.; Khayhan, K.; Theelen, B.; Groenewald, M.; Boel, E.; Multicenter Study Group; Boekhout, T. Interlaboratory comparison of sample preparation methods, database expansions, and cutoff values for identification of yeasts by matrix-assisted laser desorption ionization-time of flight mass spectrometry using a yeast test panel. J. Clin. Microbiol. 2014, 52, 3023–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koht, P.D.; Farrance, C.E. Evaluation of MALDI-TOF Mass Spectrometry for Identification of Yeasts Commonly Found During Environmental Monitoring. Am. Pharm. Rev. 2016. Available online: https://www.americanpharmaceuticalreview.com/Featured-Articles/331621-Evaluation-of-MALDI-TOF-Mass-Spectrometry-for-Identification-of-Yeasts-Commonly-Found-During-Environmental-Monitoring/ (accessed on 3 December 2020).
- Inokuma, K.; Matsuda, M.; Sasaki, D.; Hasunuma, T.; Kondo, A. Widespread effect of N-acetyl-d-glucosamine assimilation on the metabolisms of amino acids, purines, and pyrimidines in Scheffersomyces stipitis. Microb. Cell Factoriest 2018, 17, 153. [Google Scholar] [CrossRef] [PubMed]
- Dellaglio, F.; Torriani, S. DNA-DNA homology, physiological characteristics and distribution of lactic acid bacteria isolated from maize silage. J. Appl. Bacteriol. 1986, 60, 83–92. [Google Scholar] [CrossRef]
- Drihuis, F.; Oude Elferink, S.J.W.H. The impact of the quality of silage on animal health and food safety: A review. Vet. Q. 2000, 22, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Ponomarova, O.; Gabrielle, N.; Séverin, D.C.; Mülleder, M.; Zimgibl, K.; Bulyha, K.; Andrejev, S.; Kafkia, E.; Typas, A.; Sauer, U.; et al. Yeast creates a niche for symbiotic lactic acid, bacteria through nitrogen overflow. Cell Syst. 2017, 5, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Shin, J.H.; Choi, M.J.; Won, E.J.; Kee, S.J.; Kim, S.H.; Shin, M.G.; Suh, S.P. Comparison of the Bruker Biotyper and VITEK MS Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry Systems Using a Formic Acid Extraction Method to Identify Common and Uncommon Yeast Isolates. Ann. Lab. Med. 2017, 37, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Pavlovica, M.; Mewesa, A.; Maggipintoa, M.; Schmidta, W.; Messelhäußera, U.; Balsliemkeb, J.; Hörmansdorfer, S.; Buscha, U.; Huber, I. MALDI-TOF MS based identification of food-borne yeast isolates. J. Microbiol. Methods 2014, 106, 123–128. [Google Scholar] [CrossRef]
- Latouche, G.N.; Daniel, H.-M.; Lee, O.K.C.; Mitchell, T.G.; Sorrell, T.C.; Meyer, W. Comparison of use of phenotypic and genotypic characteristics for identification of species of the anamorph genus Candida and related teleomorph yeast species. J. Clin. Microbiol. 1997, 35, 3171–3180. [Google Scholar] [CrossRef] [Green Version]
- Keller, B.; Scheibl, P.; Bleckmann, E.; Hoedemaker, M. Differenzierung von Hefen in Mastitismilch. Mycoses 2000, 43 (Suppl. 1), 17–19. [Google Scholar] [PubMed]
- Hoedemaker, M.; Schmidt, A.; Keller, B.; Bleckmann, E.; Böhm, K.H. Isolation von Hefen aus Milch von Kühen mit Mastitis und aus Tupferproben von der Melkanlage. Prakt. Tierarzt. 2006, 87, 890–898. [Google Scholar]
- Buchan, B.W.; Ledeboer, N.A. Advances in identification of clinical yeast isolates by use of Matrix-Assisted Laser Desorption Ionization-Time of Flight mass spectrometry. J. Clin. Microbiol. 2013, 51, 1359–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunte, S. The Fermentation of Liquid Diets on Feeding Concept in Fattening Pigs—Potentials, but also Risks from the View of Animal Nutrition and Veterinary Medicine. Master Thesis, University of Veterinary Medicine Hannover, Hanover, Germany, 2018. [Google Scholar]
- Suutari, M.; Liukkonen, K.; Laakso, S. Temperature adaptation in yeasts: The role of fatty acids. J. Gen. Microbiol. 1990, 136, 1469–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margesin, R.; Gander, S.; Zacke, G.; Gounot, A.M.; Schinner, F. Hydrocarbon degradation and enzyme activities of cold-adapted bacteria and yeasts. Extremophiles 2003, 7, 451–458. [Google Scholar] [CrossRef]
- Rüschendorf, A. Medizinische Mykologie. Bestimmung und Differenzierung von Sproßpilzen, Schimmelopilzen, Dermatophyten und dimorphen Pilzen, 3rd ed.; Lehmanns Media: Berlin, Germany, 2014; pp. 30, 33, 37, 60. ISBN 978-3-86541-629-2. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Liquid Diet | Often Isolated Yeast Species in % |
|---|---|
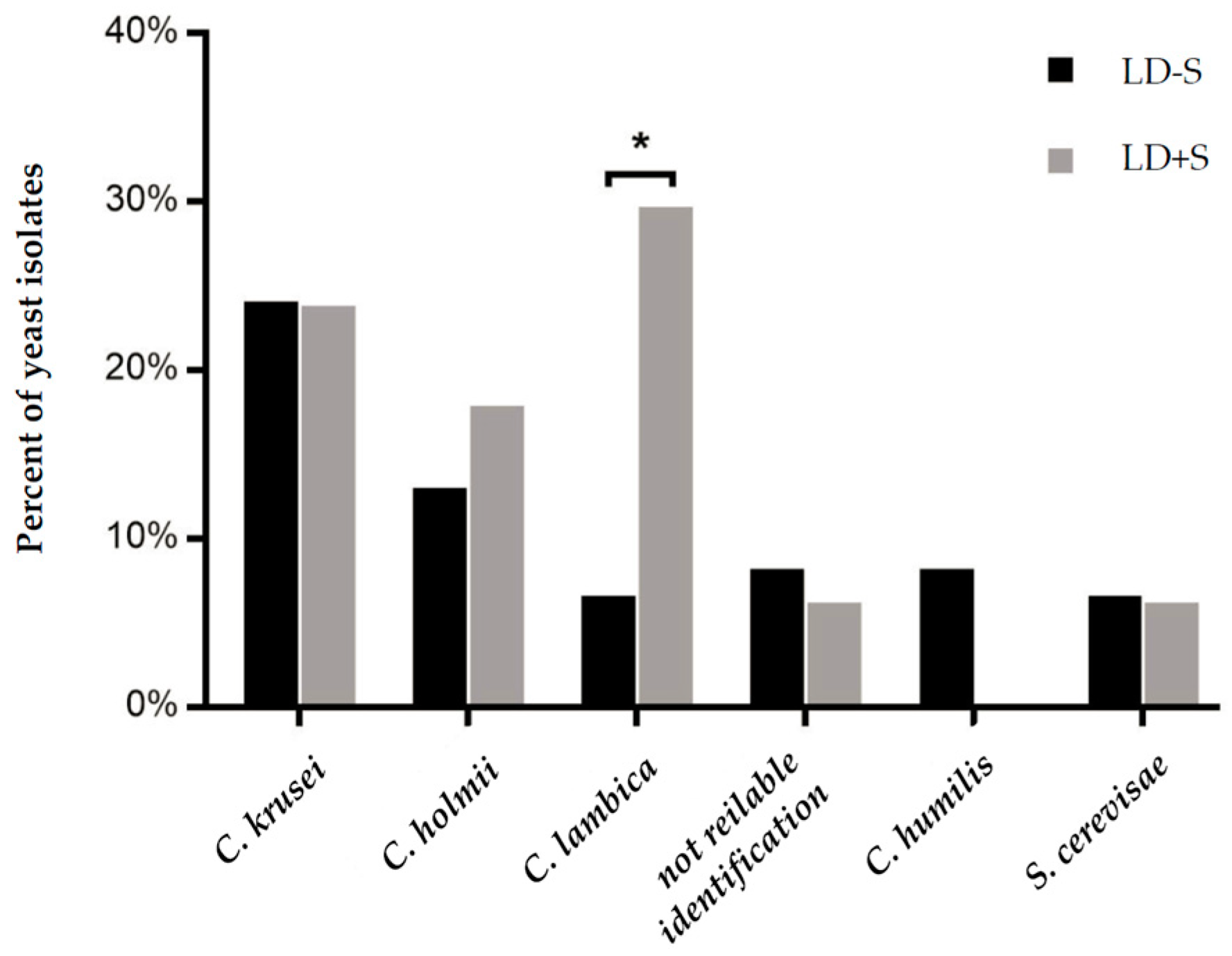
| LD – S (n = 63) | C. krusei (23.8), C. holmii (12.7), C. humilis (7.9), isolates not identified (7.9), C. lambica (6.3), S. cerevisiae (6.3), G. silvicola (6.3) |
| LD + S (n = 17) | C. lambica (29.4), C. krusei (23.5), C. holmii (17.6), C. pelliculosa (11.8), S. cerevisiae (5.9), C. valida (5.9), isolates not identified (5.9) |
| Total (n = 80) | C. krusei (23.75), C. holmii (13.75), C. lambica (11.25), isolates not identified (7.5), C. humilis (6.25), S. cerevisiae (6.25), G. silvicola (6.25) |
| Evaluation Score of MALDI-TOF | MALDI-EX | MALDI-DS | ID-32C | |||
|---|---|---|---|---|---|---|
| Highly probable species identification (>2.3) | 38.75 (n = 31) | 78.75 *,a (n = 63) | 5.0 (n = 4) | 37.5 *,b (n = 30) | 47.5 (n = 38) | 75.0 **,a (n = 60) |
| Probable species identification (2.0–2.29) | 40.0 (n = 32) | 32.5 (n = 26) | 27.5 (n = 22) | |||
| Probable genus identification (1.7–1.99) | 7.5 (n = 6) | 21.25 *** (n = 17) | 33.75 (n = 27) | 62.5 *** (n = 70) | 3.75 (n = 3) | 25.0 *** (n = 20) |
| No identification (<1.7) | 13.75 (n = 11) | 28.75 (n = 23) | 21.25 (n = 17) | |||
| Number of Isolates | MALDI-EX | ID 32C |
|---|---|---|
| 5 | Candida humilis | Candida holmii |
| 2 | Candida pararugosa | Candida rugosa |
| 1 | No identification | Rhodotorula mucilaginosa |
| 1 | Trichosporon coremiiforme | Cryptococcus curvatus |
| 1 | Trichosporon laibachii | Cryptococcus laurentii |
| MALDI-EX | ID32C | |||
|---|---|---|---|---|
| Diagnosis | Score | Diagnosis | Identification | % |
| C. pararugosa | 1.88 | C. rugosa | very good | 99.8 |
| Sap. suaveolens | 1.88 | Geotrichum spp. | very good | 99.7 |
| P. occidentalis | 1.83 | C. krusei | very good | 99.7 |
| C. holmii | 1.97 | C. holmii | very good | 99.2 |
| C. pararugosa | 1.79 | C. rugosa | very good | 99.5 |
| S. cerevisiae | 1.98 | S. cerevisiae | very good | 99.7 |
| Yeast Isolates | No Growth at 37 °C | Better Growth at 25 °C | 25 °C = 37 °C | Better Growth at 37 °C |
|---|---|---|---|---|
| LD − S (n = 63) | 10 (15.9%) | 24 (38.1%) | 13 (20.6%) | 16 (25.4%) |
| LD + S (n = 17) | 4 (23.5%) | 9 (52.9%) | 1 (5.9%) | 3 (17.6%) |
| Total (n = 80) | 14 (17.5%) | 33 (41.25%) | 14 (17.5%) | 19 (23.75%) |
| Sample | Number | <100 mbar | >800 mbar |
|---|---|---|---|
| LD − S | n = 32 | 19 (59.4%) | 13 (40.6%) |
| LD + S | n = 8 | 7 (87.5%) | 1 (12.5%) |
| total | n = 40 | 26 (65.0%) | 14 (35.0%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keller, B.; Kuder, H.; Visscher, C.; Siesenop, U.; Kamphues, J. Yeasts in Liquid Swine Diets: Identification Methods, Growth Temperatures and Gas-Formation Potential. J. Fungi 2020, 6, 337. https://doi.org/10.3390/jof6040337
Keller B, Kuder H, Visscher C, Siesenop U, Kamphues J. Yeasts in Liquid Swine Diets: Identification Methods, Growth Temperatures and Gas-Formation Potential. Journal of Fungi. 2020; 6(4):337. https://doi.org/10.3390/jof6040337
Chicago/Turabian StyleKeller, Birgit, Henrike Kuder, Christian Visscher, Ute Siesenop, and Josef Kamphues. 2020. "Yeasts in Liquid Swine Diets: Identification Methods, Growth Temperatures and Gas-Formation Potential" Journal of Fungi 6, no. 4: 337. https://doi.org/10.3390/jof6040337