Fungal Pigments: Potential Coloring Compounds for Wide Ranging Applications in Textile Dyeing

 ,
,  ,
,
Abstract
:1. Introduction
2. Historical Note on Pigments
3. Ecology of Fungal Pigments
4. Fungal Pigments
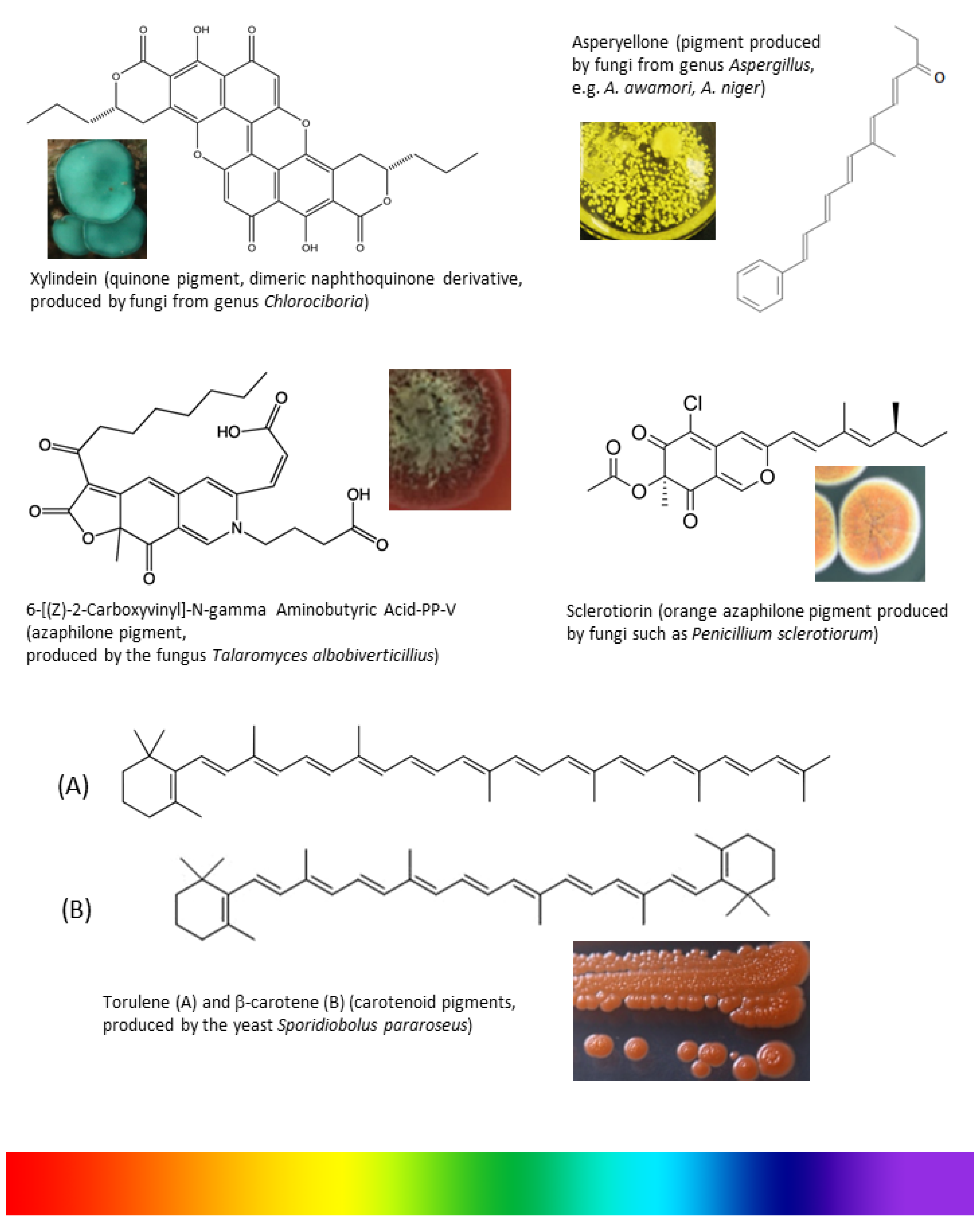
4.1. Carotenoids
4.2. Polyketides
4.3. Anthraquinones
4.4. Hydroxyanthraquinone
4.5. Naphthoquinones
4.6. Azaphilones
5. Fungal Dyes for Textile Applications
6. Toxicity Testing for Fungal Pigments
7. Biotechnology Ways for Enhanced Production
7.1. Genetic Manipulation
7.2. Agro-Waste for Clean-Up Production
8. Limits, Challenges, and Future Scope for the Dyeing of Fungal Pigments
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wrolstad, R.E.; Culver, C.A. Alternatives to those artificial FDC food colorants. Annu. Rev. Food Sci. Technol. 2012, 3, 59–77. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.; Saurabh, K.K.; Kamlesh, K.; Mali, M.K. Studies on synthesis of novel low molecular weight anthraquinone disperse dyes and their application on polyester and nylon. J. Ind. Chem. Soc. 2012, 89, 789–795. [Google Scholar]
- Lebeau, J.; Venkatachalam, M.; Fouillaud, M.; Petit, T.; Vinale, F.; Dufossé, L.; Yanis, C. Production and new extraction method of polyketide red pigments produced by Ascomycetes fungi from terrestrial and marine habitats. J. Fungi 2017, 3, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, G.; Mishra, T.; Deshmukh, S.K. Fungal pigments: Overview. In Developments in Fungal Biology and Applied Mycology; Satyanarayana, T., Deshmukh, S.K., Johri, B.N., Eds.; Springer: Singapore, 2017. [Google Scholar]
- Eisenman, H.C.; Casadevall, A. Synthesis and assembly of fungal melanin. Appl. Microbiol. Biotechnol. 2012, 93, 931–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, S.C.; Tudor, D.; Zhang, W.R.; Ng, S.; Copper, P.A. Ability of three yellow pigment producing fungi to colour wood under controlled conditions. Int. Wood Prod. J. 2014, 5, 103–107. [Google Scholar] [CrossRef]
- Bechtold, T.; Mussak, R. Hanbook of Natural Colorants; John Wiley and Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Yusof, M.; Shabbir, M.; Mohammad, F. Natural colorants: Historical, processing and sustainable prospects. Nat. Prod. Bioprospect. 2017, 7, 123–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, D.J. Is there a future for natural dyes? Color. Technol. 2008, 27, 18–25. [Google Scholar] [CrossRef]
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R.; Venil, C.K.; Dufossé, L. Multifaceted applications of microbial pigments: Current knowledge, challenges and future directions for public health implications. Microorganisms 2019, 7, 186. [Google Scholar] [CrossRef] [Green Version]
- Arifeen, M.Z.; Ma, Y.N.; Xue, Y.R.; Liu, C.H. Deep-sea fungi could be the new arsenal for bioactive compounds. Mar. Drug 2020, 18, 9. [Google Scholar] [CrossRef] [Green Version]
- Hirot, J.; Maksimenka, K.; Reichert, M.; Perovic-Ottstadt, S.; Lin, W.H.; Wray, V.; Steube, K.; Schaumann, K.; Weber, H.; Proksch, P.; et al. New natural products from the sponge derived fungus Aspergillus niger. J. Nat. Prod. 2004, 67, 1532–1543. [Google Scholar]
- Dufossé, L.; Fouillaud, M.; Caro, Y.; Mapari, S.A.S.; Sutthiwong, N. Filamentous fungi are large scale producers of pigments and colorants for the food industry. Curr. Opin. Biotechnol. 2014, 26, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Mapari, S.A.S.; Thrane, U.; Meyer, A.S. Fungal polyketide azaphilone pigments as future natural food colorants? Trends Biotechnol. 2010, 28, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Pfander, H. Carotenoids: An overview. Methods Enzymol. 1992, 213, 3–13. [Google Scholar]
- Avalos, J.; Limon, M.C. Biological roles of fungal carotenoids. Curr. Genet. 2015, 61, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Mapari, S.A.S.; Nielsen, K.F.; Larsen, T.O.; Frisvad, J.C.; Meyer, A.S.; Thrane, U. Exploring fungal biodiversity for the production of water soluble pigments as potential natural colorants. Curr. Opin. Biotechnol. 2005, 16, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Rohdich, F.; Kis, K.; Bacher, A.; Eisenreich, W. The non-mevalonate pathway of isoprenoids: Genes, enzymes and intermediates. Curr. Opin. Chem. Biol. 2001, 5, 535–540. [Google Scholar] [CrossRef]
- Zhang, C. Biosynthesis of carotenoids and apocarotenoids by microorganisms and their industrial potential. In Progress in Carotenoid Research; BioTrans: Singapore, 2018. [Google Scholar]
- Bhosale, P.; Larson, A.J.; Bernstein, P.S. Factorial analysis of tricarboxylic acid cycle intermediates for optimization of zeaxanthin production from Flavobacterium multivorum. J. Appl. Microbiol. 2004, 96, 623–629. [Google Scholar] [CrossRef]
- Ogbonna, C. Production of food colourants by filamentous fungi. Afr. J. Microbiol. Res. 2016, 10, 960–971. [Google Scholar]
- Rao, M.P.N.; Xiao, M.; Li, W.-J. Fungal and bacterial pigments: Secondary metabolites with wide applications. Front. Microbiol. 2017, 8, 1113. [Google Scholar]
- Mapari, S.A.S.; Meyer, A.S.; Thrane, U.; Frisvad, J.C. Identification of potentially safe promising fungal cell factories for the production of polyketide natural food colorants using chemotaxonomic rationale. Microb. Cell Factories 2009, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Atalla, M.M.; Elkhrisy, E.A.M.; Asem, M.A. Production of textile reddish brown dyes by fungi. Malays. J. Microbiol. 2011, 33, 40. [Google Scholar]
- Babula, P.; Adam, V.; Havel, L.; Kizek, R. Noteworthy secondary metabolites naphthoquinones–their occurrence, pharmacological properties and analysis. Curr. Pharm. Anal. 2009, 5, 47–67. [Google Scholar] [CrossRef]
- Yu, M.L.; Li, Y.X.; Banakar, S.P.; Liu, L.; Shao, C.L.; Li, Z.Y.; Wang, C.Y. New metabolites from the co-culture of marine-derived actinomycete Streptomyces rochei MB037 and fungus Rhinocladiella similis 35. Front. Microbiol. 2019, 10, 915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufossé, L. Pigments, microbial. In Encyclopedia of Microbiology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Chen, W.; He, Y.; Zhou, Y.; Shao, Y.; Feng, Y.; Li, M.; Chen, F. Edible filamentous fungi from the species monascus: Early traditional fermentations, modern molecular biology, and future genomics. Compr. Rev. Food Sci. Food Saf. 2015, 14, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.L.; Shao, Y.C.; Chen, F.S. Monascus pigments. Appl. Microbiol. Biotechnol. 2012, 96, 1421–1440. [Google Scholar] [CrossRef]
- Chen, W.; Feng, Y.; Molnar, I.; Chen, F. Nature and nurture: Confluence of pathway determinism with metabolic and chemical serendipity diversifies Monascus azaphilone pigments. Nat. Prod. Rep. 2019, 36, 561–572. [Google Scholar] [CrossRef]
- Lee, C.L.; Wen, J.Y.; Hsu, Y.W.; Pan, T.M. The blood lipid regulation of Monascus-produced monascin and ankaflavin via the suppression of low-density lipoprotein cholesterol assembly and stimulation of apolipoprotein A1 expression in the liver. J. Microbiol. Immunol. 2018, 51, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Lachenmeier, D.W.; Monakhova, Y.B.; Kuballa, T.; LobellBehrends, S.; Maixner, S.; Kohl-Himmelseher, M.; Waldner, A.; Steffen, C. NMR evaluation of total statin content and HMG-CoA reductase inhibition in red yeast rice (Monascus spp.) food supplements. Chin. Med. 2012, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Chen, R.; Liu, Q.; He, Y.; He, K.; Ding, X.; Kang, L.; Guo, X.; Xie, N.; Zhou, Y.; et al. Orange, red, yellow: Biosynthesis of azaphilone pigments in monascus fungi. Chem. Sci. 2017, 8, 4917–4925. [Google Scholar] [CrossRef] [Green Version]
- Schor, R.; Cox, R. Classical fungal natural products in the genomic age: The molecular legacy of Harold Raistrick. Nat. Prod. Rep. 2018, 35, 230–256. [Google Scholar] [CrossRef] [Green Version]
- Gessler, N.N.; Egorova, A.S.; Belozerskaya, T.A. Fungal anthraquinones. Appl. Biochem. Microbiol. 2013, 49, 85–99. [Google Scholar] [CrossRef]
- Yang, X.M.; Li, J.S.; Li, Q.Q.; Huang, G.X.; Yan, L.J. Evaluation of the potential toxicity of anthraquinone derivatives in Chinese herbal medicines by the resonance light scattering spectrum. Asian J. Chem. 2011, 23, 3631–3634. [Google Scholar]
- Fouillaud, M.; Venkatachalam, M.; Girard-Valenciennes, E.; Caro, Y.; Dufossé, L. Anthraquinones and derivatives from marine derived fungi: Structural diversity and selected biological activities. Mar. Drugs 2016, 14, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; He, Z.; Wu, J.; Yuan, J.; Wen, W.; Hu, Y.; Jiang, Y.; Lin, C.; Zhang, Q.; Lin, M.; et al. A marine anthraquinone SZ-685C overrides Adriamycin resistance in breast cancer cells through suppressing Akt signalling. Mar. Drugs 2012, 10, 694–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Nong, X.; Ren, Z.; Wang, J.; Zhang, X.; Qi, S. Anti-HSV1, antioxidant and antifouling phenolic compounds from the deep sea derived fungus Aspergillus versicolor SCSIO41502. Bioorganic Med. Chem. Lett. 2017, 15, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Hou, X.M.; Li, Z.Y.; Cao, F.; Zhang, Y.H.; Yu, J.Y.; Zhao, D.L.; Shao, C.L.; Wang, C.Y. Harzianumnones A and B: Two hydroxyanthraquinones from the coral derived fungus Trichoderma harzianum. RSC Adv. 2018, 8, 27596–27601. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Y.; Tao, L.Y.; Liang, Y.J.; Chen, L.M.; Mi, Y.J.; Zheng, L.S.; Wang, F.; She, Z.G.; Lin, Y.C. Anthracenedione derivatives as anticancer agents isolated from secondary metabolites of the mangrove endophytic fungi. Mar. Drugs 2010, 8, 1469–1481. [Google Scholar] [CrossRef]
- Awaad, A.S.; Al-Zaylaee, H.M.; Alqasoumi, S.I.; Zain, M.E.; Aloyan, E.M.; Alafeefy, A.M.; Awad, E.S.; El-Meligy, R.M. Anti-leishmanial activities of extracts and isolated compounds from Drechslera rostrata and Eurotium tonpholium. Phytother. Res. 2014, 28, 774–780. [Google Scholar] [CrossRef]
- Wellington, K.W. Understanding cancer and the anticancer activities of naphthoquinones—A review. RSC Adv. 2015, 5, 20309–20338. [Google Scholar] [CrossRef]
- Newman, A.G.; Townsend, C.A. Molecular characterization of the cercosporin biosynthetic pathway in the fungal plant pathogen Cercospora nicotianae. J. Am. Chem. Soc. 2016, 138, 4219–4228. [Google Scholar] [CrossRef] [Green Version]
- Macicella, I.A.; Coatti, G.C.; Biazi, B.I.; Zanetti, T.A.; Baranoski, A.; Marques, L.A.; Corveloni, A.C.; Lepri, S.R.; Mantovani, M.S. Molecular pathways related to the control of proliferation and cell death in 786-O cells treated with plumbagin. Mol. Biol. Rep. 2019, 46, 6071–6078. [Google Scholar]
- Li, M.; Kang, L.; Ding, X.; Liu, J.; Liu, Q.; Shao, Y.; Molnar, I.; Chen, F. Monasone naphthoquinone biosynthesis and resistance in Monascus fungi. Mol. Biol. Physiol. 2020, 11, e02676-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, S.C.; Tudor, D.; Cooper, P.A. Utilizing pigment-producing fungi to add commercial value to American beech (Fagus grandifolia). Appl. Microbiol. Biotechnol. 2012, 93, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.; Chen, H.L.; Hinsch, E.; Freitas, S.; Robinson, S. Pigments extracted from the wood staining fungi Chlorociboria aeruginosa, Scytalidium cuboideium and S. ganodermophthorum show potential for use as textile dyes. Color. Technol. 2014, 130, 445–452. [Google Scholar] [CrossRef]
- Medentsev, A.G.; Akimentko, V.K. Naphthoquinone metabolites of the fungi. Phytochemistry 1998, 47, 935–959. [Google Scholar] [CrossRef]
- Coates, C.S.; Ziegler, J.; Manz, K.; Good, J.; Kang, B.; Milikisiyants, S.; Chatterjee, R.; Hao, S.; Golbeck, J.H.; Lakshmi, K.V.J.; et al. The structure and function of quinones in biological solar energy transduction: A cyclic voltammetry, EPR, and hyperfine sub-level correlation (HYSCORE) spectroscopy study of model naphthoquinones. J. Phys. Chem. B 2013, 117, 7210–7220. [Google Scholar]
- Freire, C.P.V.; Ferreira, S.B.; Oliveira, N.S.M.; Matsuura, A.B.J.; Gama, I.L.; Silva, F.C.; Souza, M.C.B.V.; Lima, E.S.; Ferreira, V.F. Synthesis and biological evaluation of substituted α- and β-2,3-dihydrofuran naphthoquinones as potent anticandidal agents. Med. Chem. Commun. 2010, 1, 229–232. [Google Scholar] [CrossRef]
- Ferreira, M.P.S.B.C.; Cardoso, M.F.C.; Silva, F.C.; Ferreira, V.F.; Lima, E.S.; Souza, J.V.B. Antifungal activity of synthetic naphthoquinones against dermatophytes and opportunistic fungi: Preliminary mechanism-of-action tests. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Tao, H.; Chen, W.; Yang, B.; Zhou, X.; Luo, X.; Liu, Y. Recent advancements in the chemistry and biology of azaphilones. RSC Adv. 2020, 10, 10197–10220. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.M.; Yang, S.X.; Qin, J.C. Azaphilones: Chemistry and biology. Chem. Rev. 2013, 113, 4755–4811. [Google Scholar] [CrossRef]
- Osmanova, N.; Schultze, W.; Ayoub, N. Azaphilones: A class of fungal metabolites with diverse biological activities. Phytochem. Rev. 2010, 9, 315–342. [Google Scholar] [CrossRef]
- Chen, M.; Shen, N.X.; Chen, Z.Q.; Zhang, F.M.; Chen, Y. Penicilones A-D, anti-MRSA azaphilones from the marine derived fungus Penicillium janthillum HK1-6. J. Nat. Prod. 2017, 80, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.P.; Wu, Y.R.; Yang, M.H.; Li, T.X.; Wang, X.B.; Zhou, M.M.; Lei, J.L.; Kong, L.Y. Citrifurans A-D four dimeric aromatic polyketides with new carbon skeletons from the fungus Aspergillus sp. Org. Lett. 2017, 19, 4058–4061. [Google Scholar] [CrossRef] [PubMed]
- Kornsakulkarn, J.; Saepua, S.; Suvannakad, R.; Supothina, S.; Boonyuen, N.; Isaka, M.; Prabpai, S.; Kongsaeree, P.; Thongpanchang, C. Cytotoxic tropolones from the fungus Nemania sp. BCC30850. Tetrahedron 2017, 73, 3505–3512. [Google Scholar] [CrossRef]
- Fan, Z.; Sun, Z.H.; Liu, H.X.; Chen, Y.C.; Li, H.H.; Zhang, W.M. Perangustols A and B, a pair of new azaphilone epimers from a marine sediment derived fungus Cladosporium perangustm FS62. J. Asian Nat. Prod. Res. 2016, 18, 1024–1029. [Google Scholar] [CrossRef]
- Arunpanichlert, J.; Rukachaisirikul, V.; Phongpaichit, S.; Supaphon, O.; Sakayaroj, J. Xylariphilone: A new azaphilone derivative from the seagrass derived fungus Xylariales sp. PSU-ES163. Nat. Prod. Res. 2016, 30, 46–51. [Google Scholar] [CrossRef]
- Chen, Y.S.; Cheng, M.J.; Hsiao, Y.; Chan, H.Y.; Hsieh, S.Y.; Chang, C.W.; Liu, T.W.; Chang, H.S.; Chen, I.S. Chemical constituents of the endophytic fungus Hypoxylon sp. 12F0687 isolated from Taiwanese Ilex formosana. Helv. Chim. Acta 2015, 98, 1167–1176. [Google Scholar] [CrossRef]
- Wang, J.; Bai, G.; Liu, Y.; Wang, H.; Li, Y.; Yin, W.; Wang, Y.; Lu, F. Cytotoxic metabolites produced by the endophytic fungus Aspergillus clavatus. Chem. Lett. 2015, 44, 1148–1149. [Google Scholar] [CrossRef]
- Hufendiek, P.; Stoelben, S.S.M.; Kehraus, S.; Merten, N.; Harms, N.; Cruesemann, M.; Arslan, I.; Guetschow, M.; Schneider, T.; Koenig, G.M.; et al. Biosynthetic studies on Acetosellin and structure elucidation of a new acetosellin derivative. Planta Med. 2017, 83, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.J.; Wu, M.D.; Chan, H.Y.; Chang, H.S.; Wu, H.C.; Chen, J.J.; Yuan, G.F.; Weng, J.R.; Chang, C.T.; Lin, H.C.; et al. A new azaphilone derivative from the Monasucs kaoliang fermented rice. Chem. Nat. Compd. 2019, 55, 79–81. [Google Scholar] [CrossRef]
- Li, W.; Lee, C.; Bang, S.H.; Ma, J.Y.; Kim, S.; Koh, Y.S.; Shim, S.H. Isochromans and Related constituents from the endophytic fungus annulohypoxylon truncatum of zizania caduciflora and their anti-inflammatory effects. J. Nat. Prod. 2017, 80, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Orfali, R.S.; Aly, A.H.; Ebrahim, W.; Rudiyansyah, R.; Proksch, P. Isochroman and isocoumarin derivatives from hypersaline lake sediment derived fungus Penicillium sp. Phytochem. Lett. 2015, 13, 234–238. [Google Scholar] [CrossRef]
- Cheng, M.; Wu, M.; Chan, H.; Cheng, Y.C.; Chen, J.J.; Chen, I.S.; Su, Y.S.; Yuan, G.F. New metabolite isolated from the fungus Monascus pilosus. Chem. Nat. Compd. 2017, 53, 44–47. [Google Scholar] [CrossRef]
- Li, X.H.; Han, X.H.; Qin, L.L.; He, J.L.; Cao, Z.X.; Guo, D.L.; Deng, Y.; Gu, Y.C. Isochromanes from Aspergillus fumigatus, an endophytic fungus from cordyceps sinensis. Nat. Prod. Res. 2019, 33, 1870–1875. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Li, D.; Peng, J.; Zhu, T.; Gu, Q.; Li, D. Penicitols A-C and Penixanacid from the mangrove derived Penicillium chryogenum HDN11-24. J. Nat. Prod. 2015, 78, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Sperlich, J.; Hofert, S.P.; Janiak, C.; Teusch, N.; Stuhldreier, F.; Wesselborg, S.; Wang, C.; Kassack, M.U.; Dai, H.; et al. Azaphilone pigments and macrodiolides from the coprophilus fungus Coniella fragariae. Fitoterapia 2019, 137, 104249. [Google Scholar] [CrossRef]
- Wang, W.X.; Kusari, S.; Laatsch, H.; Golz, C.; Kusari, P.; Strohmann, C.; Kayser, O.; Spiteller, M. Antibacterial azaphilones from an endophytic fungus, Colletotrichum sp.BS4. J. Nat. Prod. 2016, 79, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.C.; Cheng, M.J.; Wu, M.D.; Chen, J.J.; Chen, Y.L.; Chang, H.S. Three new constituents from the fungus of Monascus purpureus and their anti-inflammatory activity. Phytochem. Lett. 2019, 31, 242–248. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, J.; Wang, X.; Zhao, L.; Liu, H.; Wei, Y.; You, X.; Cen, S.; Yu, L. Peniazaphilin A, a new azaphilone derivative produced by Penicillium sp. CPCC400786. J. Antibiot. 2018, 71, 905–907. [Google Scholar] [CrossRef]
- Xiong, X.; Zhang, X.; Wu, Z.; Wang, Z. Coupled aminophilic reaction and directed metabolic channeling to red Monascus pigments by extractive fermentation in nonionic surfactant micelle aqueous solution. Process. Biochem. 2015, 50, 180–187. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, X.; Wu, Z.; Qi, H.; Wang, Z. Perstraction of intracellular pigments by submerged cultivation of Monascus in nonionic surfactant micelle aqueous solution. Appl. Microbiol. Biotechnol. 2012, 94, 81–89. [Google Scholar] [CrossRef]
- Hinsch, E.; Robinson, S.C. Mechanical color reading of wood staining fungal pigment textile dyes: An alternative method for determining color fastness. Coating 2016, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Gupta, C.; Aggarwal, S.; Nagpal, N. Pigment extraction from fungus for textile dyeing. Indian J. Fibre Text. Res. 2012, 37, 68–73. [Google Scholar]
- Nambela, L.; Haule, L.V.; Mgani, Q. A review on source, chemistry, green synthesis and application of textile colorants. J. Clean. Prod. 2020, 246, 119036. [Google Scholar] [CrossRef]
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal pigments and their prospects in different industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef] [Green Version]
- Sajid, S.; Akber, N. Applications of fungal pigments in biotechnology. Pure Appl. Biol. 2018, 7, 922–930. [Google Scholar] [CrossRef]
- Kumar, A.; Vishwakarma, H.S.; Singh, J.; Dwivedi, S.; Kumar, M. Microbial pigments: Production and their applications in various industries. Int. J. Pharm. Chem. Biol. Sci. 2015, 5, 203–212. [Google Scholar]
- Caro, Y.; Venkatachalam, M.; Lebeau, J.; Fouillaud, M.; Dufossé, L. Pigments and colorants from filamentous fungi. In Fungal Metabolites; Merillon, J.M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 499–568. [Google Scholar]
- Räisänen, R. Handbook of Natural Colorants; John Wiley & Sons: Chichester, UK, 2009; Chapter 11; pp. 183–200. [Google Scholar]
- Perumal, K.; Stalin, V.; Chandrasekarenthiran, S.; Sumathi, E.; Saravanakumar, A. Extraction and characterization of pigment from Sclerotinia sp. and its use in dyeing cotton. Text. Res. J. 2009, 79, 1178–1187. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Afshari, M.; Shahidi, F.; Mortazavi, S.; Tabatabai, F.; Es’haghi, Z. Investigating the influence of pH, temperature and agitation speed on yellow pigment production by Penicillium aculeatum ATCC 10409. Nat. Prod. Res. 2015, 29, 1300–1306. [Google Scholar] [CrossRef]
- Nagia, F.A.; El-Mohamedy, R. Dyeing of wool with natural anthraquinone dyes from Fusarium oxysporum. Dyes Pigment. 2007, 75, 550–555. [Google Scholar] [CrossRef]
- Morales-Oyervides, L.; Oliveira, J.; Sousa-Gallagher, M.; Méndez-Zavala, A.; Montañez, J.C. Assessment of the dyeing properties of the pigments produced by Talaromyces spp. J. Fungi 2017, 3, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadni, Z.; Rahaman, M.H.; Jerin, I.; Hoque, K.M.F.; Reza, M.A. Extraction and optimization of red pigment production as secondary metabolites from Talaromyces verruculosus and its potential use in textile industries. Mycology 2017, 8, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Celestino, J.R.; Carvalho, L.; Lima, M.P.; Lima, A.M.; Ogusku, M.M.; de Souze, J.V.B. Bioprospecting of Amazon foil fungi with the potential pigment production. Process Biochem. 2014, 49, 569–575. [Google Scholar] [CrossRef]
- Hinsch, E.M.; Chen, H.L.; Weber, G.; Robinson, S.C. Colorfastness of extracted wood staining fungal pigments on fabrics: A new potential for textile dyes. J. Text. Appar. Technol. Manag. 2015, 3. [Google Scholar]
- Agurto, M.E.P.; Gutierrez, S.M.V.; Chen, H.L.; Robinson, S.C. Wood rotting fungal pigments as colorant coatings on oil based textile paints. Coating 2017, 7, 152. [Google Scholar] [CrossRef] [Green Version]
- Gupta, C.; Sharma, D.; Aggarwal, S.; Nagpal, N. Pigment production from Trichoderma sp. for dyeing of silk and wool. Int. J. Sci. Nat. 2013, 4, 351–355. [Google Scholar]
- Poorniammal, R.; Parthiban, M.; Gunasekaran, S.; Murugesan, R.; Thilagavathi, G. Natural dye production from Thermomyces sp fungi for textile application. Indian J. Fibre Text. Res. 2013, 276–279. [Google Scholar]
- Devi, S.; Karuppan, P. Reddish brown pigments from Alternaria alternate for textile dyeing and printing. Indian J. Fibre Text. Res. 2015, 40, 315–319. [Google Scholar]
- Velmurugan, P.; Kim, M.J.; Park, J.S.; Karthikeyan, K.; Lakshmanaperumalsamy, P.; Lee, K.J.; Park, Y.J.; Oh, B.T. Dyeing of cotton yarn with five water soluble fungal pigments obtained from five fungi. Fibre Polym. 2010, 11, 598–605. [Google Scholar] [CrossRef]
- Sardaryan, E.; Zihlova, H.; Strnad, R.; Cermakova, Z. Arpink Red-meet a new natural red food colorant of microbial origin. In Pigments in Food, More than Colours; Dufossé, L., Ed.; Elsevier: Quimper, France, 2004; pp. 207–208. [Google Scholar]
- Devi, A. Extraction of natural dyes from fungus—An alternate for textile dyeing. J. Nat. Sci. Res. 2014, 4, 1–6. [Google Scholar]
- Iswarya, S.; Shanuja, S.K.; Giri Dev, V.R.; Gnanamani, A. Asperyellone—A suitable coloring agent for protein based textile fabrics: An approach on production, characterization and application. J. Text. Eng. Fash. Technol. 2019, 5, 73–79. [Google Scholar]
- Hernandez, V.; Galleguillos, F.; Thibaut, R.; Muller, A. Fungal dyes for textile applications: Testing of industrial conditions for wool fabrics dyeing. J. Text. Inst. 2018, 110, 1–6. [Google Scholar] [CrossRef]
- Shibila, S.D.; Nanthini, A.U.R. Extraction and characterization of red pigment from Talaromyces australis and its application in dyeing cotton yarn. Int. Arch. Appl. Sci. Technol. 2019, 10, 81–91. [Google Scholar]
- Chiba, S.; Tsuyoshi, N.; Fudou, R.; Ojika, M.; Murakami, Y.; Ogoma, Y.; Oguchi, M.; Yamanaka, S. Magenta pigment produced by fungus. J. Gen. Appl. Microbiol. 2006, 52, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santis, D.; Moresi, M.; Gallo, A.M.; Petruccioli, M. Assessment of the dyeing properties of pigments from Monascus purpureus. J. Chem. Tech. Biotechnol. 2005, 80, 1072–1079. [Google Scholar] [CrossRef]
- Gokarneshan, N. Advances in textile printing. Int. J. Text. Sci. Eng. 2018, 01. [Google Scholar]
- Heo, Y.M.; Kim, K.; Kwon, S.L.; Na, J.; Lee, H.; Jang, S.; Kim, C.H.; Jung, J.; Kim, J.J. Investigation of filamentous fungi producing safe, functional water soluble pigmnets. Mycobiology 2018, 46, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Poorniammal, R.; Prabhu, S.; Sakthi, A.R.; Gunasekaran, S. Subacute dermal toxicity of Thermomyces sp. and Penicillium purpurogenum pigments in wistar rats. Int. J. Chem. Stud. 2019, 7, 630–634. [Google Scholar]
- Wang, W.; Liao, Y.; Chen, R.; Hou, Y.; Ke, W.; Zhang, B.; Gao, M.; Shao, Z.; Chen, J.; Li, F. Chlorinated azaphilone pigments with antimicrobial and cytotoxic activities isolated from the deep sea derived fungus Chaetomium sp. NA-S01-R1. Mar. Drugs 2018, 16, 61. [Google Scholar] [CrossRef] [Green Version]
- Pandiyarajan, S.; Premasudha, P.; Kadirvelu, K. Bio-production of novel water soluble yellow pigment from Aspergillus sp. and exploring its sustainable textile applications. 3 Biotech 2018, 8, 398. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Fan, F.; Gao, X.; Yang, C.; Bi, C.; Tang, J.; Liu, T.; Zhang, X. Balanced activation of IspG and IspH to eliminate MEP intermediate accumulation and improve isoprenoids production in Escherichia coli. Metab. Eng. 2017, 44, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Rugbjerg, P.; Naesby, M.; Mortensen, U.H.; Frandsen, R.J.N. Reconstruction of the biosynthetic pathway for the core fungal polyketide scaffold rubrofusarin in Saccharomyces cerevisiae. Microb. Cell Fact. 2013, 12, 31. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.Q.; Xu, Z.N.; Zhou, L.P.; Sung, C.K. Elimination of the mycotoxin citrinin production in the industrial important strain Monascus purpureus SM001. Metab. Eng. 2010, 12, 1–7. [Google Scholar] [CrossRef]
- Lebeau, J.; Petit, T.; Clerc, P.; Dufossé, L.; Caro, Y. Isolation of two novel purple naphthoquinone pigmnets concomitant with the bioactive red bikaverin and derivatives thereof produced by F.oxysporum. Biotechnol. Prog. 2019, 35, 2738. [Google Scholar] [CrossRef] [Green Version]
- Westphal, K.R.; Wollenberg, R.D.; Herbst, F.-A.; Sørensen, J.L.; Sondergaard, T.E.; Wimmer, R. Enhancing the production of the fungal pigment aurofusarin in Fusarium graminearum. Toxins 2018, 10, 485. [Google Scholar] [CrossRef] [Green Version]
- Klitgaard, A.; Frandsen, R.J.N.; Holm, D.K.; Knudsen, P.B.; Frisvad, J.C.; Nielsen, K.F. Combining UHPLC-high resolution MS and feeding of stable isotope labeled polyketide intermediates for linking precursors to end products. J. Nat. Prod. 2015, 78, 1518–1525. [Google Scholar] [CrossRef]
- Viggiano, A.; Salo, O.; Ali, H.; Szymanski, W.; Lankhorst, P.P.; Nygard, Y.; Bovenberg, R.A.L.; Driessen, A.J.M. Pathway for the biosynthesis of the pigment Chrysogine by Penicillium chrysogenum. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Pohl, C.; Kiel, J.A.; Driessen, A.J.; Bovenberg, R.A.; Ngard, Y. CRISPR/Cas9 based genome editing of Penicillium chrysogenum. ACS Synth. Biol. 2016, 5, 754–764. [Google Scholar] [CrossRef]
- Wollenberg, R.D.; Saei, W.; Westphal, K.R.; Klitgaard, C.S.; Nielsen, L.K.; Lysøe, E.; Gardiner, D.M.; Wimmer, R.; Sondergaard, T.E.; Sørensen, J.L.; et al. Chrysogine biosynthesis is mediated by a two-module nonribosomal peptide synthetase. J. Nat. Prod. 2017, 80, 2131–2135. [Google Scholar] [CrossRef]
- Rokas, A.; Wisecaver, J.H.; Lind, A.L. The birth, evolution and death of metabolic gene clusters in fungi. Nat. Rev. Microbiol. 2018, 16, 731–744. [Google Scholar] [CrossRef]
- Sen, T.; Barow, C.J.; Deshmukh, S.K. Microbial pigments in the food industry—Challenges and the way forward. Front. Nutr. 2019, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S.; Lee, J.H.; Lee, I. Strain improvement by overexpression of the laeA gene in monascus pilosus for the production of monascus-fermented rice. J. Microbiol. Biotechnol. 2013, 23, 959–965. [Google Scholar] [CrossRef] [Green Version]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Donohoue, P.D.; Barrangou, R.; May, A.P. Advances in industrial biotechnology using CRISPR-Cas systems. Trends Biotechnol. 2017, 36, 134–146. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.L.; Isbrandt, T.; Rasmussen, K.B.; Thrane, U.; Hoof, J.B.; Larsen, T.O.; Mortensen, U.H. Genes linked to production of secondary metabolites in Talaromyces atroroseus revealed using CRISPR-cas9. PLoS ONE 2017, 12, 2017. [Google Scholar] [CrossRef] [Green Version]
- Lopes, F.C.; Tichota, D.M.; Pereira, J.Q.; Segalin, J.; Rios Ade, O.; Brandelli, A. Pigment production by filamentous fungi on agro-industrial byproducts: An eco-friendly alternative. Appl. Biochem. Biotechnol. 2013, 171, 616–625. [Google Scholar] [CrossRef]
- Kaur, B.; Chakraborty, D.; Kaur, H. Production and stability analysis of yellowish pink pigments from Rhodotorula rubra MTCC 1446. Int. J. Microbiol. 2008, 7. [Google Scholar] [CrossRef] [Green Version]
- Subhasree, R.S.; Babu, D.; Mohan, V.P.C. Effect of carbon and nitrogen sources on stimulation of pigment production by monascus purpureus on jackfruit seeds. Int. J. Microbiol. Res. 2011.
- Srianta, I.; Novita, Y.; Kusumawati, N. Production of monascus pigments on durian seed. Effect of supplementation of carbon source. J. Pure Appl. Microbiol. 2012, 6, 59–63. [Google Scholar]
- Silveira, S.T.; Daroit, D.J.; Brandelli, A. Pigment production by Monascus purpureus in grape waste using factorial design. LWT Food Sci. Technol. 2008, 41, 170–174. [Google Scholar] [CrossRef]
- Rajeshwari, T.R.; Ponnusami, V.; Sugumaran, K.R. Production of monascus pigment in low cost fermentation. Int. J. Chem. Tech. Res. 2014, 6, 2929–2932. [Google Scholar]
- Taskin, M.; Sisman, T.; Erdal, S.; Kurbanoglu, E.B. Use of waste chicken feathers as peptone for production of carotenoids in submerged culture of Rhodotorula glutinis MT-5. Eur. Food Res. Technol. 2011, 233, 657–665. [Google Scholar] [CrossRef]
- Sanchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef]
- Mapari, S.A.S.; Meyer, A.S.; Frisvad, J.C.; Thrane, U. Production of Monascus Like Azaphilone Pigment. US Patent No. US20110250656A1, 2008. [Google Scholar]
- Pagano, M.C.; Dhar, P.P. Fungal pigments: An overview. In Fungal Bio-Molecules: Sources, Applications and Recent Developments, 1st ed.; Gupta, V.K., Mach, R.L., Sreenivasaprasad, S., Eds.; Wiley: London, UK, 2015; ISBN 978-1-118-95829-2. [Google Scholar]
- Loto, I.; Gutierrez, M.S.; Barahona, S.; Sepulveda, D.; Martinez-Moya, P.; Baeza, M.; Cifuentes, V.; Alcaino, J. Enhancement of carotenoid production by disrupting the C22-sterol desaturase gene (CYP61) in Xanthophyllomyces dendrorhous. BMC Microbiol. 2012, 12, 235. [Google Scholar] [CrossRef] [Green Version]
- Gmoser, R.; Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Filamentous ascomycetes fungi as a source of natural pigments. Fungal Biol. Biotechnol. 2017, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Dufossé, L. Microbial production of food grade pigments. Food Technol. Biotechnol. 2006, 44, 313–323. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Pigment | Color | Fabrics | References |
|---|---|---|---|---|
| Penicillium oxalicum | Anthraquinones | Arpink Red | Wool | Sardaryan et al. [97] |
| Trichoderma virens | Anthraquinones | Yellow | Silk, Wool | Sharma et al. [77] |
| Alternaria alternata | Anthraquinones | Reddish-Brown | ||
| Curvularia lunata | Anthraquinones | Black | ||
| Alternaria alternata | Anthraquinones | Reddish-Brown | Cotton | Devi and Karuppan [95] |
| Thermomyces sp. | Anthraquinones | Yellow | Cotton, Silk, Wool | Poorniammal et al. [94] |
| Trichoderma sp. | Anthraquinones | Yellow | Cotton, Silk, Silk cotton | Devi [98] |
| Trichoderma sp. | Anthraquinones | Yellow | Silk, Wool | Gupta et al. [93] |
| Penicillium oxalicum (NRC M25) | Anthraquinones | Faint Reddish- Brown | Wool | Mabrouk et al. [24] |
| Sclerotinia sp. | Anthraquinones | Pinkish-Red | Cotton | Perumal et al. [84] |
| Aspergillus sp. AN01 | Asperyellone | Yellow | Silk, Cotton, Synthetic and Wool fabrics | Iswarya et al. [99] |
| Monascus purpureus | Azaphilones | Red | Cotton | Velmurugan et al. [96] |
| Penicillium purpurogenum | Yellow | |||
| Isaria farinosa | Pink | |||
| Fusarium verticillioides | Reddish- Brown | |||
| Emericella nidulans | Red | |||
| Penicillium murcianum | Carotenoids | Yellow | Wool | Hernandez et al. [100] |
| Talaromyces australis | Red | |||
| Talaromyces australis | 2, 4-Di-tert-butylphenol | Red | Cotton fabric | Shibila and Nanthini [101] |
| Phoma herbarum | Magenta pigment | Magenta | Nylon | Chiba et al. [102] |
| Monascus purpureus | Monascorubramine | Red | Wool | De santis et al. [103] |
| Talaromyces verruculosus | Polyketide | Red | Cotton fabric | Chadni et al. [89] |
| Monascus purpureus | Rubropunctamine | Red | Wool | De santis et al. [103] |
| Chlorociboria aeruginosa | Quinones | Green | Bleached cotton, Spun polyamide, Spun polyester, Spun polyacrylic, Worsted wool | Weber et al. [48]; Hinsch et al. [91] |
| Scytalidium cuboideum | Red | |||
| Scytalidium ganodermophthorum | Yellow | |||
| Aspergillus sp. | Quinones | Brown | Cotton, Silk, Silk cotton | Devi [98] |
| Alternaria alternata | Quinones | Reddish-Brown | Cotton | Gokarneshan [104] |
| Acrostalagmus (NRC 90) | Quinones | Brown | Wool | Mabrouk et al. [24] |
| Alternaria alternata (NRC17) | Reddish-Brown | |||
| Alternaria sp. (NRC 97) | Brown | |||
| Aspergillus niger (NRC 95) | Brown | |||
| Bisporomyces sp. (NRC 63) | Deep Brown | |||
| Cunninghamella (NRC 188) | Faint Reddish-Brown | |||
| Penicillium chrysogenum (NRC 74) | Deep Brown | |||
| Penicillium italicum (NRC E11) | Brown | |||
| Penicillium regulosum (NRC 50) | Brown | |||
| Phymatotrichum sp. (NRC 151) | Reddish-Brown |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venil, C.K.; Velmurugan, P.; Dufossé, L.; Renuka Devi, P.; Veera Ravi, A. Fungal Pigments: Potential Coloring Compounds for Wide Ranging Applications in Textile Dyeing. J. Fungi 2020, 6, 68. https://doi.org/10.3390/jof6020068
Venil CK, Velmurugan P, Dufossé L, Renuka Devi P, Veera Ravi A. Fungal Pigments: Potential Coloring Compounds for Wide Ranging Applications in Textile Dyeing. Journal of Fungi. 2020; 6(2):68. https://doi.org/10.3390/jof6020068
Chicago/Turabian StyleVenil, Chidambaram Kulandaisamy, Palanivel Velmurugan, Laurent Dufossé, Ponnuswamy Renuka Devi, and Arumugam Veera Ravi. 2020. "Fungal Pigments: Potential Coloring Compounds for Wide Ranging Applications in Textile Dyeing" Journal of Fungi 6, no. 2: 68. https://doi.org/10.3390/jof6020068