
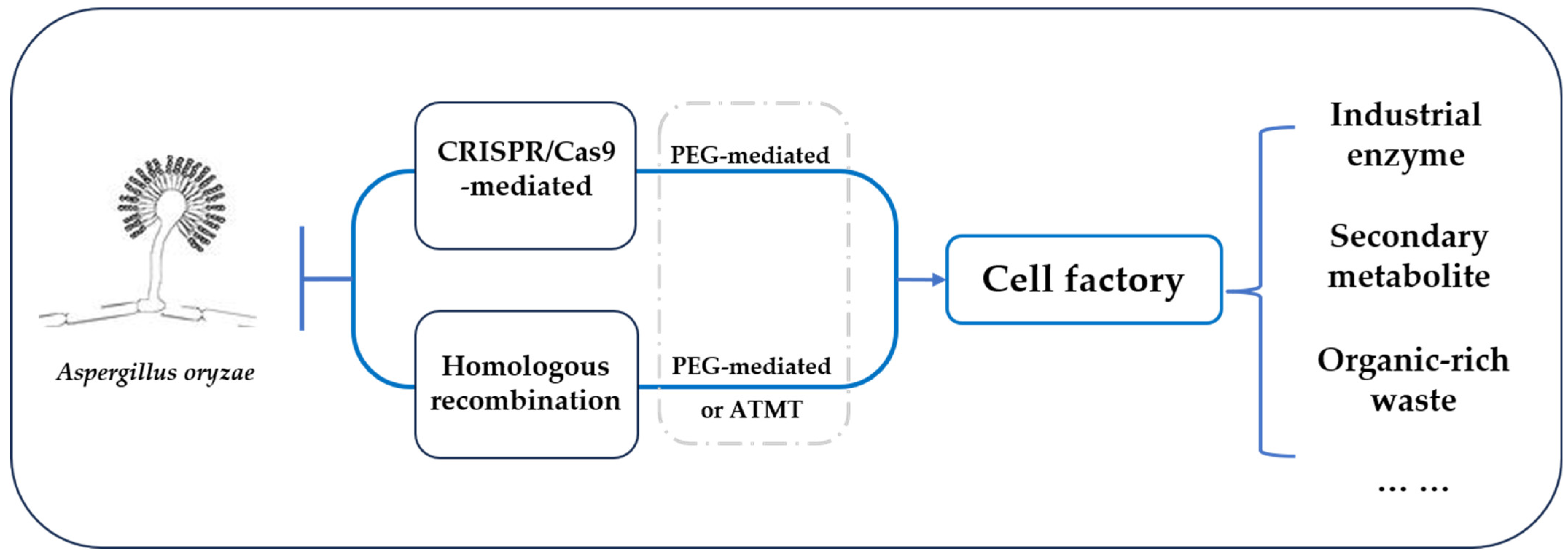
Aspergillus oryzae as a Cell Factory: Research and Applications in Industrial Production
 ,
,
Abstract
:1. Introduction
2. Cell Factory of Producing Secondary Metabolite by A. oryzae
2.1. Metabolic Engineering of KA Production in A. oryzae
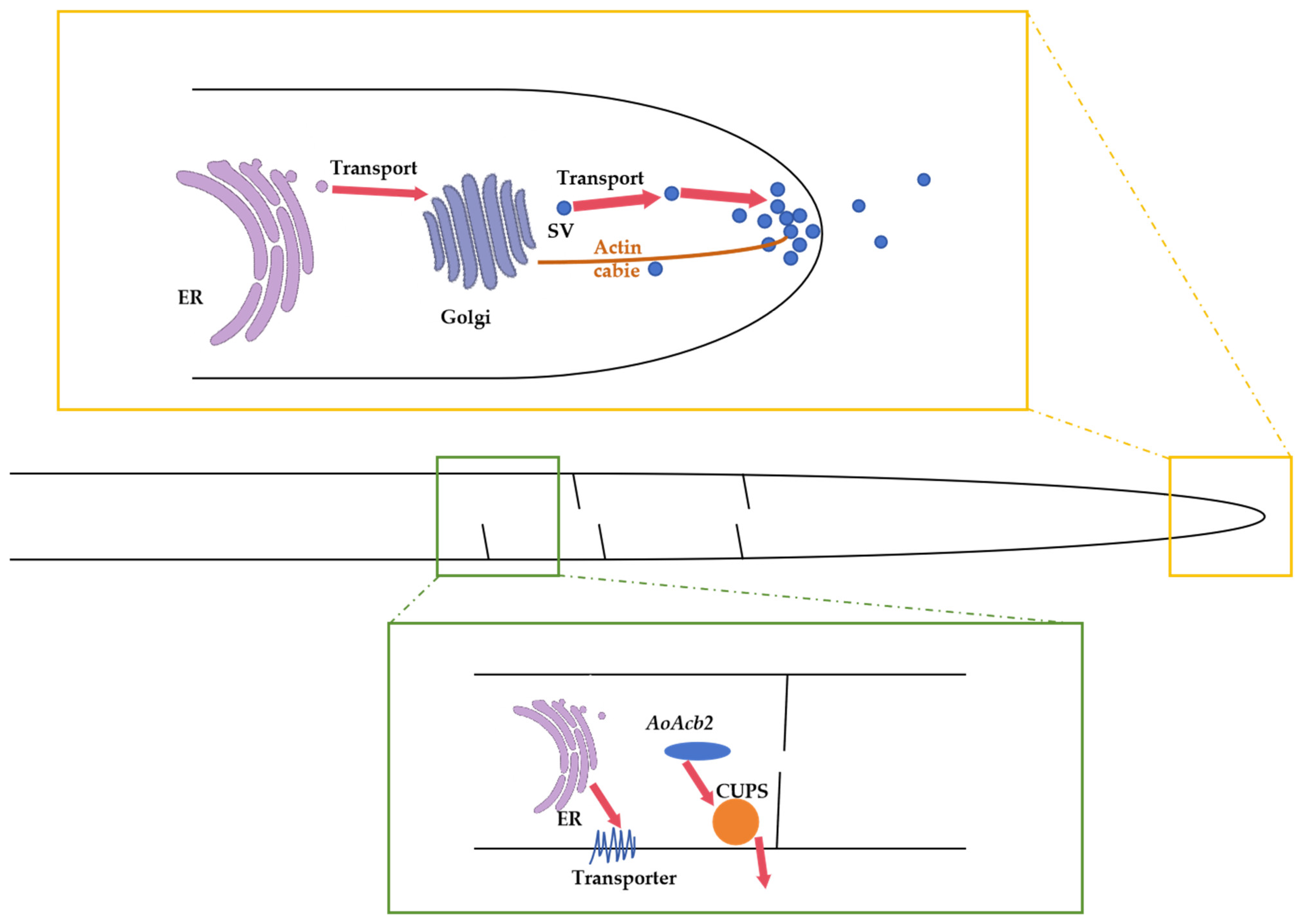
2.2. Molecular Mechanism for Secondary Metabolite Secretion in A. oryzae
3. Cell Factory for Producing Industrial Enzymes with A. oryzae
3.1. α-Amylase Production in A. oryzae
3.2. Molecular Mechanism for Protein Secretion in A. oryzae
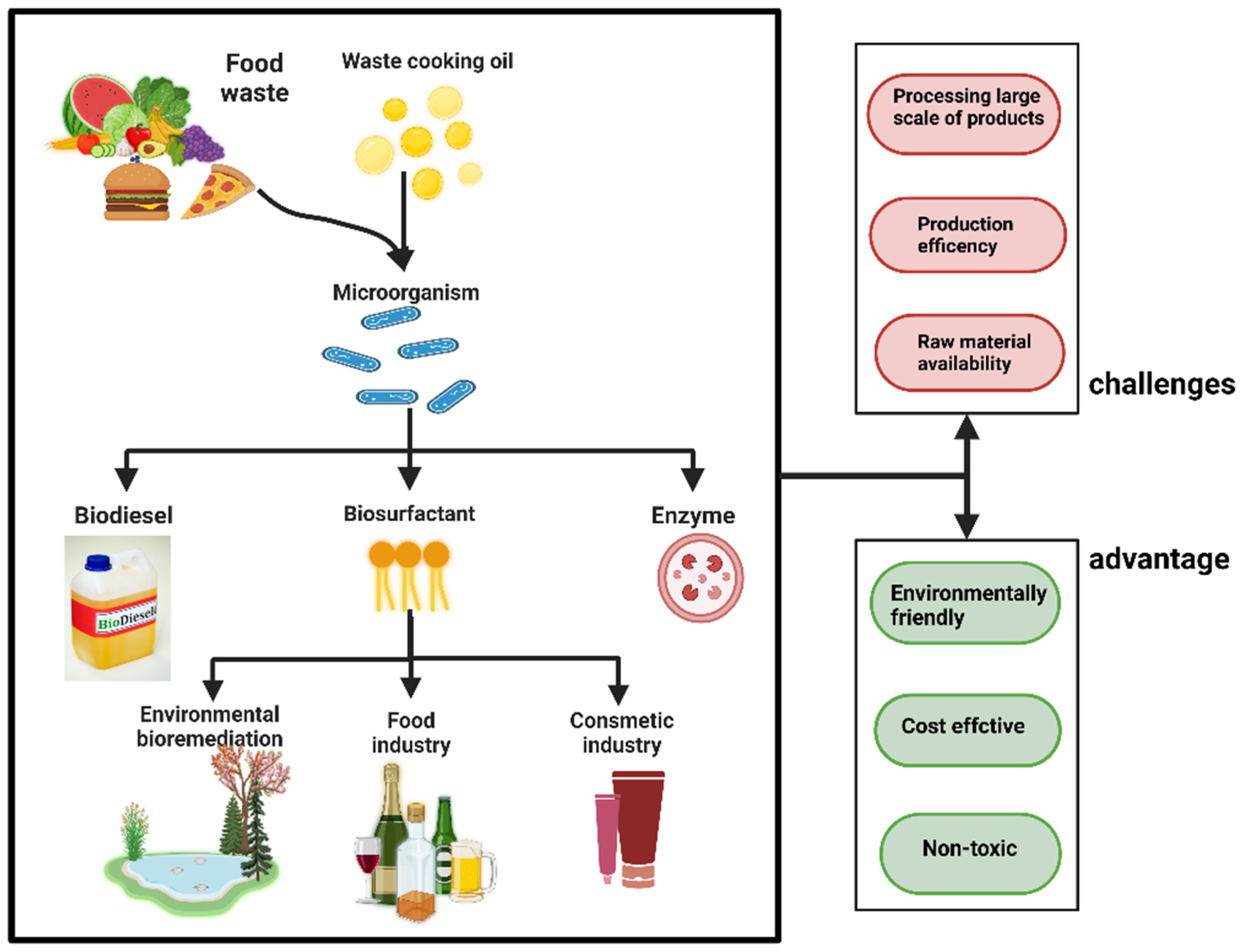
4. Cell Factory of Utilized Organic-Rich Waste by A. oryzae
4.1. Cell Factories for Processing Food Waste
4.2. Cell Factory for Processing Agricultural Waste
5. Discussion and Conclusions
6. Expectations
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jo, C.; Zhang, J.; Tam, J.M.; Church, G.M.; Khalil, A.S.; Segré, D.; Tang, T.C. Unlocking the magic in mycelium: Using synthetic biology to optimize filamentous fungi for biomanufacturing and sustainability. Mater. Today Bio 2023, 19, 100560. [Google Scholar] [CrossRef]
- Ding, Q.; Ye, C. Microbial cell factories based on filamentous bacteria, yeasts, and fungi. Microb. Cell Factories 2023, 22, 20. [Google Scholar] [CrossRef]
- Lambré, C.; Baviera, J.M.B.; Bolognesi, C.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; Mengelers, M.; Mortensen, A.; et al. Safety evaluation of the food enzyme phospholipase A1 from the genetically modified Aspergillus oryzae strain NZYM-LJ. EFSA J. 2022, 20, 7381. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Møller, L.L.H.; Larsen, T.O.; Kumar, R.; Arnau, J. Safety of the fungal workhorses of industrial biotechnology: Update on the mycotoxin and secondary metabolite potential of Aspergillus niger, Aspergillus oryzae, and Trichoderma reesei. Appl. Microbiol. Biotechnol. 2018, 102, 9481–9515. [Google Scholar] [CrossRef]
- Saqib, A.; Rashid, M.H. Random mutagenesis of koji (Aspergillus oryzae) to enhance the catalytic efficiency and thermostability of glucoamylase. Pak. J. Bot. 2023, 55, 1923–1929. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.F.; Hou, S.; Wu, C.Z.; Cao, Y.; Tong, X.; Chen, Y.J. Improving protein utilization and fermentation quality of soy sauce by adding protease. J. Food Compos. Anal. 2023, 121, 105399. [Google Scholar] [CrossRef]
- Qiu, S.K.; Liu, Q.C.; Yuan, Y.; Zhou, H.; Zeng, B. Aspergillus oryzae accelerates the conversion of ergosterol to ergosterol peroxide by efficiently utilizing cholesterol. Front. Genet. 2022, 13, 984343. [Google Scholar] [CrossRef] [PubMed]
- Nomura, R.; Tsuzuki, S.; Kojima, T.; Nagasawa, M.; Sato, Y.; Uefune, M.; Baba, Y.; Hayashi, T.; Nakano, H.; Kato, M.; et al. Administration of Aspergillus oryzae suppresses DSS-induced colitis. Food Chem. Mol. Sci. 2022, 4, 100063. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, J.; Zhang, G.; Liu, S.; Zhou, J.; Du, G.; Chen, J. Recent advances in the development of Aspergillus for protein production. Bioresour. Technol. 2022, 348, 126768. [Google Scholar] [CrossRef] [PubMed]
- Galagan, J.E.; Calvo, S.E.; Cuomo, C.; Ma, L.J.; Wortman, J.R.; Batzoglou, S.; Lee, S.I.; Bastürkmen, M.; Spevak, C.C.; Clutterbuck, J.; et al. Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 2005, 438, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.Y.; Ji, L.H.; Pan, J.Y.; Chen, X.L.; Gao, C.; Liu, J.; Wei, W.Q.; Wu, J.; Song, W.; Liu, L.M. Engineering growth phenotypes of Aspergillus oryzae for L-malate production. Bioresour. Bioprocess. 2023, 10, 25. [Google Scholar] [CrossRef]
- Zhang, X.M.; Guo, R.; Bi, F.T.; Chen, Y.; Xue, X.L.; Wang, D.P. Overexpression of kojR and the entire koj gene cluster affect the kojic acid synthesis in Aspergillus oryzae 3.042. Gene 2024, 892, 147852. [Google Scholar] [CrossRef]
- Yang, R.; Wang, Y.; Zhao, X.; Tong, Z.; Zhu, Q.; He, X.; Wang, Z.; Luo, H.; Fang, F. A facile and efficient synthesis approach of salidroside esters by whole-cell biocatalysts in organic solvents. Front. Bioeng. Biotechnol. 2022, 10, 1051117. [Google Scholar] [CrossRef]
- Umemura, M.; Koyama, Y.; Takeda, I.; Hagiwara, H.; Ikegami, T.; Koike, H.; Machida, M. Fine De Novo Sequencing of a Fungal Genome Using only SOLiD Short Read Data: Verification on Aspergillus oryzae RIB40. PLoS ONE 2013, 8, e63673. [Google Scholar] [CrossRef]
- Inglis, D.O.; Binkley, J.; Skrzypek, M.S.; Arnaud, M.B.; Cerqueira, G.C.; Shah, P.; Wymore, F.; Wortman, J.R.; Sherlock, G. Comprehensive annotation of secondary metabolite biosynthetic genes and gene clusters of Aspergillus nidulans, A. fumigatus, A. niger and A. oryzae. BMC Microbiol. 2013, 13, 91. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Oshima, K.; Noguchi, H.; Ogawa, M.; Takahashi, T.; Oguma, T.; Koyama, Y.; Itoh, T.; Hattori, M.; Hanya, Y. Draft Genome Sequencing and Comparative Analysis of Aspergillus sojae NBRC4239. DNA Res. 2011, 18, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Suryadi, H.; Irianti, M.I.; Septiarini, T.H. Methods of Random Mutagenesis of Aspergillus Strain for Increasing Kojic Acid Production. Curr. Pharm. Biotechnol. 2022, 23, 486–494. [Google Scholar] [CrossRef]
- Nodvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 System for Genetic Engineering of Filamentous Fungi. PLoS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, J.; Zhang, G.; Zhou, J.; Li, J.; Du, G.; Chen, J. CRISPR/Cas9-Mediated Multiplexed Genome Editing in Aspergillus oryzae. J. Fungi 2023, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Maruyama, J.-i. CRISPR/Cpf1-mediated mutagenesis and gene deletion in industrial filamentous fungi Aspergillus oryzae and Aspergillus sojae. J. Biosci. Bioeng. 2022, 133, 353–361. [Google Scholar] [CrossRef]
- Kumar, V.; Ahluwalia, V.; Saran, S.; Kumar, J.; Patel, A.K.; Singhania, R.R. Recent developments on solid-state fermentation for production of microbial secondary metabolites: Challenges and solutions. Bioresour. Technol. 2021, 323, 124566. [Google Scholar] [CrossRef]
- Mushtaq, S.; Abbasi, B.H.; Uzair, B.; Abbasi, R. Natural products as reservoirs of novel therapeutic agents. EXCLI J. 2018, 17, 420–451. [Google Scholar]
- Seshime, Y.; Juvvadi, P.R.; Tokuoka, M.; Koyama, Y.; Kitamoto, K.; Ebizuka, Y.; Fujii, I. Functional expression of the Aspergillus flavus PKS-NRPS hybrid CpaA involved in the biosynthesis of cyclopiazonic acid. Bioorganic Med. Chem. Lett. 2009, 19, 3288–3292. [Google Scholar] [CrossRef] [PubMed]
- Panchanawaporn, S.; Chutrakul, C.; Jeennor, S.; Anantayanon, J.; Rattanaphan, N.; Laoteng, K. Potential of Aspergillus oryzae as a biosynthetic platform for indigoidine, a non-ribosomal peptide pigment with antioxidant activity. PLoS ONE 2022, 17. [Google Scholar] [CrossRef]
- Petitgonnet, C.; Klein, G.L.; Roullier-Gall, C.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Julien-David, D.; Alexandre, H. Influence of cell-cell contact between L. thermotolerans and S. cerevisiae on yeast interactions and the exo-metabolome. Food Microbiol. 2019, 83, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Awakawa, T.; Abe, I. Reconstitution of Polyketide-Derived Meroterpenoid Biosynthetic Pathway in Aspergillus oryzae. J. Fungi 2021, 7, 486. [Google Scholar] [CrossRef] [PubMed]
- Kan, E.; Tomita, H.; Katsuyama, Y.; Maruyama, J.-I.; Koyama, Y.; Ohnishi, Y. Discovery of the 2,4′-Dihydroxy-3′-methoxypropiophenone Biosynthesis Genes in Aspergillus oryzae. ChemBioChem 2021, 22, 203–211. [Google Scholar] [CrossRef]
- Orfali, R.; Perveen, S.; Khan, M.F.; Ahmed, A.F.; Wadaan, M.A.; Al-Taweel, A.M.; Alqahtani, A.S.; Nasr, F.A.; Tabassum, S.; Luciano, P.; et al. Antiproliferative Illudalane Sesquiterpenes from the Marine Sediment Ascomycete Aspergillus oryzae. Mar. Drugs 2021, 19, 333. [Google Scholar] [CrossRef]
- Zhu, K.; Liu, X.; Qi, X.; Liu, Q.; Wang, B.; Sun, W.; Pan, B. Acaricidal activity of bioactive compounds isolated from Aspergillus oryzae against poultry red mites, Dermanyssus gallinae (Acari: Dermanyssidae). Vet. Parasitol. 2023, 320, 109983. [Google Scholar] [CrossRef]
- Han, H.; Yu, C.; Qi, J.; Wang, P.; Zhao, P.; Gong, W.; Xie, C.; Xia, X.; Liu, C. High-efficient production of mushroom polyketide compounds in a platform host Aspergillus oryzae. Microb. Cell Factories 2023, 22, 60. [Google Scholar] [CrossRef]
- Jiao, J.; Fu, J.-X.; Gai, Q.-Y.; He, X.-J.; Feng, X.; Cao, R.-Z.; Fu, Y.-J. The enhanced production and secretion of high-value cajaninstilbene acid and flavonoid aglycones in Cajanus cajan (Linn.) Millsp. cell suspension cultures elicited by Aspergillus oryzae Y-29. Process Biochem. 2023, 130, 127–137. [Google Scholar] [CrossRef]
- Kövilein, A.; Zadravec, L.; Hohmann, S.; Umpfenbach, J.; Ochsenreither, K. Effect of process mode, nitrogen source and temperature on L-malic acid production with Aspergillus oryzae DSM 1863 using acetate as carbon source. Front. Bioeng. Biotechnol. 2022, 10, 1033777. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, V.; Derenbach, L.; Ochsenreither, K. Enhanced l-Malic Acid Production by Aspergillus oryzae DSM 1863 Using Repeated-Batch Cultivation. Front. Bioeng. Biotechnol. 2022, 9, 760500. [Google Scholar] [CrossRef] [PubMed]
- Kövilein, A.; Aschmann, V.; Hohmann, S.; Ochsenreither, K. Immobilization of Aspergillus oryzae DSM 1863 for l-Malic Acid Production. Fermentation 2022, 8, 26. [Google Scholar] [CrossRef]
- Ji, L.; Wang, J.; Luo, Q.; Ding, Q.; Tang, W.; Chen, X.; Liu, L. Enhancing L-malate production of Aspergillus oryzae by nitrogen regulation strategy. Appl. Microbiol. Biotechnol. 2021, 105, 3101–3113. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, G.A.E.; Zohri, A.N.A. Amedment stable kojic acid produced by non-toxinogenic Aspergillus oryzae using five levels central composite design of response surface methodology. J. Microbiol. Biotechnol. Food Sci. 2021, 10, e2683. [Google Scholar] [CrossRef]
- Li, J.S.; Chew, Y.M.; Lin, M.C.; Lau, Y.Q.; Chen, C.S. Enhanced glucosamine production from Aspergillus oryzae NCH-42 via acidic stress under submerged fermentation. Cyta-J. Food 2021, 19, 614–624. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, S.; Sarma, S.J. Challenges and advancements in bioprocess intensification of fungal secondary metabolite: Kojic acid. World J. Microbiol. Biotechnol. 2023, 39, 140. [Google Scholar] [CrossRef]
- Lassfolk, R.; Suonpää, A.; Birikh, K.; Leino, R. Chemo-enzymatic three-step conversion of glucose to kojic acid. Chem. Commun. 2019, 55, 14737–14740. [Google Scholar] [CrossRef]
- Troiano, D.; Orsat, V.; Dumont, M.J. Status of filamentous fungi in integrated biorefineries. Renew. Sustain. Energy Rev. 2020, 117, 109472. [Google Scholar] [CrossRef]
- Chang, P.-K.; Scharfenstein, L.L.; Mahoney, N.; Kong, Q. Kojic Acid Gene Clusters and the Transcriptional Activation Mechanism of Aspergillus flavus KojR on Expression of Clustered Genes. J. Fungi 2023, 9, 259. [Google Scholar] [CrossRef]
- Marui, J.; Yamane, N.; Ohashi-Kunihiro, S.; Ando, T.; Terabayashi, Y.; Sano, M.; Ohashi, S.; Ohshima, E.; Tachibana, K.; Higa, Y.; et al. Kojic acid biosynthesis in Aspergillus oryzae is regulated by a Zn(II)2Cys6 transcriptional activator and induced by kojic acid at the transcriptional level. J. Biosci. Bioeng. 2011, 112, 40–43. [Google Scholar] [CrossRef]
- Arakawa, G.-y.; Kudo, H.; Yanase, A.; Eguchi, Y.; Kodama, H.; Ogawa, M.; Koyama, Y.; Shindo, H.; Hosaka, M.; Tokuoka, M. A unique Zn(II)2-Cys6-type protein, KpeA, is involved in secondary metabolism and conidiation in Aspergillus oryzae. Fungal Genet. Biol. 2019, 127, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Chen, Z.; Li, Y.; Zeng, B.; Zhang, Z. A Novel Major Facilitator Superfamily Transporter Gene Aokap4 near the Kojic Acid Gene Cluster Is Involved in Growth and Kojic Acid Production in Aspergillus oryzae. J. Fungi 2022, 8, 885. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, T.; Wang, H.; Jiang, C.; Liu, Y.; Wu, X.; Li, Y.; Zeng, B.; Zhang, Z. Disruption of Aokap6 near the kojic acid gene cluster affects the growth and kojic acid production in Aspergillus oryzae. World J. Microbiol. Biotechnol. 2022, 38, 175. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Arioka, M. Autophagy deficiency boosts the production of kojic acid in the filamentous fungus Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2021, 85, 2429–2433. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, J.; Long, C.; He, B.; Hu, Z.; Jiang, C.; Li, Y.; Ma, L.; Wen, J.; Zou, X.; et al. Identification and characterization of the ZRT, IRT-like protein (ZIP) family genes reveal their involvement in growth and kojic acid production in Aspergillus oryzae. J. Ind. Microbiol. Biotechnol. 2019, 46, 1769–1780. [Google Scholar] [CrossRef]
- Kudo, H.; Arakawa, G.-y.; Shirai, S.; Ogawa, M.; Shindo, H.; Hosaka, M.; Tokuoka, M. New role of a histone chaperone, HirA: Involvement in kojic acid production associated with culture conditions in Aspergillus oryzae. J. Biosci. Bioeng. 2022, 133, 235–242. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Chen, Z.; Fan, J.; Chen, T.; Xiao, Y.; Jie, J.; Zeng, B.; Zhang, Z. Overexpression of a novel gene Aokap2 affects the growth and kojic acid production in Aspergillus oryzae. Mol. Biol. Rep. 2022, 49, 2745–2754. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Z.; Zhang, F.; Chen, T.; Fan, J.; Deng, X.; Lei, X.; Zeng, B.; Zhang, Z. The C2H2-type zinc-finger regulator AoKap5 is required for the growth and kojic acid synthesis in Aspergillus oryzae. Fungal Genet. Biol. 2023, 167, 103813. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Chen, Z.; Fan, J.; Chen, T.; Zeng, B.; Zhang, Z. Identification and characterization of a novel gene Aokap1 involved in growth and kojic acid synthesis in Aspergillus oryzae. Arch. Microbiol. 2021, 204, 67. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, T.; Zhang, H.; Li, Y.; Fan, J.; Yao, L.; Zeng, B.; Zhang, Z. Functional role of a novel zinc finger protein, AoZFA, in growth and kojic acid synthesis in Aspergillus oryzae. Appl. Environ. Microbiol. 2023, 89, e00909–e00923. [Google Scholar] [CrossRef] [PubMed]
- Futagami, T.; Mori, K.; Wada, S.; Ida, H.; Kajiwara, Y.; Takashita, H.; Tashiro, K.; Yamada, O.; Omori, T.; Kuhara, S.; et al. Transcriptomic Analysis of Temperature Responses of Aspergillus kawachii during Barley Koji Production. Appl. Environ. Microbiol. 2015, 81, 1353–1363. [Google Scholar] [CrossRef]
- Kadooka, C.; Izumitsu, K.; Onoue, M.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Goto, M.; Tamaki, H.; Futagami, T. Mitochondrial Citrate Transporters CtpA and YhmA Are Required for Extracellular Citric Acid Accumulation and Contribute to Cytosolic Acetyl Coenzyme A Generation in Aspergillus luchuensis mut. kawachii. Appl. Environ. Microbiol. 2019, 85, e03136-18. [Google Scholar] [CrossRef] [PubMed]
- Kadooka, C.; Nakamura, E.; Mori, K.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Goto, M.; Tamaki, H.; Futagami, T. LaeA Controls Citric Acid Production through Regulation of the Citrate Exporter-Encoding cexA Gene in Aspergillus luchuensis mut. kawachii. Appl. Environ. Microbiol. 2020, 86, e01950-19. [Google Scholar] [CrossRef]
- Nakamura, E.; Kadooka, C.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Goto, M.; Tamaki, H.; Futagami, T. Citrate exporter enhances both extracellular and intracellular citric acid accumulation in the koji fungi Aspergillus luchuensis mut. kawachii and Aspergillus oryzae. J. Biosci. Bioeng. 2021, 131, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, B.; Khatri, M.; Singh, G.; Arya, S.K. Microbial keratinases: An overview of biochemical characterization and its eco-friendly approach for industrial applications. J. Clean. Prod. 2020, 252, 119847. [Google Scholar] [CrossRef]
- Sujitha, P.; Shanthi, C. Importance of enzyme specificity and stability for the application of proteases in greener industrial processing- a review. J. Clean. Prod. 2023, 425, 138915. [Google Scholar] [CrossRef]
- Jin, F.-J.; Hu, S.; Wang, B.-T.; Jin, L. Advances in Genetic Engineering Technology and Its Application in the Industrial Fungus Aspergillus oryzae. Front. Microbiol. 2021, 12, 644404. [Google Scholar] [CrossRef]
- Oikawa, H. Reconstitution of biosynthetic machinery of fungal natural products in heterologous hosts. Biosci. Biotechnol. Biochem. 2020, 84, 433–444. [Google Scholar] [CrossRef]
- Atalla, S.M.M.; Ahmed, N.E.; Awad, H.M.; El Gamal, N.G.; El Shamy, A.R. Statistical optimization of xylanase production, using different agricultural wastes by Aspergillus oryzae MN894021, as a biological control of faba bean root diseases. Egypt. J. Biol. Pest Control 2020, 30, 125. [Google Scholar] [CrossRef]
- de Carvalho, M.S.; de Menezes, L.H.S.; Pimentel, A.B.; Costa, F.S.; Oliveira, P.C.; dos Santos, M.M.O.; de Carvalho Tavares, I.M.; Irfan, M.; Bilal, M.; Dias, J.C.T.; et al. Application of Chemometric Methods for the Optimization Secretion of Xylanase by Aspergillus oryzae in Solid State Fermentation and Its Application in the Saccharification of Agro-industrial Waste. Waste Biomass Valori 2023, 14, 3183–3193. [Google Scholar] [CrossRef]
- Pragya; Sharma, K.K.; Singh, B. Phytase from Aspergillus oryzae SBS50: Biocatalytic reduction of anti-nutritional factor and exhibiting vanadium-dependent haloperoxidase activity. Biocatal. Agric. Biotechnol. 2023, 52, 102840. [Google Scholar] [CrossRef]
- Michel, M.R.; Gallegos, A.C.F.; Villarreal-Morales, S.L.; Aguilar-Zárate, P.; Aguilar, C.N.; Riutort, M.; Rodríguez-Herrera, R. Fructosyltransferase production by Aspergillus oryzae BM-DIA using solid-state fermentation and the properties of its nucleotide and protein sequences. Folia Microbiol. 2021, 66, 469–481. [Google Scholar] [CrossRef]
- Balakrishnan, M.; Jeevarathinam, G.; Kumar, S.K.S.; Muniraj, I.; Uthandi, S. Optimization and scale-up of α-amylase production by Aspergillus oryzae using solid-state fermentation of edible oil cakes. BMC Biotechnol. 2021, 21, 33. [Google Scholar] [CrossRef]
- Braia, M.; Cabezudo, I.; Barrera, V.L.; Anselmi, P.; Meini, M.-R.; Romanini, D. An optimization approach to the bioconversion of flour mill waste to α-amylase enzyme by Aspergillus oryzae. Process Biochem. 2021, 111, 102–108. [Google Scholar] [CrossRef]
- Han, S.; Pan, L.; Zeng, W.; Yang, L.; Yang, D.; Chen, G.; Liang, Z. Improved production of fructooligosaccharides (FOS) using a mutant strain of Aspergillus oryzae S719 overexpressing β-fructofuranosidase (FTase) genes. LWT 2021, 146, 111346. [Google Scholar] [CrossRef]
- Han, S.; Ye, T.; Leng, S.; Pan, L.; Zeng, W.; Chen, G.; Liang, Z. Purification and biochemical characteristics of a novel fructosyltransferase with a high FOS transfructosylation activity from Aspergillus oryzae S719. Protein Expr. Purif. 2020, 167, 105549. [Google Scholar] [CrossRef] [PubMed]
- Cabezudo, I.; Galetto, C.S.; Romanini, D.; Furlán, R.L.E.; Meini, M.R. Production of gallic acid and relevant enzymes by Aspergillus niger and Aspergillus oryzae in solid-state fermentation of soybean hull and grape pomace. Biomass Convers Bior 2023, 13, 14939–14947. [Google Scholar] [CrossRef]
- Fatima, B.; Javed, M.M. Kinetics, thermodynamics, and physicochemical properties of 1, 4-α-d-glucan glucohydrolase from Aspergillus oryzae NRRL1560. Biomass Convers. Biorefinery 2021, 11, 3011–3022. [Google Scholar] [CrossRef]
- Mamo, J.; Kangwa, M.; Fernandez-Lahore, H.M.; Assefa, F. Optimization of media composition and growth conditions for production of milk-clotting protease (MCP) from Aspergillus oryzae DRDFS13 under solid-state fermentation. Braz. J. Microbiol. 2020, 51, 571–584. [Google Scholar] [CrossRef]
- Mahboob, S.; Ali, S. Pectin lyase productivity by a uv-irradiated Aspergillus oryzae mutant under carrot-koji process. J. Anim. Plant Sci.-JAPS 2022, 32, 1375–1384. [Google Scholar] [CrossRef]
- Qin, Y.; Qin, B.; Zhang, J.; Fu, Y.; Li, Q.; Luo, F.; Luo, Y.; He, H. Purification and enzymatic properties of a new thermostable endoglucanase from Aspergillus oryzae HML366. Int. Microbiol. 2023, 26, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, K.; Shiono, Y.; Shintani, T.; Watanabe, A.; Kanzaki, H.; Gomi, K.; Koseki, T. Efficient production of recombinant tannase in Aspergillus oryzae using an improved glucoamylase gene promoter. J. Biosci. Bioeng. 2020, 129, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Kotani, A.; Ozaki, T.; Takino, J.; Mochizuki, S.; Akimitsu, K.; Minami, A.; Oikawa, H. Heterologous expression of a polyketide synthase ACRTS2 in Aspergillus oryzae produces host-selective ACR toxins: Coproduction of minor metabolites. Biosci. Biotechnol. Biochem. 2021, 86, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Elyasi Far, B.; Ahmadi, Y.; Yari Khosroshahi, A.; Dilmaghani, A. Microbial Alpha-Amylase Production: Progress, Challenges and Perspectives. Adv. Pharm. Bull. 2020, 10, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Marengo, M.; Pezzilli, D.; Gianquinto, E.; Fissore, A.; Oliaro-Bosso, S.; Sgorbini, B.; Spyrakis, F.; Adinolfi, S. Evaluation of Porcine and Aspergillus oryzae α-Amylases as Possible Model for the Human Enzyme. Processes 2022, 10, 780. [Google Scholar] [CrossRef]
- Gomi, K. Regulatory mechanisms for amylolytic gene expression in the koji mold Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2019, 83, 1385–1401. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Tanaka, M.; Konno, Y.; Ichikawa, T.; Ichinose, S.; Hasegawa-Shiro, S.; Shintani, T.; Gomi, K. Distinct mechanism of activation of two transcription factors, AmyR and MalR, involved in amylolytic enzyme production in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2015, 99, 1805–1815. [Google Scholar] [CrossRef]
- Bakker, R.; Mani, M.; Carthew, R.W. The Wg and Dpp morphogens regulate gene expression by modulating the frequency of transcriptional bursts. eLife 2020, 9, e56076. [Google Scholar] [CrossRef]
- Takeshita, N. Coordinated process of polarized growth in filamentous fungi. Biosci. Biotechnol. Biochem. 2016, 80, 1693–1699. [Google Scholar] [CrossRef]
- Krijgsheld, P.; Bleichrodt, R.; van Veluw, G.J.; Wang, F.; Müller, W.H.; Dijksterhuis, J.; Wösten, H.A.B. Development in Aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef]
- Yasui, M.; Oda, K.; Masuo, S.; Hosoda, S.; Katayama, T.; Maruyama, J.-i.; Takaya, N.; Takeshita, N. Invasive growth of Aspergillus oryzae in rice koji and increase of nuclear number. Fungal Biol. Biotechnol. 2020, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Tanaka, M.; Watanabe, T.; Zhan, S.; Watanabe, A.; Shintani, T.; Gomi, K. Crucial role of the intracellular α-glucosidase MalT in the activation of the transcription factor AmyR essential for amylolytic gene expression in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2021, 85, 2076–2083. [Google Scholar] [CrossRef] [PubMed]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.-I.; Arima, T.; Akita, O.; Kashiwagi, Y.; et al. Genome sequencing and analysis of Aspergillus oryzae. Nature 2005, 438, 1157–1161. [Google Scholar] [CrossRef]
- Higuchi, Y. Membrane Traffic in Aspergillus oryzae and Related Filamentous Fungi. J. Fungi 2021, 7, 534. [Google Scholar] [CrossRef]
- Chen, J.; Tonouchi, A. Copper ion (Cu2+) is involved in the transcription of the tyrosinase-encoding melB gene of Aspergillus oryzae in solid-state culture. Biosci. Biotechnol. Biochem. 2023, 88, 220–224. [Google Scholar] [CrossRef]
- Akao, T.; Yahara, A.; Sakamoto, K.; Yamada, O.; Akita, O.; Yoshida, T. Lack of endoplasmic reticulum 1,2-α-mannosidase activity that trims N-glycan Man9GlcNAc2 to Man8GlcNAc2 isomer B in a manE gene disruptant of Aspergillus oryzae. J. Biosci. Bioeng. 2012, 113, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Koseki, T.; Ishida, N.; Hirota, R.; Shiono, Y.; Makabe, K. Mutational analysis of the effects of N-glycosylation sites on the activity and thermal stability of rutinosidase from Aspergillus oryzae. Enzym. Microb. Technol. 2022, 161, 110112. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Higuchi, Y.; Tanabe, K.; Katakura, Y.; Takegawa, K. Secretory production of N-glycan-deleted glycoprotein in Aspergillus oryzae. J. Biosci. Bioeng. 2020, 129, 573–580. [Google Scholar] [CrossRef]
- Kimura, S.; Maruyama, J.-i.; Watanabe, T.; Ito, Y.; Arioka, M.; Kitamoto, K. In vivo imaging of endoplasmic reticulum and distribution of mutant α-amylase in Aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 1044–1054. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Ishikawa, E.; Shoji, J.-y.; Nakano, H.; Kitamoto, K. Septum-directed secretion in the filamentous fungus Aspergillus oryzae. Mol. Microbiol. 2011, 81, 40–55. [Google Scholar] [CrossRef]
- Srikanth, M.; Sandeep, T.S.R.S.; Sucharitha, K.; Godi, S. Biodegradation of plastic polymers by fungi: A brief review. Bioresour. Bioprocess. 2022, 9, 42. [Google Scholar] [CrossRef]
- Morilla, E.A.; Stegmann, P.M.; Tubio, G. Enzymatic cocktail production by a co-cultivation Solid-State Fermentation for detergent formulation. Food Bioprod. Process. 2023, 140, 110–121. [Google Scholar] [CrossRef]
- El-Wafai, N.A.; Farrag, A.M.I.; Abdel-Basit, H.M.; Hegazy, M.I.; Al-Goul, S.T.; Ashkan, M.F.; Al-Quwaie, D.A.; Alqahtani, F.S.; Amin, S.A.; Ismail, M.N.; et al. Eco-Friendly Degradation of Natural Rubber Powder Waste Using Some Microorganisms with Focus on Antioxidant and Antibacterial Activities of Biodegraded Rubber. Processes 2023, 11, 2350. [Google Scholar] [CrossRef]
- Varjani, S.; Upasani, V.N. Bioaugmentation of Pseudomonas aeruginosa NCIM 5514—A novel oily waste degrader for treatment of petroleum hydrocarbons. Bioresour. Technol. 2021, 319, 124240. [Google Scholar] [CrossRef] [PubMed]
- Chrysikou, L.P.; Dagonikou, V.; Dimitriadis, A.; Bezergianni, S. Waste cooking oils exploitation targeting EU 2020 diesel fuel production: Environmental and economic benefits. J. Clean. Prod. 2019, 219, 566–575. [Google Scholar] [CrossRef]
- Orjuela, A.; Clark, J. Green chemicals from used cooking oils: Trends, challenges, and opportunities. Curr. Opin. Green Sustain. Chem. 2020, 26, 100369. [Google Scholar] [CrossRef] [PubMed]
- Nasrabadi, A.E.; Ramavandi, B.; Bonyadi, Z. Recent progress in biodegradation of microplastics by Aspergillus sp. in aquatic environments. Colloid Interface Sci. Commun. 2023, 57, 100754. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Chen, X.; Siewers, V. CRISPR/Cas9-mediated point mutations improve α-amylase secretion in Saccharomyces cerevisiae. FEMS Yeast Res. 2022, 22, foac033. [Google Scholar] [CrossRef]
- Pham, T.A.; Tran, L.N.; Dam, T.H.; To, K.A. Valorization of Cassava Bagasse Using Co-culture of Aspergillus oryzae VS1 and Sporidiobolus pararoseus O1 in Solid-State Fermentation. Waste Biomass Valori 2022, 13, 3003–3012. [Google Scholar] [CrossRef]
- Song, L.; Chen, Y.; Du, Y.; Wang, X.; Guo, X.; Dong, J.; Xiao, D. Saccharomyces cerevisiae proteinase A excretion and wine making. World J. Microbiol. Biotechnol. 2017, 33, 210. [Google Scholar] [CrossRef]
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef]
- Shah, A.V.; Singh, A.; Sabyasachi Mohanty, S.; Kumar Srivastava, V.; Varjani, S. Organic solid waste: Biorefinery approach as a sustainable strategy in circular bioeconomy. Bioresour. Technol. 2022, 349, 126835. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Niu, Y.; Jin, Q.; Qin, K.; Wang, L.; Shang, Y.; Zeng, B.; Hu, Z. Identification of Six Thiolases and Their Effects on Fatty Acid and Ergosterol Biosynthesis in Aspergillus oryzae. Appl. Environ. Microbiol. 2022, 88, e02372-21. [Google Scholar] [CrossRef]
- Uwineza, C.; Mahboubi, A.; Atmowidjojo, A.; Ramadhani, A.; Wainaina, S.; Millati, R.; Wikandari, R.; Niklasson, C.; Taherzadeh, M.J. Cultivation of edible filamentous fungus Aspergillus oryzae on volatile fatty acids derived from anaerobic digestion of food waste and cow manure. Bioresour. Technol. 2021, 337, 125410. [Google Scholar] [CrossRef] [PubMed]
- Nazir, M.T.; Soufiani, A.M.; Ferreira, J.A.; Sar, T.; Taherzadeh, M.J. Production of filamentous fungal biomass with increased oil content using olive oil as a carbon source. J. Chem. Technol. Biotechnol. 2022, 97, 2626–2635. [Google Scholar] [CrossRef]
- Iwamoto, N.; Sasaki, A.; Maizawa, T.; Hamada-Sato, N. Abalone Viscera Fermented with Aspergillus oryzae 001 Prevents Pressure Elevation by Inhibiting Angiotensin Converting Enzyme. Nutrients 2023, 15, 947. [Google Scholar] [CrossRef]
- Parchami, M.; Ferreira, J.A.; Taherzadeh, M.J. Brewing Process Development by Integration of Edible Filamentous Fungi to Upgrade the Quality of Brewer’s Spent Grain (BSG). Bioresources 2021, 16, 1686–1701. [Google Scholar] [CrossRef]
- Bangar, S.P.; Sandhu, K.S.; Purewal, S.S.; Kaur, M.; Kaur, P.; Siroha, A.K.; Kumari, K.; Singh, M.; Kumar, M. Fermented barley bran: An improvement in phenolic compounds and antioxidant properties. J. Food Process. Preserv. 2022, 46, e15543. [Google Scholar] [CrossRef]
- Ikram Ul, H.; Ashraf, S.; Nawaz, A.; Arshad, Y.; Mukhtar, H. Biosynthesis of β-galactosidase from Aspergillus oryzae using milk powder as substrate. Pak. J. Bot. 2021, 53, 273–279. [Google Scholar] [CrossRef]
- Kawa-Rygielska, J.; Pietrzak, W.; Lennartsson, P.R. High-Efficiency Conversion of Bread Residues to Ethanol and Edible Biomass Using Filamentous Fungi at High Solids Loading: A Biorefinery Approach. Appl. Sci. 2022, 12, 6405. [Google Scholar] [CrossRef]
- Ozturk, A.B.; Al-Shorgani, N.K.N.; Cheng, S.; Arasoglu, T.; Gulen, J.; Habaki, H.; Egashira, R.; Kalil, M.S.; Yusoff, W.M.W.; Cross, J.S. Two-step fermentation of cooked rice with Aspergillus oryzae and Clostridium acetobutylicum YM1 for biobutanol production. Biofuels 2022, 13, 579–585. [Google Scholar] [CrossRef]
- Guo, H.-N.; Wu, S.-B.; Tian, Y.-J.; Zhang, J.; Liu, H.-T. Application of machine learning methods for the prediction of organic solid waste treatment and recycling processes: A review. Bioresour. Technol. 2021, 319, 124114. [Google Scholar] [CrossRef]
- Sirohi, R.; Kumar Gaur, V.; Kumar Pandey, A.; Jun Sim, S.; Kumar, S. Harnessing fruit waste for poly-3-hydroxybutyrate production: A review. Bioresour. Technol. 2021, 326, 124734. [Google Scholar] [CrossRef] [PubMed]
- Chilakamarry, C.R.; Sakinah, A.M.M.; Zularisam, A.W.; Pandey, A. Glycerol waste to value added products and its potential applications. Syst. Microbiol. Biomanufacturing 2021, 1, 378–396. [Google Scholar] [CrossRef]
- Chaitanya Reddy, C.; Khilji, I.A.; Gupta, A.; Bhuyar, P.; Mahmood, S.; Saeed Al-Japairai, K.A.; Chua, G.K. Valorization of keratin waste biomass and its potential applications. J. Water Process Eng. 2021, 40, 101707. [Google Scholar] [CrossRef]
- Sharma, P.; Gaur, V.K.; Sirohi, R.; Varjani, S.; Hyoun Kim, S.; Wong, J.W.C. Sustainable processing of food waste for production of bio-based products for circular bioeconomy. Bioresour. Technol. 2021, 325, 124684. [Google Scholar] [CrossRef] [PubMed]
- Banat, I.M.; Carboué, Q.; Saucedo-Castañeda, G.; de Jesús Cázares-Marinero, J. Biosurfactants: The green generation of speciality chemicals and potential production using Solid-State fermentation (SSF) technology. Bioresour. Technol. 2021, 320, 124222. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, B.; Feng, X.; Zhang, M.; Ding, T.; Zhao, Y.; Wang, C. Comparative proteome and volatile metabolome analysis of Aspergillus oryzae 3.042 and Aspergillus sojae 3.495 during koji fermentation. Food Res. Int. 2023, 165, 112527. [Google Scholar] [CrossRef]
- Robazza, A.; Welter, C.; Kubisch, C.; Baleeiro, F.C.; Ochsenreither, K.; Neumann, A. Co-Fermenting Pyrolysis Aqueous Condensate and Pyrolysis Syngas with Anaerobic Microbial Communities Enables L-Malate Production in a Secondary Fermentative Stage. Fermentation 2022, 8, 512. [Google Scholar] [CrossRef]
- Abid, K.; Jabri, J.; Yaich, H.; Malek, A.; Rekhis, J.; Kamoun, M. Nutritional value assessments of peanut hulls and valorization with exogenous fibrolytic enzymes extracted from a mixture culture of Aspergillus strains and Neurospora intermedia. Biomass Convers. Biorefinery 2022, 1–9. [Google Scholar] [CrossRef]
- Danner, C.; Mach, R.L.; Mach-Aigner, A.R. The phenomenon of strain degeneration in biotechnologically relevant fungi. Appl. Microbiol. Biotechnol. 2023, 107, 4745–4758. [Google Scholar] [CrossRef]
- Chi, Z.; Kong, C.-C.; Wang, Z.-Z.; Wang, Z.; Liu, G.-L.; Hu, Z.; Chi, Z.-M. The signaling pathways involved in metabolic regulation and stress responses of the yeast-like fungi Aureobasidium spp. Biotechnol. Adv. 2022, 55, 107898. [Google Scholar] [CrossRef] [PubMed]
- Sakekar, A.A.; Gaikwad, S.R.; Punekar, N.S. Protein expression and secretion by filamentous fungi. J. Biosci. 2021, 46, 5. [Google Scholar] [CrossRef]
- Li, Q.; Lu, J.; Liu, J.; Li, J.; Zhang, G.; Du, G.; Chen, J. High-throughput droplet microfluidics screening and genome sequencing analysis for improved amylase-producing Aspergillus oryzae. Biotechnol. Biofuels Bioprod. 2023, 16, 185. [Google Scholar] [CrossRef]
- Ntana, F.; Mortensen, U.H.; Sarazin, C.; Figge, R. Aspergillus: A Powerful Protein Production Platform. Catalysts 2020, 10, 1064. [Google Scholar] [CrossRef]
- Karaman, E.; Eyüpoğlu, A.E.; Mahmoudi Azar, L.; Uysal, S. Large-Scale Production of Anti-RNase A VHH Expressed in pyrG Auxotrophic Aspergillus oryzae. Curr. Issues Mol. Biol. 2023, 45, 4778–4795. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Gene Source | Substrate | Strain | Yield | Reference |
|---|---|---|---|---|---|
| 2,4′-dihydroxy-3′-methoxypropiophenone | Congeneric | Sucrose | RIB40 | - | [27] |
| Pyrones | Congeneric | Solid rice medium | - 1 | - | [28] |
| N, N-dimethyldecylamine N-oxide | Congeneric | Potato dextrose agar | MK674278 | - | [29] |
| Orsellinic acid | Heterologous | Maltose | NSAR1 | 340.41 mg/Kg | [30] |
| Non-ribosomal peptide pigments | Congeneric | Glucose | BCC 7051 | 265.09 ± 14.74 mg/L·d | [24] |
| Flavonoid aglycones and cajaninstilbene acid | Congeneric | Cajanus cajan cell suspension cultures | Y-29 | - | [31] |
| L-malic acid | Congeneric | Acetate | DSM 1863 | 12.08 ± 1.25 g/L | [32] |
| L-malic acid | Congeneric | Glucose | DSM 1863 | 178 g/L | [33] |
| L-malic acid | Congeneric | Glucose | DSM 1863 | - | [34] |
| L-malic acid | Congeneric | Glucose | FMME-S-38 | 164.9 g/L | [35] |
| KA | Congeneric | Glucose | - | 139.24 g/L | [36] |
| Glucosamine | Congeneric | Potato dextrose broth | NCH-42 | 0.31 g/g | [37] |
| Strain | Gene Name | Encoded Protein | Acting Site | Genetic Manipulation | KA Yield | Reference |
|---|---|---|---|---|---|---|
| 3.042 | kojR | Zn2+2Cys6 transcriptional activator | - | Overexpressed | 32.5 g/L | [12] |
| RB40 | hirA | Histone chaperon | Transcription | - | - | [48] |
| 3.042 | Aokap2 | Cell surface ferric reductase | laeA & kojA | Overexpressed | Increased | [49] |
| 3.042 | Aokap5 | C2H2-type zinc-finger protein | kojT promotor | Overexpressed | Increased | [50] |
| 3.042 | Aokap1 | Kojic acid related protein 1 | kojA, kojR and kojT | Disrupted | Increased | [51] |
| 3.042 | AozfA | Zinc finger protein | Transcriptional activator | Overexpressed | Reduced | [52] |
| RB40 | Aoatg8 | Enables phosphatidylethanolamine binding activity and protein tag | Autophagy | Disrupted | Increased | [46] |
| RB40 | AoZip2 | IRT-like protein | Response of metal ions | Overexpressed | Reduced | [47] |
| Strain | Enzyme | Source | Position | Acetive | Reference |
|---|---|---|---|---|---|
| MN894021 | Xylanase | Homologous | Extracellular | 0.37 U/mL | [61] |
| ATCC 10124 | Xylanase | Homologous | Extracellular | 11.90 U/g DS 1 | [62] |
| SBS50 | Phytase | Homologous | Extracellular | 506.12 U/g | [63] |
| BM-DIA | Fructosyltransferase | Homologous | Extracellular | 1.59 U/mL | [64] |
| - | α-amylase | Homologous | Extracellular | 9868.12 U/gds | [65] |
| NRRL695 | α-amylase | Homologous | Extracellular | 14.076 U/mL | [66] |
| S719 | β-fructofuranosidase | Homologous | Extracellular | 155.4 U/mL | [67] |
| S719 | Fructosyltransferase | Homologous | Extracellular | 12 U/mL | [68] |
| NRRL695 | Tannase | Homologous | Intracellular | - | [69] |
| NRRL1560 | 1,4-α-D-glucan glucohydrolase | Homologous | Extracellular | - | [70] |
| DRDFS13 | Milk-clotting protease | Homologous | Extracellular | 137.58 U/mL | [71] |
| ISL-9 | Pectin lyase | Homologous | Extracellular | 9.26 U/mL | [72] |
| HML366 | Endoglucanase | Homologous | Extracellular | - | [73] |
| AOK11 | Recombinant tannase | Heterogenous | Extracellular | - | [74] |
| NSPlD1 | Polyketide synthase | Heterogenous | Intracellular | - | [75] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Wu, Y.; Long, S.; Feng, S.; Jia, X.; Hu, Y.; Ma, M.; Liu, J.; Zeng, B. Aspergillus oryzae as a Cell Factory: Research and Applications in Industrial Production. J. Fungi 2024, 10, 248. https://doi.org/10.3390/jof10040248
Sun Z, Wu Y, Long S, Feng S, Jia X, Hu Y, Ma M, Liu J, Zeng B. Aspergillus oryzae as a Cell Factory: Research and Applications in Industrial Production. Journal of Fungi. 2024; 10(4):248. https://doi.org/10.3390/jof10040248
Chicago/Turabian StyleSun, Zeao, Yijian Wu, Shihua Long, Sai Feng, Xiao Jia, Yan Hu, Maomao Ma, Jingxin Liu, and Bin Zeng. 2024. "Aspergillus oryzae as a Cell Factory: Research and Applications in Industrial Production" Journal of Fungi 10, no. 4: 248. https://doi.org/10.3390/jof10040248