

New Data on Boletaceae (Agaricomycetes, Basidiomycota) from Central Vietnam with Description of Two New Species and Creation of a New Combination Based on Morphological and Phylogenetic Evidence
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collecting and Site Description
2.2. Morphological Study
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Alignment and Phylogenetic Analyses
3. Results
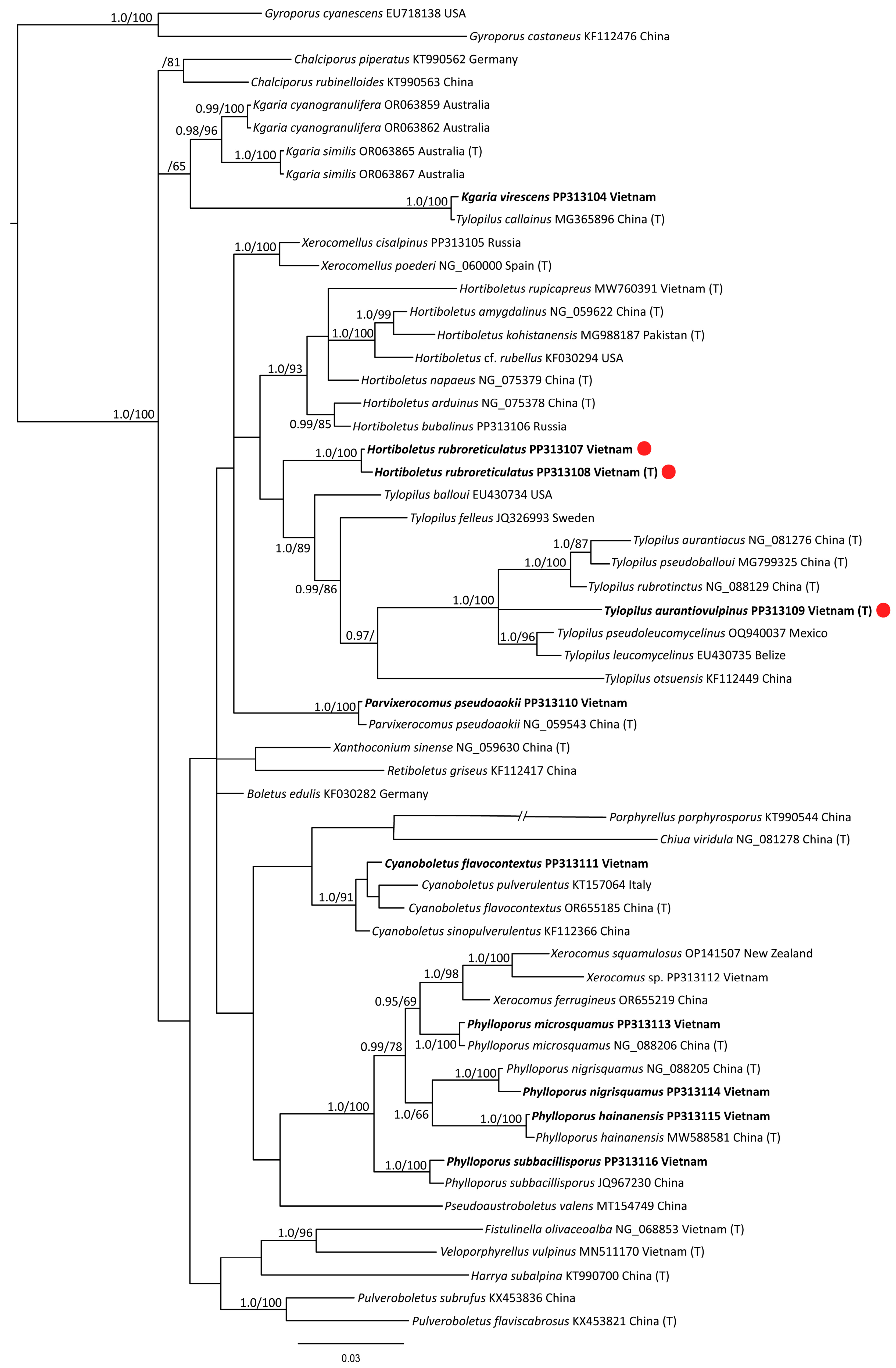
3.1. Phylogenetic Analysis
3.2. Taxonomy
3.2.1. New Species
3.2.2. Annotated List of the Species of the Boletoid Fungi Recorded for the First Time in Vietnam
- Cyanoboletus flavocontextus L. Fan, N. Mao & T.Y. Zhao, Mycosphere 14(1): 2013–2091 (2023).
- 2.
- Kgaria virescens (Har. Takah. & Taneyama) O.V. Morozova, E.S. Popov, T.H.G. Pham, comb. nov.
- 3.
- Parvixerocomus pseudoaokii G. Wu, Kuan Zhao & Zhu L. Yang, in Wu, Zhao, Li, Zeng, Feng, Halling & Yang, Fungal Diversity: 10.1007/s13225-015-0322-0, (12) (2015).
- 4.
- Phylloporus hainanensis N.K. Zeng, L.L. Wu & Zhi Q. Liang, in Xue, Zhang, Xu, Xie, Wu, Wang, Tang, Hao, Zhao, Jiang, Li, Yang, Li, Liang & Zeng, Stud. Mycol. 106: 170 (2023).
- 5.
- Phylloporus microsquamus N.K. Zeng, L.L. Wu, S. Jiang & Z.Q. Liang, in Wu, Liang, Su, Fan, Zhang, Jiang, Chen, Hao & Zeng, Mycol. Progr. 20(10): 1262 (2021).
- 6.
- Phylloporus nigrisquamus N.K. Zeng, L.L. Wu & Y.G. Fan, Mycological Progress 20 (10): 1264 (2021).
- 7.
- Phylloporus subbacillisporus Raspé, K.D. Hyde & Chuankid, Mycol. Progr. 18 (5): 608 (2019).
- 8.
- Tylopilus rubrotinctus Yan C. Li & Zhu L. Yang, The Boletes of China: Tylopilus s.l. (Singapore): 339 (2021).
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watling, R. A Manual and Source Book on the Boletes and Their Allies; Fungiflora: Oslo, Norway, 2008; 249p. [Google Scholar]
- Pham, T.H.G.; Popov, E.S.; Morozova, O.V.; Nguyen, D.H. Basidiomycetous Macromycetes of the Tay Nguyen Plateau in Vietnam: The History of Investigation. Mikol. Fitopatol. 2018, 52, 306–318. [Google Scholar]
- Pham, T.H.G.; Morozova, O.V. Boletoid fungi (Boletaceae, Basidiomycota) of the Bidoup—Nui Ba National Park (Vietnam). Turczaninowia 2020, 23, 88–98. [Google Scholar] [CrossRef]
- Tran, T.T.H.; Tran, H.T.; Le, B.D.; Nguyen, P.D.N. Diversity of Amanita genus in Kon Ka Kinh National Park in Gia Lai Province, Viet Nam. In Proceedings of the 7th National Scientific Conference on Ecology and Biological Resources, Hanoi, Vietnam, 20 October 2017; Publishing House for Science and Technology: Ha Noi, Vietnam, 2017; pp. 702–709. (In Vietnamese). [Google Scholar]
- Morozova, O.V.; Noordeloos, M.E.; Popov, E.S.; Alexandrova, A.V. Three new species within the genus Entoloma (Basidiomycota, Agaricales) with clamped basidia and a serrulatum-type lamellae edge, and their phylogenetic position. Mycol. Prog. 2018, 17, 381–392. [Google Scholar] [CrossRef]
- Malysheva, E.F.; Malysheva, V.F.; Alexandrova, A.A.; Morozova, O.V. Observations on Pluteaceae in Vietnam. 1. New species and new records of Volvariella. Phytotaxa 2019, 408, 233–254. [Google Scholar] [CrossRef]
- Malysheva, E.; Malysheva, V.; Alexandrova, A.; Morozova, O. Observations on Pluteaceae in Vietnam. 2. One new record and ten new species of Pluteus. Phytotaxa 2020, 461, 79–107. [Google Scholar] [CrossRef]
- Malysheva, E.; Popov, E.; Morozova, O.; Dudka, V.; Pham, T.H.G.; Malysheva, V. Observations on Pluteaceae in Vietnam: Four New Species and New Records of Pluteus. J. Fungi 2023, 9, 584. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.H.G.; Morozova, O.V.; Alexandrova, A.V. Boletoid fungi (Boletaceae, Basidiomycota) of protected areas of Kon Tum Plateau (Central Highlands of Vietnam). Turczaninowia 2021, 24, 65–76. [Google Scholar] [CrossRef]
- Malysheva, E.; Popov, E. Observations on Pluteaceae in Vietnam. 3. One new species of Volvariella. Phytotaxa 2022, 545, 278–286. [Google Scholar] [CrossRef]
- Morozova, O.; Popov, E.; Alexandrova, A.; Pham, T.H.G.; Noordeloos, M.E. Four new species of Entoloma (Entolomataceae, Agaricomycetes) subgenera Cyanula and Claudopus from Vietnam and their phylogenetic position. Phytotaxa 2022, 549, 1–21. [Google Scholar] [CrossRef]
- Nguyen, P.D.N.; Nguyen, H.K.; Tran, T.K.T.; Tran, T.T.H. Study of morphological features and DNA barcodes of two species of fungi belonging to the genus Phylloporus were collected at Chu Yang Sin National Park. In Proceedings of the 4th National Mycology Conference, Can Tho, Vietnam, 24–26 June 2022; Publishing House for Science and Technology: Ha Noi, Vietnam, 2022; pp. 187–198. (In Vietnamese). [Google Scholar] [CrossRef]
- Morozova, O.; Pham, T.H.G. New Species of Entoloma Subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from Central Vietnam. J. Fungi 2023, 9, 621. [Google Scholar] [CrossRef] [PubMed]
- Perreau, J.; Joly, P. Sur quelques Boletales de la flore du Viet-Nam. Bull. Soc. Mycol. France 1964, 80, 385–395. [Google Scholar]
- Le, B.D. Mycoflora of family Boletaceae of Highland area. VNU J. Sci. Nat. Sci. Technol. 2001, 17, 25–28. [Google Scholar]
- Le, B.D. Macrofungi of Tay Nguyen; Publishing House for Science and Technology: Ha Noi, Vietnam, 2003; 283p. (In Vietnamese) [Google Scholar]
- Kiet, T.T. The Larger Fungi of Vietnam; Publishing House for Science and Technology: Ha Noi, Vietnam, 2012; Volume 2, 413p. (In Vietnamese) [Google Scholar]
- Ngo, A.; Nguyen, T.P. Diversity of macromycoflora in the Phong Dien district, Thua Thien-Hue province, Vietnam. In Proceedings of the 6th National Scientific Conference on Ecology and Biological Resources, Hanoi, Vietnam, 21 October 2015; Long, K.D., Ed.; Publishing House for Science and Technology: Ha Noi, Vietnam, 2015; pp. 454–460. [Google Scholar]
- Pham, N.K.H.; Phan, H.H.; Do, T.T.L.; Le, T.C.; Truong, B.N. Preliminary investigation on macrofungal flora along the provincial road no. 723 belonging to Bidoup–Nui Ba National Park (Lam Dong Province). In Proceedings of the 6th National Scientific Conference on Ecology and Biological Resources, Hanoi, Vietnam, 21 October 2015; Long, K.D., Ed.; Publishing House for Science and Technology: Ha Noi, Vietnam, 2015; pp. 556–562. (In Vietnamese). [Google Scholar]
- Do, H.T.; Nguyen, P.D.N. Composition of edible macrofungi species of Boletus genus in Chu Yang Sin National Park, Dak Lak Province. In Proceedings of the 7th National Scientific Conference on Ecology and Biological Resources, Hanoi, Vietnam, 20 October 2017; Publishing House for Science and Technology: Ha Noi, Vietnam, 2017; pp. 1489–1495. (In Vietnamese). [Google Scholar]
- Patouillard, N. Quelques champignons de l’Annam. Bull. Soc. Mycol. Fr. 1909, 25, 1–12. [Google Scholar]
- Dörfelt, H.; Kiet, T.T.; Berg, A. Neue Makromyceten-Kollektionen von Vietnam und deren systematische und ökogeographische Bedeutung. Feddes Rep. 2004, 115, 164–177. [Google Scholar] [CrossRef]
- Pham, N.D.H.; Takahashi, H.; Fukiharu, T.; Shimizu, K.; Le, B.D.; Suzuki, A. Phlebopus spongiosus sp. nov. (Boletales, Boletinellaceae) with a sponge-like tissue. Mycotaxon 2012, 119, 27–34. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Lombard, L.; Roets, F.; Swart, W.J.; Alvarado, P.; Carnegie, A.J.; Moreno, G.; Luangsa-ard, J.; Thangavel, R.; et al. Fungal Planet description sheets: 951–1041. Persoonia 2019, 43, 223–425. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.W.; Dima, B.; Dovana, F.; et al. Fungal Planet description sheets: 1112–1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Osieck, E.R.; Jurjević, Ž.; Boers, J.; van Iperen, A.L.; Starink-Willemse, M.; Dima, B.; Balashov, S.; Bulgakov, T.S.; Johnston, P.R.; et al. Fungal Planet description sheets: 1284–1382. Persoonia 2021, 47, 178–374. [Google Scholar] [CrossRef] [PubMed]
- Haelewaters, D.; Dima, B.; Abdel-Hafiz, B.I.I.; Abdel-Wahab, M.A.; Abul-Ezz, S.R.; Acar, I.; Aguirre-Acosta, E.; Aime, M.C.; Aldemir, S.; Ali, M.; et al. Fungal Systematics and Evolution: FUSE 6. Sydowia 2020, 72, 231–356. [Google Scholar]
- Lebeuf, R.; Alexandrova, A.V.; Cerna-Mendoza, A.; Corazon-Guivin, M.A.; da Silva, G.A.; de la Sota-Ricaldi, A.M.; Dima, B.; Fryssouli, V.; Gkilas, M.; Guerrero-Abad, J.C.; et al. Fungal Systematics and Evolution: FUSE 8. Sydowia 2021, 74, 193–249. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Kuznetsov, A.N. Biodiversity and Ecological Characteristics of Bidoup—Nui Ba National Park; Natural Science and Technology Publishing House: Ha Noi, Vietnam, 2011; pp. 18–27. (In Vietnamese) [Google Scholar]
- Tordoff, A.W.; Tran, Q.B.; Nguyen, D.T.; Le, M.H. (Eds.) Sourcebook of Existing and Proposed Protected Areas in Vietnam, 2nd ed.; CD-ROOM; Birdlife International in Indochina and Ministry of Agriculture and Rural Development: Hanoi, Vietnam, 2004; Available online: https://thiennhienviet.org.vn/sourcebook/source_book/index_EN.html (accessed on 15 December 2022).
- Nguyen, T.T. Study on structural characteristics, regeneration and determination of natural forest resilience in Ta Dung National Park, Dak Nong province. Summ. Rep. Sci. Technol. Top. 2021, 22–36. (In Vietnamese) [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978; pp. 1–252. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-a sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Feng, B.; Xu, J.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nuhn, M.E.; Binder, M.; Taylor, A.F.; Halling, R.E.; Hibbett, D.S. Phylogenetic overview of the Boletineae. Fungal Biol. 2013, 117, 479–511. [Google Scholar] [CrossRef]
- Loizides, M.; Bellanger, J.M.; Assyov, B.; Moreau, P.A.; Richard, F. Present status and future of boletoid fungi (Boletaceae) on the island of Cyprus: Cryptic and threatened diversity unraveled by ten-year study. Fungal Ecol. 2019, 41, 65–81. [Google Scholar] [CrossRef]
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Han, L.H.; Cui, Y.Y.; Li, F.; Xu, J.P.; Yang, Z.L. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Mao, N.; Zhao, T.Y.; Zhang, Y.X.; Li, T.; Lv, J.C.; Fan, L. Boletaceae from Shanxi Province of northern China with descriptions of ten new species. Mycosphere 2023, 14, 2013–2091. [Google Scholar]
- Biketova, A.Y.; Rinaldi, A.C.; Simonini, G. Nomenclatural novelties: Cyanoboletus mediterraneensis Biketova, A. Rinaldi & Simonini, sp. nov. Index Fungorum 2022, 516, 1. [Google Scholar]
- Gelardi, M.; Simonini, G.; Ercole, E.; Davoli, P.; Vizzini, A. Cupreoboletus (Boletaceae, Boletineae), a new monotypic genus segregated from Boletus sect. Luridi to reassign the Mediterranean species B. poikilochromus. Mycologia 2015, 107, 1254–1269. [Google Scholar] [CrossRef]
- Gelardi, M.; Vizzini, A.; Ercole, E.; Voyron, S.; Sun, J.-Z.; Liu, X.-Z. Boletus sinopulverulentus, a new species from Shaanxi Province (central China) and notes on Boletus and Xerocomus. Sydowia 2013, 65, 45–57. [Google Scholar]
- Vadthanarat, S.; Raspé, O.; Lumyong, S. Phylogenetic affinities of the sequestrate genus Rhodactina (Boletaceae), with a new species, R rostratispora from Thailand. MycoKeys 2018, 29, 63–80. [Google Scholar] [CrossRef]
- Binder, M.; Larsson, K.H.; Matheny, P.B.; Hibbett, D.S. Amylocorticiales ord. nov. and Jaapiales ord. nov.: Early diverging clades of agaricomycetidae dominated by corticioid forms. Mycologia 2010, 102, 865–880. [Google Scholar] [CrossRef]
- Palmer, J.M.; Lindner, D.L.; Volk, T.J. Ectomycorrhizal characterization of an American chestnut (Castanea dentata)-dominated community in Western Wisconsin. Mycorrhiza 2008, 19, 27–36. [Google Scholar] [CrossRef]
- Wilson, A.W.; Binder, M.; Hibbett, D.S. Diversity and evolution of ectomycorrhizal host associations in the Sclerodermatineae (Boletales, Basidiomycota). New Phytol. 2012, 194, 1079–1095. [Google Scholar] [CrossRef]
- Kim, M.; Lee, J.S.; Park, J.Y.; Kim, C. First Report of Six Macrofungi from Daecheongdo and Socheongdo Islands, Korea. Mycobiology 2021, 49, 454–460. [Google Scholar] [CrossRef]
- Xie, H.-J.; Lin, W.-F.; Jiang, S.; Xue, R.; Wu, L.-L.; Zhang, Y.-Z.; Liang, Z.-Q.; Zeng, N.-K.; Su, M.-S. Two new species of Hortiboletus (Boletaceae, Boletales) from China. Mycol. Prog. 2020, 19, 1377–1386. [Google Scholar] [CrossRef]
- Das, K.; Chakraborty, D.; Baghela, A.; Singh, S.K.; Dentinger, B.T. New species of xerocomoid boletes (Boletaceae) from Himalayan India based on morphological and molecular evidence. Mycologia 2016, 108, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Naseer, A.; Sarwar, S.; Khalid, A.N.; Smith, M.E. Hortiboletus kohistanensis (Boletaceae), a new bolete species from temperate and subalpine oak forests of Pakistan. Phytotaxa 2019, 388, 239–246. [Google Scholar] [CrossRef]
- Halling, R.E.; Fechner, N.A.; Holmes, G.; Davoodian, N. Kgaria (Boletaceae, Boletoideae) gen. nov. in Australia: Neither a Tylopilus nor a Porphyrellus. Fungal Syst. Evol. 2023, 12, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Zhao, K.; Li, Y.C.; Zeng, N.K.; Feng, B.; Halling, R.; Yang, Z.L. Four new genera of the fungal family Boletaceae. Fungal Divers. 2015, 81, 1–24. [Google Scholar] [CrossRef]
- B Patil, P.; Gunasekaran, S.; Singh, S.K.; Vaidya, S. Parvixerocomus matheranensis (Boletaceae), a new species from India. Mycoscience 2021, 62, 244–249. [Google Scholar] [CrossRef]
- Wu, L.L.; Liang, Z.Q.; Su, M.S.; Fan, Y.G.; Zhang, P.; Jiang, S.; Chen, Y.L.; Liu, Y.; Hao, Y.J.; Zeng, N.K. Updated taxonomy of Chinese Phylloporus (Boletaceae, Boletales): Six new taxa and four redescribed species. Mycol. Prog. 2021, 20, 1243–1273. [Google Scholar] [CrossRef]
- Xue, R.; Zhang, X.; Xu, C.; Xie, H.J.; Wu, L.L.; Wang, Y.; Tang, L.P.; Hao, Y.J.; Zhao, K.; Jiang, S.; et al. The subfamily Xerocomoideae (Boletaceae, Boletales) in China. Stud. Mycol. 2023, 106, 95–197. [Google Scholar] [CrossRef]
- Chuankid, B.; Vadthanarat, S.; Hyde, K.D.; Thongklang, N.; Zhao, R.; Lumyong, S.; Raspé, O. Three new Phylloporus species from tropical China and Thailand. Mycol. Prog. 2019, 18, 603–614. [Google Scholar] [CrossRef]
- Zeng, N.K.; Tang, L.P.; Li, Y.C.; Tolgor, B.; Zhu, X.T.; Zhao, Q.; Yang, Z.L. The genus Phylloporus (Boletaceae, Boletales) from China: Morphological and multilocus DNA sequence analyses. Fungal Divers. 2013, 58, 73–101. [Google Scholar] [CrossRef]
- Li, Y.C.; Yang, Z.L. The Boletes of China: Tylopilus s.l.; Science Press & Springer Nature: Singapore, 2021. [Google Scholar] [CrossRef]
- Zeng, N.K.; Liang, Z.Q.; Tang, L.P.; Li, Y.C.; Yang, Z.L. The genus Pulveroboletus (Boletaceae, Boletales) in China. Mycologia 2017, 109, 422–442. [Google Scholar] [CrossRef]
- Halling, R.E.; Osmundson, T.W.; Neves, M.A. Pacific boletes: Implications for biogeographic relationships. Mycol. Res. 2008, 112, 437–447. [Google Scholar] [CrossRef]
- Liang, Z.-Q.; Su, M.-S.; Jiang, S.; Hong, D.; Zeng, N.-K. Tylopilus callainus, a new species with a sea-green color change of hymenophore and context from the south of China. Phytotaxa 2018, 343, 269–276. [Google Scholar] [CrossRef]
- Halling, R.E.; Nuhn, M.; Fechner, N.A.; Osmundson, T.W.; Soytong, K.; Arora, D.; Hibbett, D.S.; Binder, M. Sutorius: A new genus for Boletus eximius. Mycologia 2012, 104, 951–961. [Google Scholar] [CrossRef]
- Chakraborty, D.; Vizzini, A.; Das, K. Two new species and one new record of the genus Tylopilus (Boletaceae) from Indian Himalaya with morphological details and phylogenetic estimations. MycoKeys 2018, 33, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Vasquez, O.; Perez-Moreno, J.; Pinzon, J.P.; Garibay-Orijel, R.; Garcia-Jimenez, J.; de la Fuente, J.I.; Venegas-Barrera, C.S.; Martinez-Reyes, M.; Montoya, L.; Bandala, V.; et al. Broadening the Knowledge of Mexican Boletes: Addition of a New Genus, Seven New Species, and Three New Combinations. J. Fungi 2023, 9, 1126. [Google Scholar] [CrossRef] [PubMed]
- Farid, A.; Bessette, A.E.; Bessette, A.R.; Bolin, J.A.; Kudzma, L.V.; Franck, A.R.; Garey, J.R. Investigations in the boletes (Boletaceae) of southeastern USA: Four novel species, and three novel combinations. Mycosphere 2021, 12, 1038–1076. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M.; Le Roux, J.J.; Strasberg, D.; Edwards, J.; Roets, F.; Hubka, V.; Taylor, P.W.J.; Heykoop, M.; et al. Fungal Planet description sheets: 400–468. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef] [PubMed]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R.; et al. Fungal diversity notes 111–252—Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Taylor, A.F.; Hills, A.E.; Simonini, G.; Both, E.E.; Eberhardt, U. Detection of species within the Xerocomus subtomentosus complex in Europe using rDNA-ITS sequences. Mycol. Res. 2006, 110, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Gao, Y.; Song, H.-Y.; Hu, H.-J.; Yang, W.-J.; Zhang, W.; Liao, L.-Y.; Fang, Y.; Cheng, L.; Hu, D.-M. Phylogenetic and Morphological Evidence Reveal Five New Species of Boletes from Southern China. J. Fungi 2023, 9, 814. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization, Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.; Liang, Z.-Q.; Xue, R.; Jiang, S.; Luo, S.-H.; Wang, Y.; Wu, L.-L.; Tang, L.-P.; Chen, Y.; Hong, D.; et al. New and noteworthy boletes from subtropical and tropical China. MycoKeys 2019, 46, 55–96. [Google Scholar] [CrossRef] [PubMed]
- Peck, C.H. Report of the State Botanist 1911. Bull. N. Y. St. Mus. 1912, 157, 1–139. [Google Scholar]
- Horak, E. Revision of Malaysian species of Boletales s. l. (Basidiomycota) describeb by E. J. H. Corner (1972, 1974). Malay. For. Rec. 2011, 51, 1–283. [Google Scholar]
- Osmundson, T.W.; Halling, R.E. Tylopilus oradivensis sp. nov.: A newly described member of the Tylopilus balloui complex from Costa Rica. Mycotaxon 2010, 113, 475–483. [Google Scholar] [CrossRef]
- Osmundson, T.W.; Bougher, N.L.; Robinson, R.M.; Halling, R.E. Rubinoboletus phaseolisporus (Boletaceae) from Western Australia is a Tylopilus with bean-shaped spores. Nuytsia 2021, 32, 87–97. [Google Scholar]
- Montoya, L.; Ramos, A.; Halling, R.E.; Bandala, V.M. A new species and a new record of Tylopilus (Boletaceae) of the balloui group in lowland and montane forests from Eastern Mexico. Mycol. Progress 2023, 22, 6. [Google Scholar] [CrossRef]
- Magnago, A.C.; Reck, M.A.; Dentinger, B.T.M.; Moncalvo, J.-M.; Neves, M.A.; da Silveira, R.M.B. Two new Tylopilus species (Boletaceae) from Northeastern Atlantic Forest, Brazil. Phytotaxa 2017, 316, 250–260. [Google Scholar] [CrossRef]
- Gelardi, M.; Angelini, C.; Costanzo, F.; Dovana, F.; Ortiz-Santana, B.; Vizzini, A. Tylopilus griseiolivaceus sp. nov. and T. leucomycelinus (Boletaceae) revisited from the Dominican Republic within a comprehensive phylogeny of Tylopilus s. str. Mycol. Prog. 2019, 18, 1039–1056. [Google Scholar] [CrossRef]
- Valões-Araújo, J.C.; Barbosa-Silva, A.; Koroiva, R.; Sá, M.C.; Roy, M.; Wartchow, F. Tylopilus dunensis (Boletaceae, Basidiomycota): Notes on morphological, phylogenetical and distributional aspects. Folia Cryptogam. Est. 2024, 61, 1–11. [Google Scholar] [CrossRef]
- Halling, R.E.; Osmundson, T.W. Tylopilus balloui Group in Australia. Projects Australian Tylopilus balloui Complex (Summary PDF). February 2023. Available online: https://sweetgum.nybg.org/science/projects/boletineae/wp-content/uploads/sites/6/2023/02/Tylopilus-balloui-Oz.pdf (accessed on 5 January 2024).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | Voucher Number | Genbank Accession No. | References | ||
|---|---|---|---|---|---|---|
| ITS | LSU | tef1α | ||||
| Boletus edulis | Germany | Be3 | – | KF030282 | – | [39] |
| Boletus sp. | China | HKAS 52639 | – | – | KF112195 | [37] |
| Chalciporus amarellus | Cyprus | ML901162CA | MH011876 | – | – | [40] |
| Chalciporus piperatus | Germany | HKAS 84882 | – | KT990562 | – | [41] |
| Chalciporus piperatus | USA | Smith-2018 | MK573906 | – | – | iNaturalist # 17338127 |
| Chalciporus rubinelloides | China | HKAS 5736 | – | KT990563 | – | [41] |
| Chiua viridula | China | HKAS 74928 (T) | – | NG_081278 (KF112483) | – | [37] |
| Cyanoboletus flavocontextus | China | BJTC FM2319-A (T) | OR655149 | OR655185 | OR659986 | [42] |
| Cyanoboletus flavocontextus | Vietnam | LE F-344051 (184VN22) | PP317924 | PP313111 | PP320320 | this work |
| Cyanoboletus flavocontextus | Vietnam | LE F-344052 (48HG23) | PP317923 | – | PP320319 | this work |
| Cyanoboletus mediterraneensis | Israel | K-M000265124 | OM801199 | – | – | [43] |
| Cyanoboletus pulverulentus | Italy | MG 628a | KT157055 | KT157064 | – | [44] |
| Cyanoboletus sinopulverulentus (as Boletus sp.) | China | HKAS 59609 | – | KF112366 | – | [37] |
| Cyanoboletus sinopulverulentus (as Boletus sinopulverulentus) | China | HKAS 59609 | – | – | KF112193 | [41] |
| Cyanoboletus sinopulve-rulentus (as Boletus sinopulverulentus) | China | HMAS 266894 (T) | KC579402 | – | – | [45] |
| Cyanoboletus sp. | China | OR0257 | – | – | MG212587 | [46] |
| Cyanoboletus sp. | China | HKAS 59418 | – | – | KT990765 | [41] |
| Fistulinella olivaceoalba | Vietnam | LE 312004 (T) | – | NG_068853 (MH718396) | – | [24] |
| Gyrodon lividus | Germany | REG Gl1 | – | – | GU187701 | [47] |
| Gyroporus castaneus (as Gyroporus sp.) | China | HKAS 63505 | – | KF112476 | – | [37] |
| Gyroporus castaneus | USA | JMP0028 | EU819468 | – | – | [48] |
| Gyroporus cyanescens | USA | MB 05-001 | – | EU718138 | FJ536672 | [49] |
| Gyroporus cyanescens | USA | NAMA190 | EU819495 | – | – | [48] |
| Harrya subalpina | China | HKAS 90194 | – | KT990700 | – | [41] |
| Hortiboletus amygdalinus | China | HKAS54166 (T) | – | NG_059622 (KT990581) | KT990777 | [41] |
| Hortiboletus amygdalinus | Korea | NIBRFG0000502792 | MW578955 | – | – | [50] |
| Hortiboletus arduinus | China | FHMU 3323 (T) | – | NG_075378 (MT646432) | – | [51] |
| Hortiboletus bubalinus | Russia | LE F-315839 | PP317928 | PP313106 | PP320315 | this work |
| Hortiboletus indorubellus | India | DC 14-002 (T) | NR_154076 (KT319647) | – | – | [52] |
| Hortiboletus kohistanensis (as Xerocomus sp.) | Pakistan | AST48 (LAH35327, T) | MG988192 | MG988187 | – | [53] |
| Hortiboletus napaeus | China | FHMU3326 (T) | NR_184923 (MT646440) | NG_075379 (MT646433) | – | [51] |
| Hortiboletus cf. rubellus (=Xerocomellus cf. rubellus) | USA | MB03–033 | – | KF030294 | KF030419 | [39] |
| Hortiboletus sp. | Germany | KR-M-0044799 | MT006029 | Genbank | ||
| Hortiboletus rubroreticulatus | Vietnam | LE F-344053 (T) | PP317926 | PP313108 | PP320317 | this work |
| Hortiboletus rubroreticulatus | Vietnam | LE F-344054 | PP317927 | PP313107 | PP320316 | this work |
| Hortiboletus rufosquamosus | China | BJTC FM2649 | – | – | OR659988 | [42] |
| Hortiboletus rupicapreus | Vietnam | LE 312677 (T) | MW784161 | MW760391 | MZ424894 | [29] |
| Hortiboletus rupicapreus | Vietnam | LE 312678 | MW784162 | – | MZ424893 | [29] |
| Hortiboletus rupicapreus | Vietnam | LE F-344055 (130VN22) | PP317925 | – | – | this work |
| Hortiboletus subpaludosus | China | HKAS68158 | - | - | KT990779 | [41] |
| Hortiboletus tomentosus | China | BJTC FM2289-A (T) | – | – | OR659994 | [42] |
| Kgaria cyanogranulifera | Australia | NY1115381 (REH9189) | – | OR063859 | OR232707 | [54] |
| Kgaria cyanogranulifera | Australia | NY1194100 (REH9359) | – | OR063862 | OR232708 | [54] |
| Kgaria similis | Australia | NY1193839 (REH9406) | – | OR063867 | OR113660 | [54] |
| Kgaria similis | Australia | NY1193974 (REH9031, T) | – | OR063865 | – | [54] |
| Kgaria virescens | Vietnam | LE F-344056 (138VN23) | PP317930 | PP313104 | PP320322 | this work |
| Kgaria virescens | Vietnam | LE F-315591 (261VN16) | – | – | PP320321 | this work |
| Lanmaoa asiatica | China | HKAS 63603 | – | – | KM605153 | [55] |
| Parvixerocomus pseudoaokii | China | HKAS 80480 (T) | – | NG_059543 (KP658468) | – | [55] |
| Parvixerocomus pseudoaokii | Thailand | CMU58-ST-0504 | KX017303 | – | – | Genbank |
| Parvixerocomus pseudoaokii | China | OR0155 | – | – | MG212597 | [46] |
| Parvixerocomus pseudoaokii | Vietnam | LE F-344057 (70HG23) | PP317937 | PP313110 | – | this work |
| Parvixerocomus matheranensis | India | AMH 9976 (T) | – | – | MT140890 | [56] |
| Phylloporus hainanensis | China | Zeng2724 (FHMU1718, T) | MW588632 | MW588581 | – | [57] |
| Phylloporus hainanensis | China | N.K. Zeng 4984 (FHMU5550) | – | – | ON868501 | [58] |
| Phylloporus hainanensis | Vietnam | LE F-344058 (128VN23) | PP317934 | PP313115 | PP320324 | this work |
| Phylloporus hainanensis | Vietnam | 137VN23 | PP317935 | – | – | this work |
| Phylloporus hainanensis | Vietnam | 108VN23 | PP317933 | – | – | this work |
| Phylloporus luxiensis | Vietnam | LE 315622 | – | – | MZ424877 | [9] |
| Phylloporus microsquamus | China | FHMU 1678 (T) | NR_175702 (MW588648) | NG_088206 (MW588599) | – | [57] |
| Phylloporus microsquamus | China | OR0258 | – | – | MH580809 | [59] |
| Phylloporus microsquamus | Vietnam | LE 312685 | MZ451354 | PP313113 | – | [9] as Phyllo-porus sp.) |
| Phylloporus microsquamus | Vietnam | LE 312684 | – | – | MZ424892 | [9] |
| Phylloporus nigrisquamus | Thailand | MAN131 | JQ003649 | – | – | [57] |
| Phylloporus nigrisquamus | China | Y.G. Fan 2819 (FHMU3271, T) | – | NG_088205 (MW588590) | – | [57] |
| Phylloporus nigrisquamus | Vietnam | LE F-344059 (172VN22) | PP317936 | PP313114 | PP320325 | this work |
| Phylloporus subbacillisporus | Thailand | OR0989 | MH686277 | – | – | [59] |
| Phylloporus subbacillisporus | China | OR0436 (HMAS 279879, T) | NR_173793 (MH686274) | MH580812 | [59] | |
| Phylloporus subbacillisporus | China | HKAS 74682 | – | JQ967230 | – | [60] |
| Phylloporus subbacillisporus | Vietnam | LE F-344060 (193VN22) | PP317932 | PP313116 | PP320326 | this work |
| Porphyrellus porphyrosporus | China | HKAS 49182 (Ge687) | – | KT990544 | – | [41,61] |
| Porphyrellus porphyrosporus (as Tylopilus porphyrosporus) | China | HKAS 76671 | – | – | KF112243 | [37] |
| Pseudoaustroboletus valens | China | HKAS 82644 (LF690) | – | MT154749 | – | [61] |
| Pulveroboletus flaviscabrosus | China | HKAS83190 (T) | KX453821 | [62] | ||
| Pulveroboletus subrufus | China | HKAS84926 | – | KX453836 | – | [62] |
| Retiboletus griseus | China | HKAS:63590 | – | KF112417 | – | [37] |
| Tylopilus aurantiacus | China | HKAS59700 (Li1952, T) | – | NG_081276 (KF112458) | KF112223 | [61] |
| Tylopilus aurantiovulpinus | Vietnam | LE F-344061 (T) | PP317938 | PP313109 | PP320323 | this work |
| Tylopilus aurantiovulpi-nus (as T. aff. balloui) | Vietnam | LE 312700 | MZ451358 | – | MZ424886 | [9] |
| Tylopilus balloui | USA | NY, Halling 8292 | – | EU430734 | – | [63] |
| Tylopilus balloui | Thailand | CMU51-SL-39 | KX017306 | – | – | Genbank |
| Tylopilus callainus | China | FHMU N.K.Zeng 1459 (T) | MG365901 | MG365896 | – | [64] |
| Tylopilus callainus | China | FHMU N.K.Zeng 1464 | – | – | MG365904 | [64] |
| Tylopilus felleus | Sweden | AT2001011 | UDB000680 | JQ326993 | JQ327015 | UNITE DB; [65] |
| Tylopilus leucomycelinus | Belize | NY00796119 | – | EU430735 | – | [63] |
| Tylopilus microsporus | China | HMAS 84730 (T) | NR_137924 (KM975485) | – | – | [44] |
| Tylopilus otsuensis | China | HKAS 53401 | – | KF112449 | – | [37] |
| Tylopilus pseudoballoui | India | DC 17-35 (T) | MG799324 | MG799325 | – | [66] |
| Tylopilus pseudoballoui (as T. balloui) | China | HKAS51151 (Li714) | – | – | MW165265 | [61] |
| Tylopilus pseudoleucomycelinus | Mexico | MEXU: HO 30115 | – | OQ940037 | – | [67] |
| Tylopilus rubrotinctus | China | HKAS 80684 (KK259, T) | – | NG_088129 (MT154733) | MW165264 | [61] |
| Tylopilus rubrotinctus (as T. aff. balloui) | Vietnam | LE 312532 | MZ451357 | – | MZ424883 | [9] |
| Veloporphyrellus vulpinus | Vietnam | LE 315544 (T) | – | MN511170 | – | [25] |
| Xanthoconium sinense | China | HKAS77758 (T) | – | NG_059630 (KT990665) | – | [41] |
| Xerocomellus bolinii | USA | JAB_95 | – | – | MW737491 | [68] |
| Xerocomellus chrysenteron | Spain | AH38968 | KU355473 | – | – | Genbank |
| Xerocomellus cisalpinus | Russia | LE F-343575 | PP317929 | – | PP320318 | this work |
| Xerocomellus cisalpinus | Finland | AT2005034 | – | – | KF030417 | [39] |
| Xerocomellus cisalpinus | Russia | LE 315834 | – | PP313105 | – | this work |
| Xerocomellus poederi | Spain | AH 44050 (T) | NR_155971 (KU355475) | NG_060000 (KU355488) | – | [69] |
| Xerocomellus sarnarii | Italy | MCVE 28577 (T) | NR_138006 (KT271749) | – | – | [70] |
| Xerocomus ferrugineus | Sweden | AT1999098 | DQ066398 | – | – | [71] |
| Xerocomus ferrugineus | China | BJTC FM1245 | – | OR655219 | OR660018 | [42] |
| Xerocomus magniporus | China | HKAS 59820 | – | – | JQ967195 | [60] |
| Xerocomus sp. | Vietnam | LE F-344062 (165VN22) | PP317931 | PP313112 | PP320327 | this work |
| Xerocomus squamulosus | New Zealand | PDD 101777 | OP141473 | – | – | Genbank |
| Xerocomus squamulosus | New Zealand | JAC10883 | – | OP141507 | – | Genbank |
| Xerocomus subparvus | Vietnam | LE315595 | MT893600 | – | – | [3] |
| Xerocomus subsplendidus | China | HFJAU12011 | – | – | OQ162214 | [72] |
| Xerocomus subtomentosus | Sweden | AT2002025a | DQ066361 | – | – | [71] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, T.H.G.; Popov, E.; Alexandrova, A.; Ivanova, D.; Morozova, O. New Data on Boletaceae (Agaricomycetes, Basidiomycota) from Central Vietnam with Description of Two New Species and Creation of a New Combination Based on Morphological and Phylogenetic Evidence. J. Fungi 2024, 10, 223. https://doi.org/10.3390/jof10030223
Pham THG, Popov E, Alexandrova A, Ivanova D, Morozova O. New Data on Boletaceae (Agaricomycetes, Basidiomycota) from Central Vietnam with Description of Two New Species and Creation of a New Combination Based on Morphological and Phylogenetic Evidence. Journal of Fungi. 2024; 10(3):223. https://doi.org/10.3390/jof10030223
Chicago/Turabian StylePham, Thi Ha Giang, Eugene Popov, Alina Alexandrova, Daria Ivanova, and Olga Morozova. 2024. "New Data on Boletaceae (Agaricomycetes, Basidiomycota) from Central Vietnam with Description of Two New Species and Creation of a New Combination Based on Morphological and Phylogenetic Evidence" Journal of Fungi 10, no. 3: 223. https://doi.org/10.3390/jof10030223