
Bovine Natural Antibody Relationships to Specific Antibodies and Fasciola hepatica Burdens after Experimental Infection and Vaccination with Glutathione S-Transferase


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals, Vaccination and Sampling
2.2. Acquisition of Parasite Whole-Worm Extract and Purification of Parasitic Glutathione S-Transferase
2.3. Indirect ELISA Analysis of Natural Antibodies and Specific Antibody Responses
2.4. Statistical Analysis
3. Results
3.1. Liver Fluke Burdens and Fluke Wet Weights in Control and Vaccinated Cattle
3.2. Optimisation of NAb Assays
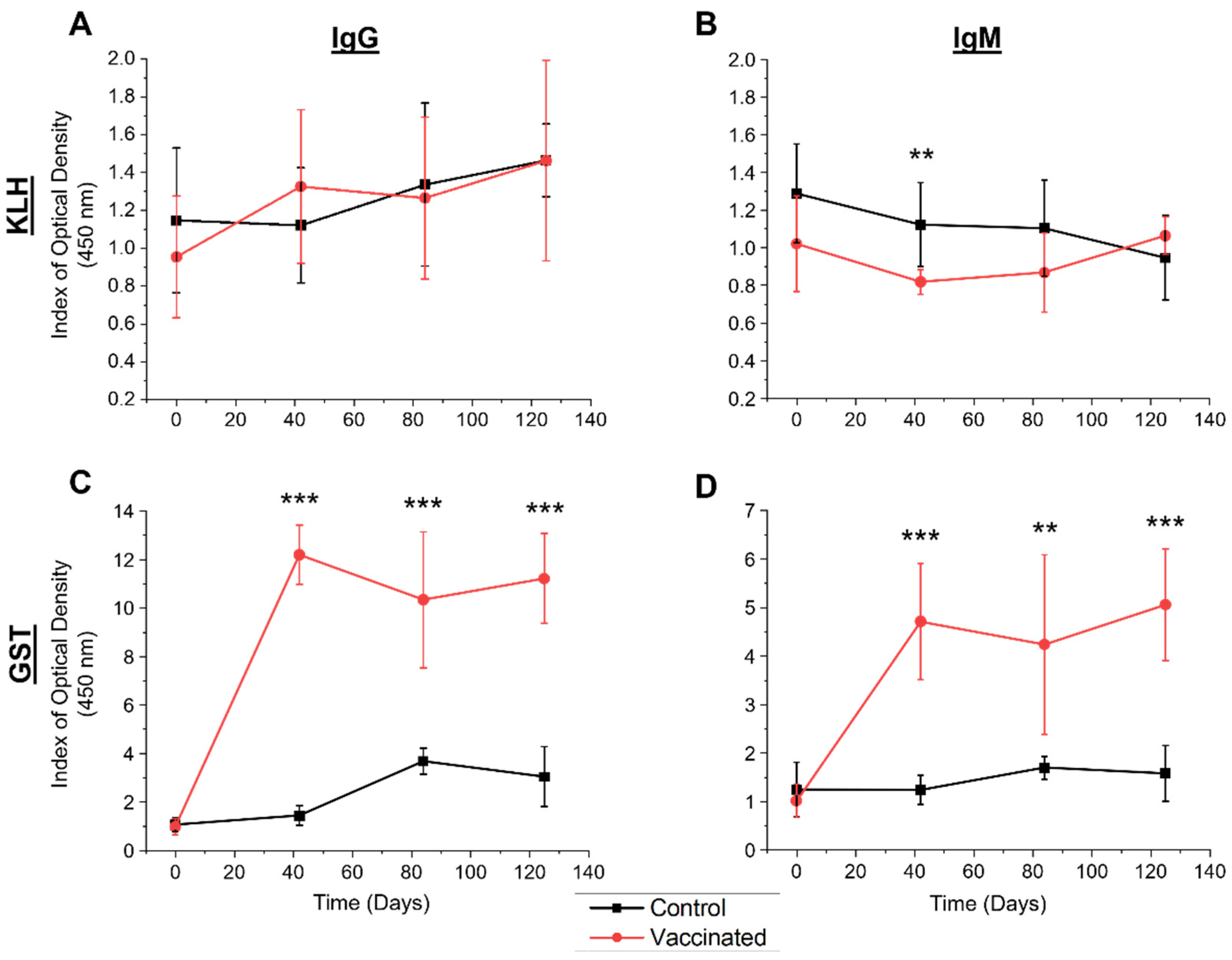
3.3. Fluctuations in Mean NAb and SpAb Levels throughout the Vaccination Trial
3.4. Relationships between Naïve NAb and Vaccine-Induced SpAb Production
3.5. Correlations between NAb and SpAb Levels to Final Liver Fluke Burdens and Wet Weights
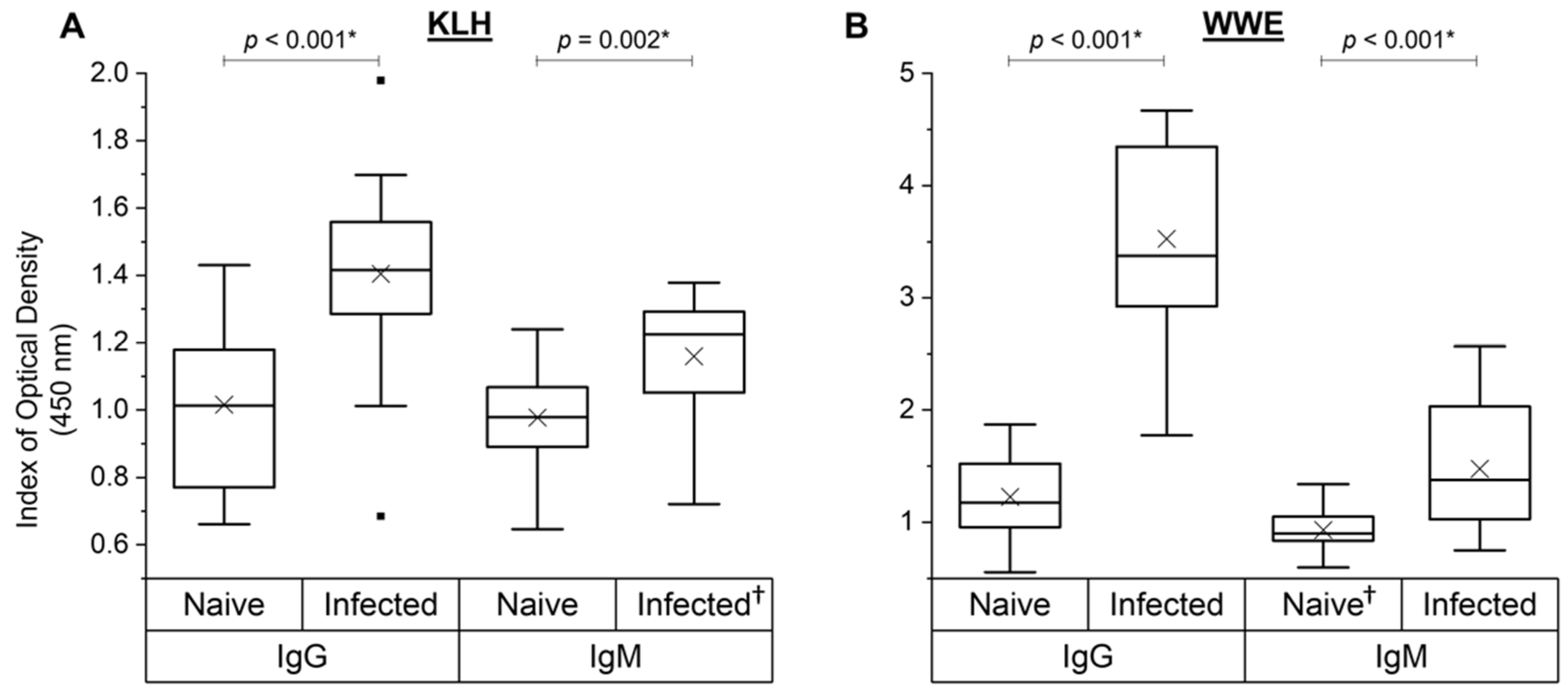
3.6. NAb and SpAb Levels to Fluke WWE before and after Experimental Infection
3.7. Correlations between Naïve NAbs and Specific WWE Production Post-Infection
3.8. Antibody Correlations to Final Liver Fluke Burdens and Wet Weights after Experimental Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruinsma, J. World Agriculture: Towards 2015/2030: An FAO Study; Earthscan: London, UK, 2017. [Google Scholar]
- Woodgate, R.; Cornell, A.; Sangster, N. Occurrence, measurement and clinical perspectives of drug resistance in important parasitic helminths of livestock. In Antimicrobial Drug Resistance; Springer: Cham, Switzerland, 2017; pp. 1305–1326. [Google Scholar]
- Mphahlele, M.; Molefe, N.; Tsotetsi-Khambule, A.; Oriel, T. Anthelmintic Resistance in Livestock. In Helminthiasis; IntechOpen: London, UK, 2019. [Google Scholar]
- Spithill, T.W. Fasciola gigantica: Epidemiology, control, immunology and molecular biology. In Fasciolosis; CABI: Wallingford, UK, 1999; pp. 465–525. [Google Scholar]
- Mehmood, K.; Zhang, H.; Sabir, A.J.; Abbas, R.Z.; Ijaz, M.; Durrani, A.Z.; Saleem, M.H.; Rehman, M.U.; Iqbal, M.K.; Wang, Y. A review on epidemiology, global prevalence and economical losses of fasciolosis in ruminants. Microb. Pathog. 2017, 109, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Kelley, J.M.; Elliott, T.P.; Beddoe, T.; Anderson, G.; Skuce, P.; Spithill, T.W. Current threat of triclabendazole resistance in Fasciola hepatica. Trends Parasitol. 2016, 32, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairweather, I.; Brennan, G.; Hanna, R.; Robinson, M.; Skuce, P. Drug resistance in liver flukes. Int. J. Parasitol. Drugs Drug Resist. 2020, 12, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Cervi, L.; Rossi, G.; Masih, D. Potential role for excretory–secretory forms of glutathione-S-transferase (GST) in Fasciola hepatica. Parasitology 1999, 119, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Piedrafita, D.; Estuningsih, E.; Pleasance, J.; Prowse, R.; Raadsma, H.W.; Meeusen, E.N.; Spithill, T.W. Peritoneal lavage cells of Indonesian thin-tail sheep mediate antibody-dependent superoxide radical cytotoxicity in vitro against newly excysted juvenile Fasciola gigantica but not juvenile Fasciola hepatica. Infect. Immun. 2007, 75, 1954–1963. [Google Scholar] [CrossRef] [Green Version]
- Piedrafita, D.; Parsons, J.C.; Sandeman, R.M.; Wood, P.; Estuningsih, S.; Partoutomo, S.; Spithill, T.W. Antibody-dependent cell-mediated cytotoxicity to newly excysted juvenile Fasciola hepatica in vitro is mediated by reactive nitrogen intermediates. Parasite Immunol. 2001, 23, 473–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brophy, P.; Patterson, L.; Brown, A.; Pritchard, D. Glutathione S-transferase (GST) expression in the human hookworm Necator americanus: Potential roles for excretory-secretory forms of GST. Acta Trop. 1995, 59, 259–263. [Google Scholar] [CrossRef]
- Toet, H.; Piedrafita, D.M.; Spithill, T.W. Liver fluke vaccines in ruminants: Strategies, progress and future opportunities. Int. J. Parasitol. 2014, 44, 915–927. [Google Scholar] [CrossRef]
- Piedrafita, D.; Spithill, T.W.; Dalton, J.P.; Brindley, P.J.; Sandeman, M.R.; Wood, P.R.; Parsons, J.C. Juvenile Fasciola hepatica are resistant to killing In Vitro by free radicals compared with larvae of Schistosoma mansoni. Parasite Immunol. 2000, 22, 287–295. [Google Scholar] [CrossRef]
- Morrison, C.A.; Colin, T.; Sexton, J.L.; Bowen, F.; Wicker, J.; Friedel, T.; Spithill, T.W. Protection of cattle against Fasciola hepatica infection by vaccination with glutathione S-transferase. Vaccine 1996, 14, 1603–1612. [Google Scholar] [CrossRef]
- Spithill, T.W.; Toet, H.; Rathinasamy, V.; Zerna, G.; Swan, J.; Cameron, T.; Smooker, P.M.; Piedrafita, D.M.; Dempster, R.; Beddoe, T. 12 Vaccines for Fasciola: New Thinking. In Fasciolosis, 2nd ed.; CAB International: Wallingford, UK, 2021. [Google Scholar]
- Baumgarth, N.; Tung, J.W.; Herzenberg, L.A. Inherent specificities in natural antibodies: A key to immune defense against pathogen invasion. In Springer Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2005; Volume 26, pp. 347–362. [Google Scholar]
- Panda, S.; Ding, J.L. Natural antibodies bridge innate and adaptive immunity. J. Immunol. 2015, 194, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boes, M. Role of natural and immune IgM antibodies in immune responses. Mol. Immunol. 2000, 37, 1141–1149. [Google Scholar] [CrossRef]
- Holodick, N.E.; Rodríguez-Zhurbenko, N.; Hernández, A.M. Defining natural antibodies. Front. Immunol. 2017, 8, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddur, M.S.; Lacroix-Desmazes, S.; Dimitrov, J.D.; Kazatchkine, M.D.; Bayry, J.; Kaveri, S.V. Natural Antibodies: From First-Line Defense Against Pathogens to Perpetual Immune Homeostasis. Clin. Rev. Allergy Immunol. 2020, 58, 213–228. [Google Scholar] [CrossRef]
- Hamilton, A.M.; Lehuen, A.; Kearney, J.F. Immunofluorescence analysis of B-1 cell ontogeny in the mouse. Int. Immunol. 1994, 6, 355–361. [Google Scholar] [CrossRef]
- Hardy, R.R.; Hayakawa, K. CD5 B cells, a fetal B cell lineage. In Advances in Immunology; Elsevier: Amsterdam, The Netherlands, 1993; Volume 55, pp. 297–339. [Google Scholar]
- Baumgarth, N.; Herman, O.C.; Jager, G.C.; Brown, L.E.; Herzenberg, L.A.; Chen, J. B-1 and B-2 cell–derived immunoglobulin M antibodies are nonredundant components of the protective response to influenza virus infection. J. Exp. Med. 2000, 192, 271–280. [Google Scholar] [CrossRef]
- Berghof, T.; Matthijs, M.; Arts, J.; Bovenhuis, H.; Dwars, R.; Van Der Poel, J.; Visker, M.; Parmentier, H. Selective breeding for high natural antibody level increases resistance to avian pathogenic Escherichia coli (APEC) in chickens. Dev. Comp. Immunol. 2019, 93, 45–57. [Google Scholar] [CrossRef]
- Sun, Y.; Parmentier, H.; Frankena, K.; Van Der Poel, J. Natural antibody isotypes as predictors of survival in laying hens. Poult. Sci. 2011, 90, 2263–2274. [Google Scholar] [CrossRef]
- Berghof, T.; Arts, J.; Bovenhuis, H.; Lammers, A.; Van Der Poel, J.; Parmentier, H. Antigen-dependent effects of divergent selective breeding based on natural antibodies on specific humoral immune responses in chickens. Vaccine 2018, 36, 1444–1452. [Google Scholar] [CrossRef]
- Thompson-Crispi, K.; Miglior, F.; Mallard, B. Genetic parameters for natural antibodies and associations with specific antibody and mastitis in Canadian Holsteins. J. Dairy Sci. 2013, 96, 3965–3972. [Google Scholar] [CrossRef] [Green Version]
- Van Altena, S.; Peen, M.; Van der Linden, F.; Parmentier, H.; Savelkoul, H.; Tijhaar, E. Bovine natural antibodies in antibody-dependent bactericidal activity against Escherichia coli and Salmonella Typhimurium and risk of mastitis. Vet. Immunol. Immunopathol. 2016, 171, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploegaert, T.; Tijhaar, E.; Lam, T.; Taverne-Thiele, A.; Van der Poel, J.; Van Arendonk, J.; Savelkoul, H.; Parmentier, H. Natural antibodies in bovine milk and blood plasma: Variability among cows, repeatability within cows, and relation between milk and plasma titers. Vet. Immunol. Immunopathol. 2011, 144, 88–94. [Google Scholar] [CrossRef] [PubMed]
- De Klerk, B.; Ducro, B.; Heuven, H.; Den Uyl, I.; Van Arendonk, J.; Parmentier, H.; Van der Poel, J. Phenotypic and genetic relationships of bovine natural antibodies binding keyhole limpet hemocyanin in plasma and milk. J. Dairy Sci. 2015, 98, 2746–2752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Knegsel, A.; de Vries Reilingh, G.; Meulenberg, S.; Van den Brand, H.; Dijkstra, J.; Kemp, B.; Parmentier, H. Natural antibodies related to energy balance in early lactation dairy cows. J. Dairy Sci. 2007, 90, 5490–5498. [Google Scholar] [CrossRef] [PubMed]
- Ochsenbein, A.F.; Zinkernagel, R.M. Natural antibodies and complement link innate and acquired immunity. Immunol. Today 2000, 21, 624–630. [Google Scholar] [CrossRef]
- Harris, J.; Markl, J. Keyhole limpet hemocyanin (KLH): A biomedical review. Micron 1999, 30, 597–623. [Google Scholar] [CrossRef]
- Brockwell, Y.; Spithill, T.; Anderson, G.; Grillo, V.; Sangster, N. Comparative kinetics of serological and coproantigen ELISA and faecal egg count in cattle experimentally infected with Fasciola hepatica and following treatment with triclabendazole. Vet. Parasitol. 2013, 196, 417–426. [Google Scholar] [CrossRef]
- McCusker, P.; Toet, H.; Rathinasamy, V.; Young, N.; Beddoe, T.; Anderson, G.; Dempster, R.; McVeigh, P.; McCammick, E.; Wells, D. Molecular characterisation and vaccine efficacy of two novel developmentally regulated surface tegument proteins of Fasciola hepatica. Vet. Parasitol. 2020, 286, 109244. [Google Scholar] [CrossRef]
- Zerna, G.; Rathinasamy, V.A.; Toet, H.; Anderson, G.; Dempster, R.; Spithill, T.W.; Beddoe, T. Evaluation of Immunogenicity and Efficacy of Fasciola hepatica Tetraspanin 2 (TSP2) Fused to E. coli Heat-Labile Enterotoxin B Subunit LTB Adjuvant Following Intranasal Vaccination of Cattle. Vaccines 2021, 9, 1213. [Google Scholar] [CrossRef]
- Swan, J.; Sakthivel, D.; Cameron, T.C.; Faou, P.; Downs, R.; Rajapaksha, H.; Piedrafita, D.; Beddoe, T. Proteomic identification of galectin-11 and-14 ligands from Fasciola hepatica. Int. J. Parasitol. 2019, 49, 921–932. [Google Scholar] [CrossRef]
- Wijffels, G.L.; Sexton, J.L.; Salvatore, L.; Pettitt, J.M.; Humphris, D.C.; Panaccio, M.; Spithill, T.W. Primary sequence heterogeneity and tissue expression of glutathione S-transferases of Fasciola hepatica. Exp. Parasitol. 1992, 74, 87–99. [Google Scholar] [CrossRef]
- Molina-Hernández, V.; Mulcahy, G.; Pérez, J.; Martínez-Moreno, Á.; Donnelly, S.; O’Neill, S.M.; Dalton, J.P.; Cwiklinski, K. Fasciola hepatica vaccine: We may not be there yet but we’re on the right road. Vet. Parasitol. 2015, 208, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyneveld, G.I.; Savelkoul, H.F.; Parmentier, H.K. Current understanding of natural antibodies and exploring the possibilities of modulation using veterinary models. A review. Front. Immunol. 2020, 11, 2139. [Google Scholar] [CrossRef] [PubMed]
- de Klerk, B.; Emam, M.; Thompson-Crispi, K.A.; Sargolzaei, M.; van der Poel, J.J.; Mallard, B.A. A genome-wide association study for natural antibodies measured in blood of Canadian Holstein cows. BMC Genom. 2018, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ploegaert, T.; Wijga, S.; Tijhaar, E.; Van Der Poel, J.; Lam, T.; Savelkoul, H.; Parmentier, H.; Van Arendonk, J. Genetic variation of natural antibodies in milk of Dutch Holstein-Friesian cows. J. Dairy Sci. 2010, 93, 5467–5473. [Google Scholar] [CrossRef] [Green Version]
- Wijga, S.; Bovenhuis, H.; Bastiaansen, J.; Van Arendonk, J.; Ploegaert, T.; Tijhaar, E.; Van Der Poel, J. Genetic parameters for natural antibody isotype titers in milk of Dutch Holstein-Friesians. Anim. Genet. 2013, 44, 485–492. [Google Scholar] [CrossRef]
- Wijga, S.; Parmentier, H.; Nieuwland, M.; Bovenhuis, H. Genetic parameters for levels of natural antibodies in chicken lines divergently selected for specific antibody response. Poult. Sci. 2009, 88, 1805–1810. [Google Scholar] [CrossRef]
- Parmentier, H.K.; Lammers, A.; Hoekman, J.J.; Reilingh, G.D.V.; Zaanen, I.T.; Savelkoul, H.F. Different levels of natural antibodies in chickens divergently selected for specific antibody responses. Dev. Comp. Immunol. 2004, 28, 39–49. [Google Scholar] [CrossRef]
- Prada Jiménez de Cisneros, J.; Stear, M.J.; Mair, C.; Singleton, D.; Stefan, T.; Stear, A.; Marion, G.; Matthews, L. An explicit immunogenetic model of gastrointestinal nematode infection in sheep. J. R. Soc. Interface 2014, 11, 20140416. [Google Scholar] [CrossRef] [Green Version]
- Stills, H.F., Jr. Adjuvants and antibody production: Dispelling the myths associated with Freund’s complete and other adjuvants. ILAR J. 2005, 46, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Dalton, J.P.; Robinson, M.W.; Mulcahy, G.; O’Neill, S.M.; Donnelly, S. Immunomodulatory molecules of Fasciola hepatica: Candidates for both vaccine and immunotherapeutic development. Vet. Parasitol. 2013, 195, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Spickler, A.R.; Roth, J.A. Adjuvants in veterinary vaccines: Modes of action and adverse effects. J. Vet. Intern. 2003, 17, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, S.; Rufrano, D.; Caputo, A. Natural antibodies and their relationship with total immunoglobulins and acquired antibody response in goat kid (Capra hircus, L. 1758) serum. Vet. Immunol. Immunopathol. 2019, 211, 38–43. [Google Scholar] [CrossRef]
- Sandmeier, F.C.; Tracy, C.R.; Dupré, S.; Hunter, K. A trade-off between natural and acquired antibody production in a reptile: Implications for long-term resistance to disease. Biol. Open 2012, 1, 1078–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seebach, J.D.; Yamada, K.; McMorrow, I.M.; Sachs, D.H.; DerSimonian, H. Xenogeneic human anti-pig cytotoxicity mediated by activated natural killer cells. Xenotransplantation 1996, 3, 188–197. [Google Scholar] [CrossRef]
- Sulaiman, A.A.; Zolnierczyk, K.; Japa, O.; Owen, J.P.; Maddison, B.C.; Emes, R.D.; Hodgkinson, J.E.; Gough, K.C.; Flynn, R.J. A trematode parasite derived growth factor binds and exerts influences on host immune functions via host cytokine receptor complexes. PLoS Path. 2016, 12, e1005991. [Google Scholar] [CrossRef] [Green Version]
- Piedrafita, D.; Raadsma, H.; Prowse, R.; Spithill, T.W. Immunology of the host–parasite relationship in fasciolosis (Fasciola hepatica and Fasciola gigantica). Can. J. Zool. 2004, 82, 233–250. [Google Scholar] [CrossRef]
- Ravidà, A.; Aldridge, A.M.; Driessen, N.N.; Heus, F.A.; Hokke, C.H.; O’Neill, S.M. Fasciola hepatica surface coat glycoproteins contain mannosylated and phosphorylated N-glycans and exhibit immune modulatory properties independent of the mannose receptor. PLoS Negl. Trop. Dis. 2016, 10, e0004601. [Google Scholar] [CrossRef] [Green Version]
- Ravidà, A.; Cwiklinski, K.; Aldridge, A.M.; Clarke, P.; Thompson, R.; Gerlach, J.Q.; Kilcoyne, M.; Hokke, C.H.; Dalton, J.P.; O’Neill, S.M. Fasciola hepatica surface tegument: Glycoproteins at the interface of parasite and host. Mol. Cell Proteom. 2016, 15, 3139–3153. [Google Scholar] [CrossRef] [Green Version]
- New, J.S.; King, R.G.; Kearney, J.F. Glycan Reactive Natural Antibodies and Viral Immunity. Viral Immunol. 2020, 33, 266–276. [Google Scholar] [CrossRef]
- Chauvin, A.; Boulard, C. Local immune response to experimental Fasciola hepatica infection in sheep. Parasite 1996, 3, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frigerio, S.; Da Costa, V.; Costa, M.; Festari, M.F.; Landeira, M.; Rodríguez-Zraquia, S.A.; Härtel, S.; Toledo, J.; Freire, T. Eosinophils Control Liver Damage by Modulating Immune Responses against Fasciola hepatica. Front. Immunol. 2020, 11, 2276. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IgG | IgM | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| KLH | GST | KLH | GST | |||||||||||||
| Day: | 0 | 42 | 84 | 125 | 0 | 42 | 84 | 125 | 0 | 42 | 84 | 125 | 0 | 42 | 84 | 125 |
| Vaccinated | ||||||||||||||||
| Fluke Burden | 0.04 (0.94) | −0.44 (0.39) | −0.20 (0.70) | −0.05 (0.93) | 0.28 (0.59) | −0.77 (0.07) | −0.78 (0.07) | −0.77 (0.07) | −0.46 (0.36) | −0.18 (0.73) | 0.24 (0.64) | −0.41 (0.42) | 0.06 (0.91) | −0.75 (0.08) | −0.66 (0.15) | −0.69 (0.13) |
| Fluke Weight | −0.03 (0.95) | −0.35 (0.50) | −0.15 (0.78) | 0.06 (0.91) | 0.20 (0.70) | −0.83 * (0.04) | −0.82 * (0.04) | −0.81 * (0.04) | −0.60 (0.22) | −0.24 (0.65) | 0.10 (0.86) | −0.30 (0.56) | −0.04 (0.94) | −0.85 * (0.03) | −0.75 (0.08) | −0.73 (0.10) |
| Control | ||||||||||||||||
| Fluke Burden | −0.29 (0.57) | −0.42 (0.41) | −0.42 (0.41) | −0.39 (0.44) | −0.23 (0.66) | 0.56 (0.25) | 0.04 (0.94) | −0.55 (0.26) | −0.36 (0.48) | −0.39 (0.46) | −0.76 (0.08) | −0.35 (0.49) | −0.31 (0.55) | 0.19 (0.73) | −0.49 (0.32) | −0.37 (0.47) |
| Fluke Weight | −0.19 (0.71) | −0.46 (0.36) | −0.44 (0.38) | −0.12 (0.82) | −0.24 (0.65) | 0.29 (0.57) | −0.08 (0.88) | −0.57 (0.24) | −0.28 (0.59) | −0.20 (0.70) | −0.48 (0.33) | −0.20 (0.70) | −0.19 (0.72) | 0.45 (0.37) | −0.26 (0.61) | −0.23 (0.66) |
| IgG | IgM | |||||||
|---|---|---|---|---|---|---|---|---|
| KLH | WWE | KLH | WWE | |||||
| Naïve | Infected | Naïve | Infected | Naïve | Infected † | Naïve † | Infected | |
| Fluke Burden | −0.08 (0.74) | 0.28 (0.24) | −0.05 (0.85) | −0.19 (0.44) | 0.10 (0.69) | 0.43 (0.07) | 0.56 * (0.02) | 0.29 (0.23) |
| Fluke Weight | −0.16 (0.51) | 0.21 (0.34) | 0.01 (0.97) | −0.09 (0.70) | 0.07 (0.77) | 0.123 (0.35) | 0.25 (0.32) | 0.118 (0.44) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zerna, G.; Cameron, T.C.; Toet, H.; Spithill, T.W.; Beddoe, T. Bovine Natural Antibody Relationships to Specific Antibodies and Fasciola hepatica Burdens after Experimental Infection and Vaccination with Glutathione S-Transferase. Vet. Sci. 2022, 9, 58. https://doi.org/10.3390/vetsci9020058
Zerna G, Cameron TC, Toet H, Spithill TW, Beddoe T. Bovine Natural Antibody Relationships to Specific Antibodies and Fasciola hepatica Burdens after Experimental Infection and Vaccination with Glutathione S-Transferase. Veterinary Sciences. 2022; 9(2):58. https://doi.org/10.3390/vetsci9020058
Chicago/Turabian StyleZerna, Gemma, Timothy C. Cameron, Hayley Toet, Terry W. Spithill, and Travis Beddoe. 2022. "Bovine Natural Antibody Relationships to Specific Antibodies and Fasciola hepatica Burdens after Experimental Infection and Vaccination with Glutathione S-Transferase" Veterinary Sciences 9, no. 2: 58. https://doi.org/10.3390/vetsci9020058