
Short Road Transport and Slaughter Stress Affects the Expression Profile of Serotonin Receptors, Adrenocortical, and Hematochemical Responses in Horses
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals and Blood Sample Collection
2.3. Transport Vehicle and Environmental Conditions
2.4. Cortisol and 5-HT Analyses
2.5. Peripheral Blood Mononuclear Cells’ (PBMC) Isolation, RNA Extraction and Reverse Transcription
2.6. Real-Time Quantitative PCR (qPCR)
2.7. Analysis of Hematochemical Parameters
2.8. Statistical Analysis
3. Results
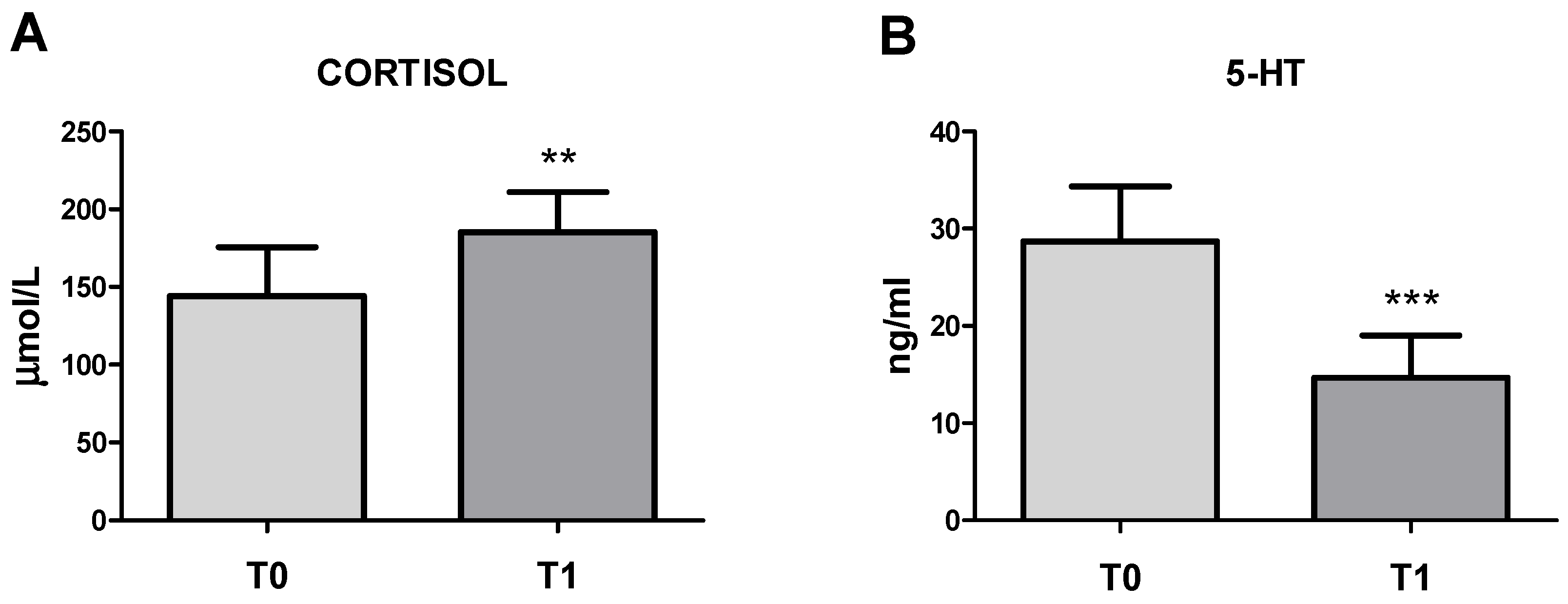
3.1. Circulating Cortisol and 5-HT Concentrations
3.2. 5-HT1B and 5-HT2A Serotoninergic Receptors’ Expression and IL-12 Expression
3.3. Hematochemical Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moberg, G.P. Biological response to stress: Implication for animal welfare. In The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; Moberg, G.P., Mench, J.A., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 1–21. [Google Scholar]
- Padalino, B.; Raidal, S.L.; Carter, N.; Celi, P.; Muscatello, G.; Jeffcott, L.; de Silva, K. Immunological, clinical, haematological and oxidative responses to long distance transportation in horses. Res. Vet. Sci. 2017, 115, 78–87. [Google Scholar] [CrossRef]
- Felici, M.; Nanni Costa, L.; Zappaterra, M.; Bozzo, G.; Di Pinto, P.; Minero, M.; Padalino, B. Journeys, Journey Conditions, and Welfare Assessment of Broken (Handled) Horses on Arrival at Italian Slaughterhouses. Animals 2022, 12, 3122. [Google Scholar] [CrossRef]
- Argüelles, J.; Echaniz, M.; Bowen, J.; Fatjó, J. The impact of a stress-reducing protocol on the quality of pre-anaesthesia in cats. Vet. Rec. 2021, 188, e138. [Google Scholar] [CrossRef]
- Placci, M.; Marliani, G.; Sabioni, S.; Gabai, G.; Mondo, E.; Borghetti, P.; De Angelis, E.; Accorsi, P.A. Natural Horse Boarding Vs Traditional Stable: A Comparison of Hormonal, Hematological and Immunological Parameters. J. Appl. Anim. Welf. Sci. 2020, 23, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Fazio, E.; Cravana, C.; Medica, P.; Ferlazzo, A. Cortisol response to road transport stress in calm and nervous stallions. J. Vet. Behav. 2013, 8, 231–237. [Google Scholar] [CrossRef]
- Marrero, M.G.; Dado-Senn, B.; Field, S.L.; Yang, G.; John, P.; Driver, J.P.; Laporta, J. Chronic heat stress delays immune system development and alters serotonin signaling in pre-weaned dairy calves. PLoS ONE 2021, 16, e0252474. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, Q.; Lu, F.; Xu, H.; Wang, P.; Feng, Y. Prognostic and immunomodulatory roles of schizophrenia-associated genes HTR2A, COMT, and PRODH in pan-cancer analysis and glioma survival prediction model. Front. Immunol. 2023, 14, 1201252. [Google Scholar] [CrossRef] [PubMed]
- Westergaard, G.C.; Suomi, S.J.; Chavanne, T.J.; Houser, L.; Hurley, A.; Cleveland, A.; Snoy, P.J.; Higley, J.D. Physiological Correlates of Aggression and Impulsivity in Free-Ranging Female Primates. Neuropsychopharmacology 2003, 28, 1045–1055. [Google Scholar] [CrossRef]
- Hunter, J.N.; Wood, E.K.; Roberg, B.L.; Neville, L.; Schwandt, M.L.; Fairbanks, L.A.; Barr, C.; Lindell, S.G.; Goldman, D.; Suomi, S.J.; et al. Mismatches in resident and stranger serotonin transporter genotypes lead to escalated aggression, and the target for aggression is mediated by sex differences in male and female rhesus monkeys (Macaca mulatta). Horm. Behav. 2022, 140, 105104. [Google Scholar] [CrossRef] [PubMed]
- Gotlib, I.H.; Joormann, J.; Minor, K.L.; Hallmayer, J. HPA Axis Reactivity: A Mechanism Underlying the Associations Among 5-HTTLPR, Stress, and Depression. Biol. Psychiatry 2008, 63, 847–851. [Google Scholar] [CrossRef]
- Bush, V.L.; Middlemiss, D.N.; Marsden, C.A.; Fone, K.C.F. Implantation of a Slow Release Corticosterone Pellet Induces Long-Term Alterations in Serotoninergic Neurochemistry in the Rat Brain. J. Neuronendocrinol. 2003, 15, 607–613. [Google Scholar] [CrossRef]
- Lucki, I. The Spectrum of Behaviors Influenced by Serotonin. Biol. Psychiatry 1998, 44, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Chaouloff, F.; Berton, O.; Mormede, P. Serotonin and Stress. Neuropsychopharmacology 1999, 21 (Suppl. S2), S28–S32. [Google Scholar] [CrossRef]
- Rouzaud Laborde, C.; Delmas, C.; Mialet-Perez, J.; Pizzinat, N.; Biendel-Picquet, C.; Boudou, N.; Dumonteil, N.; Spreux-Varoquaux, O.; Carrié, D.; Galinier, M.; et al. First Evidence of Increased Plasma Serotonin Levels in Tako-Tsubo Cardiomyopathy. Biomed. Res. Int. 2013, 2013, 847069. [Google Scholar] [CrossRef] [PubMed]
- Ayala, I.; Martos, N.F.; Silvan, G.; Gutierrez-Panizo, C.; Clavel, J.G.; Illera, J.C. Cortisol, adrenocorticotropic hormone, serotonin, adrenaline and noradrenaline serum concentrations in relation to disease and stress in the horse. Res. Vet. Sci. 2012, 93, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Bruschetta, G.; Di Pietro, P.; Fazio, E.; Ferlazzo, A.M. Plasma serotonin, tryptophan, hematological, and functional responses to horse trekking. J. Vet. Behav. 2014, 9, 248–253. [Google Scholar] [CrossRef]
- Bruschetta, G.; Medica, P.; Fazio, E.; Cravana, C.; Ferlazzo, A.M. The effect of training sessions and feeding regimes on neuromodulator role of serotonin, tryptophan and β-endorphin of horses. J. Vet. Behav. 2018, 23, 82–86. [Google Scholar] [CrossRef]
- Bruschetta, G.; D’Ascola, A.; Medica, P.; Ferlazzo, A.M. Physical Exercise Affects Serotoninergic System in Horse Leukocytes. J. Equine Vet. Sci. 2020, 88, 102969. [Google Scholar] [CrossRef] [PubMed]
- Alberghina, D.; Giannetto, C.; Piccione, G. Peripheral serotoninergic response to physical exercise in athletic horses. J. Vet. Sci. 2010, 11, 285–289. [Google Scholar] [CrossRef]
- Medica, P.; Giunta, R.P.; Bruschetta, G.; Ferlazzo, A.M. The Influence of Training and Simulated Race on Horse Plasma. J. Equine Vet. Sci. 2020, 84, 102818. [Google Scholar] [CrossRef]
- Chaouloff, F. Physiopharmacological interactions between stress hormones and central serotonergic systems. Brain Res. Brain Res. Rev. 1993, 18, 1–32. [Google Scholar] [CrossRef]
- Laaris, N.; Le Poul, E.; Hamon, M.; Lanfumey, L. Stress-induced alterations of somatodendritic 5-HT1A autoreceptor sensitivity in the rat dorsal raphe nucleus—In vitro electrophysiological evidence. Fundam. Clin. Pharmacol. 1997, 11, 206–214. [Google Scholar] [CrossRef]
- Raghupathi, R.K.; McGonigle, P. Differential Effects of Three Acute Stressors on the Serotonin 5-HT1A Receptor System in Rat Brain. Neuroendocrinology 1997, 65, 246–258. [Google Scholar] [CrossRef]
- Herr, N.; Bode, C.; Duerschmied, D. The Effects of Serotonin in Immune Cells. Front. Cardiovasc. Med. 2017, 4, 48. [Google Scholar] [CrossRef] [PubMed]
- Da Prada, M.; Cesura, A.M.; Launay, J.M.; Richards, J.G. Platelets as a model for neurones? Experientia 1988, 44, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Abeni, F.; Dal Prà, A.; Bertin, G.; Calamari, L. Serum Protein Fraction in Mature Horses and Relationship With Metabolic and Hematological Parameters. J. Equine Vet. Sci. 2013, 33, 905–911. [Google Scholar] [CrossRef]
- Noleto, P.G.; dos Santos, J.B.; Rocha, F.M.; Fasano, P.E.; Guimarães, E.C.; Mundim, A.V. Effect of a 130-km Endurance Ride on the Serum Biochemical Profile of Mangalarga Marchador Horses. J. Equine Vet. Sci. 2016, 39, 7–11. [Google Scholar] [CrossRef]
- Medica, P.; Cravana, C.; Bruschetta, G.; Ferlazzo, A.; Fazio, E. Breeding season and transport interactions on the pituitary-adrenocortical and biochemical responses of horses. J. Vet. Behav. 2018, 23, 91–96. [Google Scholar] [CrossRef]
- Medica, P.; Cravana, C.; Bruschetta, G.; Ferlazzo, A.; Fazio, E. Physiological and behavioral patterns of normal-term thoroughbred foals. J. Vet. Behav. 2018, 26, 38–42. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Cravana, C.; Aveni, F.; Ferlazzo, A. Comparative endocrinological responses to short transportation of Equidae (Equus asinus and Equus caballus). Anim. Sci. J. 2013, 84, 258–263. [Google Scholar] [CrossRef]
- Colborn, D.R.; Thompson, D.L., Jr.; Roth, T.L.; Capehart, J.S.; White, K.L. Responses of cortisol and prolactin to sexual excitement and stress in stallions and geldings. J. Anim. Sci. 1991, 69, 2556–2562. [Google Scholar] [CrossRef] [PubMed]
- Villani, M.; Cairoli, F.; Kindahl, H.; Galeati, G.; Faustini, M.; Carluccio, A.; Veronesi, M.C. Effects of Mating on Plasma Concentrations of Testosterone, Cortisol, Oestrone Sulphate and 15-Ketodihydro-PGF2α in Stallions. Reprod. Domest. Anim. 2006, 41, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Sardi, L.; Gastaldo, A.; Borciani, M.; Bertolini, A.; Musi, V.; Martelli, G.; Cavallini, D.; Rubini, G.; Nannoni, E. Identification of Possible Pre-Slaughter Indicators to Predict Stress and Meat Quality: A Study on Heavy Pigs. Animals 2020, 10, 945. [Google Scholar] [CrossRef] [PubMed]
- Sardi, L.; Gastaldo, A.; Borciani, M.; Bertolini, A.; Musi, V.; Garavaldi, A.; Martelli, G.; Cavallini, D.; Nannoni, E. Pre-Slaughter Sources of Fresh Meat Quality Variation: The Case of Heavy Pigs Intended for Protected Designation of Origin Products. Animals 2020, 10, 2386. [Google Scholar] [CrossRef] [PubMed]
- Sommavilla, R.; Faucitano, L.; Gonyou, H.; Seddon, Y.; Bergeron, R.; Widowski, T.; Crowe, T.; Connor, L.; Scheeren, B.M.; Goumon, S.; et al. Season, Transport Duration and Trailer Compartment Effects on Blood Stress Indicators in Pigs: Relationship to Environmental, Behavioral and Other Physiological Factors, and Pork Quality Traits. Animals 2017, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Schwartzkopf-Genswein, K.S.; Faucitano, L.; Dadgar, S.; Shand, P.; González, L.A.; Crowe, T.G. Road transport of cattle, swine and poultry in North America and its impact on animal welfare, carcass and meat quality: A review. Meat Sci. 2012, 92, 227–243. [Google Scholar] [CrossRef]
- Arroyo, L.; Valent, D.; Carreras, R.; Peña, R.; Sabrià, J.; Velarde, A.; Bassols, A. Housing and road transport modify the brain neurotransmitter systems of pigs: Do pigs raised in different conditions cope differently with unknown environments? PLoS ONE 2019, 14, e0210406. [Google Scholar] [CrossRef]
- Edwards, S.A. Product quality attributes associated with outdoor pig production. Livest. Prod. Sci. 2005, 94, 5–14. [Google Scholar] [CrossRef]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Clinical Biochemistry of Domestic Animals, 6th ed.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2008; pp. 1–25. [Google Scholar]
- Hart, K.A.; Wochele, D.M.; Norton, N.A.; McFarlane, D.; Wooldridge, A.A.; Frank, N. Effect of Age, Season, Body Condition, and Endocrine Status on Serum Free Cortisol Fraction and Insulin Concentration in Horses. J. Vet. Intern. Med. 2016, 30, 653–663. [Google Scholar] [CrossRef]
- Kim, J.; Yoon, M. The effect of serotonin and oxytocin on equine docility and friendliness to humans. J. Vet. Behav. 2022, 50, 18–22. [Google Scholar] [CrossRef]
- Lu, Q.; Hofferbert, B.V.; Koo, G.; Malinauskas, R.A. In Vitro Shear Stress-Induced Platelet Activation: Sensitivity of Human and Bovine Blood. Artif. Organs 2013, 37, 894–903. [Google Scholar] [CrossRef]
- Kanova, M.; Kohout, P. Serotonin—Its Synthesis and Roles in the Healthy and the Critically Ill. Int. J. Mol. Sci. 2021, 22, 4837. [Google Scholar] [CrossRef] [PubMed]
- Beltrand, P.P.; Beltrand, R.L. Serotonin release and uptake in the gastrointestinal tract. Auton. Neurosci. 2010, 153, 47–57. [Google Scholar] [CrossRef]
- Mohammad-Zadeh, L.F.; Moses, L.; Gwaltney-Brant, S.M. Serotonin: A review. J. Vet. Pharmacol. Therap. 2008, 31, 187–199. [Google Scholar] [CrossRef]
- León-Ponte, M.; Ahern, G.P.; O’Connell, P.J. Serotonin provides an accessory signal to enhance T-cell activation by signaling through the 5-HT7 receptor. Blood 2007, 109, 3139–3146. [Google Scholar] [CrossRef] [PubMed]
- Dürk, T.; Panther, E.; Müller, T.; Sorichter, S.; Ferrari, D.; Pizzirani, C.; Di Virgilio, F.; Myrtek, D.; Norgauer, J.; Idzko, M. 5-Hydroxytryptamine modulates cytokine and chemokine production in LPS-primed human monocytes via stimulation of different 5-HTR subtypes. Int. Immunol. 2005, 17, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Overgaard, K.; Fredsted, A.; Hyldal, A.; Ingemann-Hansen, T.; Gissel, H.; Clausen, T. Effects of Running Distance and Training on Ca2+ Content and Damage in Human Muscle. Med. Sci. Sports Exerc. 2004, 36, 821–829. [Google Scholar] [CrossRef]
- Wessely-Szponder, J.; Bełkot, Z.; Bobowiec, R.; Kosior-Korzecka, U.; Wójcik, M. Trasport induced inflammatory responses in horses. Pol. J. Vet. Sci. 2015, 18, 407–413. [Google Scholar] [CrossRef]
- Varberg, S.; Jönsson, L.; Lindholm, A.; Holmgren, N. Muscle histopathology and plasma aspartate aminotransferase, creatine kinase and myoglobin changes with exercise in horses with recurrent exertional rhabdomyolysis. Equine Vet. J. 1993, 25, 11–16. [Google Scholar] [CrossRef]
- Boyd, J.W. The mechanisms relating to increases in plasma enzymes and isoenzymes in diseases of animals. Vet. Clin. Pathol. 1985, 12, 9–25. [Google Scholar] [CrossRef]
- Tischner, M., Jr.; Niezgoda, J. Effect of transport on the intensity of stress reactions in mares and foals. J. Reprod. Fertil. Suppl. 2000, 56, 725–730. [Google Scholar]
- Thomassian, A.; de Carvalho, F.; Watanabe, M.J.; da Silveira, V.F.; Alves, A.L.G.; Hussni, C.A.; de Mello Nicoletti, J.L. Serum activities of aspartate aminotransferase, creatine kinase and lactate dehydrogenase in arabian horses submitted to standard incremental exercise test. Braz. J. Vet. Res. Anim. Sci. 2007, 44, 183–190. [Google Scholar] [CrossRef]
- Shaashua, L.; Sominsky, L.; Levi, B.; Sorski, L.; Reznick, M.; Page, G.G.; Ben-Eliyahu, S. In vivo suppression of plasma IL-12 levels by acute and chronic stress paradigms: Potential mediating mechanisms and sex differences. Brain Behav. Immun. 2012, 26, 996–1005. [Google Scholar] [CrossRef]
- Cloëz-Tayarani, I.; Petit-Bertron, A.-F.; Venters, H.D.; Cavaillon, J.-M. Differential effect of serotonin on cytokine production in lipopolysaccharide-stimulated human peripheral blood mononuclear cells: Involvement of 5-hydroxytryptamine2A receptors. Int. Immunol. 2003, 15, 233–240. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T0 | T1 | |
|---|---|---|
| TG (mg/dL) | 42.6 ± 9.6 | 48.9 ± 7.7 |
| TCHOL (mg/dL) | 98.0 ± 7.6 | 76.8 ± 9.5 |
| AST (U/L) | 108.2 ± 23.0 | 70.0 ± 17.3 |
| CPK (U/L) | 93.3 ± 34.6 | 120.1 ± 25.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruschetta, G.; Zanghì, G.; Giunta, R.P.; Ferlazzo, A.M.; Satué, K.; D’Ascola, A.; Fazio, E. Short Road Transport and Slaughter Stress Affects the Expression Profile of Serotonin Receptors, Adrenocortical, and Hematochemical Responses in Horses. Vet. Sci. 2024, 11, 113. https://doi.org/10.3390/vetsci11030113
Bruschetta G, Zanghì G, Giunta RP, Ferlazzo AM, Satué K, D’Ascola A, Fazio E. Short Road Transport and Slaughter Stress Affects the Expression Profile of Serotonin Receptors, Adrenocortical, and Hematochemical Responses in Horses. Veterinary Sciences. 2024; 11(3):113. https://doi.org/10.3390/vetsci11030113
Chicago/Turabian StyleBruschetta, Giuseppe, Gabriella Zanghì, Renato Paolo Giunta, Alida Maria Ferlazzo, Katiuska Satué, Angela D’Ascola, and Esterina Fazio. 2024. "Short Road Transport and Slaughter Stress Affects the Expression Profile of Serotonin Receptors, Adrenocortical, and Hematochemical Responses in Horses" Veterinary Sciences 11, no. 3: 113. https://doi.org/10.3390/vetsci11030113