
Collaborative Metabolism: Gut Microbes Play a Key Role in Canine and Feline Bile Acid Metabolism


Abstract
:Simple Summary
Abstract
1. Introduction
2. Collaborative Physiology of Bile Acid Metabolism
2.1. Bile Acids as Signaling Molecules—Moving beyond Digestion
2.2. Microbial Modifications Expand Bile Acid Pool Diversity and Signaling Potential
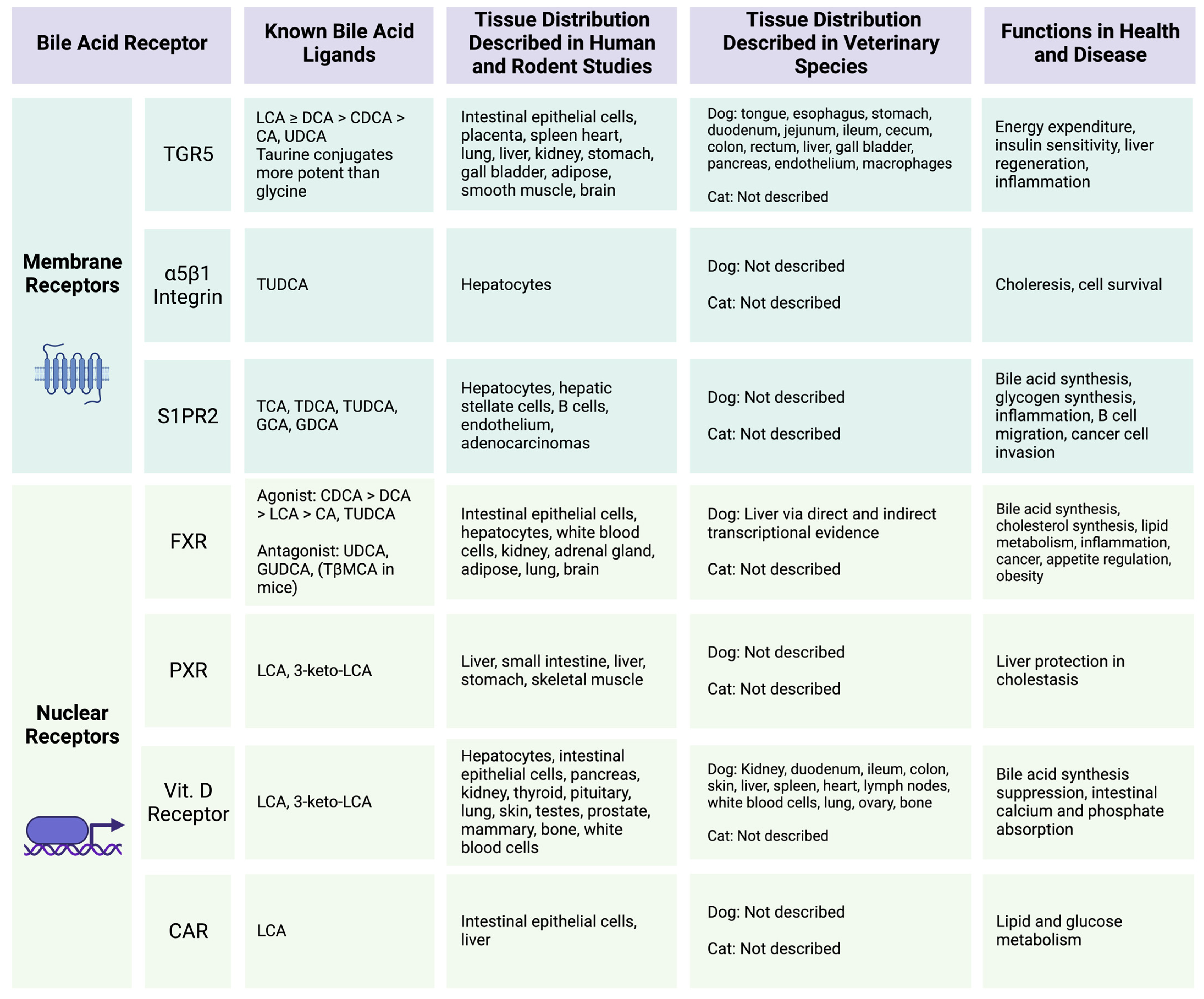
2.3. Bile Acid Reabsorption and Host Receptor Affinity

3. Canine Microbial-Derived Bile Acids
3.1. Canine Microbial-Derived Bile Acids in Health
3.2. Effects of Antimicrobials on Canine Microbial-Derived Bile Acids
3.3. Impact of Diet on Canine Microbial-Derived Bile Acids
3.4. Impact of Gastrointestinal and Pancreatic Diseases on Canine Microbial-Derived Bile Acids
3.5. Impact of Non-Gastrointestinal Disease on Canine Microbial-Derived Bile Acids

{kind=link}
{kind=link}
{kind=link}
| Author and Year | Study Context | Disease Status | Bile Acid Sample | Primary Bile Acids Reported | Secondary Bile Acids Reported | Bile Acid Assessment Method | Microbiota Assessment Method | Evidence Level |
|---|---|---|---|---|---|---|---|---|
| Honneffer et al., 2017 [139] | Healthy | HC (n = 6) | 3h post-mortem intestinal contents | None | None | GC-TOF-MS, PICRUSt | 16S rRNA amplicon | 4 |
| Blake et al., 2020 [114] | Development | HC (n = 86) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI, enteric pathogen qPCR | 2 |
| Belchik et al., 2023 [115] | Antimicrobials, diet | HC (n = 12) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI, 16S rRNA amplicon | 2 |
| Marclay et al., 2022 [59] | Antimicrobials, FMT | HC (n = 16) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI | 1 |
| Whittemore et al., 2021 [116] | Antimicrobials | HC (n = 22) | Lyophilized feces | CA | DCA, LCA | GC-TOF-MS | DI, 16S rRNA amplicon | 1 |
| Pilla et al., 2020 [117] | Antimicrobials | HC (n = 24) | Lyophilized feces | CA, CDCA | DCA, LCA | GC-MS | DI, 16S rRNA amplicon | 2 |
| Manchester et al., 2019 [118] | Antimicrobials | HC (n = 16) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI, 16S rRNA amplicon | 1 |
| Phungviwatnikul et al., 2021 [122] | Diet | HC (n = 28) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | 16S rRNA amplicon | 2 |
| Reis et al., 2021 [120] | Diet | HC (n = 8) | Voided feces | N/A | N/A | Colorimetric Total BA | None | 2 |
| Donadelli et al., 2020 [123] | Diet | HC (n = 8) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | None | 4 |
| Pezzali et al., 2020 [124] | Diet | HC (n = 12) | Fresh frozen feces | CA, CDCA | DCA, LCA | HPLC | None | 2 |
| Schmidt et al., 2018 [121] | Diet | No GI Disease (n = 46) | Lyophilized feces | All primary together | All secondary together | GC-TOF-MS | DI, 16S rRNA amplicon | 2 |
| Herstad et al., 2018 [126] | Diet | HC (n = 8) | Freeze-dried feces | CA, CDCA, GCA, GCDCA, TCA. TCDCA | DCA, GDCA, GLCA, GUDCA, LCA, TLCA, TDCA, TUDCA, UDCA | LC-MS/MS | 16S rRNA amplicon in prior publication | 4 |
| Vecchiato et al., 2023 [131] | CIE | CIE (n = 18) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI | 4 |
| Galler et al., 2022 [130] | CIE | HC (n = 26), CIE (n = 14) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI | 3 |
| Blake et al., 2019 [128] | CIE, EPI | HC (n = 34), CIE (n = 15), EPI (n = 36) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI, 16S rRNA amplicon | 3 |
| Guard et al., 2019 [127] | SRE | HC (n = 24), SRE (n = 23) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI | 3 |
| Wang et al., 2019 [52] | CIE | HC (n = 24), CIE (n = 29) | Voided feces | CA, CDCA, GCA, GCDCA, TCA, TCDCA, αMCA, βMCA | DCA, GDCA, LCA, TDCA, TLCA, γMCA, ωMCA | UPLC | Metagenomics, 16S rRNA amplicon | 2 |
| Giaretta et al., 2018 [64] | CIE | HC (n = 11), CIE (n = 24) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | DI | 3 |
| Chaitman et al., 2020 [129] | NAD | HC (n = 14), NAD (n = 18) | Lyophilized feces | CA, CDCA | DCA, LCA UDCA | GC-MS | DI, 16S rRNA amplicon | 3 |
| Phungviwatnikul et al., 2022 [119] | Diet, Overweight | OW (n = 12) | Lyophilized feces | CA, CDCA | DCA, LCA UDCA | GC-MS | 16S rRNA amplicon | 2 |
| Alexander et al., 2018 [125] | Diet, overweight | OW (n = 9) | Lyophilized feces | CA | 3-oxoCDCA, 7-oxoDCA, DCA, isoLCA, LCA | HPLC | DI, 16S rRNA amplicon | 2 |
| Li et al., 2021 [133] | MMVD | HC (n = 17), MMVD (n = 75) | Voided feces | CA, CDCA, GCA, GCDCA, TCA, TCDCA | DCA, GDCA, GLCA, GUDCA, LCA, TDCA, TLCA, TUDCA, UDCA | LC-MS/MS, UPLC | DI, 16S rRNA amplicon | 3 |
| Jergens et al., 2019 [113] | DM | HC (n = 10), DM (n = 10) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | 16S rRNA amplicon | 3 |
3.6. Conclusions Regarding Canine Microbial-Derived Bile Acids
4. Feline Microbial-Derived Bile Acids
4.1. Feline Microbial-Derived Bile Acids in Health
4.2. Feline Microbial-Derived Bile Acids in Disease
| Author and Year | Study Context | Disease Status | Bile Acid Sample | Primary Bile Acids Reported | Secondary Bile Acids Reported | Bile Acid Assessment Method | Microbiota Assessment Method | Evidence Level |
|---|---|---|---|---|---|---|---|---|
| Ephraim and Jewell, 2021 [156] | Diet, aging | HC (n = 40) | Frozen feces homogenate | CA | 7α-hydroxy cholestenone, dehydroLCA, DCA, isoUDCA, LCA, UDCA | GC-MS, LC-MS | 16S rRNA Amplicon | 2 |
| Jackson et al., 2020 [157] | Diet | HC (n = 36) | Frozen feces homogenate | * CA, CDCA, GCDCA, TCA, TCDCA | * 3-dehydroCA, 6-oxoLCA, 7-ketoDCA, 7-ketoLCA, 12-dehydroCA, dehydroLCA, DCA, GDCA, GLCA, HCA, isoHDCA, isoUDCA, LCA, TDCA, UCA | GC-MS, LC-MS | 16S rRNA Amplicon | 2 |
| Anantharaman-Barr et al., 1994 [158] | Diet | HC (n = 10) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA + HDCA | GC-MS | None | 4 |
| Whittemore et al., 2019 [160] | Antimicrobials | HC (n = 16) | Lyophilized feces | None | DCA | GC-TOF-MS | DI, 16S rRNA Amplicon | 1 |
| Whittemore et al., 2018 [159] | Antimicrobials | HC (n = 16) | Lyophilized feces | CA | DCA | GC-TOF-MS | DI, 16S rRNA Amplicon | 1 |
| Stavroulaki et al., 2022 [161] | Antimicrobials, development | URI (n = 45) | Lyophilized feces | CA, CDCA | DCA, LCA, UDCA | GC-MS | None | 3 |
| Hall et al., 2020 [166] | CKD, diet | HC (n = 10), CKD (n = 10) | Frozen feces homogenate | * CA, CDCA, GCA, TCA, TCDCA, βMCA | * 3-dehydroCA, 3β-hydroxy-5-cholenoic acid, 7-ketoDCA, 7-ketoLCA, 7α-hydroxycholestenone, 7, 12-diketoLCA, 12-dehydroCA, dehydroLCA, DCA, HCA, isoUDCA, LCA, TDCA, TLCA, TUDCA, UCA, UDCA | GC-MS, LC-MS | None | 3 |
4.3. Conclusions Regarding Feline Microbial-Derived Bile Acids
5. Overall Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ciaula, A.D.; Garruti, G.; Baccetto, R.L.; Molina-Molina, E.; Bonfrate, L.; Portincasa, P.; Wang, D.Q.H. Bile Acid Physiology. Ann. Hepatol. 2018, 16, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.F.; Small, D.M. Detergent Properties of Bile Salts: Correlation with Physiologic Function. Annu. Rev. Med. 1967, 18, 333–376. [Google Scholar] [CrossRef] [PubMed]
- Copple, B.L.; Li, T. Pharmacology of Bile Acid Receptors: Evolution of Bile Acids from Simple Detergents to Complex Signaling Molecules. Pharmacol. Res. 2016, 104, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Perino, A.; Schoonjans, K. Metabolic Messengers: Bile Acids. Nat. Metab. 2022, 4, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Claudel, T.; Staels, B.; Kuipers, F. The Farnesoid X Receptor. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2020–2030. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Jiang, C.; Cheng, J.; Krausz, K.W.; Li, T.; Ferrell, J.M.; Gonzalez, F.J.; Chiang, J.Y.L. Bile Acid Signaling in Lipid Metabolism: Metabolomic and Lipidomic Analysis of Lipid and Bile Acid Markers Linked to Anti-Obesity and Anti-Diabetes in Mice. Biochim. Et Biophys. Acta BBA Mol. Cell Biol. Lipids 2015, 1851, 19–29. [Google Scholar] [CrossRef]
- Vrieze, A.; Out, C.; Fuentes, S.; Jonker, L.; Reuling, I.; Kootte, R.S.; van Nood, E.; Holleman, F.; Knaapen, M.; Romijn, J.A.; et al. Impact of Oral Vancomycin on Gut Microbiota, Bile Acid Metabolism, and Insulin Sensitivity. J. Hepatol. 2014, 60, 824–831. [Google Scholar] [CrossRef]
- Fiorucci, S.; Cipriani, S.; Mencarelli, A.; Renga, B.; Distrutti, E.; Baldelli, F. Counter-Regulatory Role of Bile Acid Activated Receptors in Immunity and Inflammation. Curr. Mol. Med. 2010, 10, 579–595. [Google Scholar]
- Zhu, C.; Fuchs, C.D.; Halilbasic, E.; Trauner, M. Bile Acids in Regulation of Inflammation and Immunity: Friend or Foe? Clin. Exp. Rheumatol. 2016, 34, 25–31. [Google Scholar]
- Watanabe, M.; Houten, S.M.; Mataki, C.; Christoffolete, M.A.; Kim, B.W.; Sato, H.; Messaddeq, N.; Harney, J.W.; Ezaki, O.; Kodama, T.; et al. Bile Acids Induce Energy Expenditure by Promoting Intracellular Thyroid Hormone Activation. Nature 2006, 439, 484–489. [Google Scholar] [CrossRef]
- Faden, H. Review and Commentary on the Importance of Bile Acids in the Life Cycle of Clostridioides Difficile in Children and Adults. J. Pediatr. Infect. Dis. Soc. 2021, 10, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhan, S.; Tian, Z.; Liu, C.; Xie, Z.; Zhang, S.; Chen, M.; Zeng, Z.; Zhuang, X. Alterations in Bile Acid Metabolism Associated With Inflammatory Bowel Disease. Inflamm. Bowel. Dis. 2021, 27, 1525–1540. [Google Scholar] [CrossRef] [PubMed]
- Tiratterra, E.; Franco, P.; Porru, E.; Katsanos, K.H.; Christodoulou, D.K.; Roda, G. Role of Bile Acids in Inflammatory Bowel Disease. Ann. Gastroenterol. 2018, 31, 266–272. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-Omics of the Gut Microbial Ecosystem in Inflammatory Bowel Diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef] [PubMed]
- McMillin, M.; DeMorrow, S. Effects of Bile Acids on Neurological Function and Disease. FASEB J. 2016, 30, 3658–3668. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morató, J.; Matthan, N.R. Nutrition and Gastrointestinal Microbiota, Microbial-Derived Secondary Bile Acids, and Cardiovascular Disease. Curr. Atheroscler. Rep. 2020, 22, 47. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Talavera, O.; Tailleux, A.; Lefebvre, P.; Staels, B. Bile Acid Control of Metabolism and Inflammation in Obesity, Type 2 Diabetes, Dyslipidemia, and Nonalcoholic Fatty Liver Disease. Gastroenterology 2017, 152, 1679–1694.e3. [Google Scholar] [CrossRef]
- Winston, J.A.; Theriot, C.M. Diversification of Host Bile Acids by Members of the Gut Microbiota. Gut Microbes 2020, 11, 158–171. [Google Scholar] [CrossRef]
- Cai, J.; Rimal, B.; Jiang, C.; Chiang, J.Y.L.; Patterson, A.D. Bile Acid Metabolism and Signaling, the Microbiota, and Metabolic Disease. Pharmacol. Ther. 2022, 237, 108238. [Google Scholar] [CrossRef]
- Collins, S.L.; Stine, J.G.; Bisanz, J.E.; Okafor, C.D.; Patterson, A.D. Bile Acids and the Gut Microbiota: Metabolic Interactions and Impacts on Disease. Nat. Rev. Microbiol. 2023, 21, 236–247. [Google Scholar] [CrossRef]
- Suchodolski, J.S. Analysis of the Gut Microbiome in Dogs and Cats. Vet. Clin. Pathol. 2022, 50, 6–17. [Google Scholar] [CrossRef]
- Ziese, A.-L.; Suchodolski, J.S. Impact of Changes in Gastrointestinal Microbiota in Canine and Feline Digestive Diseases. Vet. Clin. Small Anim. Pract. 2021, 51, 155–169. [Google Scholar] [CrossRef]
- Russell, D.W.; Setchell, K.D.R. Bile Acid Biosynthesis. Biochemistry 1992, 31, 4737–4749. [Google Scholar] [CrossRef]
- Hagey, L.R.; Crombie, D.L.; Espinosa, E.; Carey, M.C.; Igimi, H.; Hofmann, A.F. Ursodeoxycholic Acid in the Ursidae: Biliary Bile Acids of Bears, Pandas, and Related Carnivores. J. Lipid Res. 1993, 34, 1911–1917. [Google Scholar] [CrossRef]
- Zhang, Y.; Limaye, P.B.; Renaud, H.J.; Klaassen, C.D. Effect of Various Antibiotics on Modulation of Intestinal Microbiota and Bile Acid Profile in Mice. Toxicol. Appl. Pharmacol. 2014, 277, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Botham, K.M.; Boyd, G.S. The Metabolism of Chenodeoxycholic Acid to β-Muricholic Acid in Rat Liver. Eur. J. Biochem. 1983, 134, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Vessey, D.A. The Biochemical Basis for the Conjugation of Bile Acids with Either Glycine or Taurine. Biochem. J. 1978, 174, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.; Nicolosi, R.J.; Hayes, K.C. Dietary Influence on Bile Acid Conjugation in the Cat. J. Nutr. 1976, 106, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Sasaki, S.-I.; Toyoda, A.; Wei, F.-Y.; Shirai, M.; Morishita, Y.; Ikegami, T.; Tomizawa, K.; Honda, A. Impaired Bile Acid Metabolism with Defectives of Mitochondrial-tRNA Taurine Modification and Bile Acid Taurine Conjugation in the Taurine Depleted Cats. Sci. Rep. 2020, 10, 4915. [Google Scholar] [CrossRef] [PubMed]
- Washizu, T.; Ikenaga, H.; Washizu, M.; Ishida, T.; Tomoda, I.; Kaneko, J.J. Bile Acid Composition of Dog and Cat Gall-Bladder Bile. Jpn. J. Vet. Sci. 1990, 52, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Washizu, T.; Ishida, T.; Washizu, M.; Tomoda, I.; Kaneko, J.J. Changes in Bile Acid Composition of Serum and Gallbladder Bile in Bile Duct Ligated Dogs. J. Vet. Med. Sci. 1994, 56, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Kook, P.H.; Schellenberg, S.; Rentsch, K.M.; Reusch, C.E.; Glaus, T.M. Effect of Twice-Daily Oral Administration of Hydrocortisone on the Bile Acids Composition of Gallbladder Bile in Dogs. Am. J. Vet. Res. 2011, 72, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
- Sjövall, J. Bile Acids in Man under Normal and Pathological Conditions Bile Acids and Steroids 73. Clin. Chim. Acta 1960, 5, 33–41. [Google Scholar] [CrossRef]
- Hafkenscheid, J.C.M.; Hectors, M.P.C. An Enzymic Method for the Determination of the Glycine/Taurine Ratio of Conjugated Bile Acids in Bile. Clin. Chim. Acta 1975, 65, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Ticho, A.L.; Malhotra, P.; Dudeja, P.K.; Gill, R.K.; Alrefai, W.A. Intestinal Absorption of Bile Acids in Health and Disease. Compr. Physiol. 2019, 10, 21–56. [Google Scholar] [CrossRef] [PubMed]
- Northfield, T.C.; McColl, I. Postprandial Concentrations of Free and Conjugated Bile Acids down the Length of the Normal Human Small Intestine. Gut 1973, 14, 513–518. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile Salt Biotransformations by Human Intestinal Bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef]
- Foley, M.H.; O’Flaherty, S.; Barrangou, R.; Theriot, C.M. Bile Salt Hydrolases: Gatekeepers of Bile Acid Metabolism and Host-Microbiome Crosstalk in the Gastrointestinal Tract. PLOS Pathog. 2019, 15, e1007581. [Google Scholar] [CrossRef]
- Quinn, R.A.; Melnik, A.V.; Vrbanac, A.; Fu, T.; Patras, K.A.; Christy, M.P.; Bodai, Z.; Belda-Ferre, P.; Tripathi, A.; Chung, L.K.; et al. Global Chemical Effects of the Microbiome Include New Bile-Acid Conjugations. Nature 2020, 579, 123–129. [Google Scholar] [CrossRef]
- Zhu, Q.-F.; Wang, Y.-Z.; An, N.; Hao, J.-D.; Mei, P.-C.; Bai, Y.-L.; Hu, Y.-N.; Bai, P.-R.; Feng, Y.-Q. Alternating Dual-Collision Energy Scanning Mass Spectrometry Approach: Discovery of Novel Microbial Bile-Acid Conjugates. Anal. Chem. 2022, 94, 2655–2664. [Google Scholar] [CrossRef]
- Wang, Y.-Z.; Mei, P.-C.; Bai, P.-R.; An, N.; He, J.-G.; Wang, J.; Zhu, Q.-F.; Feng, Y.-Q. A Strategy for Screening and Identification of New Amino Acid-Conjugated Bile Acids with High Coverage by Liquid Chromatography-Mass Spectrometry. Anal. Chim. Acta 2023, 1239, 340691. [Google Scholar] [CrossRef]
- Guzior, D.V.; Quinn, R.A. Review: Microbial Transformations of Human Bile Acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef]
- Dawson, P.A.; Karpen, S.J. Intestinal Transport and Metabolism of Bile Acids. J. Lipid Res. 2015, 56, 1085–1099. [Google Scholar] [CrossRef] [PubMed]
- Alnouti, Y. Bile Acid Sulfation: A Pathway of Bile Acid Elimination and Detoxification. Toxicol. Sci. 2009, 108, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Huijghebaert, S.; Parmentier, G.; Eyssen, H. Specificity of Bile Salt Sulfatase Activity in Man, Mouse and Rat Intestinal Microflora. J. Steroid Biochem. 1984, 20, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Funabashi, M.; Grove, T.L.; Wang, M.; Varma, Y.; McFadden, M.E.; Brown, L.C.; Guo, C.; Higginbottom, S.; Almo, S.C.; Fischbach, M.A. A Metabolic Pathway for Bile Acid Dehydroxylation by the Gut Microbiome. Nature 2020, 582, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B.; Bajaj, J.S. Gut Microbiota, Cirrhosis, and Alcohol Regulate Bile Acid Metabolism in the Gut. Dig. Dis. 2015, 33, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Streidl, T.; Karkossa, I.; Segura Muñoz, R.R.; Eberl, C.; Zaufel, A.; Plagge, J.; Schmaltz, R.; Schubert, K.; Basic, M.; Schneider, K.M.; et al. The Gut Bacterium Extibacter Muris Produces Secondary Bile Acids and Influences Liver Physiology in Gnotobiotic Mice. Gut Microbes 2021, 13, 1854008. [Google Scholar] [CrossRef] [PubMed]
- Vital, M.; Rud, T.; Rath, S.; Pieper, D.H.; Schlüter, D. Diversity of Bacteria Exhibiting Bile Acid-Inducible 7α-Dehydroxylation Genes in the Human Gut. Comput. Struct. Biotechnol. J. 2019, 17, 1016–1019. [Google Scholar] [CrossRef]
- Kim, K.H.; Park, D.; Jia, B.; Baek, J.H.; Hahn, Y.; Jeon, C.O. Identification and Characterization of Major Bile Acid 7α-Dehydroxylating Bacteria in the Human Gut. mSystems 2022, 7, e00455-22. [Google Scholar] [CrossRef]
- Stone, N.E.; Nunnally, A.E.; Roe, C.C.; Hornstra, H.M.; Wagner, D.M.; Sahl, J.W. Complete Genome Sequence of Peptacetobacter (Clostridium) Hiranonis Strain DGF055142, Isolated from Dog Feces from Flagstaff, Arizona, USA, 2019. Microbiol. Resour. Announc. 2021, 10, e00067-21. [Google Scholar] [CrossRef]
- Wang, S.; Martins, R.; Sullivan, M.C.; Friedman, E.S.; Misic, A.M.; El-Fahmawi, A.; De Martinis, E.C.P.; O’Brien, K.; Chen, Y.; Bradley, C.; et al. Diet-Induced Remission in Chronic Enteropathy Is Associated with Altered Microbial Community Structure and Synthesis of Secondary Bile Acids. Microbiome 2019, 7, 126. [Google Scholar] [CrossRef] [PubMed]
- AlShawaqfeh, M.; Wajid, B.; Minamoto, Y.; Markel, M.; Lidbury, J.; Steiner, J.; Serpedin, E.; Suchodolski, J. A Dysbiosis Index to Assess Microbial Changes in Fecal Samples of Dogs with Chronic Inflammatory Enteropathy. FEMS Microbiol. Ecol. 2017, 93, fix136. [Google Scholar] [CrossRef]
- Sung, C.-H.; Marsilio, S.; Chow, B.; Zornow, K.A.; Slovak, J.E.; Pilla, R.; Lidbury, J.A.; Steiner, J.M.; Park, S.Y.; Hong, M.-P.; et al. Dysbiosis Index to Evaluate the Fecal Microbiota in Healthy Cats and Cats with Chronic Enteropathies. J. Feline Med. Surg. 2022, 24, e1–e12. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.F.; Hagey, L.R. Key Discoveries in Bile Acid Chemistry and Biology and Their Clinical Applications: History of the Last Eight Decades. J. Lipid Res. 2014, 55, 1553–1595. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.; Twedt, D. Oxidative Stress and Liver Disease. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 125–135. [Google Scholar] [CrossRef]
- Setchell, K.D.; Lawson, A.M.; Tanida, N.; Sjövall, J. General Methods for the Analysis of Metabolic Profiles of Bile Acids and Related Compounds in Feces. J. Lipid Res. 1983, 24, 1085–1100. [Google Scholar] [CrossRef]
- Kakiyama, G.; Muto, A.; Takei, H.; Nittono, H.; Murai, T.; Kurosawa, T.; Hofmann, A.F.; Pandak, W.M.; Bajaj, J.S. A Simple and Accurate HPLC Method for Fecal Bile Acid Profile in Healthy and Cirrhotic Subjects: Validation by GC-MS and LC-MS. J. Lipid Res. 2014, 55, 978–990. [Google Scholar] [CrossRef]
- Marclay, M.; Dwyer, E.; Suchodolski, J.S.; Lidbury, J.A.; Steiner, J.M.; Gaschen, F.P. Recovery of Fecal Microbiome and Bile Acids in Healthy Dogs after Tylosin Administration with and without Fecal Microbiota Transplantation. Vet. Sci. 2022, 9, 324. [Google Scholar] [CrossRef]
- Hofmann, A.F. The Enterohepatic Circulation of Bile Acids in Mammals: Form and Functions. FBL 2009, 14, 2584–2598. [Google Scholar] [CrossRef]
- Dawson, P.A. Role of the Intestinal Bile Acid Transporters in Bile Acid and Drug Disposition. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 169–203. [Google Scholar] [CrossRef]
- Krag, E.; Phillips, S.F. Active and Passive Bile Acid Absorption in Man. PERFUSION STUDIES OF THE ILEUM AND JEJUNUM. J. Clin. Investig. 1974, 53, 1686–1694. [Google Scholar] [CrossRef]
- Li, M.; Wang, Q.; Li, Y.; Cao, S.; Zhang, Y.; Wang, Z.; Liu, G.; Li, J.; Gu, B. Apical Sodium-Dependent Bile Acid Transporter, Drug Target for Bile Acid Related Diseases and Delivery Target for Prodrugs: Current and Future Challenges. Pharmacol. Ther. 2020, 212, 107539. [Google Scholar] [CrossRef]
- Giaretta, P.R.; Rech, R.R.; Guard, B.C.; Blake, A.B.; Blick, A.K.; Steiner, J.M.; Lidbury, J.A.; Cook, A.K.; Hanifeh, M.; Spillmann, T.; et al. Comparison of Intestinal Expression of the Apical Sodium-Dependent Bile Acid Transporter between Dogs with and without Chronic Inflammatory Enteropathy. J. Vet. Intern. Med. 2018, 32, 1918–1926. [Google Scholar] [CrossRef]
- Jung, D.; Fantin, A.C.; Scheurer, U.; Fried, M.; Kullak-Ublick, G.A. Human Ileal Bile Acid Transporter Gene ASBT (SLC10A2) Is Transactivated by the Glucocorticoid Receptor. Gut 2004, 53, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Ticho, A.L.; Malhotra, P.; Dudeja, P.K.; Gill, R.K.; Alrefai, W.A. Bile Acid Receptors and Gastrointestinal Functions. Liver Res. 2019, 3, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Sayin, S.I.; Wahlström, A.; Felin, J.; Jäntti, S.; Marschall, H.-U.; Bamberg, K.; Angelin, B.; Hyötyläinen, T.; Orešič, M.; Bäckhed, F. Gut Microbiota Regulates Bile Acid Metabolism by Reducing the Levels of Tauro-Beta-Muricholic Acid, a Naturally Occurring FXR Antagonist. Cell Metab. 2013, 17, 225–235. [Google Scholar] [CrossRef]
- Sun, L.; Xie, C.; Wang, G.; Wu, Y.; Wu, Q.; Wang, X.; Liu, J.; Deng, Y.; Xia, J.; Chen, B.; et al. Gut Microbiota and Intestinal FXR Mediate the Clinical Benefits of Metformin. Nat. Med. 2018, 24, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Y.; Hong, S.H.; Lee, Y.J.; Chung, S.S.; Jung, H.S.; Park, S.G.; Park, K.S. Tauroursodeoxycholate (TUDCA), Chemical Chaperone, Enhances Function of Islets by Reducing ER Stress. Biochem. Biophys. Res. Commun. 2010, 397, 735–739. [Google Scholar] [CrossRef]
- Huang, C.; Wang, J.; Hu, W.; Wang, C.; Lu, X.; Tong, L.; Wu, F.; Zhang, W. Identification of Functional Farnesoid X Receptors in Brain Neurons. FEBS Lett. 2016, 590, 3233–3242. [Google Scholar] [CrossRef]
- Dehondt, H.; Marino, A.; Butruille, L.; Mogilenko, D.A.; Nzoussi Loubota, A.C.; Chávez-Talavera, O.; Dorchies, E.; Vallez, E.; Haas, J.; Derudas, B.; et al. Adipocyte-Specific FXR-Deficiency Protects Adipose Tissue from Oxidative Stress and Insulin Resistance and Improves Glucose Homeostasis. Mol. Metab. 2023, 69, 101686. [Google Scholar] [CrossRef]
- Chen, B.; Cai, H.-R.; Xue, S.; You, W.-J.; Liu, B.; Jiang, H.-D. Bile Acids Induce Activation of Alveolar Epithelial Cells and Lung Fibroblasts through Farnesoid X Receptor-Dependent and Independent Pathways. Respirology 2016, 21, 1075–1080. [Google Scholar] [CrossRef]
- Brevini, T.; Maes, M.; Webb, G.J.; John, B.V.; Fuchs, C.D.; Buescher, G.; Wang, L.; Griffiths, C.; Brown, M.L.; Scott, W.E.; et al. FXR Inhibition May Protect from SARS-CoV-2 Infection by Reducing ACE2. Nature 2023, 615, 134–142. [Google Scholar] [CrossRef]
- Wang, H.; Chen, J.; Hollister, K.; Sowers, L.C.; Forman, B.M. Endogenous Bile Acids Are Ligands for the Nuclear Receptor FXR/BAR. Mol. Cell 1999, 3, 543–553. [Google Scholar] [CrossRef]
- Winston, J.A.; Rivera, A.; Cai, J.; Patterson, A.D.; Theriot, C.M. Secondary Bile Acid Ursodeoxycholic Acid Alters Weight, the Gut Microbiota, and the Bile Acid Pool in Conventional Mice. PLoS ONE 2021, 16, e0246161. [Google Scholar] [CrossRef]
- Makishima, M.; Lu, T.T.; Xie, W.; Whitfield, G.K.; Domoto, H.; Evans, R.M.; Haussler, M.R.; Mangelsdorf, D.J. Vitamin D Receptor As an Intestinal Bile Acid Sensor. Science 2002, 296, 1313–1316. [Google Scholar] [CrossRef]
- Cartwright, J.A.; Gow, A.G.; Milne, E.; Drummond, D.; Smith, S.; Handel, I.; Mellanby, R.J. Vitamin D Receptor Expression in Dogs. J. Vet. Intern. Med. 2018, 32, 764–774. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, J.; DeLuca, H.F. Where Is the Vitamin D Receptor? Arch. Biochem. Biophys. 2012, 523, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Li, T.; Ellis, E.; Strom, S.; Chiang, J.Y.L. A Novel Bile Acid-Activated Vitamin D Receptor Signaling in Human Hepatocytes. Mol. Endocrinol. 2010, 24, 1151–1164. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, N.; Matsui, I.; Ishizuka, S.; Inoue, K.; Matsumoto, A.; Shimada, K.; Hori, S.; Lee, D.G.; Yasuda, S.; Katsuma, Y.; et al. Lithocholic Acid Increases Intestinal Phosphate and Calcium Absorption in a Vitamin D Receptor Dependent but Transcellular Pathway Independent Manner. Kidney Int. 2020, 97, 1164–1180. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L.; Goodwin, B.; Jones, S.A.; Hawkins-Brown, D.; MacKenzie, K.I.; LaTour, A.; Liu, Y.; Klaassen, C.D.; Brown, K.K.; Reinhard, J.; et al. The Nuclear Receptor PXR Is a Lithocholic Acid Sensor That Protects against Liver Toxicity. Proc. Natl. Acad. Sci. USA 2001, 98, 3369–3374. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Naito, S.; Yokoi, T. Tissue-Specific mRNA Expression Profiles of Human Nuclear Receptor Subfamilies. Drug Metab. Pharmacokinet. 2004, 19, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Moore, J.T.; Wade, L.; Staudinger, J.L.; Watson, M.A.; Jones, S.A.; McKee, D.D.; Oliver, B.B.; Willson, T.M.; Zetterström, R.H.; et al. An Orphan Nuclear Receptor Activated by Pregnanes Defines a Novel Steroid Signaling Pathway. Cell 1998, 92, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.-J.; Wang, L. Bile Acid-Activated Receptors: A Review on FXR and Other Nuclear Receptors. In Bile Acids and Their Receptors; Fiorucci, S., Distrutti, E., Eds.; Handbook of Experimental Pharmacology; Springer International Publishing: Cham, Switzerland, 2019; pp. 51–72. ISBN 978-3-030-22005-1. [Google Scholar]
- Katafuchi, T.; Makishima, M. Molecular Basis of Bile Acid-FXR-FGF15/19 Signaling Axis. Int. J. Mol. Sci. 2022, 23, 6046. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Cai, J.; Gonzalez, F.J. The Role of Farnesoid X Receptor in Metabolic Diseases, and Gastrointestinal and Liver Cancer. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Panzitt, K.; Zollner, G.; Marschall, H.-U.; Wagner, M. Recent Advances on FXR-Targeting Therapeutics. Mol. Cell. Endocrinol. 2022, 552, 111678. [Google Scholar] [CrossRef] [PubMed]
- Girisa, S.; Henamayee, S.; Parama, D.; Rana, V.; Dutta, U.; Kunnumakkara, A.B. Targeting Farnesoid X Receptor (FXR) for Developing Novel Therapeutics against Cancer. Mol. Biomed. 2021, 2, 21. [Google Scholar] [CrossRef]
- Otte, K.; Kranz, H.; Kober, I.; Thompson, P.; Hoefer, M.; Haubold, B.; Remmel, B.; Voss, H.; Kaiser, C.; Albers, M.; et al. Identification of Farnesoid X Receptor β as a Novel Mammalian Nuclear Receptor Sensing Lanosterol. Mol. Cell. Biol. 2003, 23, 864–872. [Google Scholar] [CrossRef]
- Stojancevic, M.; Stankov, K.; Mikov, M. The Impact of Farnesoid X Receptor Activation on Intestinal Permeability in Inflammatory Bowel Disease. Can. J. Gastroenterol. Hepatol. 2012, 26, 631–637. [Google Scholar] [CrossRef]
- Kakimoto, T.; Kanemoto, H.; Fukushima, K.; Ohno, K.; Tsujimoto, H. Effect of a High-Fat–High-Cholesterol Diet on Gallbladder Bile Acid Composition and Gallbladder Motility in Dogs. Am. J. Vet. Res. 2017, 78, 1406–1413. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chien, H.; van Wolferen, M.E.; Kruitwagen, H.S.; Oosterhoff, L.A.; Penning, L.C. Reduced FXR Target Gene Expression in Copper-Laden Livers of COMMD1-Deficient Dogs. Vet. Sci. 2019, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Kent, A.C.C.; Cross, G.; Taylor, D.R.; Sherwood, R.A.; Watson, P.J. Measurement of Serum 7α-Hydroxy-4-Cholesten-3-One as a Marker of Bile Acid Malabsorption in Dogs with Chronic Diarrhoea: A Pilot Study. Vet. Rec. Open 2016, 3, e000163. [Google Scholar] [CrossRef] [PubMed]
- Toresson, L.; Steiner, J.M.; Suchodolski, J.S. Cholestyramine Treatment in Two Dogs with Presumptive Bile Acid Diarrhoea: A Case Report. Canine Med. Genet. 2021, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Kawamata, Y.; Fujii, R.; Hosoya, M.; Harada, M.; Yoshida, H.; Miwa, M.; Fukusumi, S.; Habata, Y.; Itoh, T.; Shintani, Y.; et al. A G Protein-Coupled Receptor Responsive to Bile Acids. J. Biol. Chem. 2003, 278, 9435–9440. [Google Scholar] [CrossRef] [PubMed]
- Duboc, H.; Taché, Y.; Hofmann, A.F. The Bile Acid TGR5 Membrane Receptor: From Basic Research to Clinical Application. Dig. Liver Dis. 2014, 46, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Macchiarulo, A.; Thomas, C.; Gioiello, A.; Une, M.; Hofmann, A.F.; Saladin, R.; Schoonjans, K.; Pellicciari, R.; Auwerx, J. Novel Potent and Selective Bile Acid Derivatives as TGR5 Agonists: Biological Screening, Structure−Activity Relationships, and Molecular Modeling Studies. J. Med. Chem. 2008, 51, 1831–1841. [Google Scholar] [CrossRef]
- Yoneno, K.; Hisamatsu, T.; Shimamura, K.; Kamada, N.; Ichikawa, R.; Kitazume, M.T.; Mori, M.; Uo, M.; Namikawa, Y.; Matsuoka, K.; et al. TGR5 Signalling Inhibits the Production of Pro-Inflammatory Cytokines by in Vitro Differentiated Inflammatory and Intestinal Macrophages in Crohn’s Disease. Immunology 2013, 139, 19–29. [Google Scholar] [CrossRef]
- Giaretta, P.R.; Suchodolski, J.S.; Blick, A.K.; Steiner, J.M.; Lidbury, J.A.; Rech, R.R. Distribution of Bile Acid Receptor TGR5 in the Gastrointestinal Tract of Dogs. Histol. Histopathol. 2018, 34, 69–79. [Google Scholar] [CrossRef]
- Sommerfeld, A.; Reinehr, R.; Häussinger, D. Tauroursodeoxycholate Protects Rat Hepatocytes from Bile Acid-Induced Apoptosis via Β1-Integrin- and Protein Kinase A-Dependent Mechanisms. Cell. Physiol. Biochem. 2015, 36, 866–883. [Google Scholar] [CrossRef]
- Studer, E.; Zhou, X.; Zhao, R.; Wang, Y.; Takabe, K.; Nagahashi, M.; Pandak, W.M.; Dent, P.; Spiegel, S.; Shi, R.; et al. Conjugated Bile Acids Activate the Sphingosine-1-Phosphate Receptor 2 in Primary Rodent Hepatocytes. Hepatology 2012, 55, 267–276. [Google Scholar] [CrossRef]
- Sic, H.; Kraus, H.; Madl, J.; Flittner, K.-A.; von Münchow, A.L.; Pieper, K.; Rizzi, M.; Kienzler, A.-K.; Ayata, K.; Rauer, S.; et al. Sphingosine-1-Phosphate Receptors Control B-Cell Migration through Signaling Components Associated with Primary Immunodeficiencies, Chronic Lymphocytic Leukemia, and Multiple Sclerosis. J. Allergy Clin. Immunol. 2014, 134, 420–428.e15. [Google Scholar] [CrossRef]
- Xiong, Y.; Piao, W.; Brinkman, C.C.; Li, L.; Kulinski, J.M.; Olivera, A.; Cartier, A.; Hla, T.; Hippen, K.L.; Blazar, B.R.; et al. CD4 T Cell Sphingosine 1-Phosphate Receptor (S1PR)1 and S1PR4 and Endothelial S1PR2 Regulate Afferent Lymphatic Migration. Sci. Immunol. 2019, 4, eaav1263. [Google Scholar] [CrossRef]
- Yang, J.; Tang, X.; Liang, Z.; Chen, M.; Sun, L. Taurocholic Acid Promotes Hepatic Stellate Cell Activation via S1PR2/P38 MAPK/YAP Signaling under Cholestatic Conditions. Clin. Mol. Hepatol. 2023, 29, 465–481. [Google Scholar] [CrossRef]
- Liu, R.; Li, X.; Hylemon, P.B.; Zhou, H. Conjugated Bile Acids Promote Invasive Growth of Esophageal Adenocarcinoma Cells and Cancer Stem Cell Expansion via Sphingosine 1-Phosphate Receptor 2–Mediated Yes-Associated Protein Activation. Am. J. Pathol. 2018, 188, 2042–2058. [Google Scholar] [CrossRef]
- Wang, Y.-D.; Chen, W.-D.; Yu, D.; Forman, B.M.; Huang, W. The G-Protein-Coupled Bile Acid Receptor, Gpbar1 (TGR5), Negatively Regulates Hepatic Inflammatory Response through Antagonizing Nuclear Factor κ Light-Chain Enhancer of Activated B Cells (NF-κB) in Mice. Hepatology 2011, 54, 1421–1432. [Google Scholar] [CrossRef]
- Katsuma, S.; Hirasawa, A.; Tsujimoto, G. Bile Acids Promote Glucagon-like Peptide-1 Secretion through TGR5 in a Murine Enteroendocrine Cell Line STC-1. Biochem. Biophys Res. Commun. 2005, 329, 386–390. [Google Scholar] [CrossRef]
- Cao, W.; Tian, W.; Hong, J.; Li, D.; Tavares, R.; Noble, L.; Moss, S.F.; Resnick, M.B. Expression of Bile Acid Receptor TGR5 in Gastric Adenocarcinoma. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G322–G327. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M. TGR5 as a Therapeutic Target for Treating Obesity. Curr. Top. Med. Chem. 2010, 10, 386–396. [Google Scholar] [CrossRef] [PubMed]
- van Nierop, F.S.; Scheltema, M.J.; Eggink, H.M.; Pols, T.W.; Sonne, D.P.; Knop, F.K.; Soeters, M.R. Clinical Relevance of the Bile Acid Receptor TGR5 in Metabolism. Lancet Diabetes Endocrinol. 2017, 5, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Eggink, H.M.; Soeters, M.R.; Pols, T.W. TGR5 Ligands as Potential Therapeutics in Inflammatory Diseases. Int. J. Interferon Cytokine Mediat. Res. 2014, 6, 27–38. [Google Scholar] [CrossRef]
- Manchester, A.C.; Wheat, W.H.; Chow, L.; Dow, S. Differential modulation of innate immune response by primary and secondary bile acids in dogs. 2021 ACVIM Forum Research Abstract Program. J. Vet. Intern. Med. 2021, 35, 2943–3079. [Google Scholar] [CrossRef]
- Jergens, A.E.; Guard, B.C.; Redfern, A.; Rossi, G.; Mochel, J.P.; Pilla, R.; Chandra, L.; Seo, Y.-J.; Steiner, J.M.; Lidbury, J.; et al. Microbiota-Related Changes in Unconjugated Fecal Bile Acids Are Associated With Naturally Occurring, Insulin-Dependent Diabetes Mellitus in Dogs. Front. Vet. Sci. 2019, 6, 199. [Google Scholar] [CrossRef] [PubMed]
- Blake, A.B.; Cigarroa, A.; Klein, H.L.; Khattab, M.R.; Keating, T.; Van De Coevering, P.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. Developmental Stages in Microbiota, Bile Acids, and Clostridial Species in Healthy Puppies. J. Vet. Intern. Med. 2020, 34, 2345–2356. [Google Scholar] [CrossRef] [PubMed]
- Belchik, S.E.; Oba, P.M.; Wyss, R.; Asare, P.T.; Vidal, S.; Miao, Y.; Adesokan, Y.; Suchodolski, J.S.; Swanson, K.S. Effects of a Milk Oligosaccharide Biosimilar on Fecal Characteristics, Microbiota, and Bile Acid, Calprotectin, and Immunoglobulin Concentrations of Healthy Adult Dogs Treated with Metronidazole. J. Anim. Sci. 2023, 110, skad011. [Google Scholar] [CrossRef] [PubMed]
- Whittemore, J.C.; Price, J.M.; Moyers, T.; Suchodolski, J.S. Effects of Synbiotics on the Fecal Microbiome and Metabolomic Profiles of Healthy Research Dogs Administered Antibiotics: A Randomized, Controlled Trial. Front. Vet. Sci. 2021, 8, 665713. [Google Scholar] [CrossRef] [PubMed]
- Pilla, R.; Gaschen, F.P.; Barr, J.W.; Olson, E.; Honneffer, J.; Guard, B.C.; Blake, A.B.; Villanueva, D.; Khattab, M.R.; AlShawaqfeh, M.K.; et al. Effects of Metronidazole on the Fecal Microbiome and Metabolome in Healthy Dogs. J. Vet. Intern. Med. 2020, 34, 1853–1866. [Google Scholar] [CrossRef] [PubMed]
- Manchester, A.C.; Webb, C.B.; Blake, A.B.; Sarwar, F.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. Long-Term Impact of Tylosin on Fecal Microbiota and Fecal Bile Acids of Healthy Dogs. J. Vet. Intern. Med. 2019, 33, 2605–2617. [Google Scholar] [CrossRef] [PubMed]
- Phungviwatnikul, T.; Lee, A.H.; Belchik, S.E.; Suchodolski, J.S.; Swanson, K.S. Weight Loss and High-Protein, High-Fiber Diet Consumption Impact Blood Metabolite Profiles, Body Composition, Voluntary Physical Activity, Fecal Microbiota, and Fecal Metabolites of Adult Dogs. J. Anim. Sci. 2022, 100, skab379. [Google Scholar] [CrossRef]
- Reis, L.G.; Morris, T.; Quilliam, C.; Rodrigues, L.A.; Loewen, M.E.; Weber, L.P. The Effects of Fermentation of Low or High Tannin Fava Bean-Based Diets on Glucose Response, Cardiovascular Function, and Fecal Bile Acid Excretion during a 28-Day Feeding Period in Dogs: Comparison with Commercial Diets with Normal vs. High Protein. Metabolites 2021, 11, 878. [Google Scholar] [CrossRef]
- Schmidt, M.; Unterer, S.; Suchodolski, J.S.; Honneffer, J.B.; Guard, B.C.; Lidbury, J.A.; Steiner, J.M.; Fritz, J.; Kölle, P. The Fecal Microbiome and Metabolome Differs between Dogs Fed Bones and Raw Food (BARF) Diets and Dogs Fed Commercial Diets. PLoS ONE 2018, 13, e0201279. [Google Scholar] [CrossRef]
- Phungviwatnikul, T.; Alexander, C.; Do, S.; He, F.; Suchodolski, J.S.; de Godoy, M.R.C.; Swanson, K.S. Effects of Dietary Macronutrient Profile on Apparent Total Tract Macronutrient Digestibility and Fecal Microbiota, Fermentative Metabolites, and Bile Acids of Female Dogs after Spay Surgery. J. Anim. Sci. 2021, 99, skab225. [Google Scholar] [CrossRef] [PubMed]
- Donadelli, R.A.; Pezzali, J.G.; Oba, P.M.; Swanson, K.S.; Coon, C.; Varney, J.; Pendlebury, C.; Shoveller, A.K. A Commercial Grain-Free Diet Does Not Decrease Plasma Amino Acids and Taurine Status but Increases Bile Acid Excretion When Fed to Labrador Retrievers. Transl. Anim. Sci. 2020, 4, txaa141. [Google Scholar] [CrossRef] [PubMed]
- Pezzali, J.G.; Acuff, H.L.; Henry, W.; Alexander, C.; Swanson, K.S.; Aldrich, C.G. Effects of Different Carbohydrate Sources on Taurine Status in Healthy Beagle Dogs. J. Anim. Sci. 2020, 98, skaa010. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.; Cross, T.-W.L.; Devendran, S.; Neumer, F.; Theis, S.; Ridlon, J.M.; Suchodolski, J.S.; de Godoy, M.R.C.; Swanson, K.S. Effects of Prebiotic Inulin-Type Fructans on Blood Metabolite and Hormone Concentrations and Faecal Microbiota and Metabolites in Overweight Dogs. Br. J. Nutr. 2018, 120, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Herstad, K.M.V.; Rønning, H.T.; Bakke, A.M.; Moe, L.; Skancke, E. Changes in the Faecal Bile Acid Profile in Dogs Fed Dry Food vs High Content of Beef: A Pilot Study. Acta Vet. Scand. 2018, 60, 29. [Google Scholar] [CrossRef]
- Guard, B.C.; Honneffer, J.B.; Jergens, A.E.; Jonika, M.M.; Toresson, L.; Lawrence, Y.A.; Webb, C.B.; Hill, S.; Lidbury, J.A.; Steiner, J.M.; et al. Longitudinal Assessment of Microbial Dysbiosis, Fecal Unconjugated Bile Acid Concentrations, and Disease Activity in Dogs with Steroid-Responsive Chronic Inflammatory Enteropathy. J. Vet. Intern. Med. 2019, 33, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Blake, A.B.; Guard, B.C.; Honneffer, J.B.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. Altered Microbiota, Fecal Lactate, and Fecal Bile Acids in Dogs with Gastrointestinal Disease. PLoS ONE 2019, 14, e0224454. [Google Scholar] [CrossRef]
- Chaitman, J.; Ziese, A.-L.; Pilla, R.; Minamoto, Y.; Blake, A.B.; Guard, B.C.; Isaiah, A.; Lidbury, J.A.; Steiner, J.M.; Unterer, S.; et al. Fecal Microbial and Metabolic Profiles in Dogs With Acute Diarrhea Receiving Either Fecal Microbiota Transplantation or Oral Metronidazole. Front. Vet. Sci. 2020, 7, 192. [Google Scholar] [CrossRef]
- Galler, A.I.; Suchodolski, J.S.; Steiner, J.M.; Sung, C.-H.; Hittmair, K.M.; Richter, B.; Burgener, I.A. Microbial Dysbiosis and Fecal Metabolomic Perturbations in Yorkshire Terriers with Chronic Enteropathy. Sci. Rep. 2022, 12, 12977. [Google Scholar] [CrossRef]
- Vecchiato, C.G.; Pinna, C.; Sung, C.-H.; Borrelli De Andreis, F.; Suchodolski, J.S.; Pilla, R.; Delsante, C.; Sportelli, F.; Mammi, L.M.E.; Pietra, M.; et al. Fecal Microbiota, Bile Acids, Sterols, and Fatty Acids in Dogs with Chronic Enteropathy Fed a Home-Cooked Diet Supplemented with Coconut Oil. Animals 2023, 13, 502. [Google Scholar] [CrossRef]
- Research Communications of the 31st ECVIM-CA Online Congress. J. Vet. Intern. Med. 2021, 35, 3080–3204. [CrossRef]
- Li, Q.; Larouche-Lebel, É.; Loughran, K.A.; Huh, T.P.; Suchodolski, J.S.; Oyama, M.A. Gut Dysbiosis and Its Associations with Gut Microbiota-Derived Metabolites in Dogs with Myxomatous Mitral Valve Disease. mSystems 2021, 6, e00111-21. [Google Scholar] [CrossRef]
- Xiong, J.; Hu, H.; Xu, C.; Yin, J.; Liu, M.; Zhang, L.; Duan, Y.; Huang, Y. Development of Gut Microbiota along with Its Metabolites of Preschool Children. BMC Pediatr. 2022, 22, 25. [Google Scholar] [CrossRef]
- Berry, A.S.F.; Kelly, B.J.; Barnhart, D.; Kelly, D.J.; Beiting, D.P.; Baldassano, R.N.; Redding, L.E. Gut Microbiota Features Associated with Clostridioides Difficile Colonization in Puppies. PLoS ONE 2019, 14, e0215497. [Google Scholar] [CrossRef]
- Thanissery, R.; Winston, J.A.; Theriot, C.M. Inhibition of Spore Germination, Growth, and Toxin Activity of Clinically Relevant C. Difficile Strains by Gut Microbiota Derived Secondary Bile Acids. Anaerobe 2017, 45, 86–100. [Google Scholar] [CrossRef]
- Rousseau, C.; Levenez, F.; Fouqueray, C.; Doré, J.; Collignon, A.; Lepage, P. Clostridium Difficile Colonization in Early Infancy Is Accompanied by Changes in Intestinal Microbiota Composition. J. Clin. Microbiol. 2020, 49, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Weese, J.S.; Finley, R.; Reid-Smith, R.R.; Janecko, N.; Rousseau, J. Evaluation of Clostridium Difficile in Dogs and the Household Environment. Epidemiol. Infect. 2010, 138, 1100–1104. [Google Scholar] [CrossRef] [PubMed]
- Honneffer, J.B.; Steiner, J.M.; Lidbury, J.A.; Suchodolski, J.S. Variation of the Microbiota and Metabolome along the Canine Gastrointestinal Tract. Metabolomics 2017, 13, 26. [Google Scholar] [CrossRef]
- Herstad, K.M.V.; Gajardo, K.; Bakke, A.M.; Moe, L.; Ludvigsen, J.; Rudi, K.; Rud, I.; Sekelja, M.; Skancke, E. A Diet Change from Dry Food to Beef Induces Reversible Changes on the Faecal Microbiota in Healthy, Adult Client-Owned Dogs. BMC Vet. Res. 2017, 13, 147. [Google Scholar] [CrossRef] [PubMed]
- Pilla, R.; Suchodolski, J.S. The Role of the Canine Gut Microbiome and Metabolome in Health and Gastrointestinal Disease. Front. Vet. Sci. 2020, 6, 498. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of Diet on the Gut Microbiome and Implications for Human Health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Jergens, A.E.; Heilmann, R.M. Canine Chronic Enteropathy—Current State-of-the-Art and Emerging Concepts. Front. Vet. Sci. 2022, 9, 923013. [Google Scholar] [CrossRef]
- Félix, A.P.; Souza, C.M.M.; de Oliveira, S.G. Biomarkers of Gastrointestinal Functionality in Dogs: A Systematic Review and Meta-Analysis. Anim. Feed. Sci. Technol. 2022, 283, 115183. [Google Scholar] [CrossRef]
- Rudinsky, A.J.; Howard, J.P.; Bishop, M.A.; Sherding, R.G.; Parker, V.J.; Gilor, C. Dietary Management of Presumptive Protein-Losing Enteropathy in Yorkshire Terriers. J. Small Anim. Pract. 2017, 58, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Allenspach, K.; Wieland, B.; Gröne, A.; Gaschen, F. Chronic Enteropathies in Dogs: Evaluation of Risk Factors for Negative Outcome. J. Vet. Intern. Med. 2007, 21, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y.L.; Ferrell, J.M. Bile Acid Receptors FXR and TGR5 Signaling in Fatty Liver Diseases and Therapy. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G554–G573. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhu, C.; Shao, L.; Ye, J.; Shen, Y.; Ren, Y. Role of Bile Acids in Dysbiosis and Treatment of Nonalcoholic Fatty Liver Disease. Mediat. Inflamm. 2019, 2019, e7659509. [Google Scholar] [CrossRef] [PubMed]
- Evangelakos, I.; Heeren, J.; Verkade, E.; Kuipers, F. Role of Bile Acids in Inflammatory Liver Diseases. Semin. Immunopathol. 2021, 43, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Andreu-Sánchez, S.; Kuipers, F.; Fu, J. Gut Microbiome and Bile Acids in Obesity-Related Diseases. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101493. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.-K.; Lee, S.-H.; Kim, S.J.; Jo, M.-J.; Kumar, H.; Han, I.-B.; Sohn, S. Anti-Inflammatory Effects of Ursodeoxycholic Acid by Lipopolysaccharide-Stimulated Inflammatory Responses in RAW 264.7 Macrophages. PLoS ONE 2017, 12, e0180673. [Google Scholar] [CrossRef] [PubMed]
- O’Kell, A.L.; Garrett, T.J.; Wasserfall, C.; Atkinson, M.A. Untargeted Metabolomic Analysis in Naturally Occurring Canine Diabetes Mellitus Identifies Similarities to Human Type 1 Diabetes. Sci. Rep. 2017, 7, 9467. [Google Scholar] [CrossRef]
- Ferrell, J.M.; Chiang, J.Y.L. Understanding Bile Acid Signaling in Diabetes: From Pathophysiology to Therapeutic Targets. Diabetes Metab. J. 2019, 43, 257–272. [Google Scholar] [CrossRef]
- Rainer, P.P.; Primessnig, U.; Harenkamp, S.; Doleschal, B.; Wallner, M.; Fauler, G.; Stojakovic, T.; Wachter, R.; Yates, A.; Groschner, K.; et al. Bile Acids Induce Arrhythmias in Human Atrial Myocardium—Implications for Altered Serum Bile Acid Composition in Patients with Atrial Fibrillation. Heart 2013, 99, 1685–1692. [Google Scholar] [CrossRef]
- Vasavan, T.; Ferraro, E.; Ibrahim, E.; Dixon, P.; Gorelik, J.; Williamson, C. Heart and Bile Acids–Clinical Consequences of Altered Bile Acid Metabolism. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2018, 1864, 1345–1355. [Google Scholar] [CrossRef]
- Ephraim, E.; Jewell, D.E. Effect of Nutrition on Age-Related Metabolic Markers and the Gut Microbiota in Cats. Microorganisms 2021, 9, 2430. [Google Scholar] [CrossRef]
- Jackson, M.I.; Waldy, C.; Jewell, D.E. Dietary Resistant Starch Preserved through Mild Extrusion of Grain Alters Fecal Microbiome Metabolism of Dietary Macronutrients While Increasing Immunoglobulin A in the Cat. PLoS ONE 2020, 15, e0241037. [Google Scholar] [CrossRef]
- Anantharaman-Barr, G.; Ballèvre, O.; Gicquello, P.; Bracco-Hammer, I.; Vuichoud, J.; Montigon, F.; Fern, E. Fecal Bile Acid Excretion and Taurine Status in Cats Fed Canned and Dry Diets. J. Nutr. 1994, 124, 2546S–2551S. [Google Scholar] [CrossRef]
- Whittemore, J.C.; Stokes, J.E.; Laia, N.L.; Price, J.M.; Suchodolski, J.S. Short and Long-Term Effects of a Synbiotic on Clinical Signs, the Fecal Microbiome, and Metabolomic Profiles in Healthy Research Cats Receiving Clindamycin: A Randomized, Controlled Trial. PeerJ 2018, 6, e5130. [Google Scholar] [CrossRef] [PubMed]
- Whittemore, J.C.; Stokes, J.E.; Price, J.M.; Suchodolski, J.S. Effects of a Synbiotic on the Fecal Microbiome and Metabolomic Profiles of Healthy Research Cats Administered Clindamycin: A Randomized, Controlled Trial. Gut Microbes 2019, 10, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Stavroulaki, E.M.; Suchodolski, J.S.; Pilla, R.; Fosgate, G.T.; Sung, C.-H.; Lidbury, J.; Steiner, J.M.; Xenoulis, P.G. The Serum and Fecal Metabolomic Profiles of Growing Kittens Treated with Amoxicillin/Clavulanic Acid or Doxycycline. Animals 2022, 12, 330. [Google Scholar] [CrossRef] [PubMed]
- Summers, S.; Quimby, J.; Winston, J. Fecal primary and secondary bile acids in cats with chronic kidney disease. 2020 ACVIM Forum On Demand Research Abstract Program. J. Vet. Intern. Med. 2020, 34, 2830–2989. [Google Scholar] [CrossRef]
- Sung, C.; Marsilio, S.; Chow, B.; Zornow, K.; Slovak, J.; Pilla, R.; Lidbury, J.; Steiner, J.; Hill, S.; Suchodolski, J. Fecal Bile Acids Profiles in Cats with Chronic Enteropathy. 2022 ACVIM Forum Research Abstract Program. J. Vet. Intern. Med. 2022, 36, 2282–2454. [Google Scholar] [CrossRef]
- Pilla, R.; Suchodolski, J.S. The Gut Microbiome of Dogs and Cats, and the Influence of Diet. Vet. Clin. Small Anim. Pract. 2021, 51, 605–621. [Google Scholar] [CrossRef]
- Doden, H.; Sallam, L.A.; Devendran, S.; Ly, L.; Doden, G.; Daniel, S.L.; Alves, J.M.P.; Ridlon, J.M. Metabolism of Oxo-Bile Acids and Characterization of Recombinant 12α-Hydroxysteroid Dehydrogenases from Bile Acid 7α-Dehydroxylating Human Gut Bacteria. Appl. Environ. Microbiol. 2018, 84, e00235-18. [Google Scholar] [CrossRef]
- Hall, J.A.; Jewell, D.E.; Ephraim, E. Changes in the Fecal Metabolome Are Associated with Feeding Fiber Not Health Status in Cats with Chronic Kidney Disease. Metabolites 2020, 10, 281. [Google Scholar] [CrossRef]
- Stavroulaki, E.M.; Suchodolski, J.S.; Pilla, R.; Fosgate, G.T.; Sung, C.-H.; Lidbury, J.A.; Steiner, J.M.; Xenoulis, P.G. Short- and Long-Term Effects of Amoxicillin/Clavulanic Acid or Doxycycline on the Gastrointestinal Microbiome of Growing Cats. PLoS ONE 2021, 16, e0253031. [Google Scholar] [CrossRef] [PubMed]
- Deusch, O.; O’Flynn, C.; Colyer, A.; Swanson, K.S.; Allaway, D.; Morris, P. A Longitudinal Study of the Feline Faecal Microbiome Identifies Changes into Early Adulthood Irrespective of Sexual Development. PLoS ONE 2015, 10, e0144881. [Google Scholar] [CrossRef] [PubMed]
- Marsilio, S.; Chow, B.; Hill, S.L.; Ackermann, M.R.; Estep, J.S.; Sarawichitr, B.; Pilla, R.; Lidbury, J.A.; Steiner, J.M.; Suchodolski, J.S. Untargeted Metabolomic Analysis in Cats with Naturally Occurring Inflammatory Bowel Disease and Alimentary Small Cell Lymphoma. Sci. Rep. 2021, 11, 9198. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Wang, W.; Garey, K.W. Heterogeneity and Lyophilization Comparison of Stool Processing for Gastrointestinal Bile Acid Measurement by LC-MS/MS. J. Chromatogr. B 2023, 1214, 123569. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rowe, J.C.; Winston, J.A. Collaborative Metabolism: Gut Microbes Play a Key Role in Canine and Feline Bile Acid Metabolism. Vet. Sci. 2024, 11, 94. https://doi.org/10.3390/vetsci11020094
Rowe JC, Winston JA. Collaborative Metabolism: Gut Microbes Play a Key Role in Canine and Feline Bile Acid Metabolism. Veterinary Sciences. 2024; 11(2):94. https://doi.org/10.3390/vetsci11020094
Chicago/Turabian StyleRowe, John C., and Jenessa A. Winston. 2024. "Collaborative Metabolism: Gut Microbes Play a Key Role in Canine and Feline Bile Acid Metabolism" Veterinary Sciences 11, no. 2: 94. https://doi.org/10.3390/vetsci11020094