
The Impact of the Virulence of Pseudomonas aeruginosa Isolated from Dogs
 ,
,  , ,
, ,  , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and Bacterial Isolates
2.2. Antimicrobial Susceptibility Testing
2.3. Biofilm Formation and Biomass Quantification
2.4. DNA Extraction
2.5. Antimicrobial Resistance and Virulence Genes
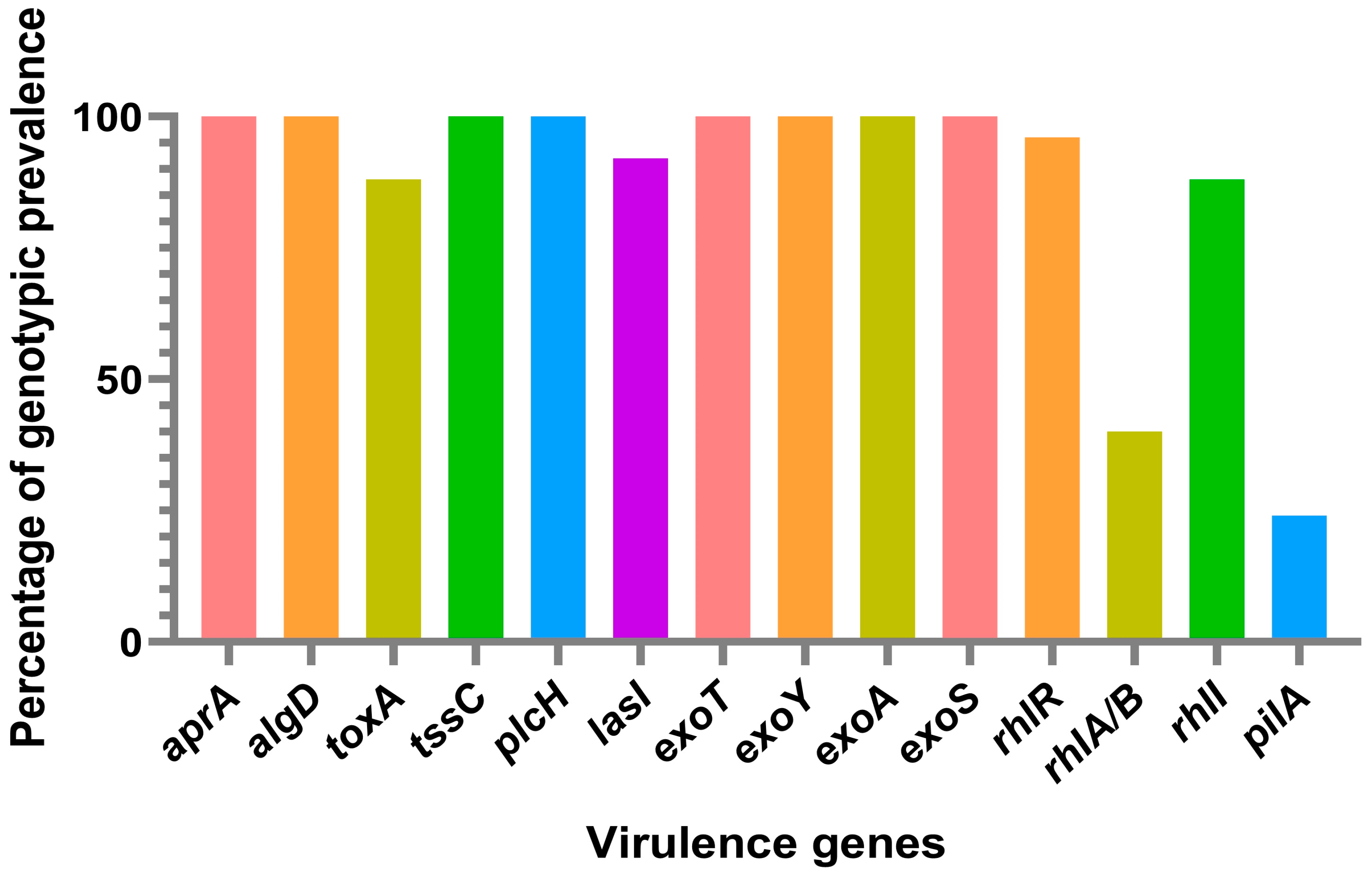
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, T.; Hebraud, M.; Dapkevicius, M.; Maltez, L.; Pereira, J.E.; Capita, R.; Alonso-Calleja, C.; Igrejas, G.; Poeta, P. Genomic and Metabolic Characteristics of the Pathogenicity in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 12892. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.; Gdynia, A.; Tielen, P.; Rosenau, F.; Jaeger, K.E. The autotransporter esterase EstA of Pseudomonas aeruginosa is required for rhamnolipid production, cell motility, and biofilm formation. J. Bacteriol. 2007, 189, 6695–6703. [Google Scholar] [CrossRef] [PubMed]
- Rybtke, M.; Berthelsen, J.; Yang, L.; Hoiby, N.; Givskov, M.; Tolker-Nielsen, T. The LapG protein plays a role in Pseudomonas aeruginosa biofilm formation by controlling the presence of the CdrA adhesin on the cell surface. Microbiologyopen 2015, 4, 917–930. [Google Scholar] [CrossRef]
- Llor, C.; Bjerrum, L. Background for different use of antibiotics in different countries. Clin. Infect. Dis. 2005, 40, 333. [Google Scholar] [CrossRef]
- Taylor, P.K.; Yeung, A.T.; Hancock, R.E. Antibiotic resistance in Pseudomonas aeruginosa biofilms: Towards the development of novel anti-biofilm therapies. J. Biotechnol. 2014, 191, 121–130. [Google Scholar] [CrossRef]
- Hoiby, N.; Ciofu, O.; Johansen, H.K.; Song, Z.J.; Moser, C.; Jensen, P.O.; Molin, S.; Givskov, M.; Tolker-Nielsen, T.; Bjarnsholt, T. The clinical impact of bacterial biofilms. Int. J. Oral Sci. 2011, 3, 55–65. [Google Scholar] [CrossRef]
- Ma, Q.; Zhai, Y.; Schneider, J.C.; Ramseier, T.M.; Saier, M.H., Jr. Protein secretion systems of Pseudomonas aeruginosa and P fluorescens. Biochim. Biophys. Acta (BBA)-Biomembr. 2003, 1611, 223–233. [Google Scholar] [CrossRef]
- Manago, A.; Becker, K.A.; Carpinteiro, A.; Wilker, B.; Soddemann, M.; Seitz, A.P.; Edwards, M.J.; Grassme, H.; Szabo, I.; Gulbins, E. Pseudomonas aeruginosa pyocyanin induces neutrophil death via mitochondrial reactive oxygen species and mitochondrial acid sphingomyelinase. Antioxid. Redox Signal. 2015, 22, 1097–1110. [Google Scholar] [CrossRef]
- Hassett, D.J.; Charniga, L.; Bean, K.; Ohman, D.E.; Cohen, M.S. Response of Pseudomonas aeruginosa to pyocyanin: Mechanisms of resistance, antioxidant defenses, and demonstration of a manganese-cofactored superoxide dismutase. Infect. Immun. 1992, 60, 328–336. [Google Scholar] [CrossRef]
- Bleves, S.; Viarre, V.; Salacha, R.; Michel, G.P.; Filloux, A.; Voulhoux, R. Protein secretion systems in Pseudomonas aeruginosa: A wealth of pathogenic weapons. Int. J. Med. Microbiol. 2010, 300, 534–543. [Google Scholar] [CrossRef] [PubMed]
- DeBritto, S.; Gajbar, T.D.; Satapute, P.; Sundaram, L.; Lakshmikantha, R.Y.; Jogaiah, S.; Ito, S.I. Isolation and characterization of nutrient dependent pyocyanin from Pseudomonas aeruginosa and its dye and agrochemical properties. Sci. Rep. 2020, 10, 1542. [Google Scholar] [CrossRef] [PubMed]
- Dear, J.D. Bacterial Pneumonia in Dogs and Cats: An Update. Vet. Clin. Small Anim. Pract. 2020, 50, 447–465. [Google Scholar] [CrossRef]
- van Knapen, F.; Overgaauw, P. Dogs and Transmission of Infection to Man,“Respected Member of the Family?”. In Zoonoses-Infections Affecting Humans and Animals: Focus on Public Health Aspects; Springer: Dordrecht, The Netherlands, 2015; pp. 575–585. [Google Scholar]
- Haenni, M.; Hocquet, D.; Ponsin, C.; Cholley, P.; Guyeux, C.; Madec, J.Y.; Bertrand, X. Population structure and antimicrobial susceptibility of Pseudomonas aeruginosa from animal infections in France. BMC Vet. Res. 2015, 11, 9. [Google Scholar] [CrossRef]
- Sabath, L.D.; Jago, M.; Abraham, E.P. Cephalosporinase and penicillinase activities of a beta-lactamase from Pseudomonas pyocyanea. Biochem. J. 1965, 96, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Kimura, N.; Mima, T.; Mizushima, T.; Tsuchiya, T. Roles of MexXY- and MexAB-multidrug efflux pumps in intrinsic multidrug resistance of Pseudomonas aeruginosa PAO1. J. Gen. Appl. Microbiol. 2001, 47, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Johnson, N.V.; Kreamer, N.N.K.; Barnes, S.W.; Walker, J.R.; Woods, A.L.; Six, D.A.; Dean, C.R. Defects in Efflux (oprM), beta-Lactamase (ampC), and Lipopolysaccharide Transport (lptE) Genes Mediate Antibiotic Hypersusceptibility of Pseudomonas aeruginosa Strain Z61. Antimicrob. Agents Chemother. 2019, 63, e00784-19. [Google Scholar] [CrossRef] [PubMed]
- Sindeldecker, D.; Stoodley, P. The many antibiotic resistance and tolerance strategies of Pseudomonas aeruginosa. Biofilm 2021, 3, 100056. [Google Scholar] [CrossRef]
- Poole, K. Aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2005, 49, 479–487. [Google Scholar] [CrossRef]
- Meletis, G.; Exindari, M.; Vavatsi, N.; Sofianou, D.; Diza, E. Mechanisms responsible for the emergence of carbapenem resistance in Pseudomonas aeruginosa. Hippokratia 2012, 16, 303–307. [Google Scholar]
- Jeannot, K.; Hagart, K.; Dortet, L.; Kostrzewa, M.; Filloux, A.; Plesiat, P.; Larrouy-Maumus, G. Detection of Colistin Resistance in Pseudomonas aeruginosa Using the MALDIxin Test on the Routine MALDI Biotyper Sirius Mass Spectrometer. Front. Microbiol. 2021, 12, 725383. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Han, M.L.; Zhao, J.; Zhu, Y.; Rao, G.; Forrest, A.; Song, J.; Kaye, K.S.; Hertzog, P.; Purcell, A.; et al. Synergistic Combination of Polymyxin B and Enrofloxacin Induced Metabolic Perturbations in Extensive Drug-Resistant Pseudomonas aeruginosa. Front. Pharmacol. 2019, 10, 1146. [Google Scholar] [CrossRef] [PubMed]
- Karruli, A.; Catalini, C.; D’Amore, C.; Foglia, F.; Mari, F.; Harxhi, A.; Galdiero, M.; Durante-Mangoni, E. Evidence-Based Treatment of Pseudomonas aeruginosa Infections: A Critical Reappraisal. Antibiotics 2023, 12, 399. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Animal Health and Welfare (AHAW); Nielsen, S.S.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortazar, C.; Herskin, M.; et al. Assessment of listing and categorisation of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Antimicrobial-resistant Pseudomonas aeruginosa in dogs and cats. EFSA J. 2022, 20, e07310. [Google Scholar] [CrossRef]
- Guardabassi, L.; Schwarz, S.; Lloyd, D.H. Pet animals as reservoirs of antimicrobial-resistant bacteria. J. Antimicrob. Chemother. 2004, 54, 321–332. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, e2437. [Google Scholar] [CrossRef]
- Dashti, A.A.; Jadaon, M.M.; Abdulsamad, A.M.; Dashti, H.M. Heat treatment of bacteria: A simple method of DNA extraction for molecular techniques. Kuwait Med. J. 2009, 41, 117–122. [Google Scholar]
- Amoon, R.H.; Abdallha, A.H.; Sharif, A.O.; Moglad, E.H.; Altyb, H.N.; Elzaki, S.G.; Salih, M.A. Molecular characterization of Pseudomonas aeruginosa isolates from Sudanese patients: A cross-sectional study. F1000Research 2018, 7, 1135. [Google Scholar] [CrossRef]
- Belaaouaj, A.; Lapoumeroulie, C.; Caniça, M.M.; Vedel, G.; Névot, P.; Krishnamoorthy, R.; Paul, G. Nucleotide sequences of the genes coding for the TEM-like β-lactamases IRT-1 and IRT-2 (formerly called TRI-1 and TRI-2). FEMS Microbiol. Lett. 1994, 120, 75–80. [Google Scholar]
- Steward, C.D.; Rasheed, J.K.; Hubert, S.K.; Biddle, J.W.; Raney, P.M.; Anderson, G.J.; Williams, P.P.; Brittain, K.L.; Oliver, A.; McGowan, J.E., Jr. Characterization of clinical isolates of Klebsiella pneumoniae from 19 laboratories using the National Committee for Clinical Laboratory Standards extended-spectrum β-lactamase detection methods. J. Clin. Microbiol. 2001, 39, 2864–2872. [Google Scholar] [CrossRef]
- Pagani, L.; Dell’Amico, E.; Migliavacca, R.; D’Andrea, M.M.; Giacobone, E.; Amicosante, G.; Romero, E.; Rossolini, G.M. Multiple CTX-M-type extended-spectrum β-lactamases in nosocomial isolates of Enterobacteriaceae from a hospital in northern Italy. J. Clin. Microbiol. 2003, 41, 4264–4269. [Google Scholar] [CrossRef]
- Weldhagen, G.F.; Poirel, L.; Nordmann, P. Ambler class A extended-spectrum β-lactamases in Pseudomonas aeruginosa: Novel developments and clinical impact. Antimicrob. Agents Chemother. 2003, 47, 2385–2392. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.S.; Kim, K.; Huh, J.Y.; Jung, B.; Kang, M.S.; Hong, S.G. Multiplex PCR for rapid detection of genes encoding class A carbapenemases. Ann. Lab. Med. 2012, 32, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Neyestanaki, D.K.; Mirsalehian, A.; Rezagholizadeh, F.; Jabalameli, F.; Taherikalani, M.; Emaneini, M. Determination of extended spectrum beta-lactamases, metallo-beta-lactamases and AmpC-beta-lactamases among carbapenem resistant Pseudomonas aeruginosa isolated from burn patients. Burns 2014, 40, 1556–1561. [Google Scholar] [CrossRef]
- Ellington, M.J.; Kistler, J.; Livermore, D.M.; Woodford, N. Multiplex PCR for rapid detection of genes encoding acquired metallo-β-lactamases. J. Antimicrob. Chemother. 2007, 59, 321–322. [Google Scholar] [CrossRef]
- Amudhan, M.S.; Sekar, U.; Kamalanathan, A.; Balaraman, S. blaIMP and blaVIM mediated carbapenem resistance in Pseudomonas and Acinetobacter species in India. J. Infect. Dev. Ctries. 2012, 6, 757–762. [Google Scholar] [CrossRef]
- Franco, M.R.G.; Caiaffa-Filho, H.H.; Burattini, M.N.; Rossi, F. Metallo-beta-lactamases among imipenem-resistant Pseudomonas aeruginosa in a Brazilian university hospital. Clinics 2010, 65, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Van De Klundert, J.; Vliegenthart, J. Nomenclature of aminoglycoside resistance genes: A comment. Antimicrob. Agents Chemother. 1993, 37, 927–928. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, J.; Ju, Z.; Chang, W.; Sun, S. Molecular characterization of antimicrobial resistance in Escherichia coli from rabbit farms in Tai’an, China. BioMed Res. Int. 2018, 2018, 8607647. [Google Scholar] [CrossRef] [PubMed]
- Terzi, H.A.; Kulah, C.; Atasoy, A.R.; Ciftci, I.H. Investigation of OprD porin protein levels in carbapenem-resistant Pseudomonas aeruginosa isolates. Jundishapur J. Microbiol. 2015, 8, e25952. [Google Scholar]
- Fazeli, N.; Momtaz, H. Virulence gene profiles of multidrug-resistant Pseudomonas aeruginosa isolated from Iranian hospital infections. Iran. Red Crescent Med. J. 2014, 16, e15722. [Google Scholar] [CrossRef] [PubMed]
- Petit, S.M.-C.; Lavenir, R.; Colinon-Dupuich, C.; Boukerb, A.M.; Cholley, P.; Bertrand, X.; Freney, J.; Doléans-Jordheim, A.; Nazaret, S.; Laurent, F. Lagooning of wastewaters favors dissemination of clinically relevant Pseudomonas aeruginosa. Res. Microbiol. 2013, 164, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Khattab, M.; Nour, M.; ElSheshtawy, N.M. Genetic identification of Pseudomonas aeruginosa virulence genes among different isolates. J. Microb. Biochem. Technol. 2015, 7, 274–277. [Google Scholar]
- Zhang, L.; Hinz, A.J.; Nadeau, J.-P.; Mah, T.-F. Pseudomonas aeruginosa tssC1 links type VI secretion and biofilm-specific antibiotic resistance. J. Bacteriol. 2011, 193, 5510–5513. [Google Scholar] [CrossRef] [PubMed]
- Schaber, J.A.; Carty, N.L.; McDonald, N.A.; Graham, E.D.; Cheluvappa, R.; Griswold, J.A.; Hamood, A.N. Analysis of quorum sensing-deficient clinical isolates of Pseudomonas aeruginosa. J. Med. Microbiol. 2004, 53, 841–853. [Google Scholar] [CrossRef]
- Faraji, F.; Mahzounieh, M.; Ebrahimi, A.; Fallah, F.; Teymournejad, O.; Lajevardi, B. Molecular detection of virulence genes in Pseudomonas aeruginosa isolated from children with Cystic Fibrosis and burn wounds in Iran. Microb. Pathog. 2016, 99, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Ajayi, T.; Allmond, L.R.; Sawa, T.; Wiener-Kronish, J.P. Single-nucleotide-polymorphism mapping of the Pseudomonas aeruginosa type III secretion toxins for development of a diagnostic multiplex PCR system. J. Clin. Microbiol. 2003, 41, 3526–3531. [Google Scholar] [CrossRef]
- Zhu, H.; Bandara, R.; Conibear, T.C.; Thuruthyil, S.J.; Rice, S.A.; Kjelleberg, S.; Givskov, M.; Willcox, M.D. Pseudomonas aeruginosa with lasI quorum-sensing deficiency during corneal infection. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1897–1903. [Google Scholar] [CrossRef]
- MSD Animal Health Portugal. 2023. Available online: https://www.msd-animal-health.pt/ (accessed on 20 March 2023).
- Harada, K.; Arima, S.; Niina, A.; Kataoka, Y.; Takahashi, T. Characterization of Pseudomonas aeruginosa isolates from dogs and cats in Japan: Current status of antimicrobial resistance and prevailing resistance mechanisms. Microbiol. Immunol. 2012, 56, 123–127. [Google Scholar] [CrossRef]
- Shahini, N.; Shahini, N.; Ala, S. Determining of resistance and sensitivity of Pseudomonas aeruginosa in Iran in 2010–2011. Res. Pharm. Sci. 2012, 7, 884. [Google Scholar]
- Kocsis, B.; Gulyas, D.; Szabo, D. Diversity and Distribution of Resistance Markers in Pseudomonas aeruginosa International High-Risk Clones. Microorganisms 2021, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Hattab, J.; Mosca, F.; Di, C.E.F.; Aste, G.; Marruchella, G.; Guardiani, P.; Tiscar, P.G. Occurrence, antimicrobial susceptibility, and pathogenic factors of Pseudomonas aeruginosa in canine clinical samples. Vet. World 2021, 14, 978. [Google Scholar] [CrossRef] [PubMed]
- Mader, R.; Damborg, P.; Amat, J.-P.; Bengtsson, B.; Bourély, C.; Broens, E.M.; Busani, L.; Crespo-Robledo, P.; Filippitzi, M.-E.; Fitzgerald, W. Building the European antimicrobial resistance surveillance network in veterinary medicine (EARS-Vet). Eurosurveillance 2021, 26, 2001359. [Google Scholar] [CrossRef]
- Poonsuk, K.; Chuanchuen, R. Contribution of the MexXY multidrug efflux pump and other chromosomal mechanisms on aminoglycoside resistance in Pseudomonas aeruginosa isolates from canine and feline infections. J. Vet. Med. Sci. 2012, 74, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Haenni, M.; Bour, M.; Châtre, P.; Madec, J.-Y.; Plésiat, P.; Jeannot, K. Resistance of animal strains of Pseudomonas aeruginosa to carbapenems. Front. Microbiol. 2017, 8, 1847. [Google Scholar] [CrossRef]
- Li, H.; Luo, Y.-F.; Williams, B.J.; Blackwell, T.S.; Xie, C.-M. Structure and function of OprD protein in Pseudomonas aeruginosa: From antibiotic resistance to novel therapies. Int. J. Med. Microbiol. 2012, 302, 63–68. [Google Scholar] [CrossRef]
- Tamber, S.; Ochs, M.M.; Hancock, R.E. Role of the novel OprD family of porins in nutrient uptake in Pseudomonas aeruginosa. J. Bacteriol. 2006, 188, 45–54. [Google Scholar] [CrossRef]
- Davies, J.C.; Alton, E.; Simbo, A.; Murphy, R.; Seth, I.; Williams, K.; Somerville, M.; Jolly, L.; Morant, S.; Guest, C. Training dogs to differentiate Pseudomonas aeruginosa from other cystic fibrosis bacterial pathogens: Not to be sniffed at? Eur. Respir. J. 2019, 54, 1900970. [Google Scholar] [CrossRef]
- Ledbetter, E.C.; Mun, J.J.; Kowbel, D.; Fleiszig, S.M. Pathogenic phenotype and genotype of Pseudomonas aeruginosa isolates from spontaneous canine ocular infections. Investig. Ophthalmol. Vis. Sci. 2009, 50, 729–736. [Google Scholar] [CrossRef]
- Pottier, M.; Castagnet, S.; Gravey, F.; Leduc, G.; Sévin, C.; Petry, S.; Giard, J.-C.; Le Hello, S.; Léon, A. Antimicrobial Resistance and Genetic Diversity of Pseudomonas aeruginosa Strains Isolated from Equine and Other Veterinary Samples. Pathogens 2022, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Koo, S.H. Epidemiology, molecular characteristics, and virulence factors of carbapenem-resistant Pseudomonas aeruginosa isolated from patients with urinary tract infections. Infect. Drug Resist. 2022, 15, 141–151. [Google Scholar] [CrossRef]
- O’Connor, K.; Zhao, C.Y.; Diggle, S.P. Frequency of quorum sensing mutations in Pseudomonas aeruginosa strains isolated from different environments. bioRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Abidi, S.H.; Sherwani, S.K.; Siddiqui, T.R.; Bashir, A.; Kazmi, S.U. Drug resistance profile and biofilm forming potential of Pseudomonas aeruginosa isolated from contact lenses in Karachi-Pakistan. BMC Ophthalmol. 2013, 13, 57. [Google Scholar] [CrossRef] [PubMed]
- Musken, M.; Di Fiore, S.; Romling, U.; Haussler, S. A 96-well-plate-based optical method for the quantitative and qualitative evaluation of Pseudomonas aeruginosa biofilm formation and its application to susceptibility testing. Nat. Protoc. 2010, 5, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Plokarz, D.; Czopowicz, M.; Bierowiec, K.; Rypula, K. Virulence Genes as Markers for Pseudomonas aeruginosa Biofilm Formation in Dogs and Cats. Animals 2022, 12, 422. [Google Scholar] [CrossRef]
- Pye, C.C.; Yu, A.A.; Weese, J.S. Evaluation of biofilm production by Pseudomonas aeruginosa from canine ears and the impact of biofilm on antimicrobial susceptibility in vitro. Vet. Dermatol. 2013, 24, 446-e99. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, P.; Wang, S.; Li, X.; Peng, L.; Fang, R.; Xiong, J.; Li, H.; Mei, C.; Gao, J. Pseudomonas aeruginosa biofilm dispersion by the mouse antimicrobial peptide CRAMP. Vet. Res. 2022, 53, 80. [Google Scholar] [CrossRef]


{kind=link}
| Isolate | Sex | Age (Year) | Collection | Sample |
|---|---|---|---|---|
| D1 | F | 15 | November | Chronic ulcerative Dermatitis |
| D2 | M | 12 | November | Urine |
| D3 | M | November | Ear exudate | |
| D4 | M | 2 | November | Ear exudate |
| D5 | F | 4 M | November | Skin exudate |
| D6 | F | 10 | November | Ear exudate |
| D7 | F | 15 | November | Ear exudate |
| D8 | M | - | November | Skin exudate |
| D9 | F | - | November | Ear exudate |
| D10 | F | 2 | December | Vaginal exudate |
| D11 | F | 7 | December | Skin exudate |
| D12 | F | 7 | December | Urine |
| D13 | M | - | December | Ear exudate |
| D14 | M | 8 | December | Lip injury |
| D15 | F | December | Ear exudate | |
| D16 | M | 10 | December | Ear exudate |
| D17 | F | 6 | December | Ear exudate |
| D18 | M | 7 | December | Ear exudate |
| D19 | M | 2 | December | Ear exudate |
| D20 | M | 2 | December | Ear exudate |
| D21 | M | 8 | December | Ear exudate |
| D22 | F | 12 | December | Ear exudate |
| D23 | M | 9 | December | Ear exudate |
| D24 | M | 6 | December | Ear exudate |
| D25 | F | 12 | December | Ear exudate |
| D26 | F | 10 | December | Ear exudate |
| D27 | F | 8 | December | Ear exudate |
| Name | Sequence (5′ → 3′) | Length (bp) | References |
|---|---|---|---|
| blaTEM | F: ATTCTTGAAGACGAAAGGGC R: ACGCTCAGTGGAACGAAAAC | 1150 | [31] |
| blaSHV | F: CACTCAAGGATGTATTGTG R: TTAGCGTTGCCAGTGCTCG | 885 | [32] |
| blaCTX | F: CGATGTGCAGTACCAGTAA R: TTAGTGACCAGAATCAGCGG | 585 | [33] |
| blaPER | F: ATGAATGTCATTATAAAAGC R: AATTTGGGCTTAGGGCAGAA | 920 | [34] |
| blaSME | F: ACTTTGATGGGAGGATTGGC R: ACGAATTCGAGCATCACCAG | 551 | [35] |
| blaKPC | F: GTATCGCCGTCTAGTTCTGC R: GGTCGTGTTTCCCTTTAGCC | 638 | [36] |
| blaIMP | F: GTTTATGTTCATACTCG R: GGTTTAAAAAACAACCAC | 432 | [36] |
| blaSmp | F: AAAATCTGGGTACGCAAACG R: ACATTATCCGCTGGAACAGG | 271 | [37] |
| blaVim | F: TTTGGTCGCATATCGCAACG R: CCATTCAGCCAGATCGGCAT | 500 | [38] |
| blaVim-2 | F: AAAGTTATGCCGCACTCACC R: TGCAACTTCATGTTATGCCG | 815 | [39] |
| blaNDM | F: GGTTTGGCGATCTGGTTTTC R: CGGAATGGCTCATCACGATC | 621 | [36] |
| blaOXA | F: CCAAAGACGTGGATG R: GTTAAATTCGACCCCAAGTT | 813 | [32] |
| aac(6′)-Ie-aph(2″)-Ia | F: CCAAGAGCAATAAGGGCATA R: CACTATCATAACCACTACCG | 220 | [40] |
| aph(3′)-IIIa | F: GCCGATGTGGATTGCGAAAA R: GCTTGATCCCCAGTAAGTCA | 292 | [40] |
| aac(3)-II | F: ACTGTGATGGGATACGCGTC R: CTCCGTCAGCGTTTCAGCTA | 237 | [41] |
| aac(3)-III | F: CACAAGAACGTGGTCCGCTA R: AACAGGTAAGCATCCGCATC | 195 | [41] |
| aac(3)-IV | F: CTTCAGGATGGCAAGTTGGT R: TACTCTCGTTCTCCGCTCAT | 286 | [40] |
| ant(4′)-Ia | F: GCAAGGACCGACAACATTTC R: TGGCACAGATGGTCATAACC | 165 | [40] |
| ant(2′)-I | F: ATGTTACGCAGCAGGGCAGTCG R: CGTCAGATCAATATCATCGTGC | 188 | [41] |
| oprD | F: TCCGCAGGTAGCACTCAGTTC R: AAGCCGGATTCATAGGTGGTG | 191 | [42] |
| pilB | F: TCGAACTGATGATCGTGG R: CTTTCGGAGTGAACATCG | 408 | [43] |
| pilA | F: ACAGCATCCAACTGAGCG R: TTGACTTCCTCCAGGCTG | 1675 | [43] |
| aprA | F: ACCCTGTCCTATTCGTTCC R: GATTGCAGCGACAACTTGG | 140 | [44] |
| toxA | F: GGTAACCACGTCAGCCACAT R: TGATGTCCAGGTCATGCTTC | 352 | [45] |
| tssC | F: CTCCAACGACGCGATCAAGT R: TCGGTGTTGTTGACCAGGTA | 150 | [46] |
| plcH | F: GCACGTGGTCATCCTGATGC R: TCCGTAGGCGTCGACGTAC | 608 | [43] |
| lasA | F: GCAGCACAAAAGATCCC R: GAAATGCAGGTGCGGTC | 1075 | [43] |
| lasB | F: GGAATGAACGAAGCGTTCTC R: GGTCCAGTAGTAGCGGTTGG | 284 | [43] |
| lasR | F: CGGGTATCGTACTAGGTGCATCA R: GACGGGAAAGCCAGGAAACTT | 1100 | [44] |
| lasI | F: ATGATCGTACAAATTGGTCGGC R: GTCATGAAACCGCCAGTCG | 605 | [47] |
| exoU | F: ATGCATATCCAATCGTTG R: TCATGTGAACTCCTTATT | 2000 | [44] |
| exoS | F: CGTCGTGTTCAAGCAGATGGTGCTG R: CCGAACCGCTTCACCAGGC | 444 | [48] |
| exoA | F: GACAACGCCCTCAGCATCACCAGC R: CGCTGGCCCATTCGCTCCAGCGCT | 396 | [49] |
| exoY | F: CGGATTCTATGGCAGGGAGG R: GCCCTTGATGCACTCGACCA | 289 | [49] |
| exoT | F: AATCGCCGTCCAACTGCATGCG R: TGTTCGCCGAGGTACTGCTC | 159 | [49] |
| rhlR | F: CAATGAGGAATGACGGAGGC R: GCTTCAGATGAGGCCCAGC | 730 | [47] |
| rhlI | F: CTTGGTCATGATCGAATTGCTC R: ACGGCTGACGACCTCACAC | 625 | [47] |
| rhlA/B | F: TCATGGAATTGTCACAACCGC R: ATACGGCAAAATCATGGCAAC | 151 | [50] |
| algD | F: CGTCTGCCGCGAGATCGGCT R: GACCTCGACGGTCTTGCGGA | 313 | [43] |
| Class and⁄or Antimicrobial | Breakpoints (mm; S≥/R<) | Dogs (n = 27) | |
|---|---|---|---|
| Number of Resistant Strains | Percentage of Resistant Strains (%) | ||
| β-Lactams | |||
| Cefepime | 50/21 | 0 | |
| Ceftiofur | ** | 16 | 59 |
| Cefovecin | ** | 20 | 74 |
| Aztreonam | 50/18 | 0 | |
| Ceftazidime * | 18/15 * | 8 | 30 |
| Doripenem | 50/22 | 0 | |
| Imipenem | 50/20 | 8 | 30 |
| Meropenem | 20/14 | 1 | 4 |
| Ticarcillin-clavulanic acid | 50/18 | 1 | 4 |
| Fluoroquinolones | |||
| Ciprofloxacin | 50/26 | 2 | 7 |
| Enrofloxacin | ** | 2 | 7 |
| Marbofloxacin | ** | 1 | 4 |
| Aminoglycosides | |||
| Amikacin | 15/15 | 0 | |
| Tobramicyn | 18/18 | 0 | |
| Gentamicin | 15/15 | 2 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa, T.; Garcês, A.; Silva, A.; Lopes, R.; Alegria, N.; Hébraud, M.; Igrejas, G.; Poeta, P. The Impact of the Virulence of Pseudomonas aeruginosa Isolated from Dogs. Vet. Sci. 2023, 10, 343. https://doi.org/10.3390/vetsci10050343
de Sousa T, Garcês A, Silva A, Lopes R, Alegria N, Hébraud M, Igrejas G, Poeta P. The Impact of the Virulence of Pseudomonas aeruginosa Isolated from Dogs. Veterinary Sciences. 2023; 10(5):343. https://doi.org/10.3390/vetsci10050343
Chicago/Turabian Stylede Sousa, Telma, Andreia Garcês, Augusto Silva, Ricardo Lopes, Nuno Alegria, Michel Hébraud, Gilberto Igrejas, and Patricia Poeta. 2023. "The Impact of the Virulence of Pseudomonas aeruginosa Isolated from Dogs" Veterinary Sciences 10, no. 5: 343. https://doi.org/10.3390/vetsci10050343