
Transcriptome Profile Analysis Identifies Candidate Genes for the Melanin Pigmentation of Skin in Tengchong Snow Chickens
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experimentation Ethical Statement
2.2. Chicken, Diet, and Housing
2.3. Determination of Luminance Values and Melanin Content
2.4. Extraction of Total RNA
2.5. Construction and Sequencing of RNA-Seq Libraries
2.6. RNA-Seq Data Acquisition, Quality Control, and Processing Raw Data Quantification
2.7. Enrichment Analysis
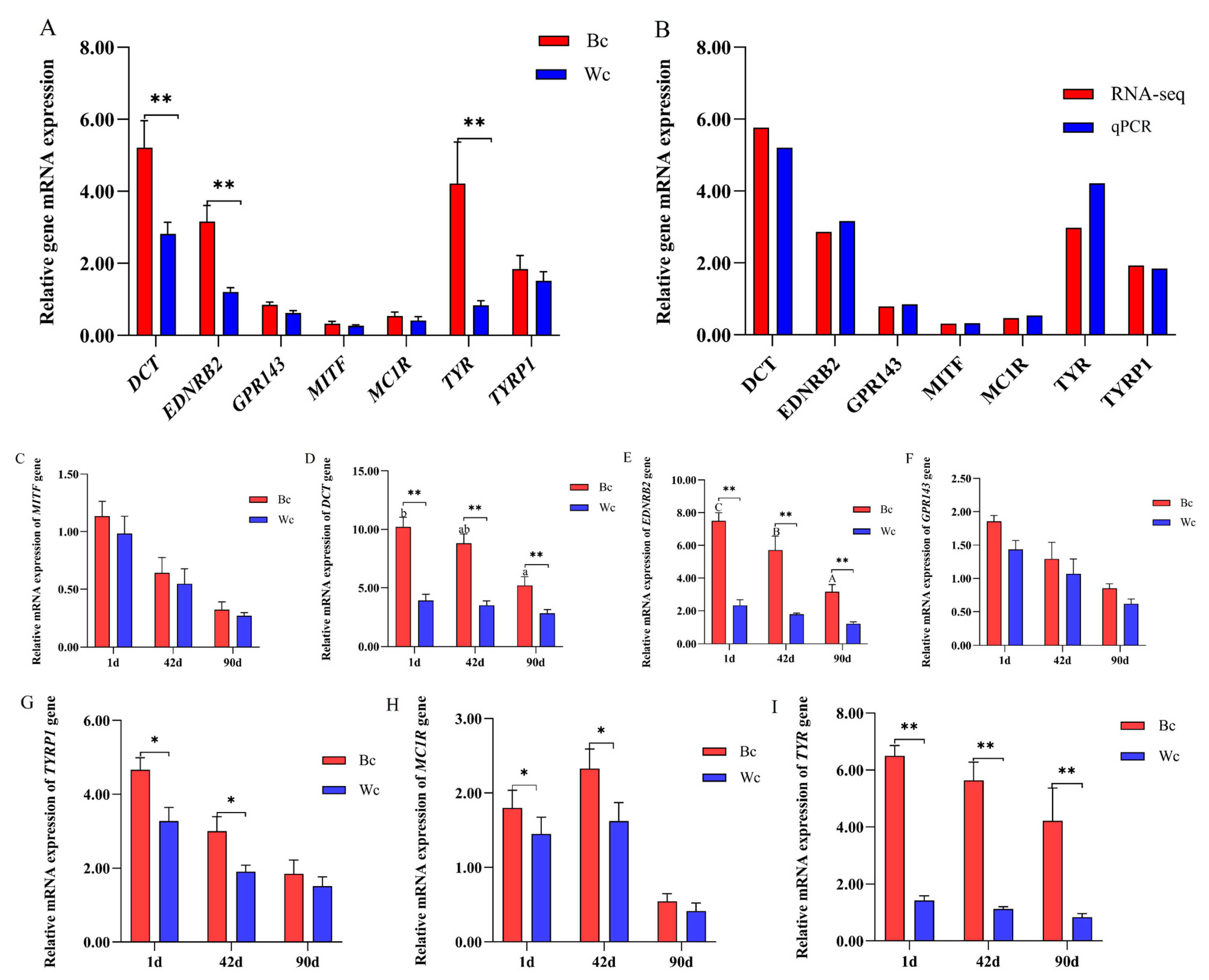
2.8. Verification of the RNA-Seq Results
2.9. Statistical Analysis
3. Results
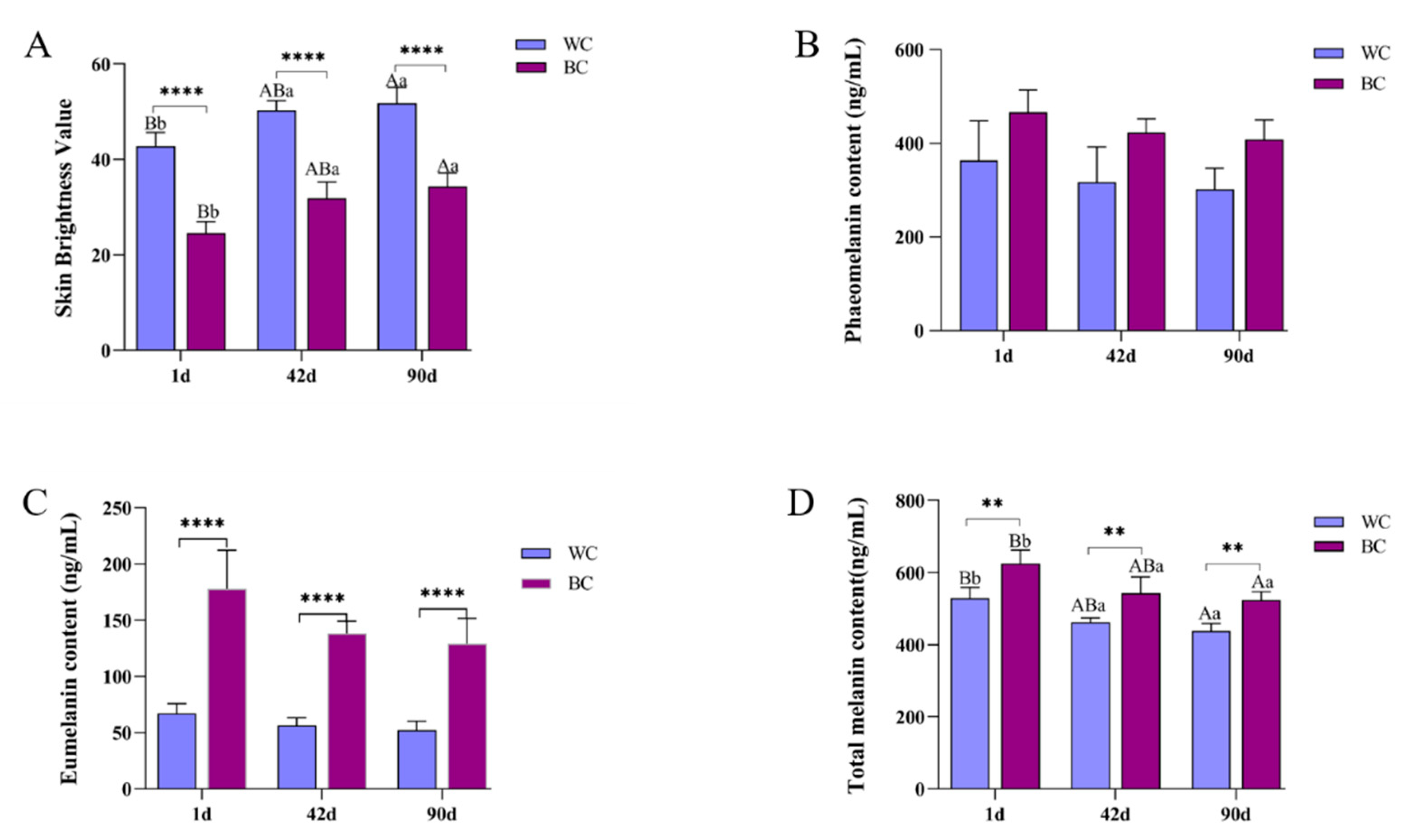
3.1. Comparative Analysis of the Luminance Value and Melanin Content
3.2. Correlation Analysis between Luminance Value and Melanin Content
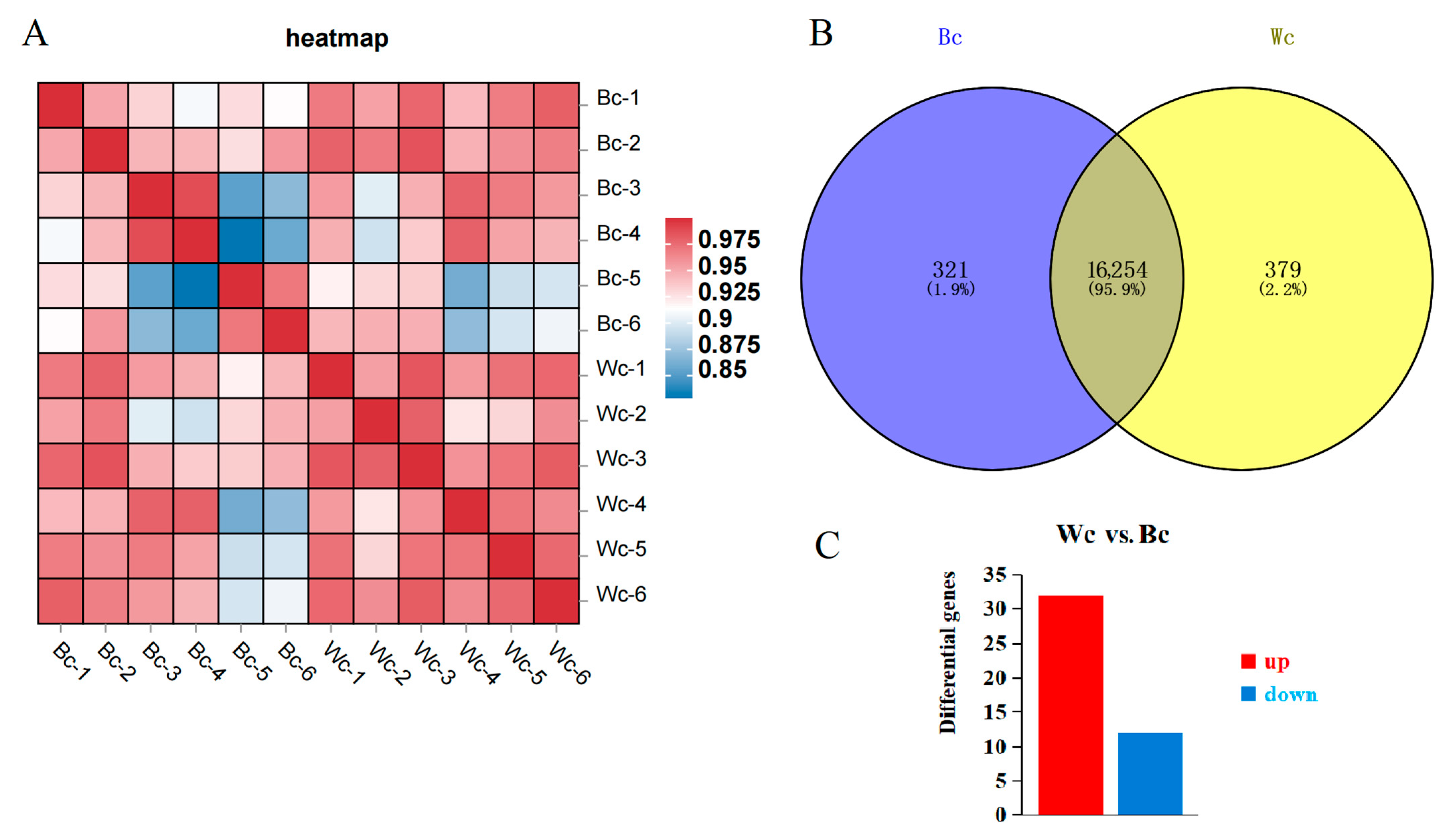
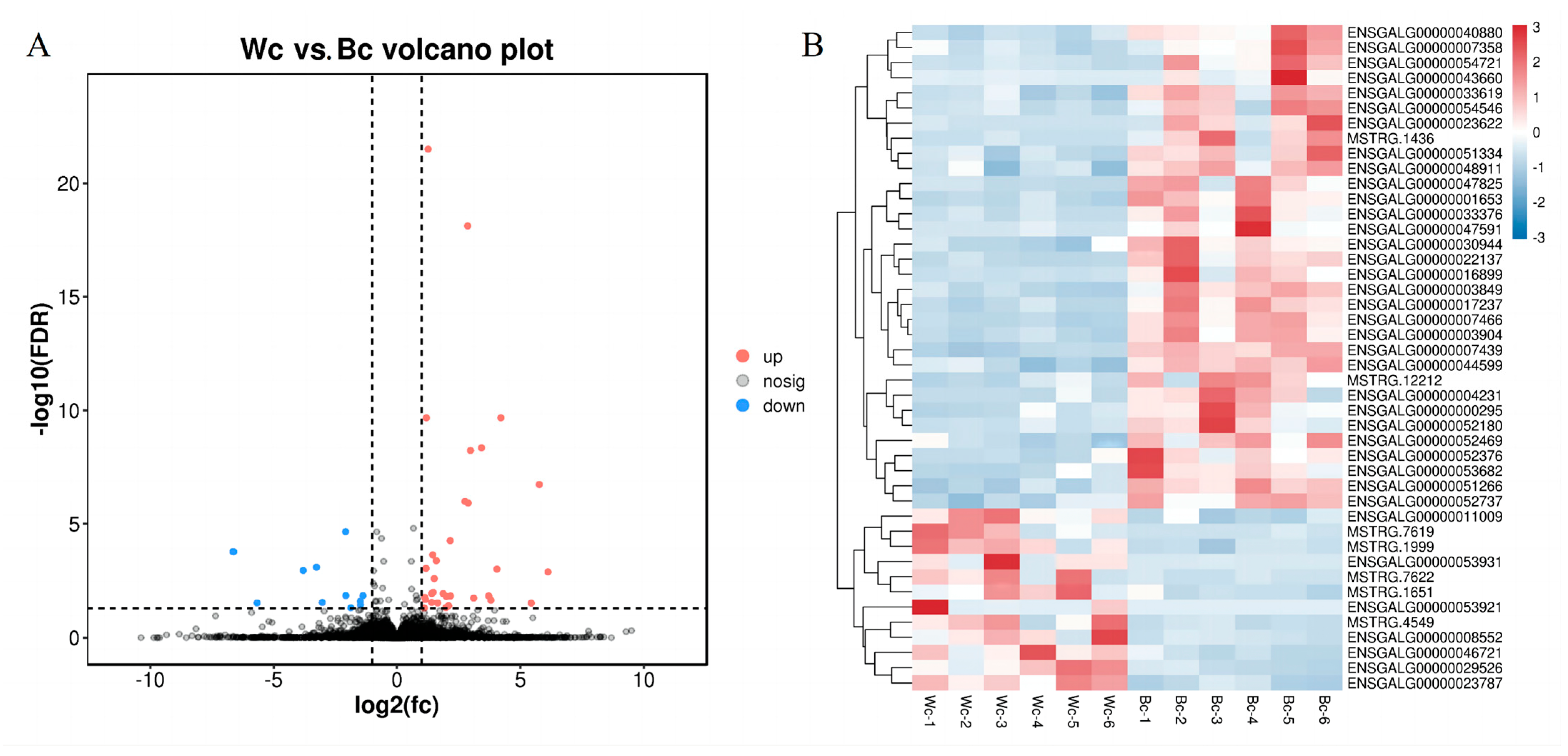
3.3. Skin Transcriptome Analysis: Differentially Expressed Genes and KEGG Enrichment Analysis
3.3.1. Functional Enrichment Analysis of Differentially Expressed Genes
3.3.2. GO Functional Enrichment Analysis
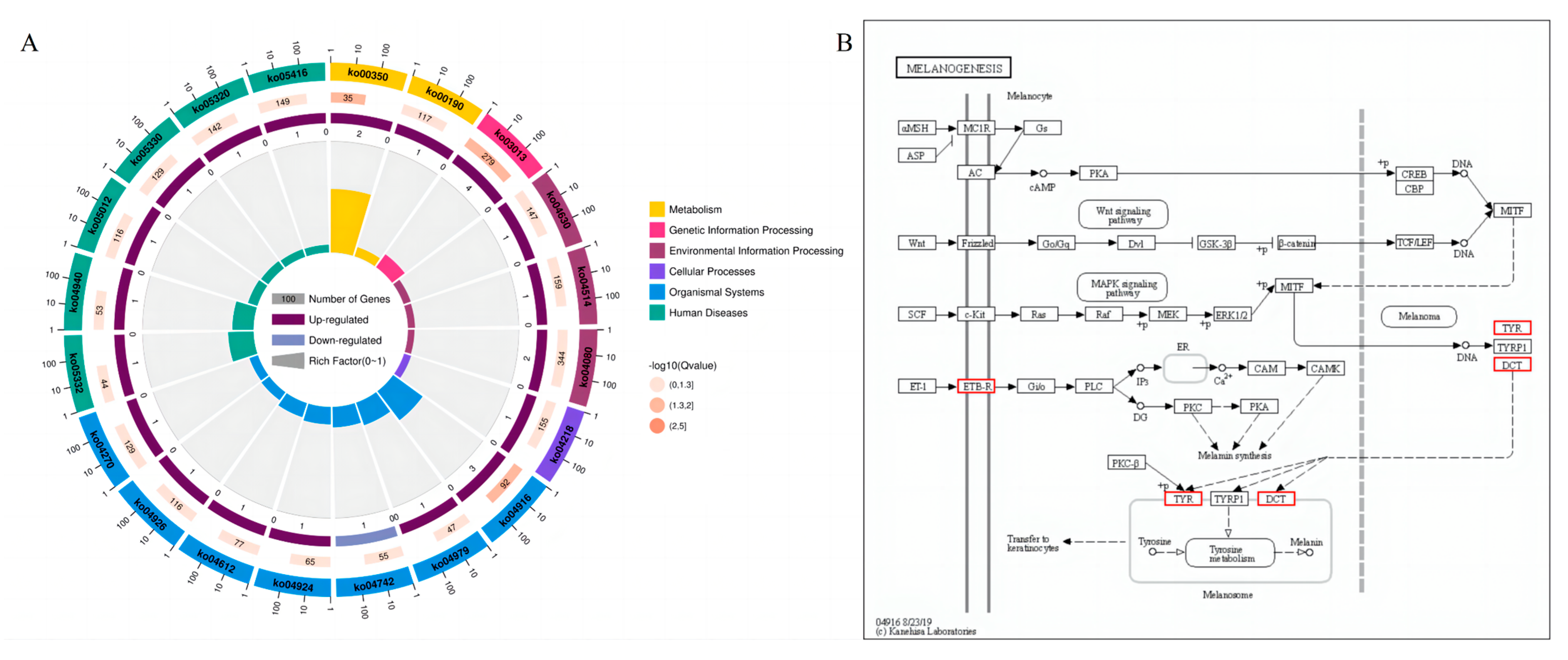
3.3.3. KEGG Pathway Analysis
3.3.4. Reactome Enrichment Analysis
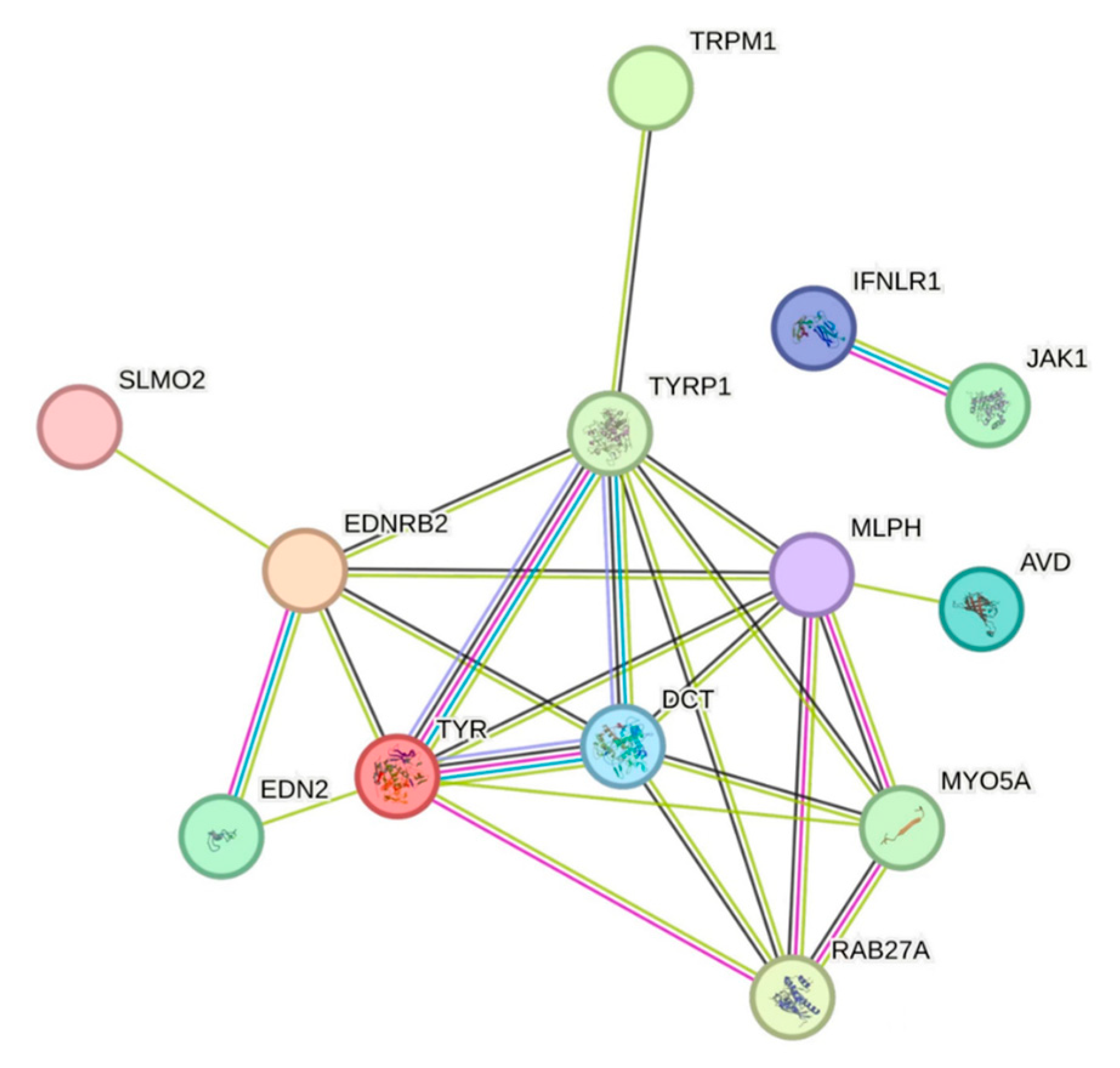
3.3.5. Protein Interaction Network Analysis
3.4. Validation of Differentially Expressed mRNA in Chicken Skin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, W.Q.; Li, H.F.; Wang, J.Y.; Shu, J.T.; Zhu, C.H.; Song, W.T.; Song, C.; Ji, G.G.; Liu, H.X. Molecular genetic diversity and maternal origin of Chinese black-bone chicken breeds. Genet. Mol. Res. 2014, 13, 3275–3282. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xie, M.; Wang, W.; Wu, H.; Fu, Z.; Lin, L. Determination of carnosine in Black-Bone Silky Fowl (Gallus gallus domesticus Brisson) and common chicken by HPLC. Eur. Food Res. Technol. 2007, 226, 311–314. [Google Scholar] [CrossRef]
- Li, D.; Wang, X.; Fu, Y.; Zhang, C.; Cao, Y.; Wang, J.; Zhang, Y.; Li, Y.; Chen, Y.; Li, Z.; et al. Transcriptome analysis of the breast muscle of Xichuan Black-Bone Chickens under tyrosine supplementation revealed the mechanism of tyrosine-induced melanin deposition. Front. Genet. 2019, 10, 457. [Google Scholar] [CrossRef] [PubMed]
- Nofsinger, J.B.; Weinert, E.E.; Simon, J.D. Establishing structure-function relationships for eumelanin. Biopolymers 2002, 67, 302–305. [Google Scholar] [CrossRef]
- Mohagheghpour, N.; Waleh, N.; Garger, S.J.; Dousman, L.; Grill, L.K.; Tusé, D. Synthetic melanin suppresses production of proinflammatory cytokines. Cell. Immunol. 2000, 199, 25–36. [Google Scholar] [CrossRef]
- Yu, S.; Wang, G.; Liao, J. Association of a novel SNP in the ASIP gene with skin color in black-bone chicken. Anim. Genet. 2019, 50, 283–286. [Google Scholar] [CrossRef]
- Prota, G. The role of peroxidase in melanogenesis revisited. Pigment Cell Res. 1992, 3 (Suppl. S2), 25–31. [Google Scholar] [CrossRef]
- Dou, T.; Yan, S.; Liu, L.; Wang, K.; Jian, Z.; Xu, Z.; Zhao, J.; Wang, Q.; Sun, S.; Talpur, M.Z.; et al. Integrative analysis of transcriptomics and metabolomics to reveal the melanogenesis pathway of muscle and related meat characters in Wuliangshan black-boned chickens. BMC Genom. 2022, 23, 173. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, F.; Cao, J.; Liu, X. Skin transcriptome profiles associated with skin color in chickens. PLoS ONE 2015, 10, e0127301. [Google Scholar] [CrossRef]
- Zhang, P.; Cao, Y.; Fu, Y.; Zhu, H.; Xu, S.; Zhang, Y.; Li, W.; Sun, G.; Jiang, R.; Han, R.; et al. Revealing the regulatory mechanism of lncRNA-LMEP on melanin deposition based on high-throughput sequencing in Xichuan Chicken skin. Genes 2022, 13, 2143. [Google Scholar] [CrossRef]
- Khumpeerawat, P.; Duangjinda, M.; Phasuk, Y. Factors affecting gene expression associated with the skin color of black-bone chicken in Thailand. Poult. Sci. 2021, 100, 101440. [Google Scholar] [CrossRef]
- Jian, H.; Zu, P.; Rao, Y.; Li, W.; Mou, T.; Lin, J.; Zhang, F. Comparative analysis of melanin deposition between Chishui silky fowl and Taihe silky fowl. J. Appl. Anim. Res. 2021, 49, 366–373. [Google Scholar] [CrossRef]
- Liao, G.Z.; Wang, G.Y.; Cheng, Z.B.; Jia, J.J.; Ge, C.R. Proteomic analysis of Tengchong snowcock muscles in Yunnan. Meat Res. 2013, 27, 1–5. [Google Scholar]
- Wang, H.; Cahaner, A.; Lou, L.; Zhang, L.; Ge, Y.; Li, Q.; Zhang, X. Genetics and breeding of a black-bone and blue eggshell chicken line. 1. Body weight, skin color, and their combined selection. Poult. Sci. 2021, 100, 101035. [Google Scholar] [CrossRef]
- Kriangwanich, W.; Piboon, P.; Sakorn, W.; Buddhachat, K.; Kochagul, V.; Pringproa, K.; Mekchay, S.; Nganvongpanit, K. Consistency of dark skeletal muscles in Thai native black-bone chickens (Gallus gallus domesticus). PeerJ 2021, 9, e10728. [Google Scholar] [CrossRef]
- Nganvongpanit, K.; Kaewkumpai, P.; Kochagul, V.; Pringproa, K.; Punyapornwithaya, V.; Mekchay, S. Distribution of Melanin Pigmentation in 33 Organs of Thai Black-Bone Chickens (Gallus gallus domesticus). Animals 2020, 10, 777. [Google Scholar] [CrossRef]
- Clarys, P.; Alewaeters, K.; Lambrecht, R.; Barel, A.O. Skin color measurements: Comparison between three instruments: The Chromameter(R), the DermaSpectrometer(R) and the Mexameter(R). Ski. Res. Technol. 2000, 6, 230–238. [Google Scholar] [CrossRef]
- Liu, W.B.; Chen, S.R.; Zheng, J.X.; Qu, L.J.; Xu, G.Y.; Yang, N. Developmental phenotypic-genotypic associations of tyrosinase and melanocortin 1 receptor genes with changing profiles in chicken plumage pigmentation. Poult. Sci. 2010, 89, 1110–1114. [Google Scholar] [CrossRef]
- Mackintosh, J.A. The antimicrobial properties of melanocytes, melanosomes and melanin and the evolution of black skin. J. Theor. Biol. 2001, 211, 101–113. [Google Scholar] [CrossRef]
- Wang, S.; Chen, X.; Hu, S.; Wu, Z. The influence of several factors on the aggregate ratio of melanin in Taihe Silky Fowl. J. Biobased Mater. Bioenergy 2012, 6, 710–713. [Google Scholar] [CrossRef]
- Nishimura, S.; Oshima, I.; Ono, Y.; Tabata, S.; Ishibashi, A.; Iwamoto, H. Age-related changes in the intramuscular distribution of melanocytes in the Silky fowl. Br. Poult. Sci. 2006, 47, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Wang, S.; Hu, Y.; Zhang, Y.; Dong, X.; Yang, Z.; Wang, J.; Li, J.; Deng, X. Hyperpigmentation results in aberrant immune development in silky fowl (Gallus gallus domesticus Brisson). PLoS ONE 2015, 10, e0125686. [Google Scholar] [CrossRef] [PubMed]
- Duffy, D.L.; Zhao, Z.Z.; Sturm, R.A.; Hayward, N.K.; Martin, N.G.; Montgomery, G.W. Multiple pigmentation gene polymorphisms account for a substantial proportion of risk of cutaneous malignant melanoma. J. Investig. Dermatol. 2010, 130, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Nan, H.; Kraft, P.; Hunter, D.J.; Han, J. Genetic variants in pigmentation genes, pigmentary phenotypes, and risk of skin cancer in Caucasians. Int. J. Cancer 2009, 125, 909–917. [Google Scholar] [CrossRef]
- Fan, R.; Xie, J.; Bai, J.; Wang, H.; Tian, X.; Bai, R.; Jia, X.; Yang, L.; Song, Y.; Herrid, M.; et al. Skin transcriptome profiles associated with coat color in sheep. BMC Genom. 2013, 14, 389. [Google Scholar] [CrossRef]
- Yu, S.; Wang, G.; Liao, J.; Tang, M. Transcriptome profile analysis identifies candidate genes for the melanin pigmentation of breast muscle in Muchuan black-boned chicken. Poult. Sci. 2018, 97, 3446–3455. [Google Scholar] [CrossRef]
- Fan, Y.; Wu, X.; Li, Y.; Han, H.; Zhang, Y.; Yang, J.; Liu, Y. Effect of polymorphisms in the 5′-flanking sequence of MC1R on feather color in Taihang chickens. Poult. Sci. 2022, 101, 102192. [Google Scholar] [CrossRef]
- Cullinane, A.R.; Vilboux, T.; O’Brien, K.; Curry, J.A.; Maynard, D.M.; Carlson-Donohoe, H.; Ciccone, C.; Markello, T.C.; Gunay-Aygun, M.; Huizing, M.; et al. Homozygosity mapping and whole-exome sequencing to detect SLC45A2 and G6PC3 mutations in a single patient with oculocutaneous albinism and neutropenia. J. Investig. Dermatol. 2011, 131, 2017–2025. [Google Scholar] [CrossRef]
- Lopez, V.M.; Decatur, C.L.; Stamer, W.D.; Lynch, R.M.; McKay, B.S. L-DOPA is an endogenous ligand for OA1. PLoS Biol. 2008, 6, e236. [Google Scholar] [CrossRef]
- Costin, G.E.; Valencia, J.C.; Vieira, W.D.; Lamoreux, M.L.; Hearing, V.J. Tyrosinase processing and intracellular trafficking is disrupted in mouse primary melanocytes carrying the underwhite (uw) mutation. A model for oculocutaneous albinism (OCA) type 4. J. Cell Sci. 2003, 116, 3203–3212. [Google Scholar] [CrossRef]
- Cho, E.; Kim, M.; Manjula, P.; Cho, S.H.; Seo, D.; Lee, S.S.; Lee, J.H. A retroviral insertion in the tyrosinase (TYR) gene is associated with the recessive white plumage color in the Yeonsan Ogye chicken. J. Anim. Sci. Technol. 2021, 63, 751–758. [Google Scholar] [CrossRef]
- Nam, I.S.; Oh, M.G.; Nam, M.S.; Kim, W.S. Specific mutations in the genes of MC1R and TYR have an important influence on the determination of pheomelanin pigmentation in Korean native chickens. J. Adv. Vet. Anim. Res. 2021, 8, 266–273. [Google Scholar] [CrossRef]
- Yang, C.W.; Ran, J.S.; Yu, C.L.; Qiu, M.H.; Zhang, Z.R.; Du, H.R.; Li, Q.Y.; Xiong, X.; Song, X.Y.; Xia, B.; et al. Polymorphism in MC1R, TYR and ASIP genes in different colored feather chickens. 3 Biotech. 2019, 9, 203. [Google Scholar] [CrossRef]
- Ludwig, A.; Rehberg, S.; Wegner, M. Melanocyte-specific expression of dopachrome tautomerase is dependent on synergistic gene activation by the Sox10 and Mitf transcription factors. FEBS Lett. 2004, 556, 236–244. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, X.; Zhang, Q.; Wang, Y.; Wang, S.; Li, Y.; Zhang, Y.; Jing, J.; Qiu, J.; Wang, Z.; et al. Comparative transcriptome profiles of Lindian chicken eyelids identify melanin genes controlling eyelid pigmentation. Br. Poult. Sci. 2019, 60, 15–22. [Google Scholar] [CrossRef]
- Binstock, M.; Hafeez, F.; Metchnikoff, C.; Arron, S.T. Single-nucleotide polymorphisms in pigment genes and nonmelanoma skin cancer predisposition: A systematic review. Br. J. Dermatol. 2014, 171, 713–721. [Google Scholar] [CrossRef]
- Kawasaki-Nishihara, A.; Nishihara, D.; Nakamura, H.; Yamamoto, H. ET3/Ednrb2 signaling is critically involved in regulating melanophore migration in Xenopus. Dev. Dyn. 2011, 240, 1454–1466. [Google Scholar] [CrossRef]
- Kinoshita, K.; Akiyama, T.; Mizutani, M.; Shinomiya, A.; Ishikawa, A.; Younis, H.H.; Tsudzuki, M.; Namikawa, T.; Matsuda, Y. Endothelin receptor B2 (EDNRB2) is responsible for the tyrosinase-independent recessive white (mow) and mottled (mo) plumage phenotypes in the chicken. PLoS ONE 2014, 9, e86361. [Google Scholar] [CrossRef]
- Wu, N.; Qin, H.; Wang, M.; Bian, Y.; Dong, B.; Sun, G.; Zhao, W.; Chang, G.; Xu, Q.; Chen, G. Variations in endothelin receptor B subtype 2 (EDNRB2) coding sequences and mRNA expression levels in 4 Muscovy duck plumage colour phenotypes. Br. Poult. Sci. 2017, 58, 116–121. [Google Scholar] [CrossRef]
- Shi, H.; Fu, J.; He, Y.; Li, Z.; Kang, J.; Hu, C.; Zi, X.; Liu, Y.; Zhao, J.; Dou, T.; et al. Hyperpigmentation inhibits early skeletal muscle development in Tengchong Snow Chicken breed. Genes 2022, 13, 2253. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhou, C.; Xie, H.; Huang, S.; Wang, J.; Liu, S. NGS-based targeted sequencing identified two novel variants in Southwestern Chinese families with oculocutaneous albinism. BMC Genom. 2022, 23, 332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.D.; Wang, H.H.; Zhang, C.X.; Li, Q.H.; Chen, X.H.; Lou, L.F. Analysis of skin color change and related gene expression after crossing of Dongxiang black chicken and ISA layer. Genet. Mol. Res. 2015, 14, 11551–11561. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, A.G.; McKay, B.S. GPR143 signaling and retinal degeneration. Adv. Exp. Med. Biol. 2019, 1185, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Giordano, F.; Bonetti, C.; Surace, E.M.; Marigo, V.; Raposo, G. The ocular albinism type 1 (OA1) G-protein-coupled receptor functions with MART-1 at early stages of melanogenesis to control melanosome identity and composition. Hum. Mol. Genet. 2009, 18, 4530–4545. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, R.; Peng, Y.; Zhang, C.; Li, L.; Lu, C.; Li, X. Feather follicles transcriptome profiles in Bashang long-tailed chickens with different plumage colors. Genes Genom. 2019, 41, 1357–1367. [Google Scholar] [CrossRef]
- Chen, T.Z.; Zhao, B.L.; Liu, Y.; Zhao, Y.Y.; Wang, H.D.; Fan, R.W.; Wang, P.C.; Dong, C.S. Expression and localization of GPR143 in sheep skin. Yi Chuan 2016, 38, 658–665. [Google Scholar] [CrossRef]
- Wu, J.; Lin, Z.; Chen, G.; Luo, Q.; Nie, Q.; Zhang, X.; Luo, W. Characterization of chicken skin yellowness and exploration of genes involved in skin yellowness deposition in chicken. Front. Physiol. 2021, 12, 585089. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L Value | Total Melanin | Eumelanin | Phaeomelanosome | |
|---|---|---|---|---|
| L value | 1 | |||
| Total melanin | −0.987 ** | 1 | ||
| Eumelanin | −0.663 * | 0.617 * | 1 | |
| Phaeomelanosome | −0.728 * | 0.767 * | 0.031 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zi, X.; Ge, X.; Zhu, Y.; Liu, Y.; Sun, D.; Li, Z.; Liu, M.; You, Z.; Wang, B.; Kang, J.; et al. Transcriptome Profile Analysis Identifies Candidate Genes for the Melanin Pigmentation of Skin in Tengchong Snow Chickens. Vet. Sci. 2023, 10, 341. https://doi.org/10.3390/vetsci10050341
Zi X, Ge X, Zhu Y, Liu Y, Sun D, Li Z, Liu M, You Z, Wang B, Kang J, et al. Transcriptome Profile Analysis Identifies Candidate Genes for the Melanin Pigmentation of Skin in Tengchong Snow Chickens. Veterinary Sciences. 2023; 10(5):341. https://doi.org/10.3390/vetsci10050341
Chicago/Turabian StyleZi, Xiannian, Xuehai Ge, Yixuan Zhu, Yong Liu, Dawei Sun, Zijian Li, Mengqian Liu, Zhengrong You, Bo Wang, Jiajia Kang, and et al. 2023. "Transcriptome Profile Analysis Identifies Candidate Genes for the Melanin Pigmentation of Skin in Tengchong Snow Chickens" Veterinary Sciences 10, no. 5: 341. https://doi.org/10.3390/vetsci10050341