
Impact of Two Commercial S. cerevisiae Strains on the Aroma Profiles of Different Regional Musts
 ,
,  , , , , , ,
, , , , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Inoculum Preparation
2.3. Fermentation Trials
2.4. Volatile Aroma Profile
2.5. Statistical Analysis
- (a)
- The table of standardized effects, which reports on the statistical weight (Fisher test and p-value) of the kind of strain, or must, as well as on their interaction.
- (b)
- The decomposition of the statistical hypothesis, which is a mathematical function showing the correlation of each factor (must, strain, must × strain) with the dependent variable.
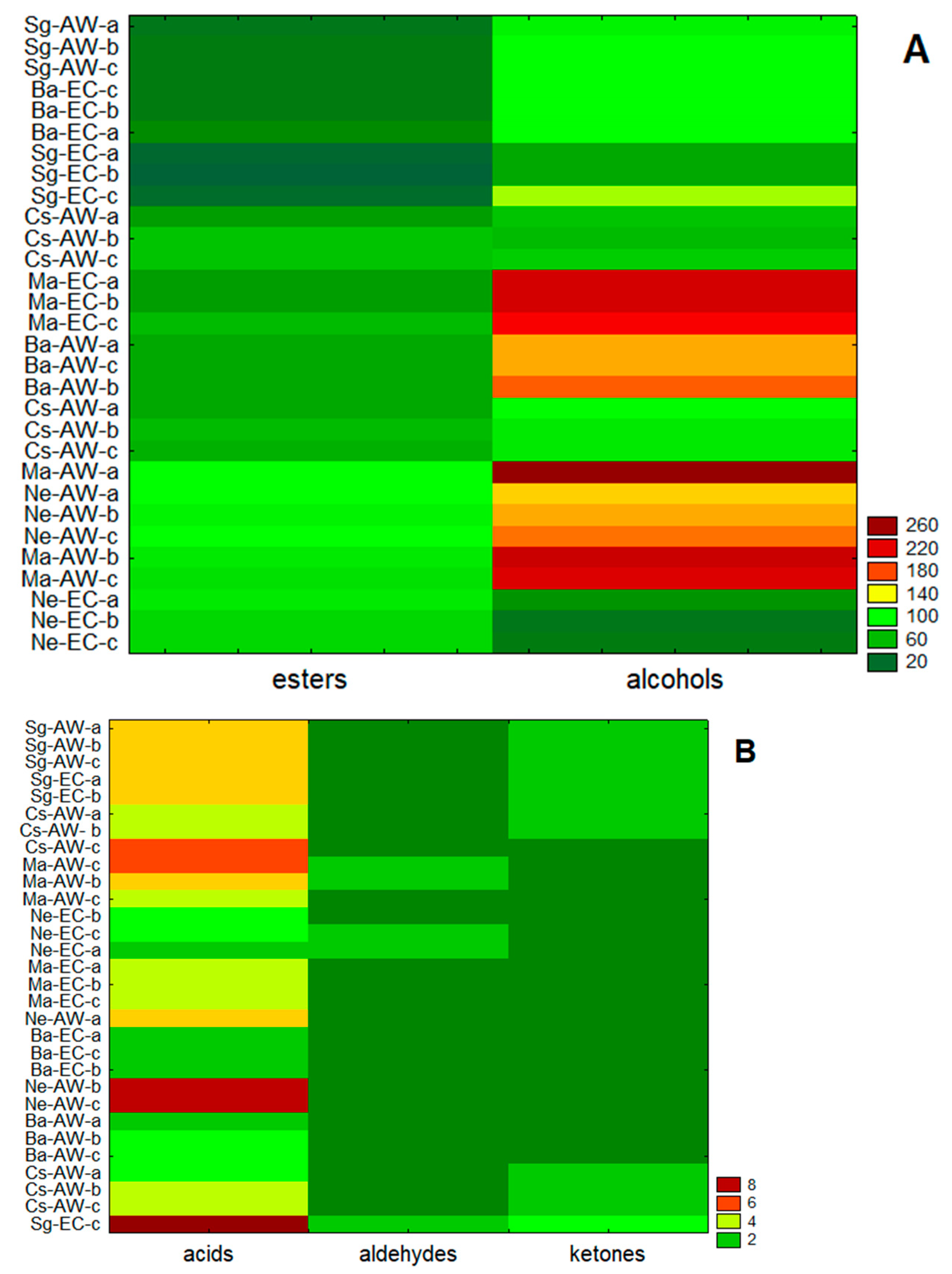
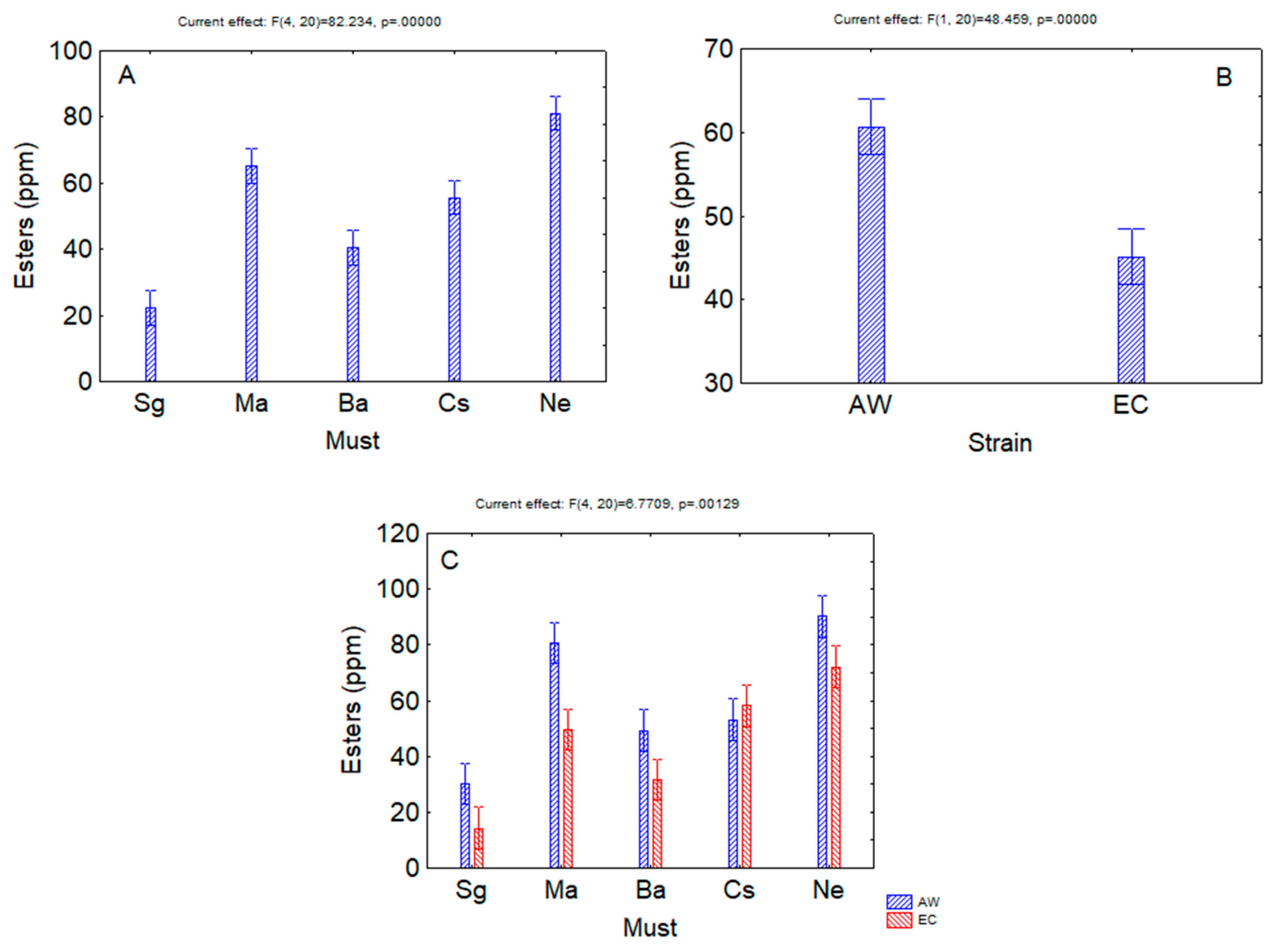
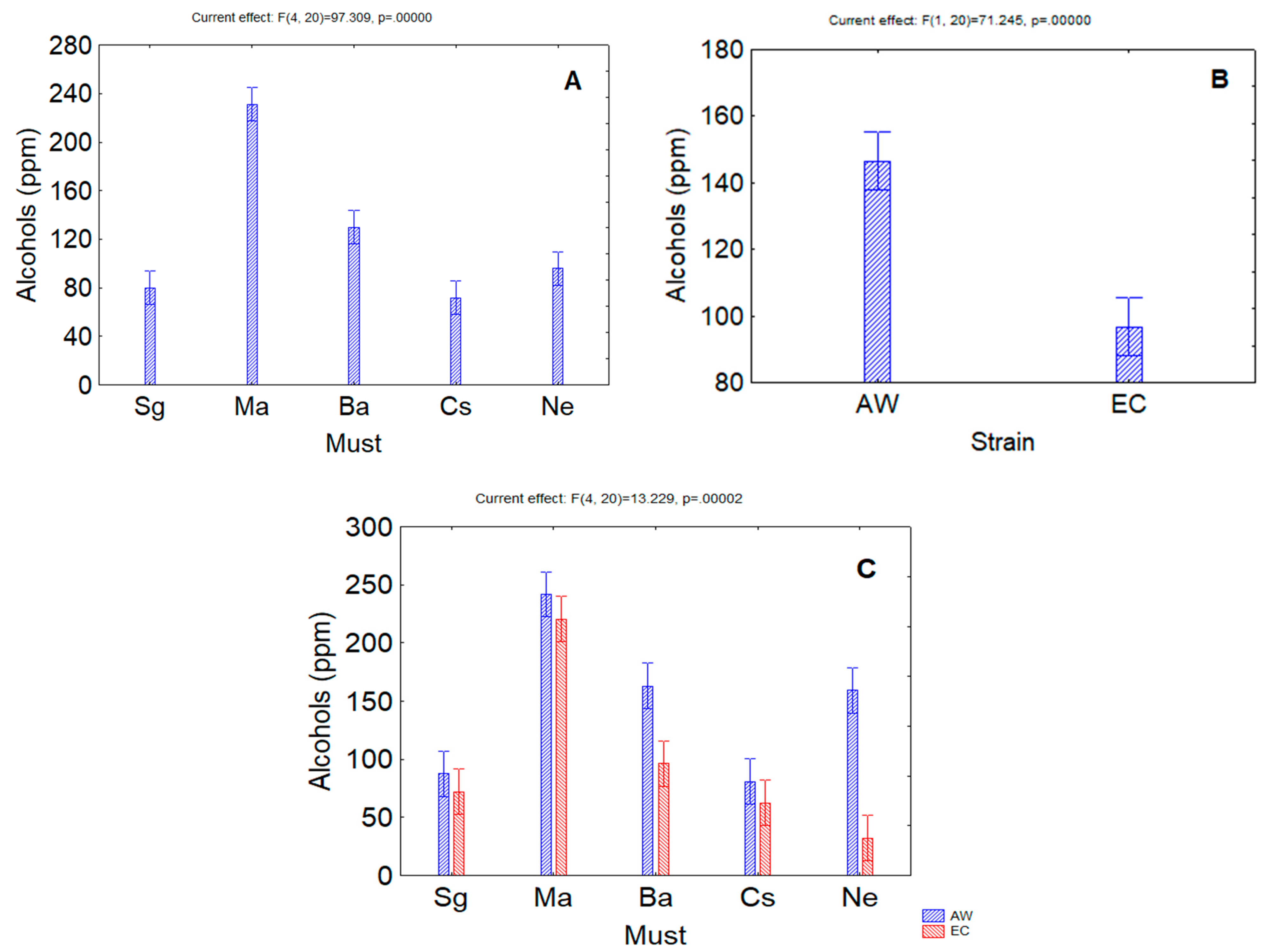
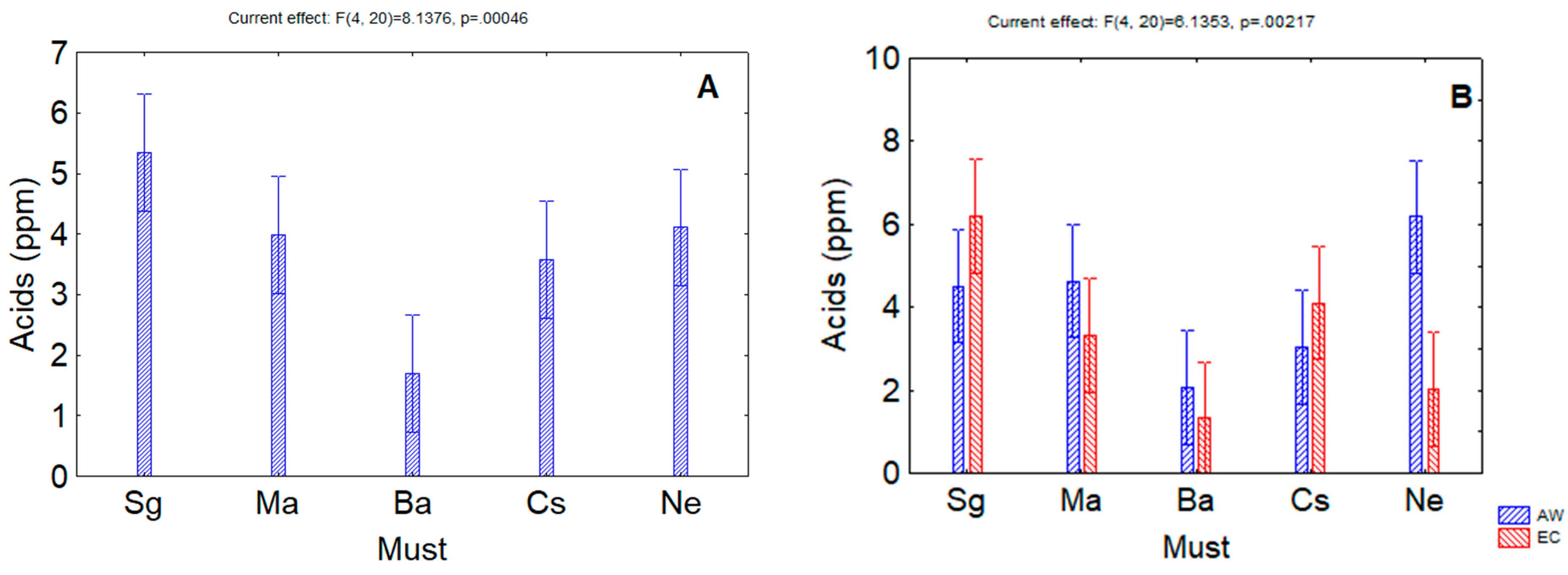
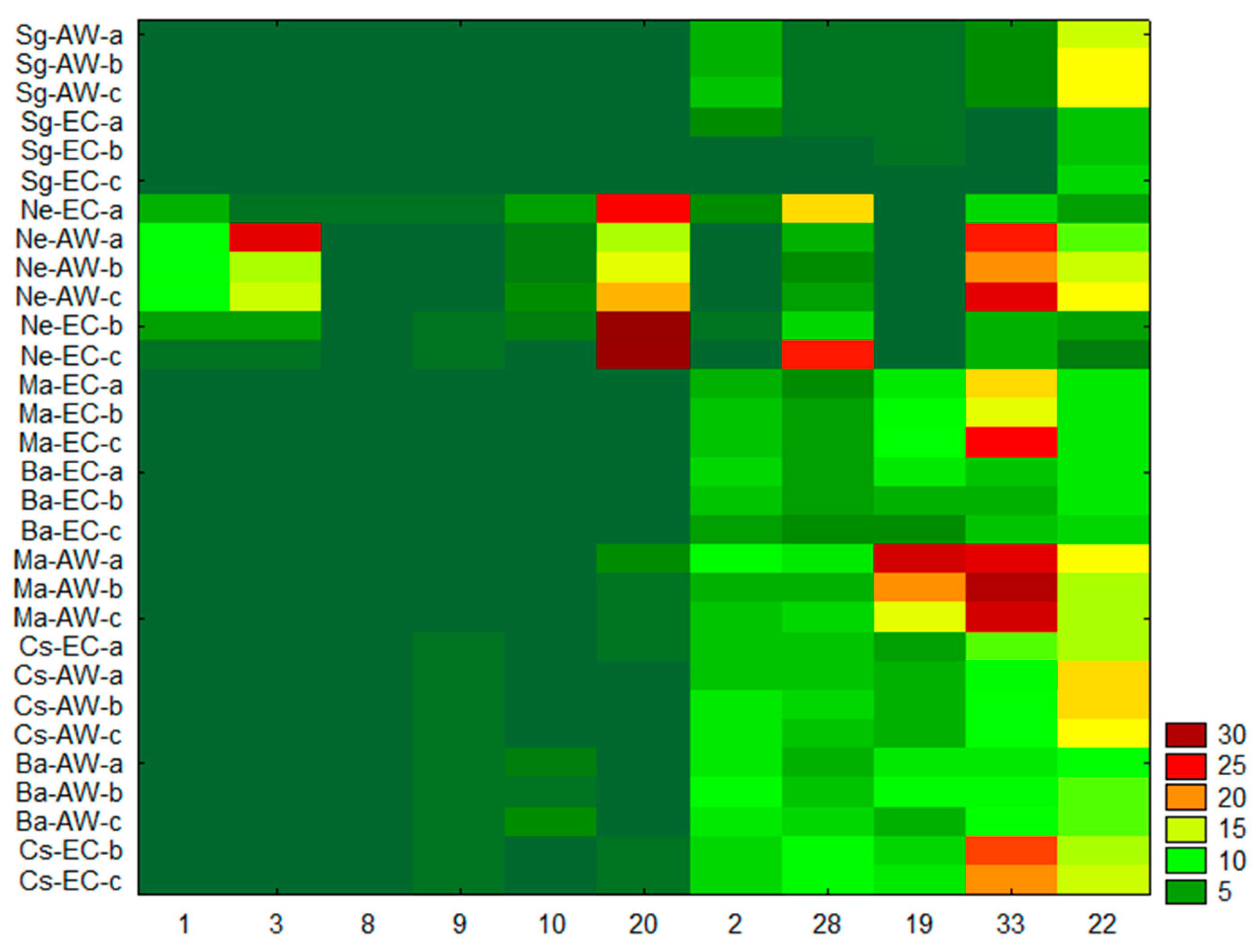
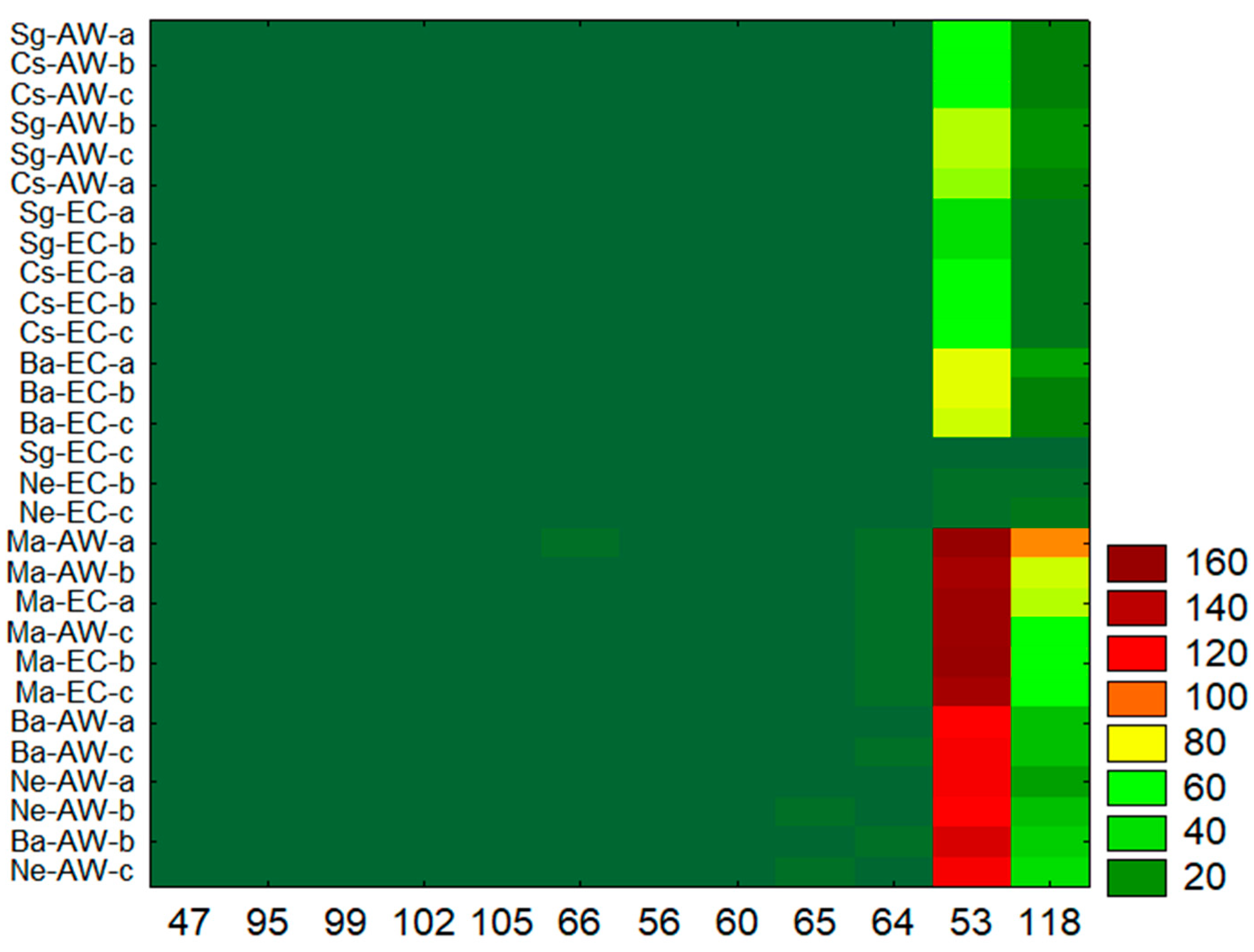
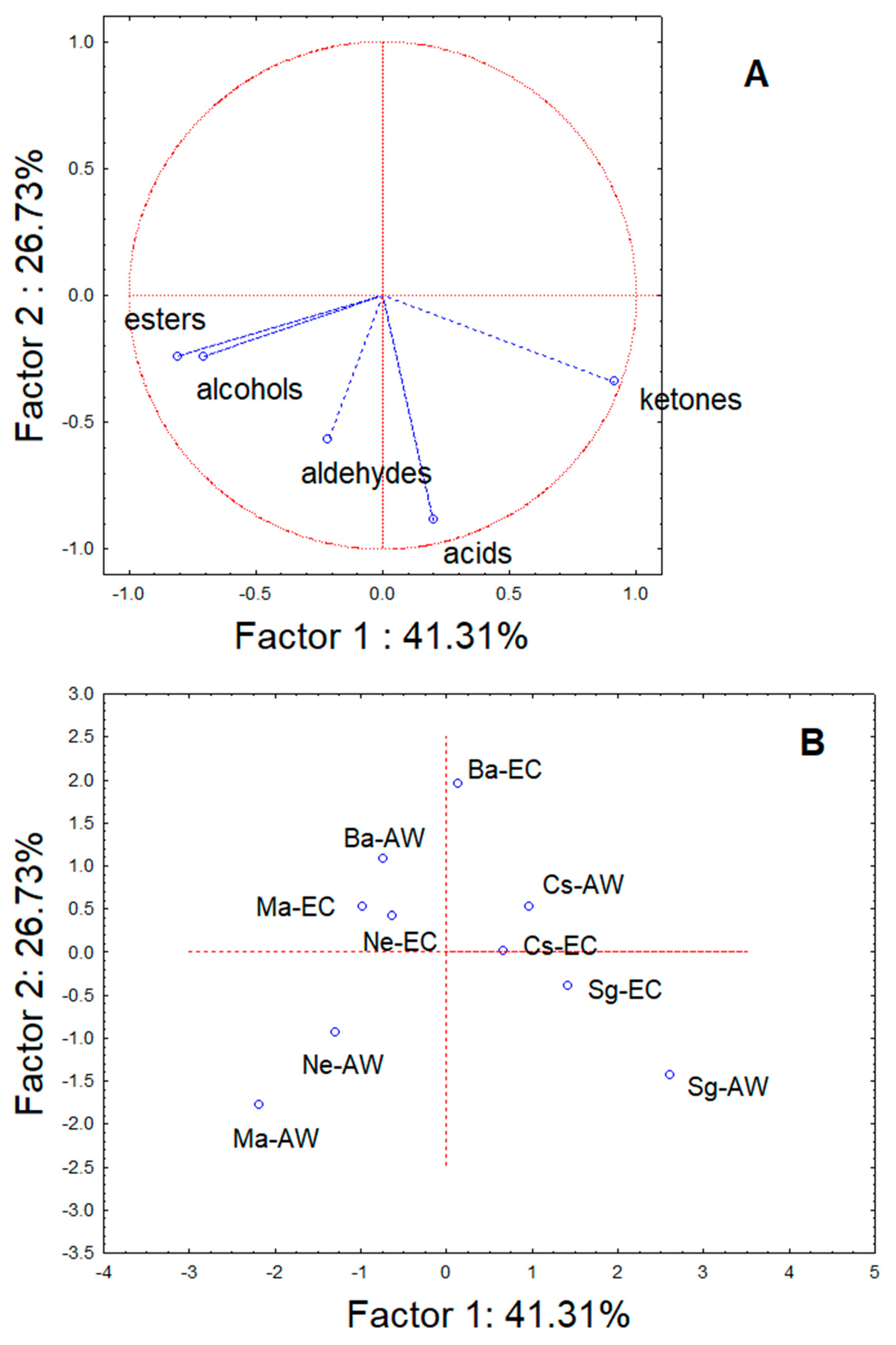
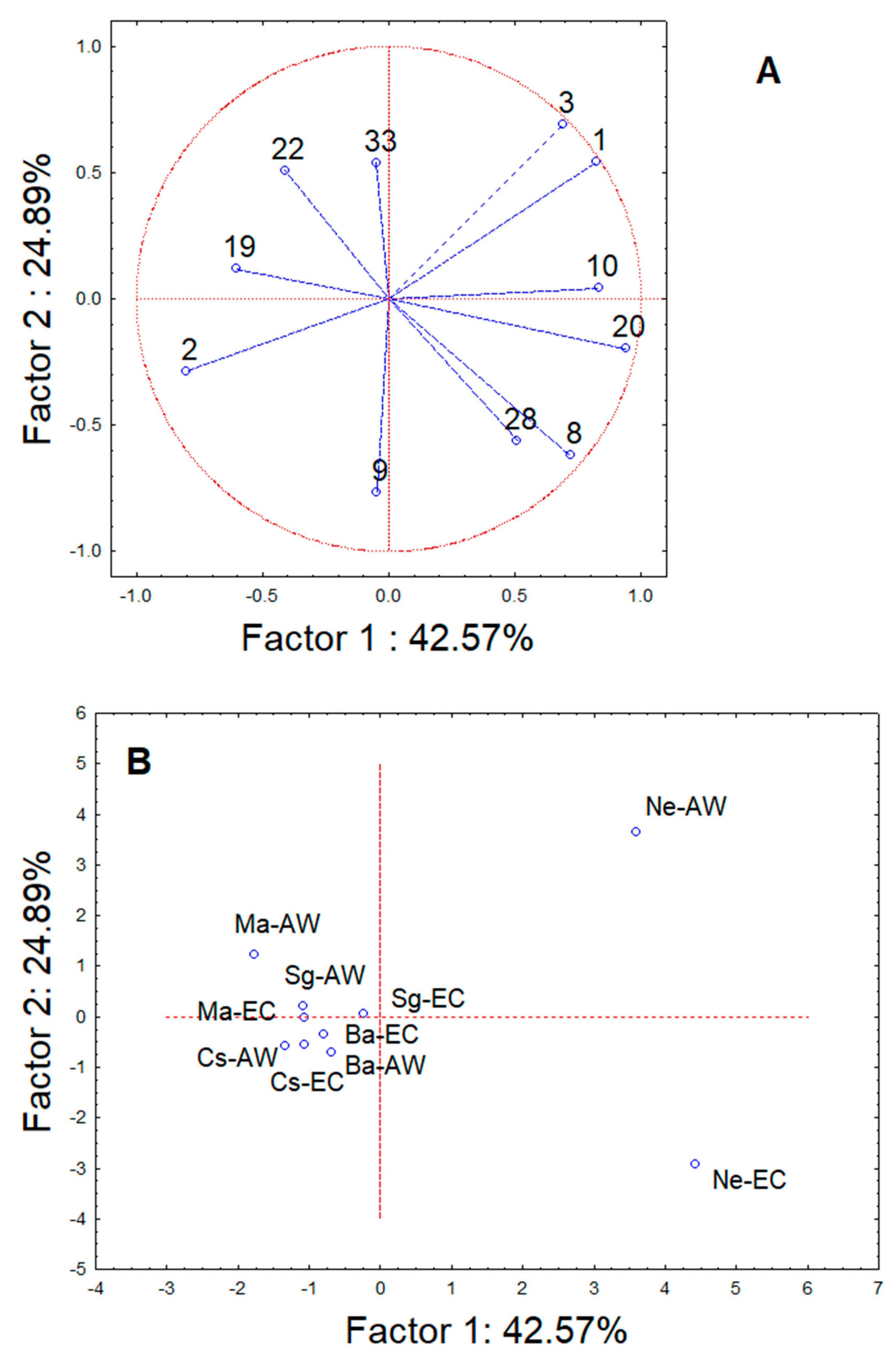
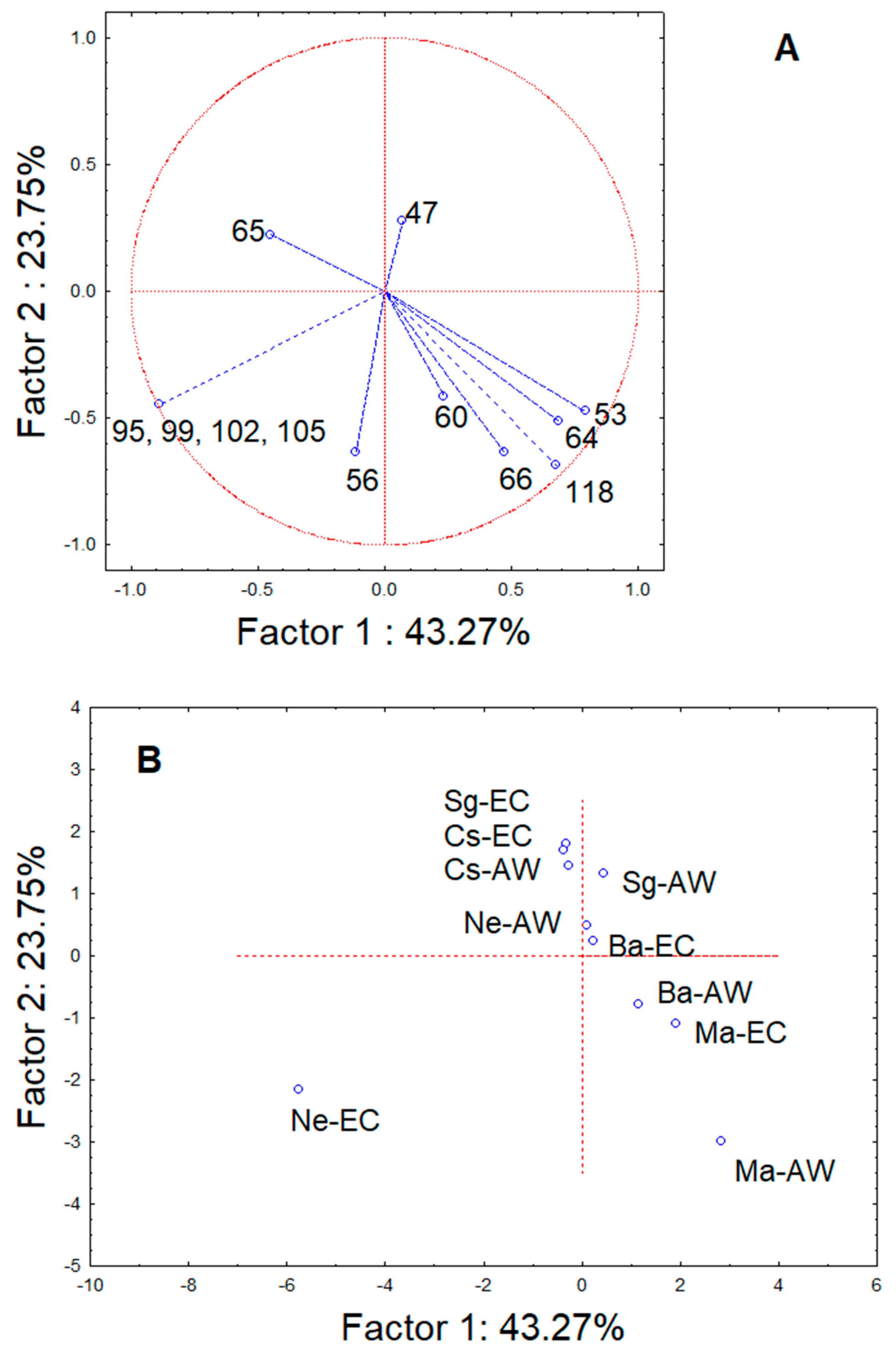
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Barbosa, C.; Lage, P.; Vilela, A.; Mendes-Faia, A.; Mendes-Ferreira, A. Phenotypic and metabolic traits of commercial Saccharomyces cerevisiae yeasts. AMB Express 2014, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Roullier-Gall, C.; David, V.; Hemmler, D.; Schmitt-Kopplin, P.; Alexandre, H. Exploring yeast interactions through metabolic profiling. Sci. Rep. 2020, 10, 6073. [Google Scholar] [CrossRef]
- Capece, A.; Romano, P. Genetic improvement of wine yeasts. In Yeasts in the Production of Wine; Romano, P., Ciani, M., Fleet, G.H., Eds.; Springer: New York, NY, USA, 2019; pp. 315–342. [Google Scholar] [CrossRef]
- Marsit, S.; Dequin, S. Diversity and adaptive evolution of Saccharomyces wine yeast: A review. FEMS Yeast Res. 2015, 15, fov067. [Google Scholar] [CrossRef] [PubMed]
- Feghali, N.; Albertin, W.; Tabet, E.; Rizk, Z.; Bianco, A.; Zara, G.; Masneuf-Pomarede, I.; Budroni, M. Genetic and Phenotypic Characterisation of a Saccharomyces cerevisiae Population of ‘Merwah’White Wine. Microorganisms 2019, 7, 492. [Google Scholar] [CrossRef] [PubMed]
- Mercado, L.; Sturm, M.E.; Rojo, M.C.; Ciklic, I.; Martínez, C.; Combina, M. Biodiversity of Saccharomyces cerevisiae populations in Malbec vineyards from the “Zona Alta del Río Mendoza” region in Argentina. Int. J. Food Microbiol. 2011, 151, 319–326. [Google Scholar] [CrossRef]
- Guillamón, J.M.; Barrio, E. Genetic polymorphism in wine yeasts: Mechanisms and methods for its detection. Front. Microbiol. 2017, 8, 806. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Kievit, R.L.; Siebert, T.; Lattey, K.A.; Bramley, B.R.; Francis, I.L.; King, E.S.; Pretorius, I.S. The influence of yeast on the aroma of Sauvignon Blanc wine. Food Microbiol. 2009, 26, 204–211. [Google Scholar] [CrossRef]
- Francesca, N.; Chiurazzi, M.; Romano, R.; Aponte, M.; Settanni, L.; Moschetti, G. Indigenous yeast communities in the environment of “Rovello bianco” grape variety and their use in commercial white wine fermentation. World J. Microbiol. Biotechnol. 2010, 26, 337–351. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Diversity of Saccharomyces cerevisiae yeasts associated to spontaneously fermenting grapes from an Italian “heroic vine-growing area”. Food Microbiol. 2012, 31, 159–166. [Google Scholar] [CrossRef]
- Puertas, B.; Jimenez-Hierro, M.; Cantos-Villar, E.; Marrufo-Curtido, A.; Carbú, M.; Cuevas, F.; Moreno-Rojas, J.; González-Rodríguez, V.; Cantoral, J.; Ruiz-Moreno, M. The influence of yeast on chemical composition and sensory properties of dry white wines. Food Chem. 2018, 253, 227–235. [Google Scholar] [CrossRef]
- De Filippis, F.; Aponte, M.; Piombino, P.; Lisanti, M.T.; Moio, L.; Ercolini, D.; Blaiotta, G. Influence of microbial communities on the chemical and sensory features of Falanghina sweet passito wines. Food Res. Int. 2019, 120, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Cotea, V.V.; Focea, M.C.; Luchian, C.E.; Colibaba, L.C.; Scutarașu, E.C.; Marius, N.; Zamfir, C.I.; Popîrdă, A. Influence of different commercial yeasts on volatile fraction of sparkling wines. Foods 2021, 10, 247. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145. [Google Scholar] [CrossRef] [PubMed]
- Duarte, F.L.; Baleiras-Couto, M.M. Survey of Inoculated Commercial Saccharomyces cerevisiae in Winery-Based Trials. Fermentation 2021, 7, 176. [Google Scholar] [CrossRef]
- Romano, P.; Braschi, G.; Siesto, G.; Patrignani, F.; Lanciotti, R. Role of Yeasts on the Sensory Component of Wines. Foods 2022, 11, 1921. [Google Scholar] [CrossRef]
- Torrens, J.; Urpí, P.; Riu-Aumatell, M.; Vichi, S.; López-Tamames, E.; Buxaderas, S. Different commercial yeast strains affecting the volatile and sensory profile of cava base wine. Int. J. Food Microbiol. 2008, 124, 48–57. [Google Scholar] [CrossRef]
- Perestrelo, R.; Silva, C.; Câmara, J.S. Madeira wine volatile profile. A platform to establish madeira wine aroma descriptors. Molecules 2019, 24, 3028. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascués, E.; Marquina, D.; Santos, A. Unraveling the enzymatic basis of wine “flavorome”: A phylo-functional study of wine related yeast species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Hirst, M.B.; Richter, C.L. Review of aroma formation through metabolic pathways of Saccharomyces cerevisiae in beverage fermentations. Am. J. Enol. Vitic. 2016, 67, 361–370. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Li, J. Aroma compounds in wine. In Grape and Wine Biotechnology; IntechOpen: Rijeka, Croatia, 2016; pp. 273–283. [Google Scholar] [CrossRef]
- Marín-San Román, S.; Rubio-Bretón, P.; Pérez-Álvarez, E.P.; Garde-Cerdán, T. Advancement in analytical techniques for the extraction of grape and wine volatile compounds. Food Res. Int. 2020, 137, 109712. [Google Scholar] [CrossRef] [PubMed]
- Mina, M.; Tsaltas, D. Contribution of yeast in wine aroma and flavour. In Yeast-Industrial Applications; Morata, A., Loira, I., Eds.; IntechOpen: Rijeka, Croatia, 2017; pp. 117–134. [Google Scholar] [CrossRef]
- Eldarov, M.; Kishkovskaia, S.; Tanaschuk, T.; Mardanov, A. Genomics and biochemistry of Saccharomyces cerevisiae wine yeast strains. Biochemistry 2016, 81, 1650–1668. [Google Scholar] [CrossRef] [PubMed]
- Tetik, M.A.; Sevindik, O.; Kelebek, H.; Selli, S. Screening of key odorants and anthocyanin compounds of cv. Okuzgozu (Vitis vinifera L.) red wines with a free run and pressed pomace using GC-MS-Olfactometry and LC-MS-MS. J. Mass Spectrom. 2018, 53, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Jeleń, H.H.; Szczurek, A. Solid phase microextraction for profiling volatile compounds in liquered white wines. Acta Sci. Pol. Technol. Aliment. 2010, 9, 23–32. [Google Scholar]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial contribution to wine aroma and its intended use for wine quality improvement. Molecules 2017, 22, 189. [Google Scholar] [CrossRef]
- Bisson, L.F.; Karpel, J.E. Genetics of yeast impacting wine quality. Annu. Rev. Food Sci. Technol. 2010, 1, 139–162. [Google Scholar] [CrossRef]
- Cordente, A.G.; Curtin, C.D.; Varela, C.; Pretorius, I.S. Flavour-active wine yeasts. Appl. Microbiol. Biotechnol. 2012, 96, 601–618. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavor. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar] [CrossRef]
- Saerens, S.M.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef]
- Barrajón, N.; Arévalo-Villena, M.; Úbeda, J.; Briones, A. Enological properties in wild and commercial Saccharomyces cerevisiae yeasts: Relationship with competition during alcoholic fermentation. World J. Microbiol. Biotechnol. 2011, 27, 2703–2710. [Google Scholar] [CrossRef]
- Kłosowski, G.; Mikulski, D.; Rolbiecka, A.; Czupryński, B. Changes in the concentration of carbonyl compounds during the alcoholic fermentation process carried out with Saccharomyces cerevisiae yeast. Pol. J. Microbiol. 2017, 66, 327–334. [Google Scholar] [CrossRef]
- Synos, K.; Reynolds, A.G.; Bowen, A.J. Effect of yeast strain on aroma compounds in Cabernet franc icewines. LWT-Food Sci. Technol. 2015, 64, 227–235. [Google Scholar] [CrossRef]
- Bely, M.; Rinaldi, A.; Dubourdieu, D. Influence of assimilable nitrogen on volatile acidity production by Saccharomyces cerevisiae during high sugar fermentation. J. Biosci. Bioeng. 2003, 96, 507–512. [Google Scholar] [CrossRef]
- Chidi, B.S.; Bauer, F.F.; Rossouw, D. Organic acid metabolism and the impact of fermentation practices on wine acidity: A review. S. Afr. J. Enol. Vitic. 2018, 39, 1–15. [Google Scholar] [CrossRef]
- Romano, P.; Siesto, G.; Capece, A.; Pietrafesa, R.; Lanciotti, R.; Patrignani, F.; Granchi, L.; Galli, V.; Bevilacqua, A.; Campaniello, D. Validation of a Standard Protocol to Assess the Fermentative and Chemical Properties of Saccharomyces cerevisiae Wine Strains. Front. Microbiol. 2022, 13, 830277. [Google Scholar] [CrossRef]
- Anonymous, 2009 Codex—Active Dry Yeasts; Modification Resolution OIV-OENO 329/Zagreb; International Organization of Vine and Wine General: Paris, France, 2009.
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL medium to profile native flora fermentations. Am. J. Enol. Vitic. 2001, 52, 198–203. [Google Scholar]
- Boudreau, T.F.; Peck, G.M.; O’Keefe, S.F.; Stewart, A.C. Free amino nitrogen concentration correlates to total yeast assimilable nitrogen concentration in apple juice. Food Sci. Nutr. 2018, 6, 119–123. [Google Scholar] [CrossRef]
- Patrignani, F.; Montanari, C.; Serrazanetti, D.I.; Braschi, G.; Vernocchi, P.; Tabanelli, G.; Parpinello, G.P.; Versari, A.; Gardini, F.; Lanciotti, R. Characterisation of yeast microbiota, chemical and sensory properties of organic and biodynamic Sangiovese red wines. Ann. Microbiol. 2017, 67, 99–109. [Google Scholar] [CrossRef]
- Parpinello, G.P.; Ricci, A.; Folegatti, B.; Patrignani, F.; Lanciotti, R.; Versari, A. Unraveling the potential of cryotolerant Saccharomyces eubayanus in Chardonnay white wine production. LWT-Food Sci. Technol. 2020, 134, 110183. [Google Scholar] [CrossRef]
- Antalick, G.; Šuklje, K.; Blackman, J.W.; Meeks, C.; Deloire, A.; Schmidtke, L.M. Influence of grape composition on red wine ester profile: Comparison between Cabernet Sauvignon and Shiraz cultivars from Australian warm climate. J. Agric. Food Chem. 2015, 63, 4664–4672. [Google Scholar] [CrossRef]
- Liu, P.; Ivanova-Petropulos, V.; Duan, C.; Yan, G. Effect of unsaturated fatty acids on intra-metabolites and aroma compounds of Saccharomyces cerevisiae in wine fermentation. Foods 2021, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.L.; Dunn, B.; Sherlock, G.; Pugh, T. Comparative metabolic footprinting of a large number of commercial wine yeast strains in Chardonnay fermentations. FEMS Yeast Res. 2013, 13, 394–410. [Google Scholar] [CrossRef] [PubMed]
- Nguela, J.M.; Vernhet, A.; Julien-Ortiz, A.; Sieczkowski, N.; Mouret, J.R. Effect of grape must polyphenols on yeast metabolism during alcoholic fermentation. Food Res. Int. 2019, 121, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Kiene, F.; Belda, I.; Fracassetti, D.; Marquina, D.; Navascués, E.; Calderón, F.; Benito, A.; Rauhut, D.; Santos, A.; et al. Effects on varietal aromas during wine making: A review of the impact of varietal aromas on the flavor of wine. Appl. Microbiol. Biotechnol. 2019, 103, 7425–7450. [Google Scholar] [CrossRef]
- Borren, E.; Tian, B. The important contribution of non-Saccharomyces yeasts to the aroma complexity of wine: A review. Foods 2020, 10, 13. [Google Scholar] [CrossRef]
- Casu, F.; Pinu, F.R.; Fedrizzi, B.; Greenwood, D.R.; Villas-Boas, S.G. The effect of linoleic acid on the Sauvignon blanc fermentation by different wine yeast strains. FEMS Yeast Res. 2016, 16, fow050. [Google Scholar] [CrossRef]
- Pinu, F.R.; Edwards, P.J.; Jouanneau, S.; Kilmartin, P.A.; Gardner, R.C.; Villas-Boas, S.G. Sauvignon blanc metabolomics: Grape juice metabolites affecting the development of varietal thiols and other aroma compounds in wines. Metabolomics 2014, 10, 556–573. [Google Scholar] [CrossRef]
- Pereira, C.; Mendes, D.; Dias, T.; Garcia, R.; da Silva, M.G.; Cabrita, M.J. Revealing the yeast modulation potential on amino acid composition and volatile profile of Arinto white wines by a combined chromatographic-based approach. J. Chromatogr. 2021, 1641, 461991. [Google Scholar] [CrossRef]
- Mandl, K.; Silhavy-Richter, K.; Wendelin, S.; Prinz, M.; Patzl-Fischerleitner, E.; Eder, R. Influence of different commercial yeasts on the phenol pattern of rosé wine. BIO Web Conf. 2017, 9, 02016. [Google Scholar] [CrossRef] [Green Version]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with non-Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef]
- Rollero, S.; Bloem, A.; Brand, J.; Ortiz-Julien, A.; Camarasa, C.; Divol, B. Nitrogen metabolism in three non-conventional wine yeast species: A tool to modulate wine aroma profiles. Food Microbiol. 2021, 94, 103650. [Google Scholar] [CrossRef] [PubMed]
- Carrau, F.M.; Medina, K.; Farina, L.; Boido, E.; Henschke, P.A.; Dellacassa, E. Production of fermentation aroma compounds by Saccharomyces cerevisiae wine yeasts: Effects of yeast assimilable nitrogen on two model strains. FEMS Yeast Res. 2008, 8, 1196–1207. [Google Scholar] [CrossRef] [PubMed]
- Loscos, N.; Hernandez-Orte, P.; Cacho, J.; Ferreira, V. Release and formation of varietal aroma compounds during alcoholic fermentation from nonfloral grape odorless flavor precursors fractions. J. Agric. Food Chem. 2007, 55, 6674–6684. [Google Scholar] [CrossRef] [PubMed]
- Crandles, M.; Reynolds, A.G.; Khairallah, R.; Bowen, A. The effect of yeast strain on odor active compounds in Riesling and Vidal blanc icewines. LWT-Food Sci. Technol. 2015, 64, 243–258. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Mayr, C.M.; Geue, J.P.; Holt, H.E.; Pearson, W.P.; Jeffery, D.W.; Francis, I.L. Characterization of the key aroma compounds in Shiraz wine by quantitation, aroma reconstitution, and omission studies. J. Agric. Food Chem. 2014, 62, 4528–4536. [Google Scholar] [CrossRef]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma-a review of flavour-active esters and higher alcohols produced by the brewing yeast. App. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Must | Strain | Must × Strain | |
|---|---|---|---|
| Esters | <0.00001 | <0.00001 | 0.00129 |
| Alcohols | <0.00001 | <0.00001 | 0.00002 |
| Acids | 0.00004 | ns | 0.00217 |
| Aldehydes | 0.00002 | ns | <0.00001 |
| Ketones | <0.00001 | ns | 0.00258 |
| IUPAC Name | Must | Strain | Must × Strain |
|---|---|---|---|
| ethyl undec-10-enoate | <0.00001 | <0.00001 | <0.00001 |
| 3-methylbutyl acetate | <0.00001 | 0.00993 | 0.00396 |
| [(2E)-3,7-dimethylocta-2,6-dienyl] acetate | <0.00001 | 0.00182 | 0.00002 |
| methyl non-7-ynoate | 0.00004 | 0.00251 | 0.00004 |
| 2-phenylethyl acetate | <0.00001 | 0.00113 | <0.00001 |
| 3-hydroxy-2,2-dimethoxypropyl acetate | <0.00001 | ns | 0.03931 |
| ethyl decanoate | <0.00001 | 0.00818 | 0.00094 |
| ethyl dodecanoate | <0.00001 | 0.00036 | <0.00001 |
| ethyl acetate | <0.00001 | <0.00001 | 0.00006 |
| ethyl hexanoate | 0.00001 | ns | 0.00053 |
| ethyl octanoate | <0.00001 | <0.00001 | <0.00001 |
| 2-methylbutane-1,3-diol | <0.00001 | 0.00008 | <0.00001 |
| 3-methylbutan-1-ol | <0.00001 | <0.00001 | <0.00001 |
| hexan-1-ol | 0.00066 | ns | ns |
| propan-1-ol | 0.000485 | 0.00004 | ns |
| 2-methylpropan-1-ol | <0.00001 | 0.00991 | 0.00192 |
| 2-methyl-2-nitropropan-1-ol | <0.00001 | ns | ns |
| 3-methylsulfanylpropan-1-ol | <0.00001 | 0.00004 | <0.00001 |
| 3-methylhexan-3-ol | <0.00001 | <0.00001 | <0.00001 |
| 4-(methoxymethoxy)-3-nitropentan-2-ol | <0.00001 | <0.00001 | <0.00001 |
| 5-methylsulfanyl-3H-1,3,4-thiadiazole-2-thione | <0.00001 | <0.00001 | <0.00001 |
| [2-(2-aminopropoxy)-3-methylphenyl] methanol | <0.00001 | 0.00009 | <0.00001 |
| 2-phenylethanol | <0.00001 | 0.00043 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patrignani, F.; Siesto, G.; Gottardi, D.; Vigentini, I.; Toffanin, A.; Englezos, V.; Blaiotta, G.; Grieco, F.; Lanciotti, R.; Speranza, B.; et al. Impact of Two Commercial S. cerevisiae Strains on the Aroma Profiles of Different Regional Musts. Beverages 2022, 8, 59. https://doi.org/10.3390/beverages8040059
Patrignani F, Siesto G, Gottardi D, Vigentini I, Toffanin A, Englezos V, Blaiotta G, Grieco F, Lanciotti R, Speranza B, et al. Impact of Two Commercial S. cerevisiae Strains on the Aroma Profiles of Different Regional Musts. Beverages. 2022; 8(4):59. https://doi.org/10.3390/beverages8040059
Chicago/Turabian StylePatrignani, Francesca, Gabriella Siesto, Davide Gottardi, Ileana Vigentini, Annita Toffanin, Vasileios Englezos, Giuseppe Blaiotta, Francesco Grieco, Rosalba Lanciotti, Barbara Speranza, and et al. 2022. "Impact of Two Commercial S. cerevisiae Strains on the Aroma Profiles of Different Regional Musts" Beverages 8, no. 4: 59. https://doi.org/10.3390/beverages8040059