
Computational Modeling of Motile Cilia-Driven Cerebrospinal Flow in the Brain Ventricles of Zebrafish Embryo


Abstract
:1. Introduction
2. Materials and Methods
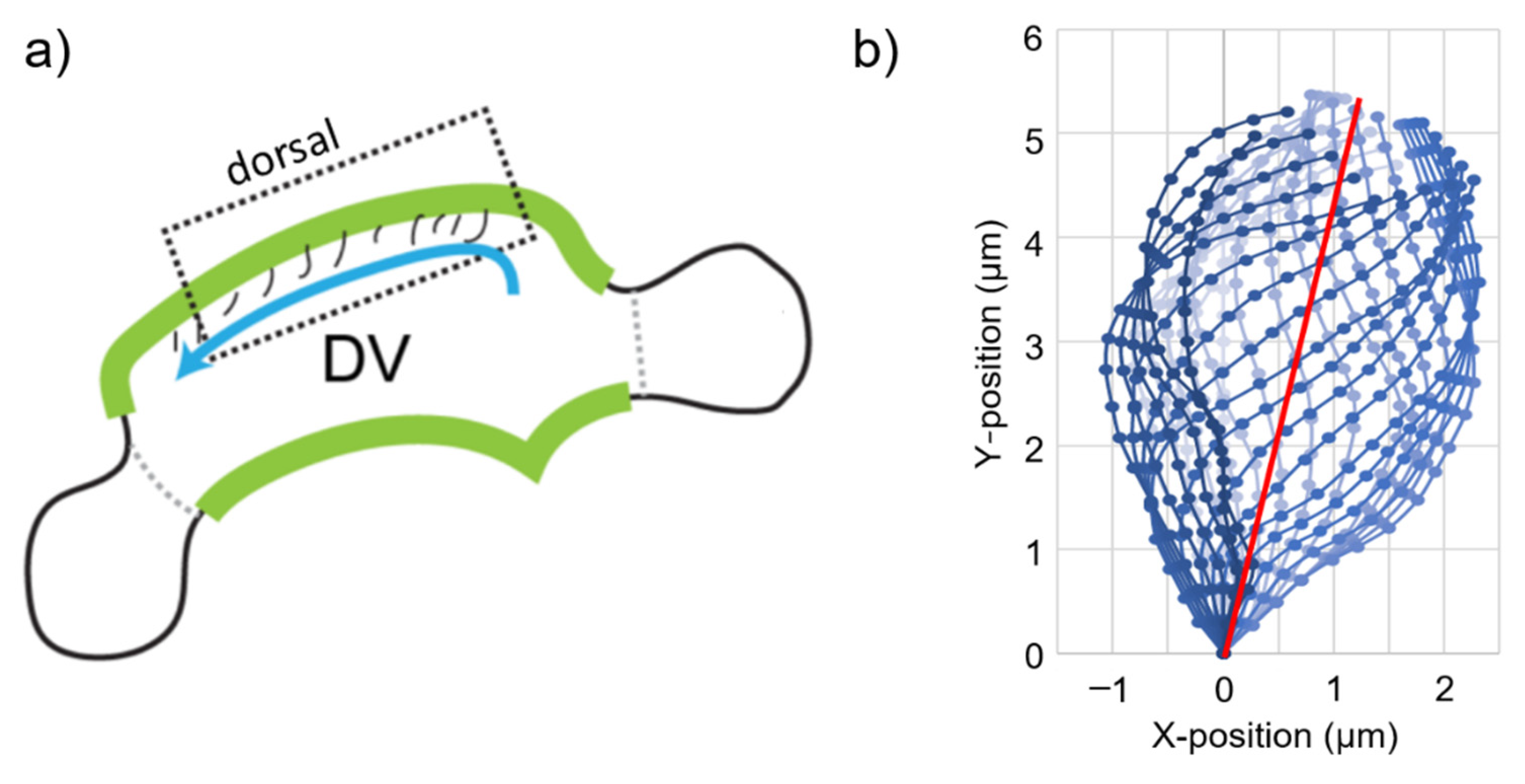
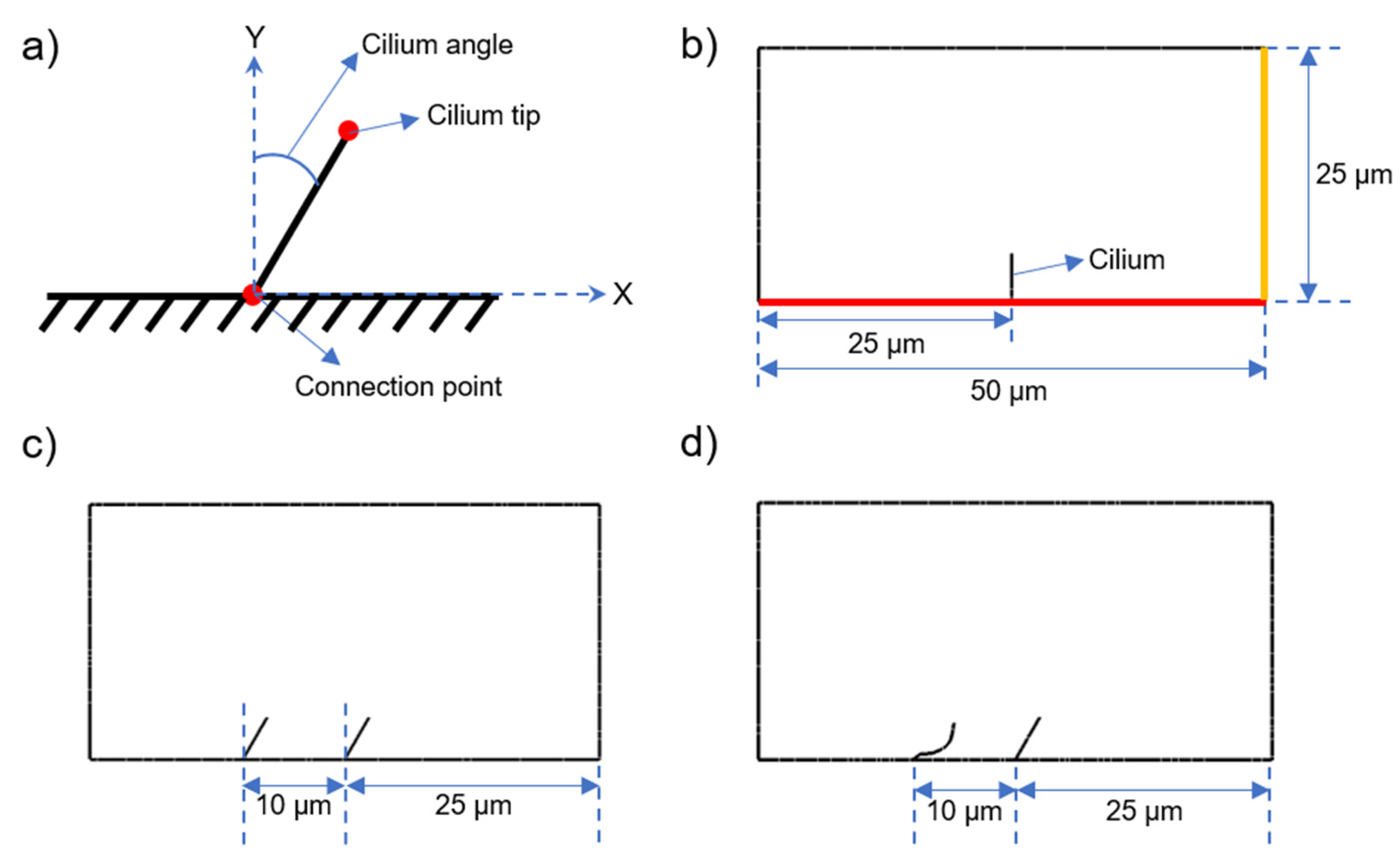
2.1. Determination of Cyclic Cilia Motion
2.2. Model Geometry of CFD Models
2.3. CFD Models
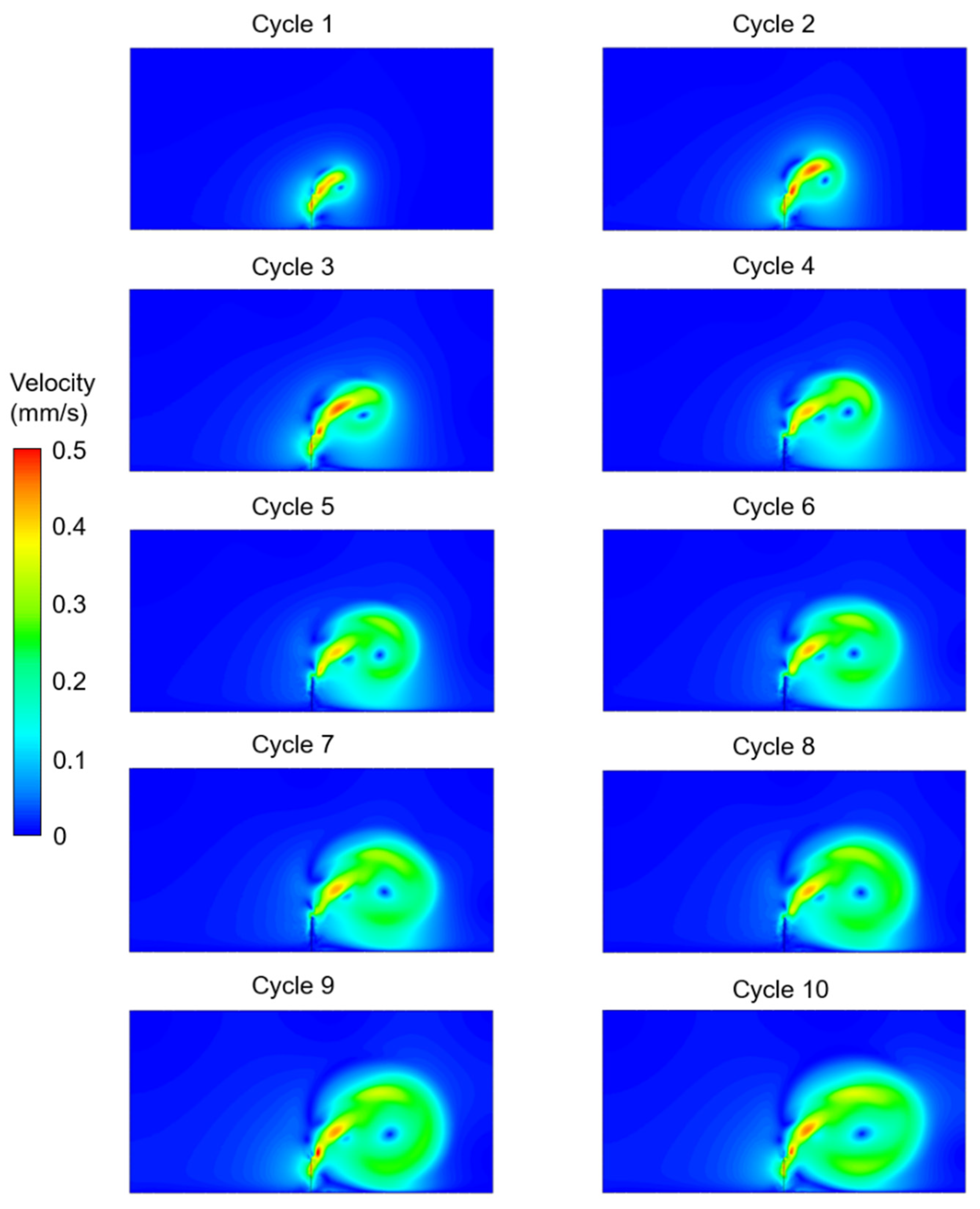
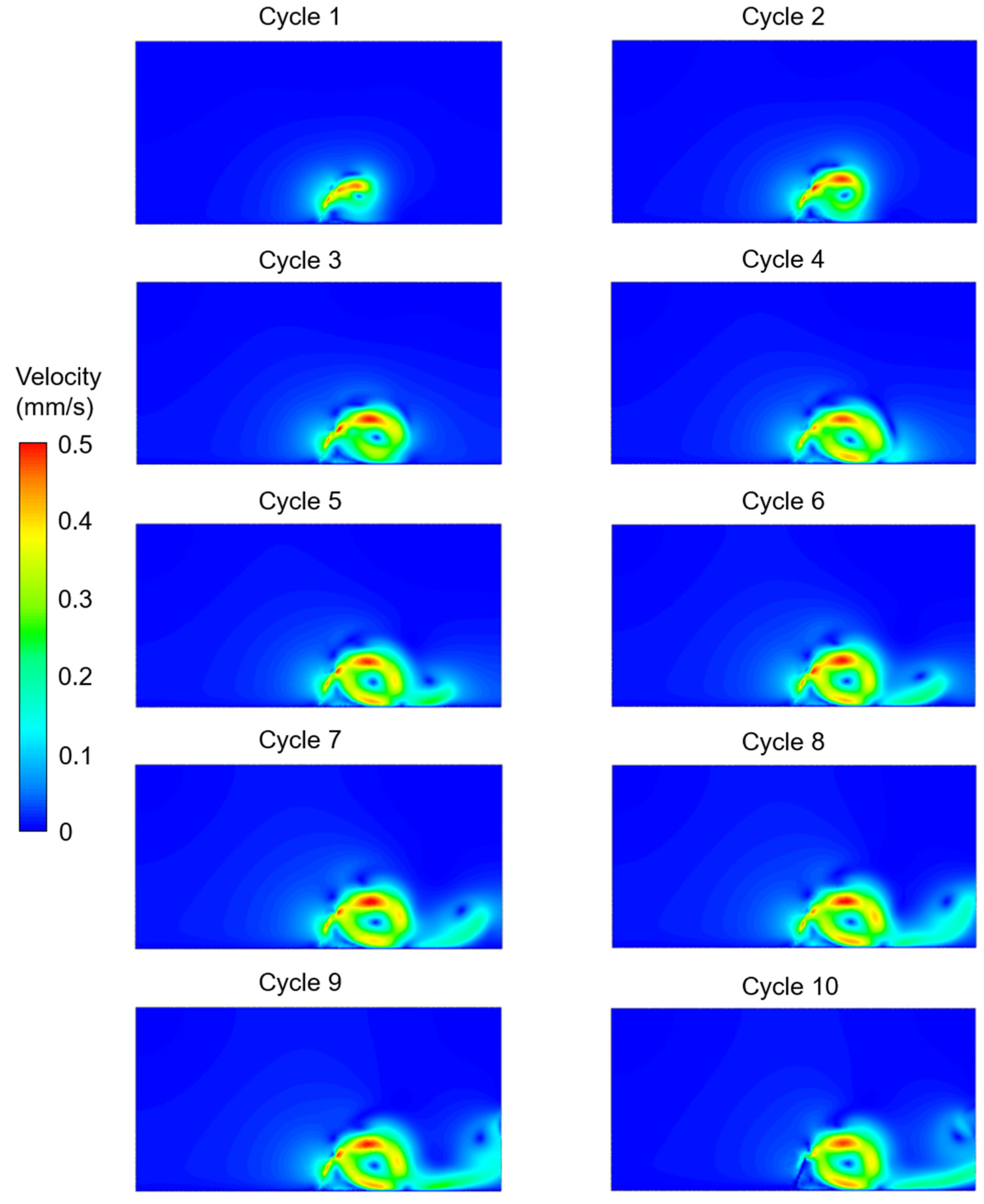
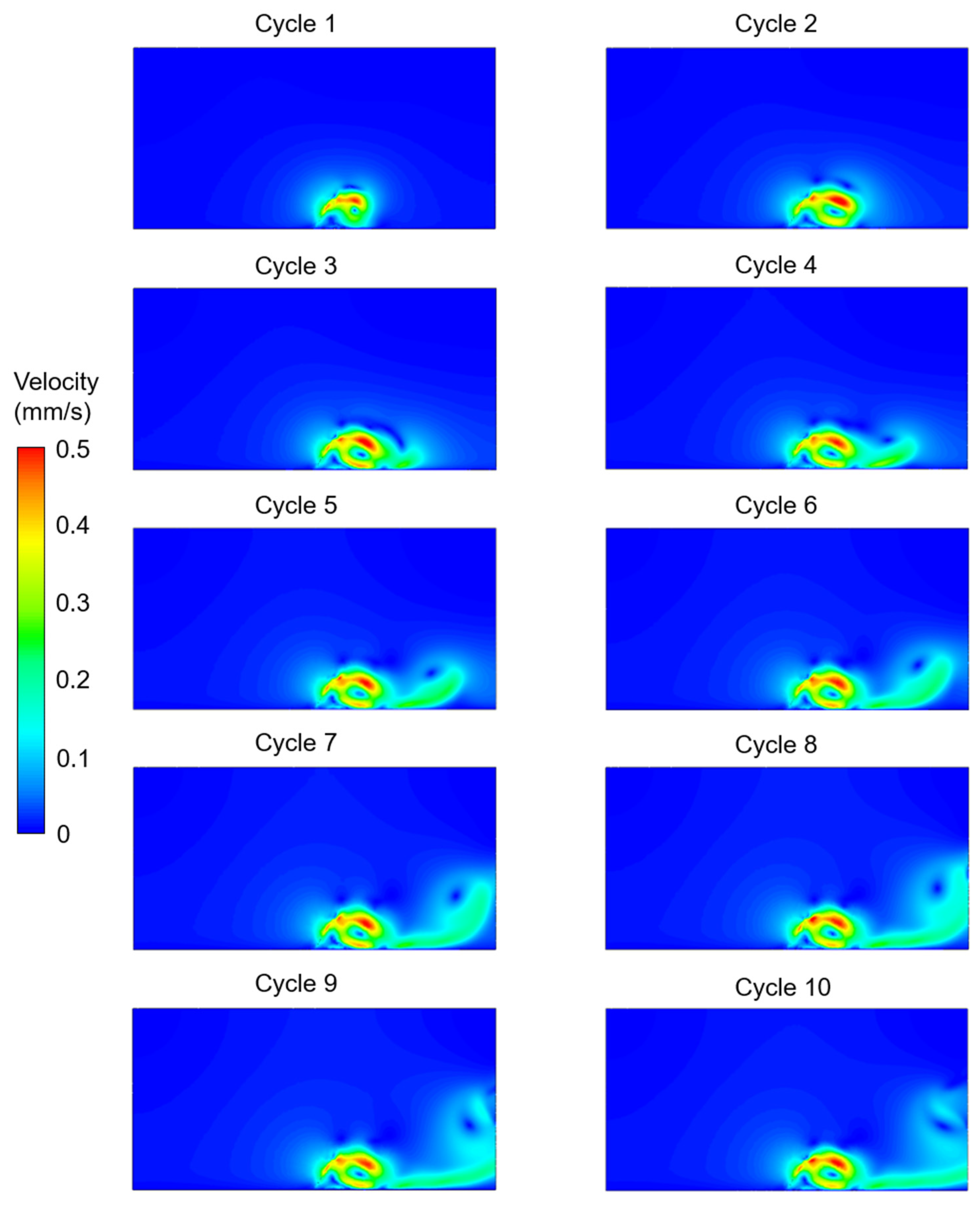
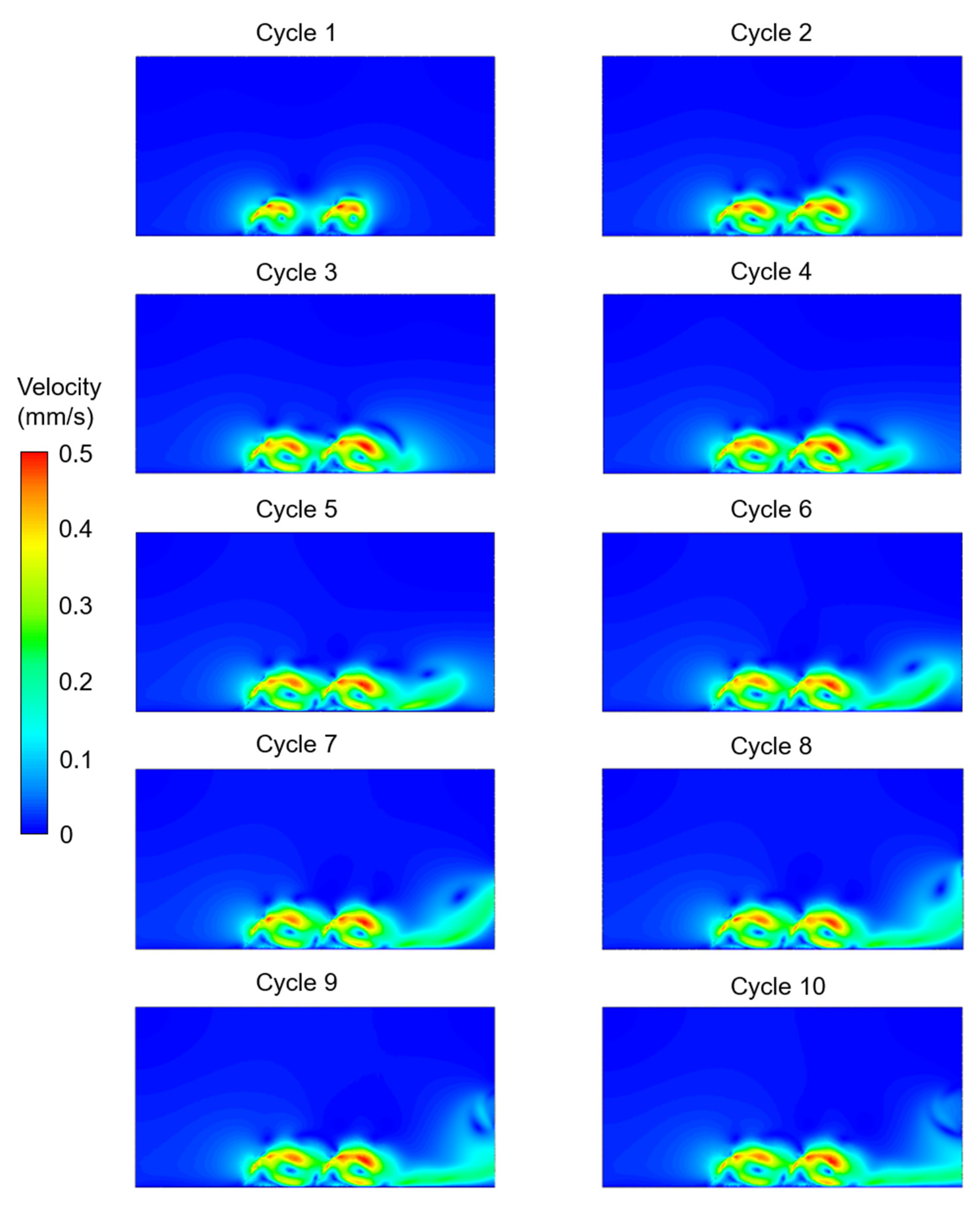
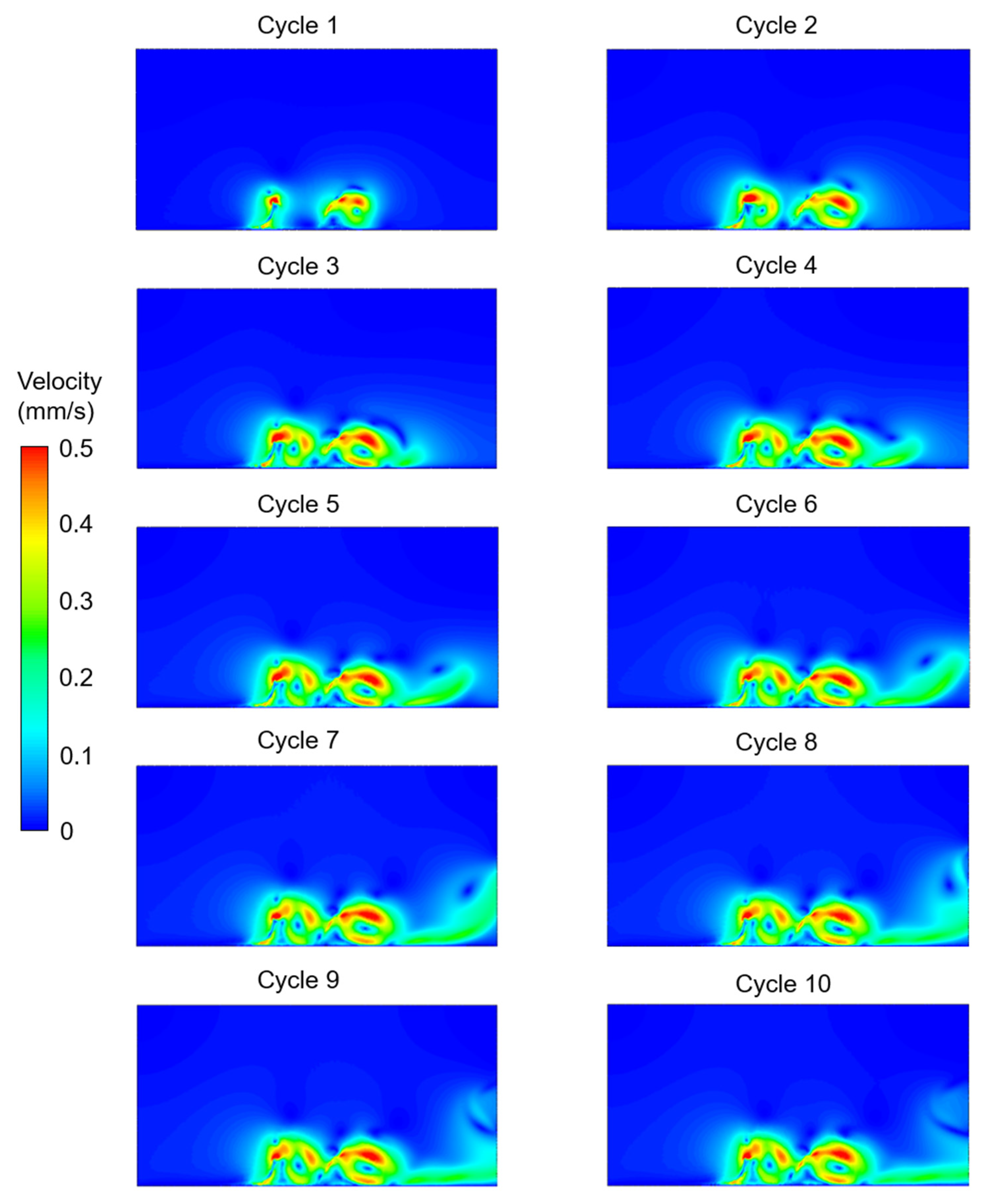
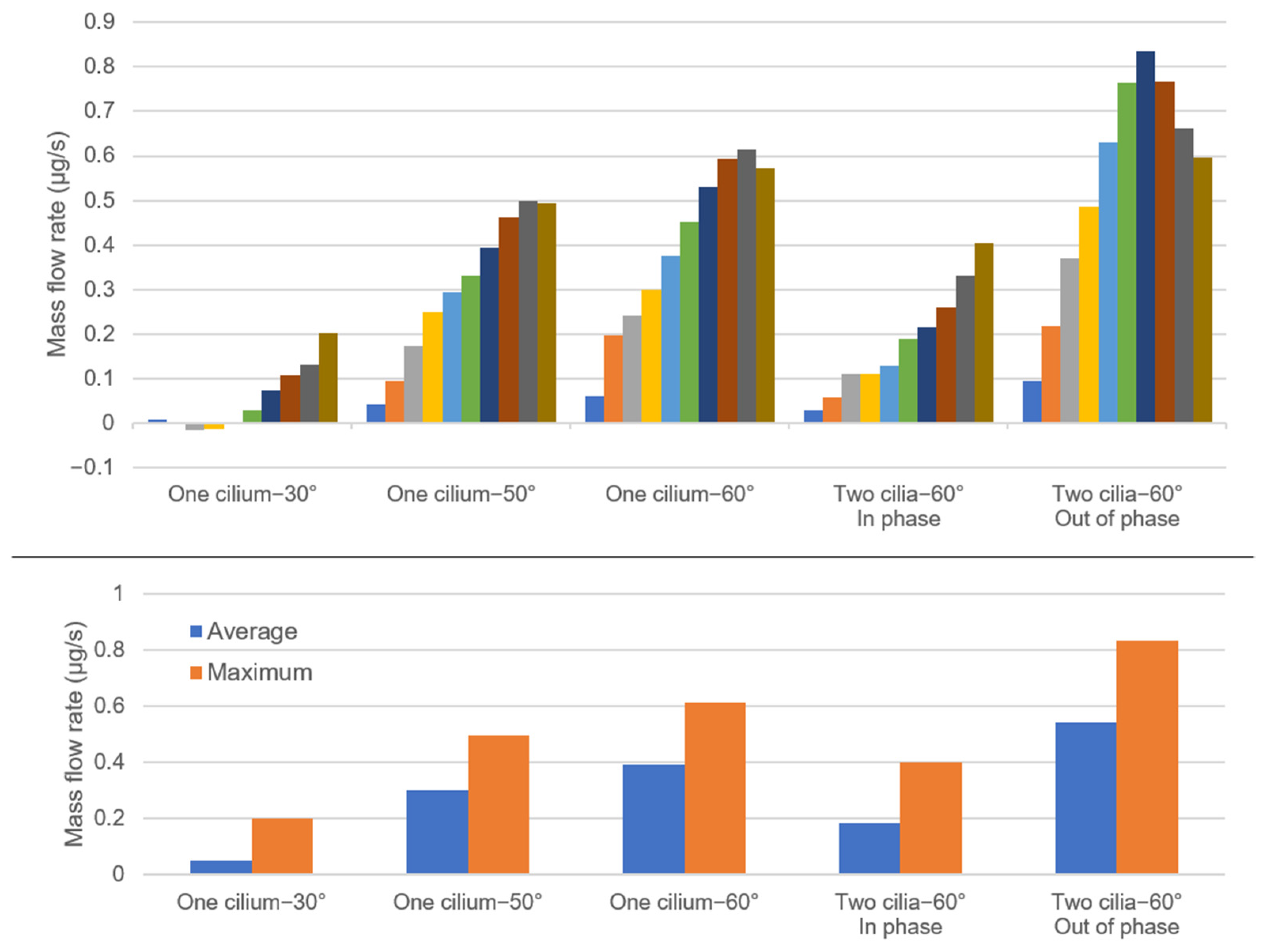
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, H.; Zhu, H.; Veerapaneni, S. Simulating Cilia-Driven Mixing and Transport in Complex Geometries. Phys. Rev. Fluids 2020, 5, 053103. [Google Scholar] [CrossRef]
- Ishikawa, T. Axoneme Structure from Motile Cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028076. [Google Scholar] [CrossRef]
- Satir, P.; Heuser, T.; Sale, W.S. A Structural Basis for How Motile Cilia Beat. Bioscience 2014, 64, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Geyer, V.F.; Howard, J.; Sartori, P. Ciliary Beating Patterns Map onto a Low-Dimensional Behavioural Space. Nat. Phys. 2022, 18, 332–337. [Google Scholar] [CrossRef]
- Gilpin, W.; Bull, M.S.; Prakash, M. The Multiscale Physics of Cilia and Flagella. Nat. Rev. Phys. 2020, 2, 74–88. [Google Scholar] [CrossRef]
- Satir, P.; Christensen, S.T. Overview of Structure and Function of Mammalian Cilia. Annu. Rev. Physiol. 2007, 69, 377–400. [Google Scholar] [CrossRef]
- Ueno, H.; Ishikawa, T.; Bui, K.H.; Gonda, K.; Ishikawa, T.; Yamaguchi, T. Mouse Respiratory Cilia with the Asymmetric Axonemal Structure on Sparsely Distributed Ciliary Cells Can Generate Overall Directional Flow. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 1081–1087. [Google Scholar] [CrossRef]
- Olstad, E.W.; Ringers, C.; Hansen, J.N.; Wens, A.; Brandt, C.; Wachten, D.; Yaksi, E.; Jurisch-Yaksi, N. Ciliary Beating Compartmentalizes Cerebrospinal Fluid Flow in the Brain and Regulates Ventricular Development. Curr. Biol. 2019, 29, 229–241. [Google Scholar] [CrossRef]
- Thouvenin, O.; Keiser, L.; Belarif, Y.C.; Carbo-Tano, M.; Verweij, F.; Jurisch-Yaksi, N.; Bardet, P.-L.; Van Niel, G.; Gallaire, F.; Wyart, C. Origin and Role of the Cerebrospinal Fluid Bidirectional Flow in the Central Canal. eLife 2020, 9, e47699. [Google Scholar] [CrossRef]
- Ringers, C.; Olstad, E.W.; Jurisch-Yaksi, N. The Role of Motile Cilia in the Development and Physiology of the Nervous System. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190156. [Google Scholar] [CrossRef] [Green Version]
- Wallmeier, J.; Nielsen, K.G.; Kuehni, C.E.; Lucas, J.S.; Leigh, M.W.; Zariwala, M.A.; Omran, H. Motile Ciliopathies. Nat. Rev. Dis. Prim. 2020, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Fauci, L.J.; Dillon, R. Biofluidmechanics of Reproduction. Annu. Rev. Fluid Mech. 2005, 38, 371–394. [Google Scholar] [CrossRef]
- Faubel, R.; Westendorf, C.; Bodenschatz, E.; Eichele, G. Cilia-Based Flow Network in the Brain Ventricles. Science 2016, 353, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, H.M.; Valente, E.M. Motile and Non-Motile Cilia in Human Pathology: From Function to Phenotypes. J. Pathol. 2017, 241, 294–309. [Google Scholar] [CrossRef]
- Boutin, C.; Kodjabachian, L. Biology of Multiciliated Cells. Curr. Opin. Genet. Dev. 2019, 56, 1–7. [Google Scholar] [CrossRef]
- Hirokawa, N.; Tanaka, Y.; Okada, Y.; Takeda, S. Nodal Flow and the Generation of Left-Right Asymmetry. Cell 2006, 125, 33–45. [Google Scholar] [CrossRef]
- McGrath, J.; Brueckner, M. Cilia are at the Heart of Vertebrate Left–Right Asymmetry. Curr. Opin. Genet. Dev. 2003, 13, 385–392. [Google Scholar] [CrossRef]
- Stannard, W.; O’Callaghan, C. Ciliary Function and the Role of Cilia in Clearance. J. Aerosol Med. 2006, 19, 110–115. [Google Scholar] [CrossRef]
- Brody, S.L.; Yan, X.H.; Wuerffel, M.K.; Song, S.K.; Shapiro, S.D. Ciliogenesis and Left–Right Axis Defects in Forkhead Factor HFH-4–Null Mice. Am. J. Respir. Cell Mol. Biol. 2000, 23, 45–51. [Google Scholar] [CrossRef]
- Nonaka, S.; Tanaka, Y.; Okada, Y.; Takeda, S.; Harada, A.; Kanai, Y.; Kido, M.; Hirokawa, N. Randomization of Left–Right Asymmetry due to Loss of Nodal Cilia Generating Leftward Flow of Extraembryonic Fluid in Mice Lacking KIF3B Motor Protein. Cell 1998, 95, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Okada, Y.; Takeda, S.; Tanaka, Y.; Belmonte, J.C.I.; Hirokawa, N. Mechanism of Nodal Flow: A Conserved Symmetry Breaking Event in Left-Right Axis Determination. Cell 2005, 121, 633–644. [Google Scholar] [CrossRef] [PubMed]
- D’Gama, P.P.; Qiu, T.; Cosacak, M.I.; Rayamajhi, D.; Konac, A.; Hansen, J.N.; Ringers, C.; Acuña-Hinrichsen, F.; Hui, S.P.; Olstad, E.W.; et al. Diversity and Function of Motile Ciliated Cell Types within Ependymal Lineages of the Zebrafish Brain. Cell Rep. 2021, 37, 109775. [Google Scholar] [CrossRef] [PubMed]
- Worthington, W.C., Jr.; Cathcart, R.S., 3rd; Cooper, P.; Goldring, I.; Klein, M. Ependymal Cilia: Distribution and Activity in the Adult Human Brain. Science 1963, 139, 221–222. [Google Scholar] [CrossRef]
- Hogg, C. Primary ciliary dyskinesia. In Ciliopathies: A Reference for Clinicians, 1st ed.; Kenny, T.D., Beales, P.L., Eds.; Oxford University Press: Oxford, UK, 2014; pp. 218–237. [Google Scholar]
- Braun, D.A.; Hildebrandt, F. Ciliopathies. Cold Spring Harb. Perspect. Biol. 2017, 9, a028191. [Google Scholar] [CrossRef]
- Tilley, A.E.; Walters, M.S.; Shaykhiev, R.; Crystal, R.G. Cilia Dysfunction in Lung Disease. Annu. Rev. Physiol. 2015, 77, 379–406. [Google Scholar] [CrossRef]
- Lukens, S.; Yang, X.; Fauci, L. Using Lagrangian Coherent Structures to Analyze Fluid Mixing by Cilia. Chaos Interdiscip. J. Nonlinear Sci. 2010, 20, 017511. [Google Scholar] [CrossRef] [PubMed]
- Jayathilake, P.G.; Le, D.V.; Tan, Z.; Lee, H.P.; Khoo, B.C. A Numerical Study of Muco-Ciliary Transport under the Condition of Diseased Cilia. Comput. Methods Biomech. Biomed. Eng. 2015, 18, 944–951. [Google Scholar] [CrossRef]
- Chatelin, R.; Poncet, P. A parametric study of mucociliary transport by numerical simulations of 3D non-homogeneous mucus. J. Biomech. 2016, 49, 1772–1780. [Google Scholar] [CrossRef]
- Juan, G.R.S.; Mathijssen, A.J.T.M.; He, M.; Jan, L.; Marshall, W.; Prakash, M. Multi-Scale Spatial Heterogeneity Enhances Particle Clearance in Airway Ciliary Arrays. Nat. Phys. 2020, 16, 958–964. [Google Scholar] [CrossRef]
- Buceta, J.; Ibañes, M.; Rasskin-Gutman, D.; Okada, Y.; Hirokawa, N.; Izpisúa-Belmonte, J.C. Nodal Cilia Dynamics and the Specification of the Left/Right Axis in Early Vertebrate Embryo Development. Biophys. J. 2005, 89, 2199–2209. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.J.; Montenegro-Johnson, T.D.; Lopes, S.S. Symmetry-Breaking Cilia-Driven Flow in Embryogenesis. Annu. Rev. Fluid Mech. 2019, 51, 105–128. [Google Scholar] [CrossRef]
- Ferreira, R.R.; Pakula, G.; Klaeyle, L.; Fukui, H.; Vilfan, A.; Supatto, W.; Vermot, J. Chiral Cilia Orientation in the Left–Right Organizer. Cell Rep. 2018, 25, 2008–2016.e4. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Yoshiba, S.; Watanabe, D.; Ikeuchi, S.; Goto, T.; Marshall, W.F.; Hamada, H. De Novo Formation of Left–Right Asymmetry by Posterior Tilt of Nodal Cilia. PLoS Biol. 2005, 3, e268. [Google Scholar] [CrossRef]
- Guirao, B.; Meunier, A.; Mortaud, S.; Aguilar, A.; Corsi, J.M.; Strehl, L.; Hirota, Y.; Desoeuvre, A.; Boutin, C.; Han, Y.G.; et al. Coupling Between Hydrodynamic Forces and Planar Cell Polarity Orients Mammalian Motile Cilia. Nat. Cell Biol. 2010, 12, 341–350. [Google Scholar] [CrossRef]
- Pellicciotta, N.; Hamilton, E.; Kotar, J.; Faucourt, M.; Delgehyr, N.; Spassky, N.; Cicuta, P. Entrainment of Mammalian Motile Cilia in the Brain with Hydrodynamic Forces. Proc. Natl. Acad. Sci. USA 2020, 117, 8315–8325. [Google Scholar] [CrossRef] [PubMed]
- Date, P.; Ackermann, P.; Furey, C.; Fink, I.B.; Jonas, S.; Khokha, M.K.; Kahle, K.T.; Deniz, E. Visualizing Flow in an Intact CSF Network Using Optical Coherence Tomography: Implications for Human Congenital Hydrocephalus. Sci. Rep. 2019, 9, 6196. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef]
- MacAulay, N.; Keep, R.F.; Zeuthen, T. Cerebrospinal Fluid Production by the Choroid Plexus: A Century of Barrier Research Revisited. Fluids Barriers CNS 2022, 19, 26. [Google Scholar] [CrossRef]
- Simon, M.J.; Iliff, J.J. Regulation of Cerebrospinal Fluid (CSF) Flow in Neurodegenerative, Neurovascular and Neuroinflammatory Disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2016, 1862, 442–451. [Google Scholar] [CrossRef]
- Dreha-Kulaczewski, S.; Joseph, A.A.; Merboldt, K.-D.; Ludwig, H.-C.; Gärtner, J.; Frahm, J. Inspiration Is the Major Regulator of Human CSF Flow. J. Neurosci. 2015, 35, 2485–2491. [Google Scholar] [CrossRef]
- Proulx, S.T. Cerebrospinal fluid outflow: A review of the Historical and Contemporary Evidence for Arachnoid Villi, Perineural Routes, and Dural Lymphatics. Cell Mol. Life Sci. 2021, 78, 2429–2457. [Google Scholar] [CrossRef] [PubMed]
- Siyahhan, B.; Knobloch, V.; de Zelicourt, D.; Asgari, M.; Daners, M.S.; Poulikakos, D.; Kurtcuoglu, V. Flow Induced by Ependymal Cilia Dominates Near-Wall Cerebrospinal Fluid Dynamics in the Lateral Ventricles. J. R. Soc. Interface 2014, 11, 20131189. [Google Scholar] [CrossRef] [PubMed]
- Del Bigio, M.R. Ependymal Cells: Biology and Pathology. Acta Neuropathol. 2010, 119, 55–73. [Google Scholar] [CrossRef]
- Paul, A.; Chaker, Z.; Doetsch, F. Hypothalamic Regulation of Regionally Distinct Adult Neural Stem Cells and Neurogenesis. Science 2017, 356, 1383–1386. [Google Scholar] [CrossRef]
- Sakka, L.; Coll, G.; Chazal, J. Anatomy and Physiology of Cerebrospinal Fluid. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2011, 128, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Meunier, A.; Azimzadeh, J. Multiciliated Cells in Animals. Cold Spring Harb. Perspect. Biol. 2016, 8, a028233. [Google Scholar] [CrossRef]
- Brooks, E.R.; Wallingford, J.B. Multiciliated Cells. Curr. Biol. 2014, 24, R973–R982. [Google Scholar] [CrossRef]
- Reiten, I.; Uslu, F.E.; Fore, S.; Pelgrims, R.; Ringers, C.; Verdugo, C.D.; Hoffman, M.; Lal, P.; Kawakami, K.; Pekkan, K.; et al. Motile-Cilia-Mediated Flow Improves Sensitivity and Temporal Resolution of Olfactory Computations. Curr. Biol. 2016, 27, 166–174. [Google Scholar] [CrossRef]
- Machemer, H. Ciliary Activity and the Origin of Metachrony in Paramecium: Effects of Increased Viscosity. J. Exp. Biol. 1972, 57, 239–259. [Google Scholar] [CrossRef]
- Vanaki, S.M.; Holmes, D.; Jayathilake, P.G.; Brown, R. Three-Dimensional Numerical Analysis of Periciliary Liquid Layer: Ciliary Abnormalities in Respiratory Diseases. Appl. Sci. 2019, 9, 4033. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.S.; Marquez, J.; Date, P.P.; Ventrella, R.; Mitchell, B.J.; Khokha, M.K. Mechanical Stretch Scales Centriole Number to Apical Area via Piezo1 in Multiciliated Cells. eLife 2021, 10, e66076. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.; Stubbs, J.L.; Huisman, F.; Taborek, P.; Yu, C.; Kintner, C. The PCP Pathway Instructs the Planar Orientation of Ciliated Cells in the Xenopus Larval Skin. Curr. Biol. 2009, 19, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Nawroth, J.C.; Guo, H.; Koch, E.; Heath-Heckman, E.A.C.; Hermanson, J.C.; Ruby, E.G.; Dabiri, J.O.; Kanso, E.; McFall-Ngai, M. Motile Cilia Create Fluid-Mechanical Microhabitats for the Active recruitment of the Host Microbiome. Proc. Natl. Acad. Sci. USA 2017, 114, 9510–9516. [Google Scholar] [CrossRef]
- Ringers, C.; Bialonski, S.; Hansen, J.N.; Ege, M.; Friedrich, B.M.; Jurisch-Yaksi, N. Locally Synchronized Ciliary Domains and Tissue-Scale Cilia Alignment underlie Global Metachronal Wave Patterns. bioRxiv 2021. [Google Scholar] [CrossRef]
- Ferreira, R.R.; Vilfan, A.; Jülicher, F.; Supatto, W.; Vermot, J. Physical Limits of Flow Sensing in the Left-Right Organizer. eLife 2017, 6, e25078. [Google Scholar] [CrossRef] [PubMed]
- Solovev, A.; Friedrich, B.M. Synchronization in cilia carpets: Multiple Metachronal Waves are Stable, but One Wave Dominates. New J. Phys. 2022, 24, 013015. [Google Scholar] [CrossRef]
- Meng, F.; Bennett, R.R.; Uchida, N.; Golestanian, R. Conditions for Metachronal Coordination in Arrays of Model Cilia. Proc. Natl. Acad. Sci. USA 2021, 118, e2102828118. [Google Scholar] [CrossRef]
- Brumley, D.R.; Polin, M.; Pedley, T.J.; Goldstein, R.E. Metachronal Waves in the Flagellar Beating of Volvox and their HydroDynamic Origin. J. R. Soc. Interface 2015, 12, 20141358. [Google Scholar] [CrossRef]
- Elgeti, J.; Gompper, G. Emergence of Metachronal Waves in Cilia Arrays. Proc. Natl. Acad. Sci. USA 2013, 110, 4470–4475. [Google Scholar] [CrossRef]
- Guirao, B.; Joanny, J.-F. Spontaneous Creation of Macroscopic Flow and Metachronal Waves in an Array of Cilia. Biophys. J. 2007, 92, 1900–1917. [Google Scholar] [CrossRef] [Green Version]
- Alser, M.; Salman, H.E.; Naïja, A.; Seers, T.D.; Khan, T.; Yalcin, H.C. Blood Flow Disturbance and Morphological Alterations Following the Right Atrial Ligation in the Chick Embryo. Front. Physiol. 2022, 13, 499. [Google Scholar] [CrossRef] [PubMed]
- Salman, H.E.; Kamal, R.Y.; Hijazi, Z.M.; Yalcin, H.C. Hemodynamic and Structural Comparison of Human Fetal Heart Development Between Normally Growing and Hypoplastic Left Heart Syndrome-Diagnosed Hearts. Front. Physiol. 2022, 13, 856879. [Google Scholar] [CrossRef] [PubMed]
- Polanczyk, A.; Strzelecki, M.; Wozniak, T.; Szubert, W.; Stefanczyk, L. 3D Blood Vessels Reconstruction Based on Segmented CT Data for Further Simulations of Hemodynamic in Human Artery Branches. Found. Comput. Decis. Sci. 2017, 42, 359–371. [Google Scholar] [CrossRef]
- Polanczyk, A.; Piechota-Polanczyk, A.; Huk, I.; Neumayer, C.; Strzelecki, M. Computational Fluid Dynamics as an Engineering Tool for the Reconstruction of Endovascular Prosthesis Endoleaks. IEEE Access 2022, 10, 18873–18885. [Google Scholar] [CrossRef]
- Polanczyk, A.; Piechota-Polanczyk, A.; Huk, I.; Neumayer, C.; Balcer, J.; Strzelecki, M. Computational Fluid Dynamic Technique for Assessment of How Changing Character of Blood Flow and Different Value of Hct Influence Blood Hemodynamic in Dissected Aorta. Diagnostics 2021, 11, 1866. [Google Scholar] [CrossRef] [PubMed]
- Salman, H.E.; Ramazanli, B.; Yavuz, M.M.; Yalcin, H.C. Biomechanical Investigation of Disturbed Hemodynamics-Induced Tissue Degeneration in Abdominal Aortic Aneurysms Using Computational and Experimental Techniques. Front. Bioeng. Biotechnol. 2019, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Fame, R.M.; Chang, J.T.; Hong, A.; Aponte-Santiago, N.A.; Sive, H. Directional Cerebrospinal Fluid Movement between Brain Ventricles in Larval Zebrafish. Fluids Barriers CNS 2016, 13, 11. [Google Scholar] [CrossRef] [PubMed]
- Jurisch-Yaksi, N.; Yaksi, E.; Kizil, C. Radial Glia in the Zebrafish Brain: Functional, Structural, and Physiological Comparison with the Mammalian Glia. Glia 2020, 68, 2451–2470. [Google Scholar] [CrossRef]
- Fame, R.M.; Lehtinen, M.K. Emergence and Developmental Roles of the Cerebrospinal Fluid System. Dev. Cell 2020, 52, 261–275. [Google Scholar] [CrossRef]
- Eichele, G.; Bodenschatz, E.; Ditte, Z.; Günther, A.-K.; Kapoor, S.; Wang, Y.; Westendorf, C. Cilia-Driven Flows in the Brain Third Ventricle. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190154. [Google Scholar] [CrossRef] [Green Version]
- Fame, R.M.; Cortés-Campos, C.; Sive, H.L. Brain Ventricular System and Cerebrospinal Fluid Development and Function: Light at the End of the Tube. BioEssays 2020, 42, 1900186. [Google Scholar] [CrossRef] [PubMed]
- Khaderi, S.; Hussong, J.; Westerweel, J.; Toonder, J.D.; Onck, P. Fluid Propulsion using Magnetically-Actuated Artificial Cilia—Experiments and Simulations. RSC Adv. 2013, 3, 12735–12742. [Google Scholar] [CrossRef]
- Zhu, P.; Chen, D.; Xu, Y. Simulation Study on the Mass Transport in PCL Based on the Ciliated Dynamic System of the Respiratory Tract. J. Phys. Conf. Ser. 2019, 1300, 012068. [Google Scholar] [CrossRef]
- Borovina, A.; Superina, S.; Voskas, D.; Ciruna, B. Vangl2 Directs the Posterior Tilting and Asymmetric Localization of Motile Primary cilia. Nat. Cell Biol. 2010, 12, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.N.; Rassmann, S.; Jikeli, J.F.; Wachten, D. SpermQ—A Simple Analysis Software to Comprehensively Study Flagellar Beating and Sperm Steering. bioRxiv 2018, 449173. [Google Scholar] [CrossRef]
- Lui, A.C.P.; Polis, T.Z.; Cicutti, N.J. Densities of Cerebrospinal Fluid and Spinal Anaesthetic Solutions in Surgical Patients at Body Temperature. Can. J. Anaesth. 1998, 45, 297–303. [Google Scholar] [CrossRef]
- Bloomfield, I.G.; Johnston, I.H.; Bilston, L.E. Effects of Proteins, Blood Cells and Glucose on the Viscosity of Cerebrospinal Fluid. Pediatr. Neurosurg. 1998, 28, 246–251. [Google Scholar] [CrossRef]
- Supatto, W.; Fraser, S.E.; Vermot, J. An All-Optical Approach for Probing Microscopic Flows in Living Embryos. Biophys. J. 2008, 95, L29–L31. [Google Scholar] [CrossRef]
- Hoffmann, F.; Cortez, R. Numerical Computation of Doubly-Periodic Stokes Flow Bounded by a Plane with Applications to Nodal Cilia. Commun. Comput. Phys. 2017, 22, 620–642. [Google Scholar] [CrossRef]
- Ding, Y.; Nawroth, J.C.; McFall-Ngai, M.J.; Kanso, E. Mixing and Transport by Ciliary Carpets: A Numerical Study. J. Fluid Mech. 2014, 743, 124–140. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quantity of Cilia | Cilium Tilt Angle | Phase between the Cilia | |
|---|---|---|---|
| Case 1 | 1 | 30° | |
| Case 2 | 1 | 50° | |
| Case 3 | 1 | 60° | |
| Case 4 | 2 | 60° | In phase |
| Case 5 | 2 | 60° | Out of phase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salman, H.E.; Jurisch-Yaksi, N.; Yalcin, H.C. Computational Modeling of Motile Cilia-Driven Cerebrospinal Flow in the Brain Ventricles of Zebrafish Embryo. Bioengineering 2022, 9, 421. https://doi.org/10.3390/bioengineering9090421
Salman HE, Jurisch-Yaksi N, Yalcin HC. Computational Modeling of Motile Cilia-Driven Cerebrospinal Flow in the Brain Ventricles of Zebrafish Embryo. Bioengineering. 2022; 9(9):421. https://doi.org/10.3390/bioengineering9090421
Chicago/Turabian StyleSalman, Huseyin Enes, Nathalie Jurisch-Yaksi, and Huseyin Cagatay Yalcin. 2022. "Computational Modeling of Motile Cilia-Driven Cerebrospinal Flow in the Brain Ventricles of Zebrafish Embryo" Bioengineering 9, no. 9: 421. https://doi.org/10.3390/bioengineering9090421