
Magnetic Nanoparticles as a Component of Peptide-Based DNA Delivery System for Suicide Gene Therapy of Uterine Leiomyoma



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Peptide Carrier, Expression Plasmids and Magnetic Nanoparticles
2.3. Formation of R6p-cRGD/DNA/MNPs Polyplexes
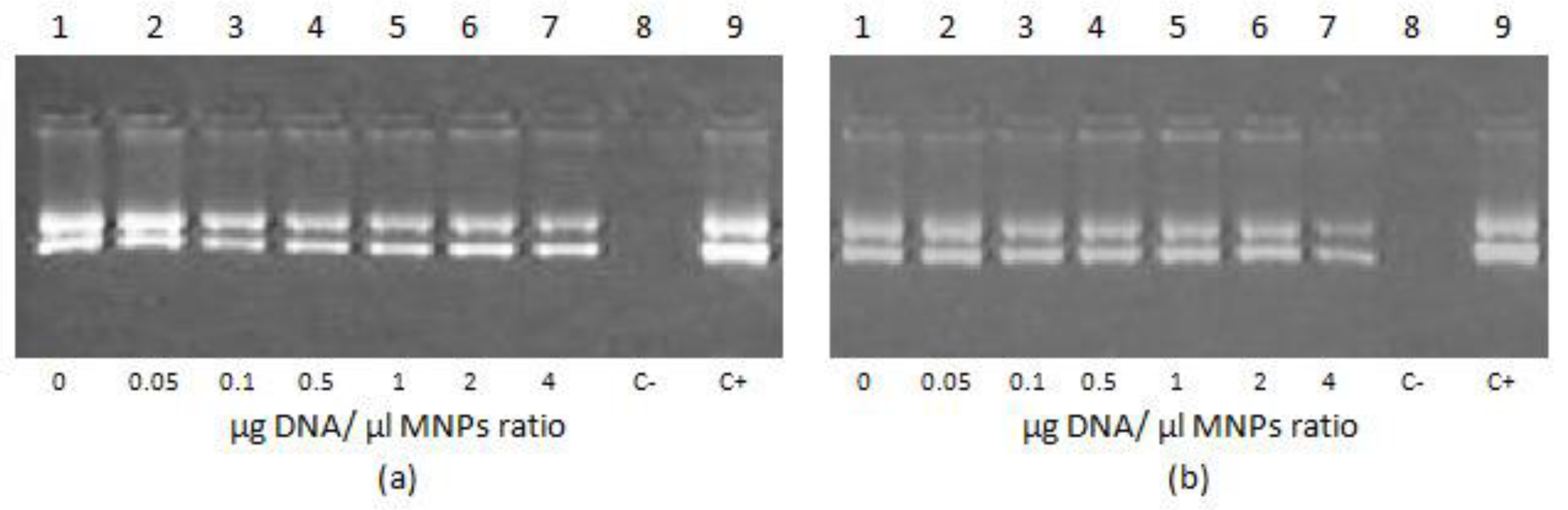
2.4. DNA Binding and DNAse I Protection Assay
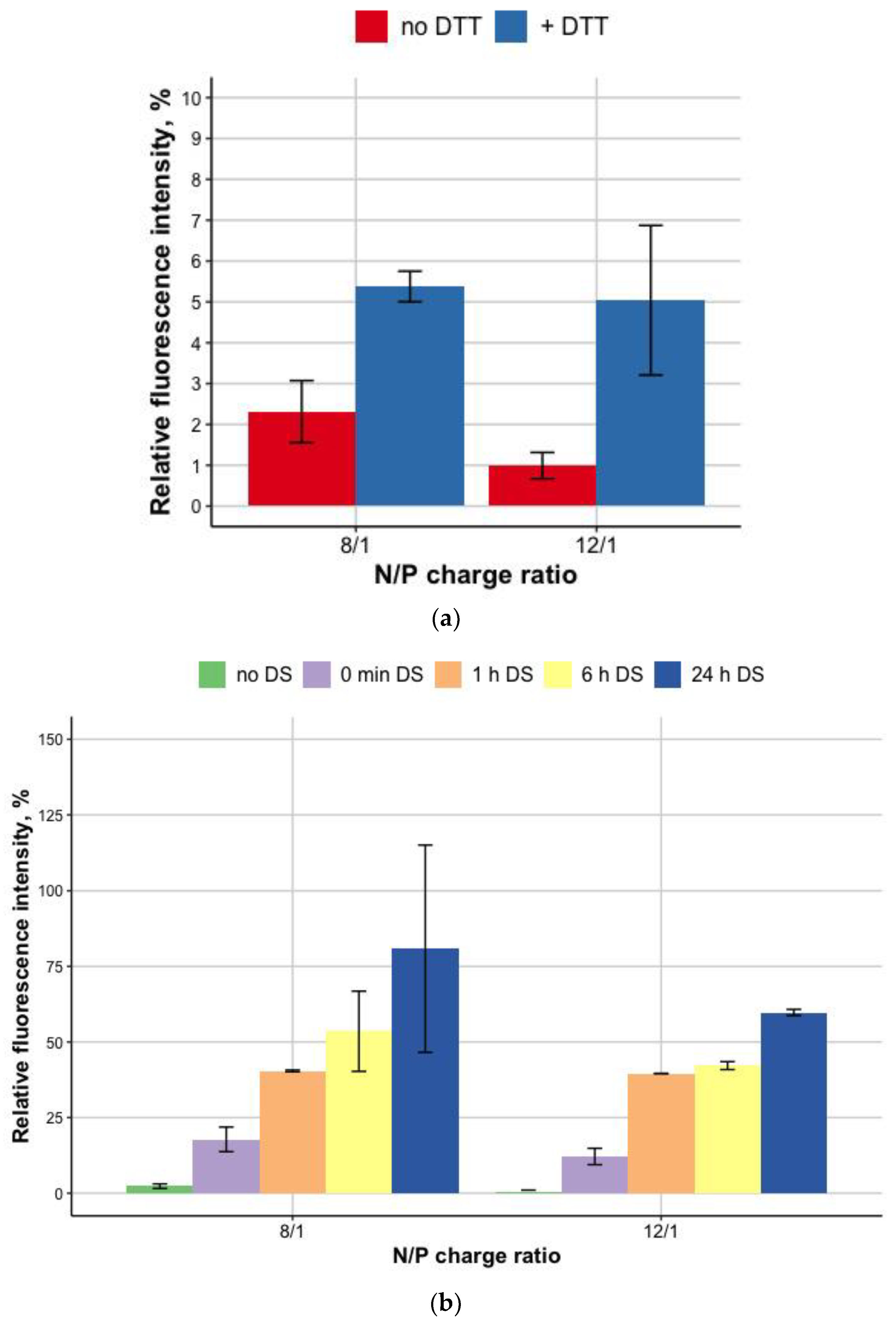
2.5. Relaxation of Carrier/DNA Polyplexes by Dextran-Sulfate and DTT Destabilization
2.6. Size and z-Potential Measurement of Peptide/DNA/MNPs Complexes
2.7. Transmission Electronic Microscopy
2.8. Transfection of PANC-1 Cells with DNA/Peptide Polyplexes with MNPs
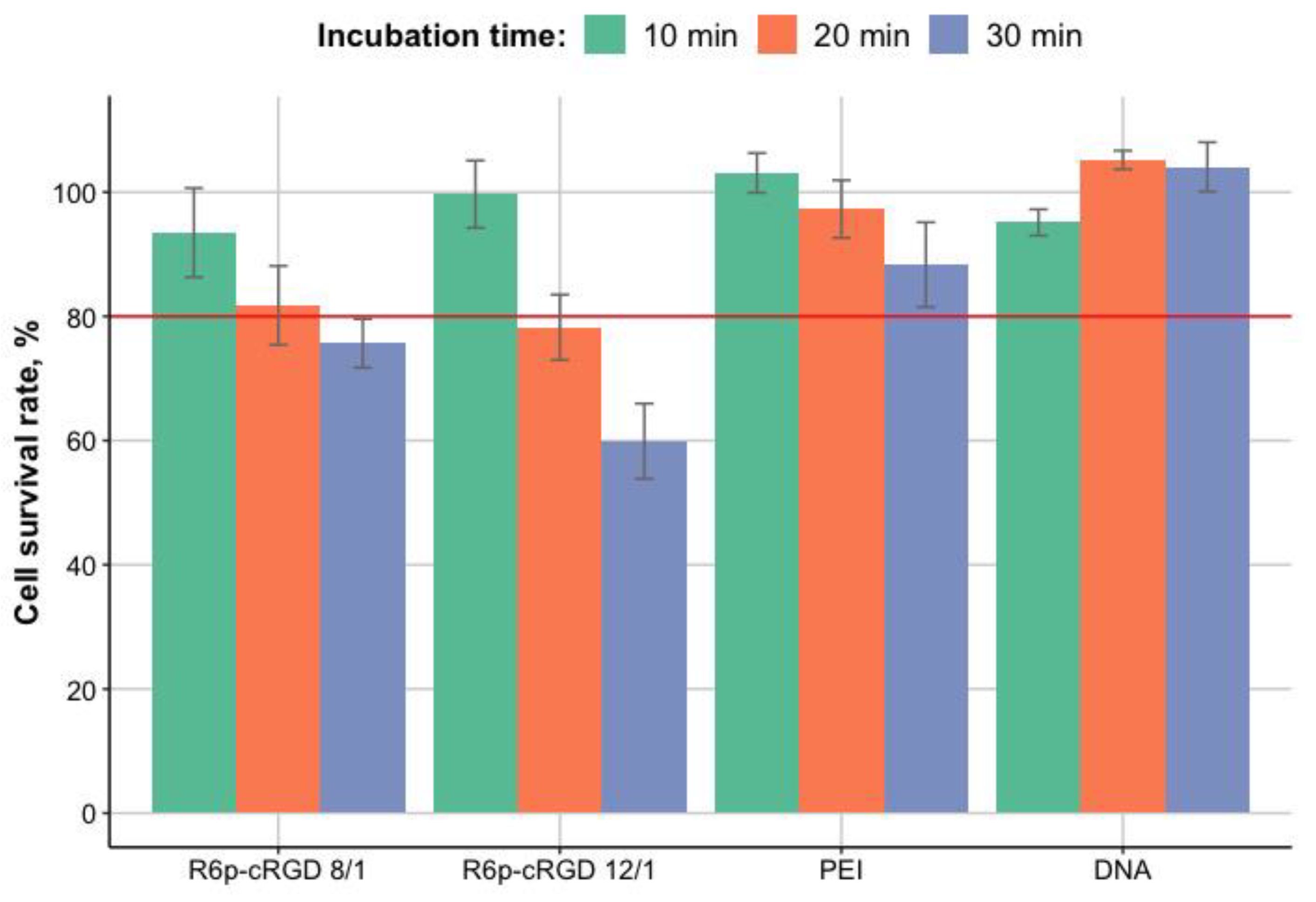
2.9. Cytotoxicity Assay
2.10. Suicide Gene Therapy of Primary UL Cells
2.11. Data Analysis and Statistical Comparisons
3. Results
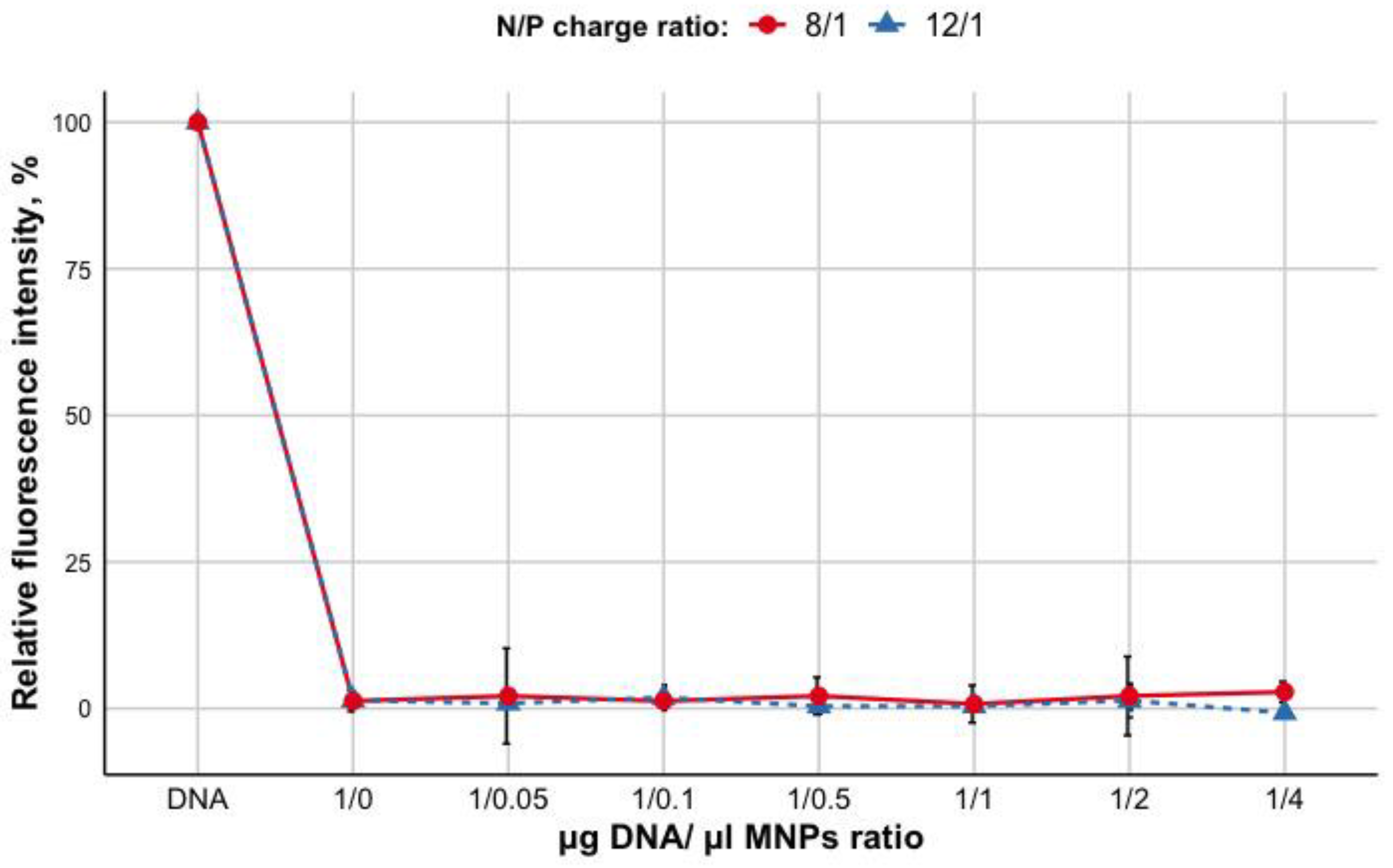
3.1. Evaluation of R6p-cRGD/DNA Polyplexes Stability and Protecting Properties after Non-Covalent Bindingwith MNPs
3.2. DNA Release after DTT and DS Treatment of the Polyplexes
3.3. Size and ʐ-Potential of the Carrier/DNA/MNP Polyplexes
3.4. Cytotoxicity Evaluation of R6p-cRGD/DNA/MNPs Polyplexes
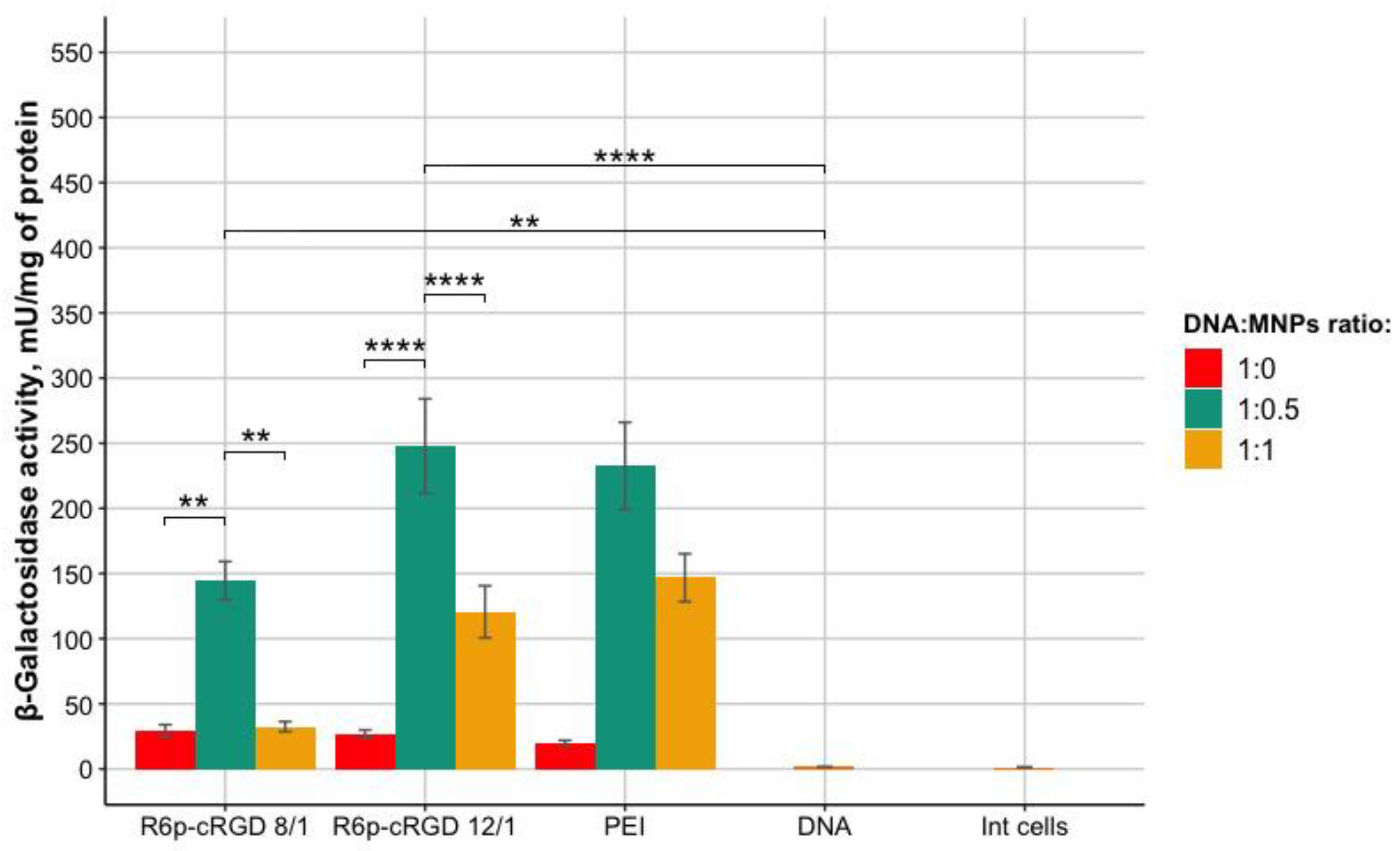
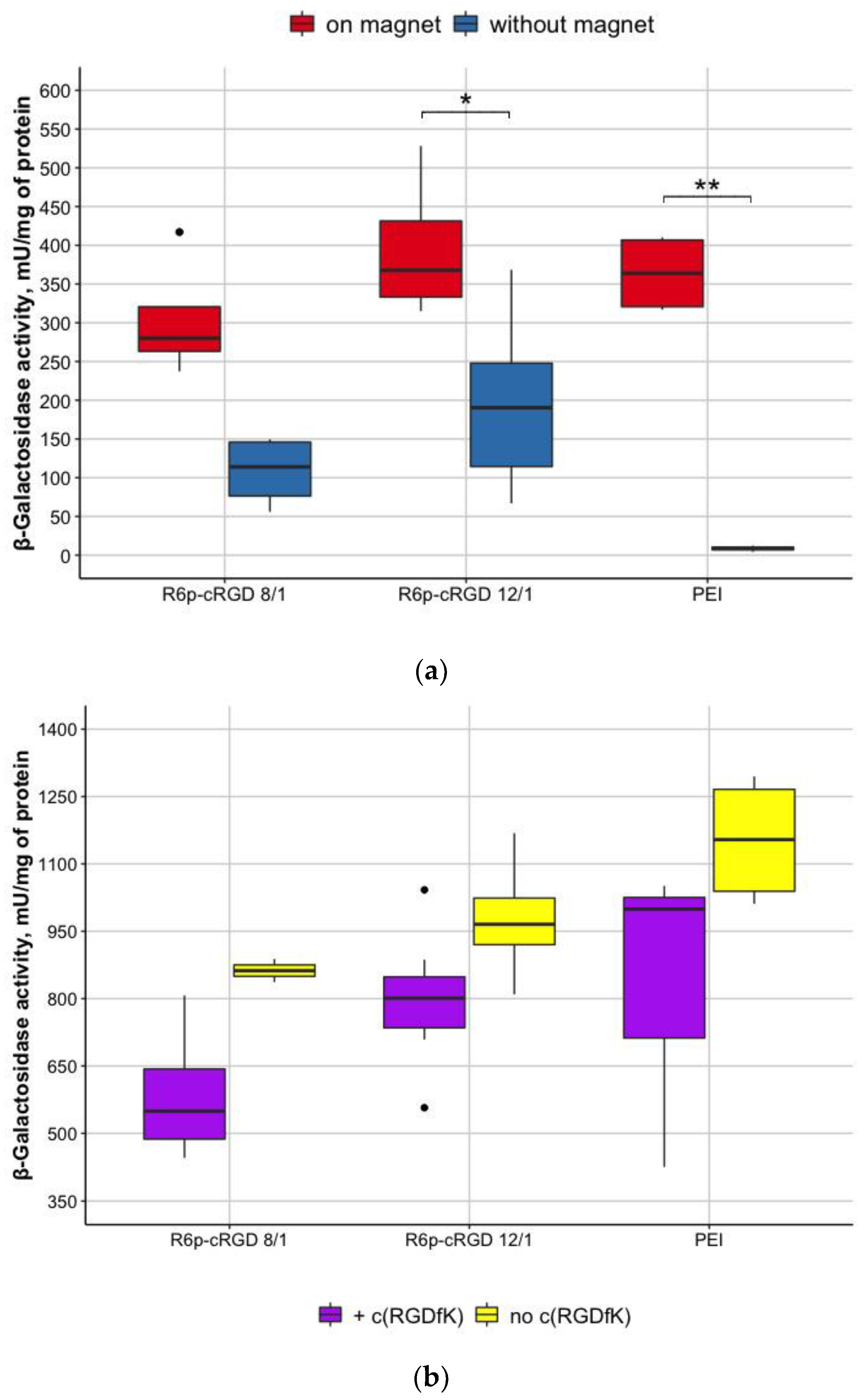
3.5. Evaluation of Magnetofection Efficacy Using R6p-cRGD/DNA/MNPs Polyplexes
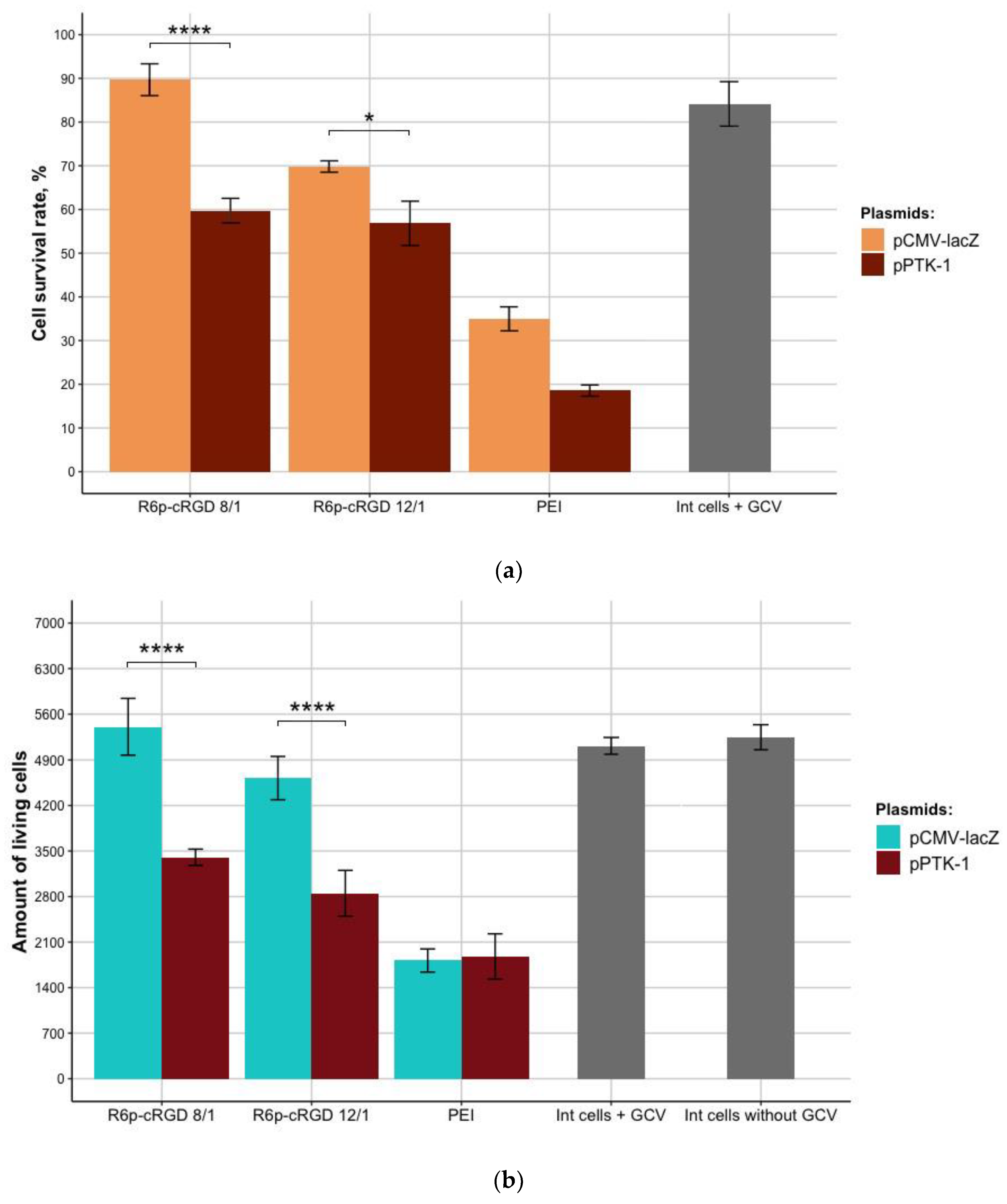
3.6. The Therapeutic Effect of pPTK-1/R6p-cRGD/MNPs Polyplexes after Ganciclovir (GCV) Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murase, E.; Siegelman, E.S.; Outwater, E.K.; Perez-Jaffe, L.A.; Tureck, R.W. Uterine Leiomyomas: Histopathologic Features, MR Imaging Findings, Differential Diagnosis, and Treatment. Radio Graph. 1999, 19, 1179–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, C.L. Uterine Fibroids: The Elephant in the Room. Science 2005, 308, 1589–1592. [Google Scholar] [CrossRef] [Green Version]
- Stewart, E.A. Uterine fibroids. Lancet 2001, 357, 293–298. [Google Scholar] [CrossRef]
- Packenham, J.P.; Du Manoir, S.; Schrock, E.; Risinger, J.I.; Dixon, D.; Denz, D.N.; Evans, J.A.C.; Berchuck, A.; Barrett, J.C.; Devereux, T.R.; et al. Analysis of Genetic Alterations in Uterine Leiomyomas and Leiomyosarcomas by Comparative Genomic Hybridization. Mol. Carcinog. 1997, 19, 273–279. [Google Scholar] [CrossRef]
- Ali, M.; Raslan, M.; Ciebiera, M.; Zaręba, K.; Al-Hendy, A. Current approaches to overcome the side effects of GnRH analogs in the treatment of patients with uterine fibroids. Expert Opin. Drug Saf. 2021, 1–10. [Google Scholar] [CrossRef]
- Sayed, N.; Allawadhi, P.; Khurana, A.; Singh, V.; Navik, U.; Pasumarthi, S.K.; Khurana, I.; Banothu, A.K.; Weiskirchen, R.; Bharani, K.K. Gene therapy: Comprehensive overview and therapeutic applications. Life Sci. 2022, 294, 120375. [Google Scholar] [CrossRef]
- Hassan, M.H.; Salama, S.A.; Zhang, D.; Arafa, H.M.M.; Hamada, F.M.A.; Fouad, H.; Walker, C.C.; Al-Hendy, A. Gene therapy targeting leiomyoma: Adenovirus-mediated delivery of dominant-negative estrogen receptor gene shrinks uterine tumors in Eker rat model. Fertil. Steril. 2010, 93, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Lazennec, G.; Alcorn, J.L.; Katzenellenbogen, B.S. Adenovirus-Mediated Delivery of a Dominant Negative Estrogen Receptor Gene Abrogates Estrogen-Stimulated Gene Expression and Breast Cancer Cell Proliferation. Mol. Endocrinol. 1999, 13, 969–980. [Google Scholar] [CrossRef]
- Navarro, S.A.; Carrillo, E.; Griñán-Lisón, C.; Martín, A.; Perán, M.; Marchal, J.A.; Boulaiz, H. Cancer suicide gene therapy: A patent review. Expert Opin. Ther. Pat. 2016, 26, 1095–1104. [Google Scholar] [CrossRef]
- Niculescu-Duvaz, I.; Springer, C.J. Introduction to the background, principles, and state of the art in suicide gene therapy. Mol. Biotechnol. 2005, 30, 71–88. [Google Scholar] [CrossRef]
- Duarte, S.; Carle, G.; Faneca, H.; de Lima, M.C.P.; Pierrefite-Carle, V. Suicide gene therapy in cancer: Where do we stand now? Cancer Lett. 2012, 324, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Curiel, D.T.; Rajaratnam, V.; Thota, C.; Al-Hendy, A. Targeting adenoviral vectors for enhanced gene therapy of uterine leiomyomas. Hum. Reprod. 2013, 28, 2398–2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelaziz, M.; Sherif, L.; ElKhiary, M.; Nair, S.; Shalaby, S.; Mohamed, S.; Eziba, N.; El-Lakany, M.; Curiel, D.; Ismail, N.; et al. Targeted Adenoviral Vector Demonstrates Enhanced Efficacy for In Vivo Gene Therapy of Uterine Leiomyoma. Reprod. Sci. 2016, 23, 464–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.S.; Ciavattini, A.; Petraglia, F.; Castellucci, M.; Ciarmela, P. Extracellular matrix in uterine leiomyoma pathogenesis: A potential target for future therapeutics. Hum. Reprod. Update 2018, 24, 59–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majidi, S.; Zeinali Sehrig, F.; Samiei, M.; Milani, M.; Abbasi, E.; Dadashzadeh, K.; Akbarzadeh, A. Magnetic nanoparticles: Applications in gene delivery and gene therapy. Artif. Cells Nanomed. Biotechnol. 2015, 44, 1186–1193. [Google Scholar] [CrossRef]
- Scherer, F.; Anton, M.; Schillinger, U.; Henke, J.; Bergemann, C.; Krü Ger, A.; Gänsbacher, B.; Plank, C. Magnetofection: Enhancing and targeting gene delivery by magnetic force in vitro and in vivo. Gene Ther. 2002, 9, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Dobson, J. Gene therapy progress and prospects: Magnetic nanoparticle-based gene delivery. Gene Ther. 2006, 13, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Blokpoel Ferreras, L.A.; Chan, S.Y.; Vazquez Reina, S.; Dixon, J.E. Rapidly Transducing and Spatially Localized Magnetofection Using Peptide-Mediated Non-Viral Gene Delivery Based on Iron Oxide Nanoparticles. ACS Appl. Nano Mater. 2021, 4, 167–181. [Google Scholar] [CrossRef]
- Shalaby, S.M.; Khater, M.K.; Perucho, A.M.; Mohamed, S.A.; Helwa, I.; Laknaur, A.; Lebedyeva, I.; Liu, Y.; Diamond, M.P.; Al-Hendy, A.A. Magnetic nanoparticles as a new approach to improve the efficacy of gene therapy against differentiated human uterine fibroid cells and tumor-initiating stem cells. Fertil. Steril. 2016, 105, 1638–1648.e8. [Google Scholar] [CrossRef] [Green Version]
- Egorova, A.; Shtykalova, S.; Selutin, A.; Shved, N.; Maretina, M.; Selkov, S.; Baranov, V.; Kiselev, A. Development of irgd-modified peptide carriers for suicide gene therapy of uterine leiomyoma. Pharmaceutics 2021, 13, 202. [Google Scholar] [CrossRef]
- Egorova, A.; Shtykalova, S.; Maretina, M.; Selutin, A.; Shved, N.; Deviatkin, D.; Selkov, S.; Baranov, V.; Kiselev, A. Polycondensed Peptide Carriers Modified with Cyclic RGD Ligand for Targeted Suicide Gene Delivery to Uterine Fibroid Cells. Int. J. Mol. Sci. 2022, 23, 1164. [Google Scholar] [CrossRef] [PubMed]
- Egorova, A.A.; Shtykalova, S.V.; Maretina, M.A.; Selyutin, A.V.; Shved, N.Y.; Krylova, N.V.; Ilina, A.V.; Pyankov, I.A.; Freund, S.A.; Selkov, S.A.; et al. Cys-Flanked Cationic Peptides For Cell Delivery of the Herpes Simplex Virus Thymidine Kinase Gene for Suicide Gene Therapy of Uterine Leiomyoma. Mol. Biol. 2020, 54, 436–448. [Google Scholar] [CrossRef]
- Shved, N.; Egorova, A.; Osinovskaya, N.; Kiselev, A. Development of primary monolayer cell model and organotypic model of uterine leiomyoma. Methods Protoc. 2022, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, A.V.; Il’ina, P.L.; Egorova, A.A.; Baranov, A.N.; Guryanov, I.A.; Bayanova, N.V.; Tarasenko, I.I.; Lesina, E.A.; Vlasov, G.P.; Baranov, V.S. Lysine dendrimers as vectors for delivery of genetic constructs to eukaryotic cells. Russ. J. Genet. 2007, 43, 593–600. [Google Scholar] [CrossRef]
- Egorova, A.; Bogacheva, M.; Shubina, A.; Baranov, V.; Kiselev, A. Development of a receptor-targeted gene delivery system using CXCR4 ligand-conjugated cross-linking peptides. J. Gene Med. 2014, 16, 336–351. [Google Scholar] [CrossRef]
- Niidome, T.; Ohmori, N.; Ichinose, A.; Wada, A.; Mihara, H.; Hirayama, T.; Aoyagi, H. Binding of Cationic α-Helical Peptides to Plasmid DNA and Their Gene Transfer Abilities into Cells. J. Biol. Chem. 1997, 272, 15307–15312. [Google Scholar] [CrossRef] [Green Version]
- Kiselev, A.; Egorova, A.; Laukkanen, A.; Baranov, V.; Urtti, A. Characterization of reducible peptide oligomers as carriers for gene delivery. Int. J. Pharm. 2013, 441, 736–747. [Google Scholar] [CrossRef]
- Ruponen, M.; Honkakoski, P.; Tammi, M.; Urtti, A. Cell-surface glycosaminoglycans inhibit cation-mediated gene transfer. J. Gene Med. 2004, 6, 405–414. [Google Scholar] [CrossRef]
- Huth, S.; Hoffmann, F.; von Gersdorff, K.; Laner, A.; Reinhardt, D.; Rosenecker, J.; Rudolph, C. Interaction of polyamine gene vectors with RNA leads to the dissociation of plasmid DNA-carrier complexes. J. Gene Med. 2006, 8, 1416–1424. [Google Scholar] [CrossRef]
- Mounkes, L.C.; Zhong, W.; Cipres-Palacin, G.; Heath, T.D.; Debs, R.J. Proteoglycans mediate cationic liposome-DNA complex-based gene delivery in vitro and in vivo. J. Biol. Chem. 1998, 273, 26164–26170. [Google Scholar] [CrossRef] [Green Version]
- Egorova, A.; Kiselev, A. Peptide modules for overcoming barriers of nucleic acids transport to cells. Curr. Top. Med. Chem. 2016, 16, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Moghimi, S.M.; Symonds, P.; Murray, J.C.; Hunter, A.C.; Debska, G.; Szewczyk, A. A two-stage poly(ethylenimine)-mediated cytotoxicity: Implications for gene transfer/therapy. Mol. Ther. 2005, 11, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Egorova, A.; Petrosyan, M.; Maretina, M.; Balashova, N.; Polyanskih, L.; Baranov, V.; Kiselev, A. Anti-angiogenic treatment of endometriosis via anti-VEGFA siRNA delivery by means of peptide-based carrier in a rat subcutaneous model. Gene Ther. 2018, 25, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Simari, R.D.; Zimmermann, E.M.; Christman, G.M. Nonviral Vector-Mediated Thymidine Kinase Gene Transfer and Ganciclovir Treatment in Leiomyoma Cells. Obstet. Gynecol. 1998, 91, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Salama, S.A.; Kamel, M.; Christman, G.; Wang, H.Q.; Fouad, H.M.; Al-Hendy, A. Gene Therapy of Uterine Leiomyoma: Adenovirus-Mediated Herpes Simplex Virus Thymidine Kinase/Ganciclovir Treatment Inhibits Growth of Human and Rat Leiomyoma Cells in vitro and in a Nude Mouse Model. Gynecol. Obstet. Investig. 2007, 63, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Su, D.; Liu, J.; Saha, R.; Wang, J.P. Magnetic nanoparticles in nanomedicine: A review of recent advances. Nanotechnology 2019, 30, 502003. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.-Y.; Liu, Z.-H.; Weng, W.-H.; Chang, C.-W. Magnetic nanocomplexes for gene delivery applications. J. Mater. Chem. B 2021, 9, 4267–4286. [Google Scholar] [CrossRef]
- Sizikov, A.A.; Nikitin, P.I.; Nikitin, M.P. Magnetofection In Vivo by Nanomagnetic Carriers Systemically Administered into the Bloodstream. Pharmaceutics 2021, 13, 1927. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carrier | Charge Ratio | Size (nm) ± S.D. | ʐ-Potential (mV) ± S.D. | References |
|---|---|---|---|---|
| R6p-cRGD | 8/1 | 104.5 ± 0.15 | 32 ± 1.2 | Adapted from ref. [21] |
| 12/1 | 178.4 ± 24.3 | 31.5 ± 0.6 | ||
| R6p-cRGD/MNPs | 8/1 | 1814.7 ± 175.5 | −5.3 ± 4.2 | Current study |
| 12/1 | 1481.0 ± 182.6 | 4.0 ± 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shtykalova, S.; Egorova, A.; Maretina, M.; Baranov, V.; Kiselev, A. Magnetic Nanoparticles as a Component of Peptide-Based DNA Delivery System for Suicide Gene Therapy of Uterine Leiomyoma. Bioengineering 2022, 9, 112. https://doi.org/10.3390/bioengineering9030112
Shtykalova S, Egorova A, Maretina M, Baranov V, Kiselev A. Magnetic Nanoparticles as a Component of Peptide-Based DNA Delivery System for Suicide Gene Therapy of Uterine Leiomyoma. Bioengineering. 2022; 9(3):112. https://doi.org/10.3390/bioengineering9030112
Chicago/Turabian StyleShtykalova, Sofia, Anna Egorova, Marianna Maretina, Vladislav Baranov, and Anton Kiselev. 2022. "Magnetic Nanoparticles as a Component of Peptide-Based DNA Delivery System for Suicide Gene Therapy of Uterine Leiomyoma" Bioengineering 9, no. 3: 112. https://doi.org/10.3390/bioengineering9030112