

The Osteogenic Potential of Falciform Ligament-Derived Stromal Cells—A Comparative Analysis between Two Osteogenic Induction Programs

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
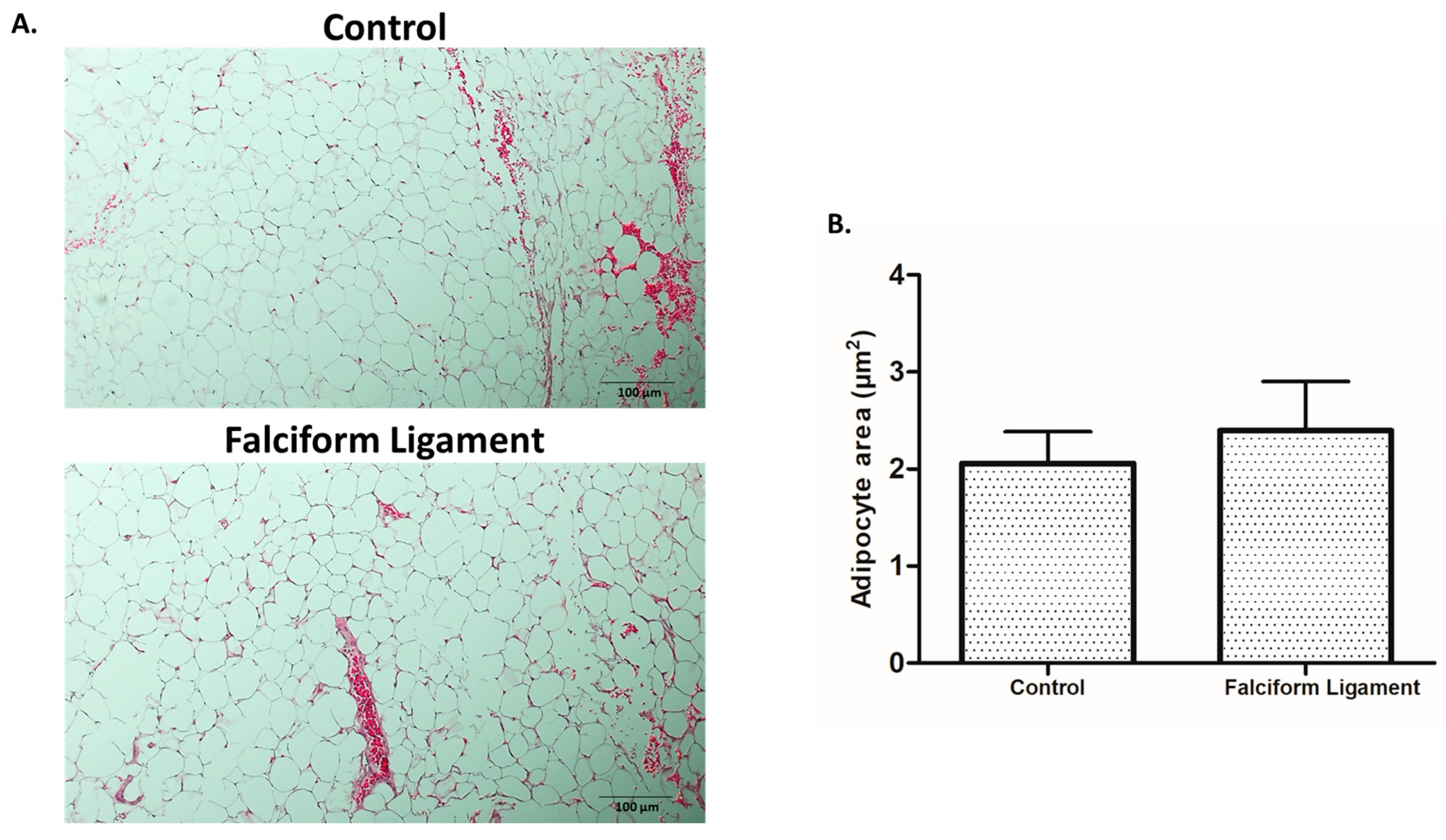
2.2. Adipose Tissue Histological Characterization
2.3. Isolation of ADSCs
2.4. Osteogenic Differentiation of ADSCs
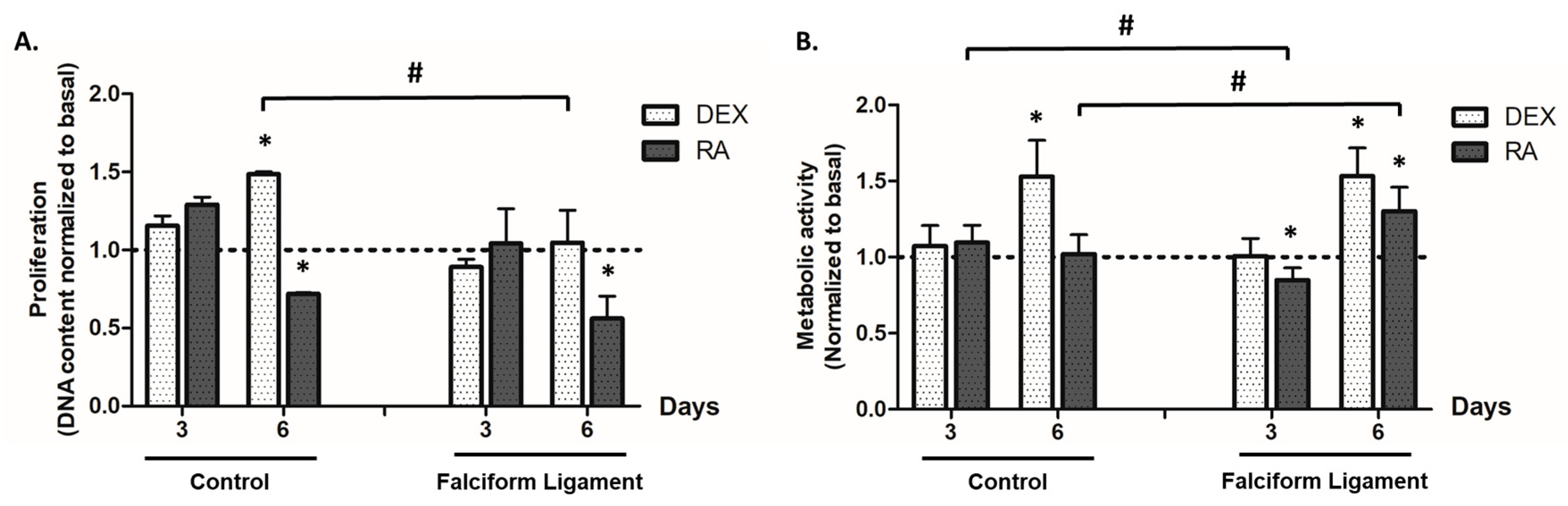
2.5. Cell Proliferation
2.6. Metabolic Activity
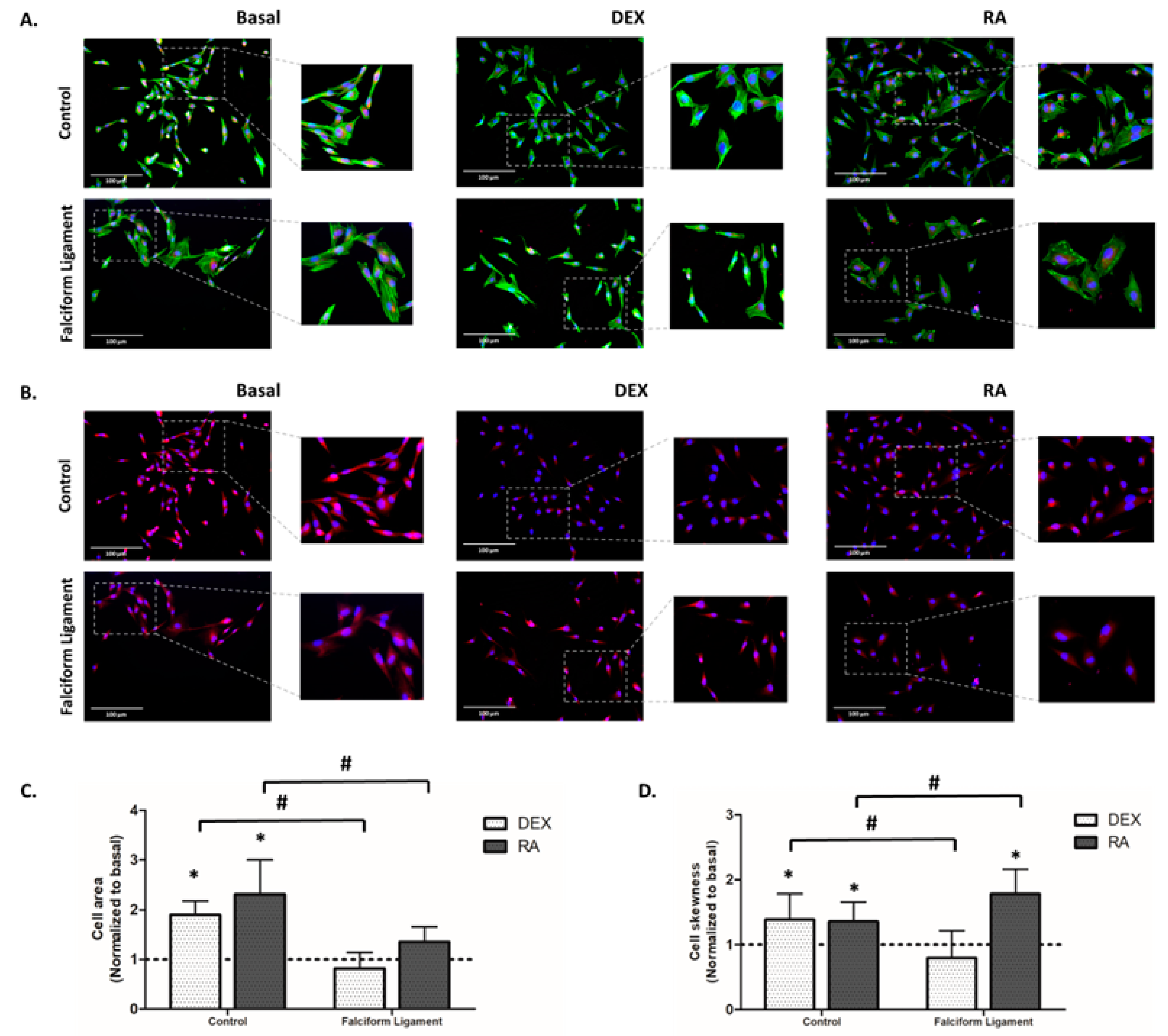
2.7. Immunostaining of F-Actin Cytoskeleton, Mitochondria, and Nucleus
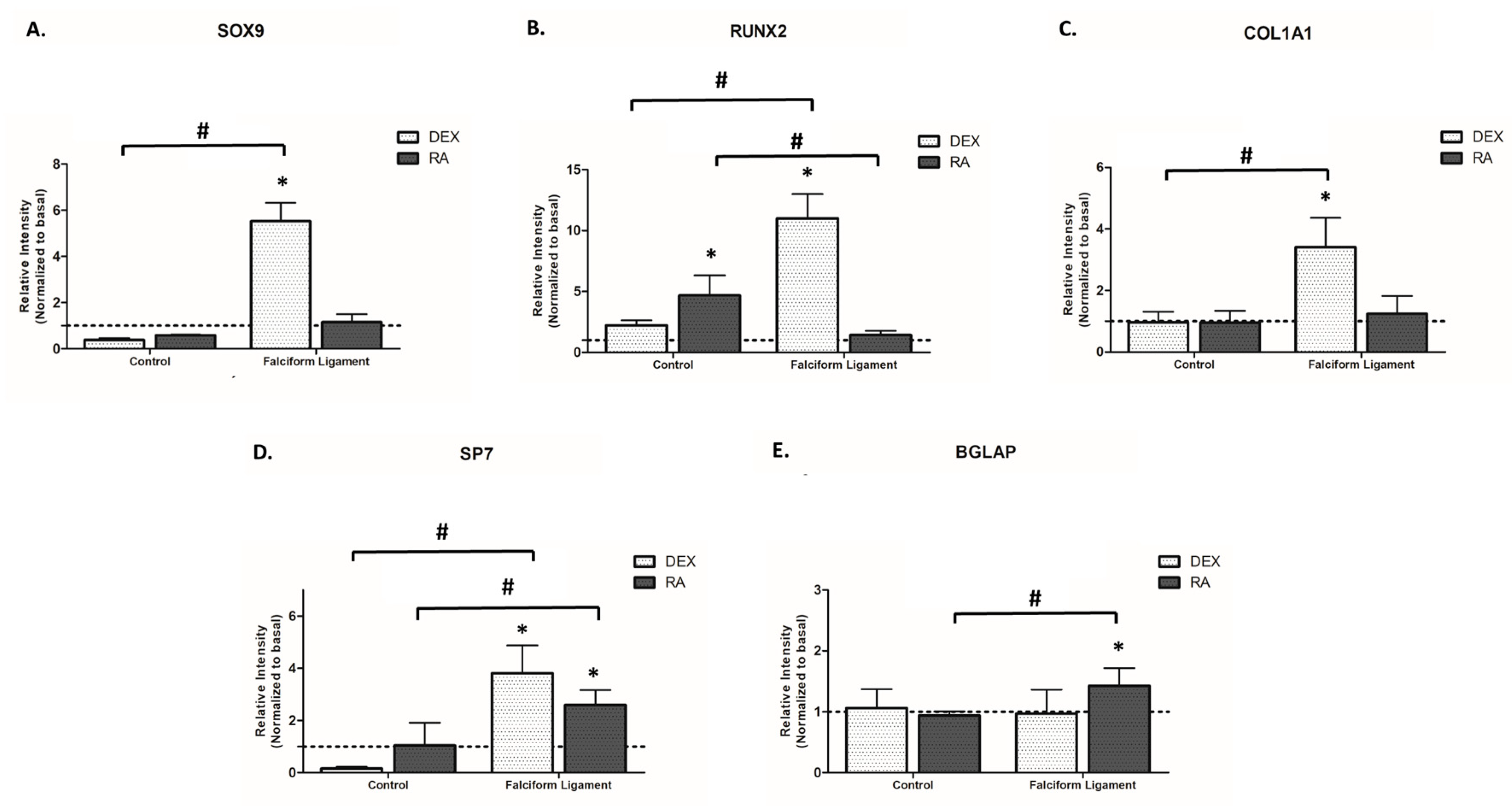
2.8. Gene Expression Assessment
2.9. Cytochemical Staining of Collagen
2.10. Cytochemical Staining of Alkaline Phosphatase
2.11. Statistical Analysis
3. Results
3.1. Histological Analysis of Adipose Tissue
3.2. Cell Proliferation
3.3. Metabolic Activity
3.4. Immunostaining of F-Actin Cytoskeleton, Mitochondrial, and Nucleus
3.5. Gene Expression Analysis
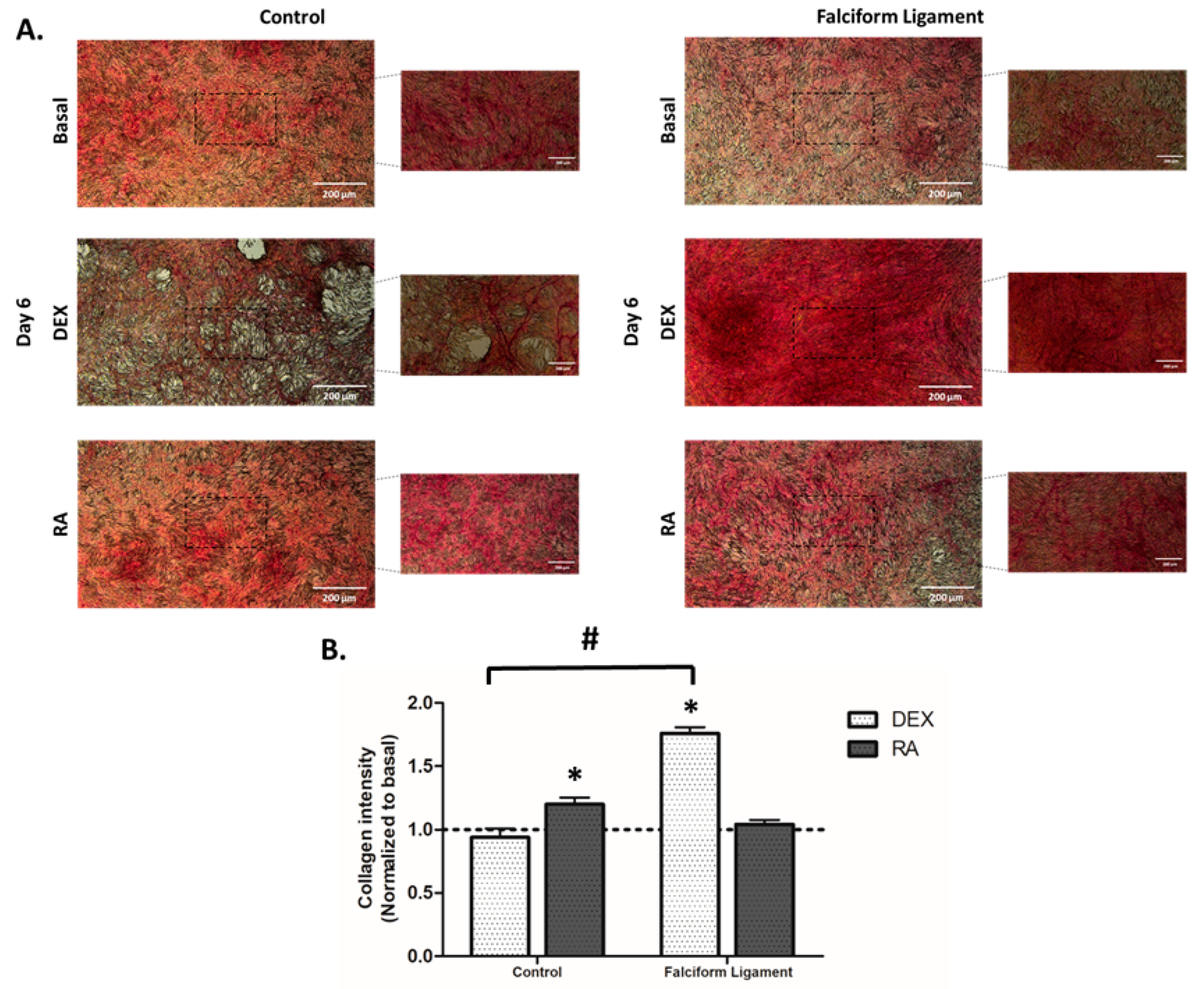
3.6. Cytochemical Staining of Collagen
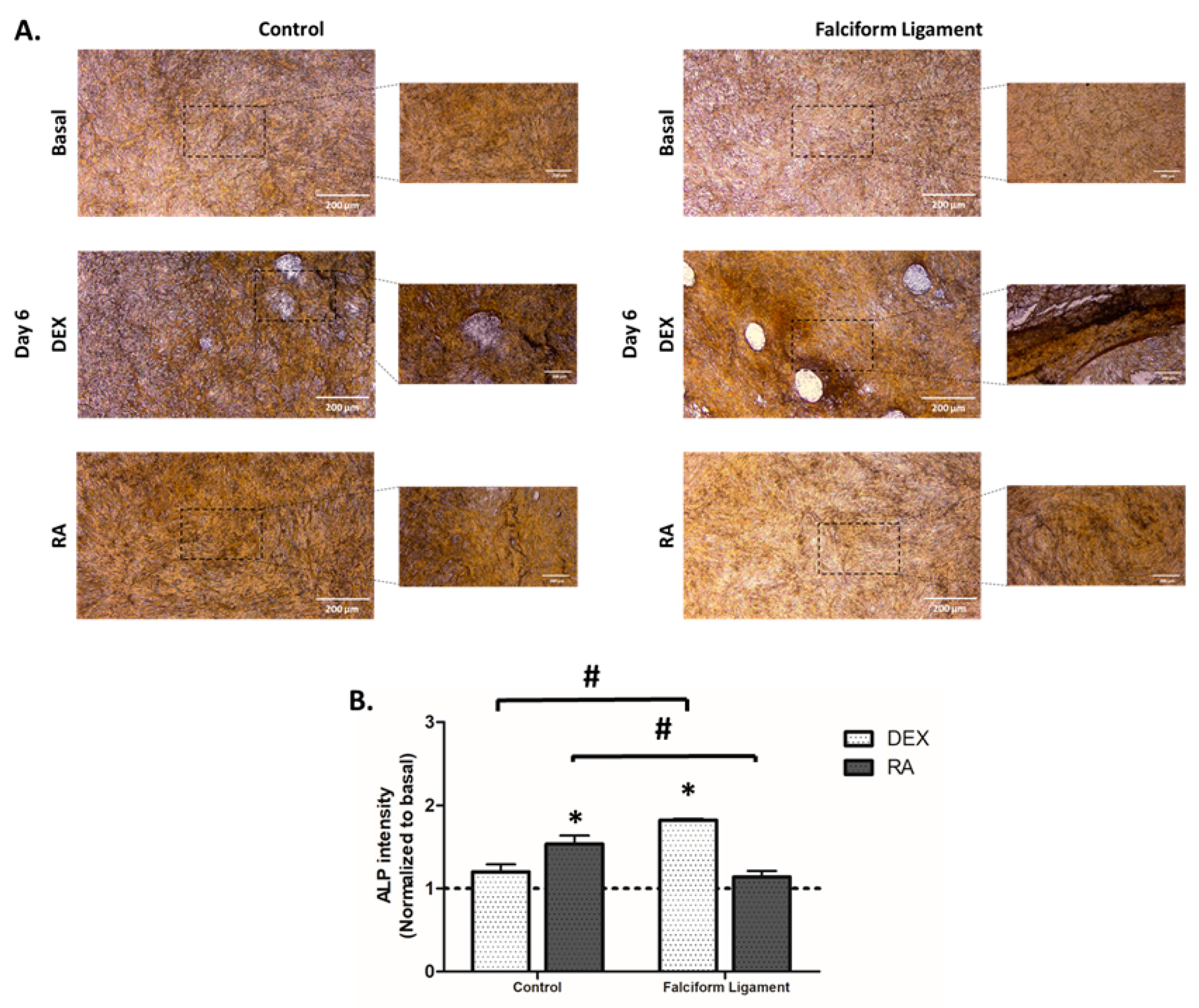
3.7. Cytochemical Staining of Alkaline Phosphatase
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, F.; Wang, Q.; Ouyang, L.; Wu, H.; Yang, Z.; Fu, X.; Liu, X.; Yan, L.; Cao, Y.; Xiao, R. Comparison of concentrated fresh mononuclear cells and cultured mesenchymal stem cells from bone marrow for bone regeneration. Stem Cells Transl. Med. 2021, 10, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Requicha, J.F.; Viegas, C.A.; Albuquerque, C.M.; Azevedo, J.M.; Reis, R.L.; Gomes, M.E. Effect of Anatomical Origin and Cell Passage Number on the Stemness and Osteogenic Differentiation Potential of Canine Adipose-Derived Stem Cells. Stem Cell Rev. Rep. 2012, 8, 1211–1222. [Google Scholar] [CrossRef]
- Voga, M.; Kovač, V.; Majdic, G. Comparison of Canine and Feline Adipose-Derived Mesenchymal Stem Cells/Medicinal Signaling Cells with Regard to Cell Surface Marker Expression, Viability, Proliferation, and Differentiation Potential. Front. Vet. Sci. 2021, 7, 610240. [Google Scholar] [CrossRef] [PubMed]
- Baer, P.C. Adipose-derived mesenchymal stromal/stem cells: An update on their phenotype in vivo and in vitro. World J. Stem Cells 2014, 6, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Szydlarska, J.; Weiss, C.; Marycz, K. The Effect of Methyl-β-cyclodextrin on Apoptosis, Proliferative Activity, and Oxidative Stress in Adipose-Derived Mesenchymal Stromal Cells of Horses Suffering from Metabolic Syndrome (EMS). Molecules 2018, 23, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safarova, Y.; Umbayev, B.; Hortelano, G.; Askarova, S. Mesenchymal stem cells modifications for enhanced bone targeting and bone regeneration. Regen. Med. 2020, 15, 1579–1594. [Google Scholar] [CrossRef] [Green Version]
- Connolly, J.F. Clinical use of marrow osteoprogenitor cells to stimulate osteogenesis. Clin. Orthop. Relat. Res. 1998, 355, S257–S266. [Google Scholar] [CrossRef]
- Hernigou, P.; Beaujean, F. Treatment of osteonecrosis with autologous bone marrow grafting. Clin. Orthop. Relat. Res. 2002, 405, 14–23. [Google Scholar] [CrossRef]
- Morishita, T.; Honoki, K.; Ohgushi, H.; Kotobuki, N.; Matsushima, A.; Takakura, Y. Tissue engineering approach to the treatment of bone tumors: Three cases of cultured bone grafts derived from patients’ mesenchymal stem cells. Artif. Organs 2006, 30, 115–118. [Google Scholar] [CrossRef]
- Quarto, R.; Mastrogiacomo, M.; Cancedda, R.; Kutepov, S.M.; Mukhachev, V.; Lavroukov, A.; Kon, E.; Marcacci, M. Repair of large bone defects with the use of autologous bone marrow stromal cells. N. Engl. J. Med. 2001, 344, 385–386. [Google Scholar] [CrossRef]
- Qin, Y.; Guan, J.; Zhang, C. Mesenchymal stem cells: Mechanisms and role in bone regeneration. Postgrad. Med. J. 2014, 90, 643–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Zhou, H.; Yan, C.; Wang, Y.; Xiao, C.; Gu, P.; Fan, X. In vitro osteogenic induction of bone marrow stromal cells with encapsulated gene-modified bone marrow stromal cells and in vivo implantation for orbital bone repair. Tissue Eng. 2014, 20, 2019–2029. [Google Scholar] [CrossRef]
- Zhang, L.; Tang, J.; Sun, L.; Zheng, T.; Pu, X.; Chen, Y.; Yang, K. Three-dimensional printed tissue engineered bone for canine mandibular defects. Genes Dis. 2020, 7, 138–149. [Google Scholar] [CrossRef]
- Bearden, R.N.; Huggins, S.S.; Cummings, K.J.; Smith, R.; Gregory, C.A.; Saunders, W.B. In-vitro characterization of canine multipotent stromal cells isolated from synovium, bone marrow, and adipose tissue: A donor-matched comparative study. Stem Cell Res. Ther. 2017, 8, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, B.-J.; Ryu, H.-H.; Park, S.S.; Koyama, Y.; Kikuchi, M.; Woo, H.-M.; Kim, W.H.; Kweon, O.-K. Comparing the osteogenic potential of canine mesenchymal stem cells derived from adipose tissues, bone marrow, umbilical cord blood, and Wharton’s jelly for treating bone defects. J. Vet. Sci. 2012, 13, 299–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaneselli, K.M.; Kuhl, C.P.; Terraciano, P.B.; de Oliveira, F.S.; Pizzato, S.B.; Pazza, K.; Magrisso, A.B.; Torman, V.; Rial, A.; Moreno, M.; et al. Comparison of the characteristics of canine adipose tissue-derived mesenchymal stem cells extracted from different sites and at different passage numbers. J. Vet. Sci. 2018, 19, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.-Y.; Wang, B.-Y.; Li, S.-C.; Luo, D.-Z.; Zhan, X.; Chen, S.-F.; Chen, Z.-S.; Liu, C.-Y.; Ji, H.-Q.; Bai, Y.-S.; et al. Evaluation of the Curative Effect of Umbilical Cord Mesenchymal Stem Cell Therapy for Knee Arthritis in Dogs Using Imaging Technology. Stem Cells Int. 2018, 2018, 1983025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisiel, A.H.; McDuffee, L.A.; Masaoud, E.; Bailey, T.R.; Gonzalez, B.P.E.; Nino-Fong, R. Isolation, characterization, and in vitro proliferation of canine mesenchymal stem cells derived from bone marrow, adipose tissue, muscle, and periosteum. Am. J. Vet. Res. 2012, 73, 1305–1317. [Google Scholar] [CrossRef]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef]
- Bhatt, A.; Robinson, E.; Cunningham, S.C. Spontaneous inflammation and necrosis of the falciform and round ligaments: A case report and review of the literature. J. Med. Case Rep. 2020, 14, 17. [Google Scholar] [CrossRef]
- Lee, S.W.; Chong, J.U.; Min, S.O.; Bak, S.Y.; Kim, K.S. Are Adipose-Derived Stem Cells from Liver Falciform Ligaments Another Possible Source of Mesenchymal Stem Cells? Cell Transplant. 2017, 26, 855–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.P.; Xu, D.C.; Tan, H.Y.; Li, C.L. Anatomical study on the morphology and blood supply of the falciform ligament and its clinical significance. Surg. Radiol. Anat. SRA 2004, 26, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Ozmen, M.M.; Coskun, F.; Ziraman, I. Falciform ligament in the management of the residual cavity for liver hydatidosis: New surgical technique. World J. Surg. 2006, 30, 1722–1728. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.O.; Gordon-Evans, W.J.; Fredericks, L.P.; Kiefer, K.; Conzemius, M.G.; Griffon, D.J. Comparison of Mesenchymal Stem Cell Surface Markers from Bone Marrow Aspirates and Adipose Stromal Vascular Fraction Sites. Front. Vet. Sci. 2015, 2, 82. [Google Scholar] [CrossRef] [Green Version]
- Hendawy, H.; Uemura, A.; Ma, D.; Namiki, R.; Samir, H.; Ahmed, M.F.; Elfadadny, A.; El-Husseiny, H.M.; Chieh-Jen, C.; Tanaka, R. Tissue Harvesting Site Effect on the Canine Adipose Stromal Vascular Fraction Quantity and Quality. Animals 2021, 11, 460. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Zhao, L.; Li, L. Current understanding of adipose-derived mesenchymal stem cell-based therapies in liver diseases. Stem Cell Res. Ther. 2019, 10, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satilmis, B.; Cicek, G.S.; Cicek, E.; Akbulut, S.; Sahin, T.T.; Yilmaz, S. Adipose-derived stem cells in the treatment of hepatobiliary diseases and sepsis. World J. Clin. Cases 2022, 10, 4348–4356. [Google Scholar] [CrossRef]
- Gao, J.; Dennis, J.E.; Solchaga, L.A.; Awadallah, A.S.; Goldberg, V.M.; Caplan, A.I. Tissue-Engineered Fabrication of an Osteochondral Composite Graft Using Rat Bone Marrow-Derived Mesenchymal Stem Cells. Tissue Eng. 2001, 7, 363–371. [Google Scholar] [CrossRef]
- Vieira, N.M.; Brandalise, V.; Zucconi, E.; Secco, M.; Strauss, B.E.; Zatz, M. Isolation, Characterization, and Differentiation Potential of Canine Adipose-Derived Stem Cells. Cell Transplant. 2010, 19, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Baptista, C.; Queirós, A.; Ferreira, R.; Fernandes, M.H.; Gomes, P.S.; Colaço, B. Retinoic acid induces the osteogenic differentiation of cat adipose tissue-derived stromal cells from distinct anatomical sites. J. Anat. 2022, 00, 1–12. [Google Scholar] [CrossRef]
- Song, H.M.; Nacamuli, R.P.; Xia, W.; Bari, A.S.; Shi, Y.-Y.; Fang, T.D.; Longaker, M.T. High-Dose Retinoic Acid Modulates Rat Calvarial Osteoblast Biology. J. Cell. Physiol. 2005, 202, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.C.; Shi, Y.-Y.; Nacamuli, R.P.; Quarto, N.; Lyons, K.M.; Longaker, M.T. Osteogenic differentiation of mouse adipose-derived adult stromal cells requires retinoic acid and bone morphogenetic protein receptor type IB signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 12335–12340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, B.; Nelson, E.R.; Brown, K.; James, A.W.; Xu, D.; Dunlevie, R.; Wu, J.C.; Lee, M.; Wu, B.; Commons, G.W.; et al. Differences in Osteogenic Differentiation of Adipose-Derived Stromal Cells from Murine, Canine, and Human Sources In Vitro and In Vivo. Plast. Reconstr. Surg. 2011, 128, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Freeman, L.; Becvarova, I.; Cave, N.; MacKay, C.; Nguyen, P.; Rama, B.; Takashima, G.; Tiffin, R.; Tsjimoto, H.; Beukelen, P.V. WSAVA nutritional assessment guidelines. J. Small Anim. Pract. 2011, 52, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, C.M.; Lima, P.S.P.; Zanette, D.L.; Oliveira, T.Y.K.; Pereira, F.d.A.; Lorenzi, J.C.C.; Santos, J.L.D.; Prata, K.L.; Neto, J.M.P.; de Paula, F.J.A.; et al. Osteoblastic differentiation of bone marrow mesenchymal stromal cells in Bruck Syndrome. BMC Med. Genet. 2016, 17, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, S.; Ghosh, J.; Dhara, S.K. Osteogenic differentiation potential of porcine bone marrow mesenchymal stem cell subpopulations selected in different basal media. Biol. Open 2020, 9, bio053280. [Google Scholar] [CrossRef] [PubMed]
- Capak, H.; Bottegaro, N.B.; Manojlovic, A.; Smolec, O.; Vnuk, D. Review of 166 Gunshot Injury Cases in Dogs. Top. Companion Anim. Med. 2016, 31, 146–151. [Google Scholar] [CrossRef]
- Kumar, K.; Mogha, I.V.; Aithal, H.P.; Kinjavdekar, P.; Singh, G.R.; Pawde, A.M.; Kushwaha, R.B. Occurrence and Pattern of Long Bone Fractures in Growing Dogs with Normal and Osteopenic Bones. J. Vet. Med. 2007, 54, 484–490. [Google Scholar] [CrossRef]
- Minar, M.; Hwang, Y.; Park, M.; Kim, G. Retrospective study on fractures in dogs. J. Biomed. Res. 2013, 14, 140–144. [Google Scholar] [CrossRef]
- Ali, L.M.B. Incidence, Occurrence, Classification and Outcome of Small Animal Fractures: A Retrospective Study (2005–2010). Anim. Vet. Sci. 2013, 7, 191–196. [Google Scholar]
- Jain, R.; Shukla, B.P.; Nema, S.; Shukla, S.; Chabra, D.; Karmore, S.K. Incidence of fracture in dog: A retrospective study. Vet. Pract. 2018, 19, 63–65. [Google Scholar]
- Libardoni, R.D.N.; Costa, D.D.; Menezes, F.B.; Cavalli, L.G.; Pedrotti, L.F.; Kohlrausch, P.R.; Minto, B.W.; Silva, M.A.M. Classification, fixation techniques, complications and outcomes of femur fractures in dogs and cats: 61 cases (2015–2016). Clin. Surg. 2018, 48, e20170028. [Google Scholar] [CrossRef]
- Uwagie-Ero, E.A.; Abiaezute, C.N.; Okorie-Kanu, O.J.; Odigie, E.A.; Asemota, O.D. Retrospective evaluation of canine fractures in southern Nigeria. Comp. Clin. Pathol. 2018, 27, 1127–1132. [Google Scholar] [CrossRef]
- Boston, S.E.; Vinayak, A.; Lu, X.; Larue, S.; Bacon, N.J.; Bleedorn, J.A.; Souza, C.H.M.; Ehrhart, N.P. Outcome and complications in dogs with appendicular primary bone tumors treated with stereotactic radiotherapy and concurrent surgical stabilization. Vet. Surg. 2017, 46, 829–837. [Google Scholar] [CrossRef]
- Frese, L.; Dijkman, P.E.; Hoerstrup, S.P. Adipose Tissue-Derived Stem Cells in Regenerative Medicine. Transfus. Med. Hemotherapy 2016, 43, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Alves, E.G.L.; Serakides, R.; Boeloni, J.N.; Rosado, I.R.; Ocarino, N.M.; Oliveira, H.P.; Góes, A.M.; Rezende, C.M.F. Comparison of the osteogenic potential of mesenchymal stem cells from the bone marrow and adipose tissue of young dogs. BMC Vet. Res. 2014, 10, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinello, T.; Bronzini, I.; Maccatrozzo, L.; Mollo, A.; Sampaolesi, M.; Mascarello, F.; Decaminada, M.; Patruno, M. Canine adipose-derived-mesenchymal stem cells do not lose stem features after a long-term cryopreservation. Res. Vet. Sci. 2011, 91, 18–24. [Google Scholar] [CrossRef]
- Schwarz, C.; Leicht, U.; Rothe, C.; Drosse, I.; Luibl, V.; Röcken, M.; Schieker, M. Effects of different media on proliferation and differentiation capacity of canine, equine and porcine adipose derived stem cells. Res. Vet. Sci. 2012, 93, 457–462. [Google Scholar] [CrossRef]
- Weatherall, E.L.; Avilkina, V.; Cortes-Araya, Y.; Dan-Jumbo, S.; Stenhouse, C.; Donadeu, F.X.; Esteves, C.L. Differentiation Potential of Mesenchymal Stem/Stromal Cells Is Altered by Intrauterine Growth Restriction. Front. Vet. Sci. 2020, 7, 558905. [Google Scholar] [CrossRef] [PubMed]
- Chun, K.-H. Mouse model of the adipose organ: The heterogeneous anatomical characteristics. Arch. Pharmacal Res. 2021, 44, 857–875. [Google Scholar] [CrossRef]
- Hung, C.-S.; Lee, J.-K.; Yang, C.-Y.; Hsieh, H.-R.; Ma, W.-Y.; Lin, M.-S.; Liu, P.-H.; Shih, S.-R.; Liou, J.-M.; Chuang, L.-M.; et al. Measurement of Visceral Fat: Should We Include Retroperitoneal Fat? PLoS ONE 2014, 9, e112355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: Structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Sackmann-Sala, L.; Berryman, D.E.; Munn, R.D.; Lubbers, E.R.; Kopchick, J.J. Heterogeneity among white adipose tissue depots in male C57BL/6J mice. Obesity 2012, 20, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Cárcamo-Orive, I.; Gaztelumendi, A.; Delgado, J.; Tejados, N.; Dorronsoro, A.; Fernández-Rueda, J.; Pennington, D.J.; Trigueros, C. Regulation of Human Bone Marrow Stromal Cell Proliferation and Differentiation Capacity by Glucocorticoid Receptor and AP-1 Crosstalk. J. Bone Miner. Metab. 2010, 25, 2115–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neganova, I.; Lako, M. G1 to S phase cell cycle transition in somatic and embryonic stem cells. J. Anat. 2008, 213, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Fan, Y.; Loor, J.J.; Liang, Y.; Sun, X.; Jia, H.; Zhao, C.; Xu, C. All-trans retinoic acid controls differentiation, proliferation, and lipolysis in isolated subcutaneous adipocytes from peripartal Holstein cows. J. Dairy Sci. 2021, 104, 4999–5008. [Google Scholar] [CrossRef]
- Guercio, A.; Bella, S.D.; Casella, S.; Marco, P.D.; Russo, C.; Piccione, G. Canine mesenchymal stem cells (MSCs): Characterization in relation to donor age and adipose tissue-harvesting site. Cell Biol. Int. 2013, 37, 789–798. [Google Scholar] [CrossRef]
- Neupane, M.; Chang, C.-C.; Kiupel, M.; Yuzbasiyan-Gurkan, V. Isolation and Characterization of Canine Adipose–Derived Mesenchymal Stem Cells. Tissue Eng. Part A 2008, 14, 1007–1015. [Google Scholar] [CrossRef]
- Langenfeld, J.; Kiyokawa, H.; Sekula, D.; Boyle, J.; Dmitrovsky, E. Posttranslational regulation of cyclin D1 by retinoic acid: A chemoprevention mechanism. Proc. Natl. Acad. Sci. USA 1997, 94, 12070–12074. [Google Scholar] [CrossRef] [Green Version]
- Bahamondes, F.; Flores, E.; Cattaneo, G.; Bruna, F.; Conget, P. Omental adipose tissue is a more suitable source of canine Mesenchymal stem cells. BMC Vet. Res. 2017, 13, 166. [Google Scholar] [CrossRef] [Green Version]
- Arrigoni, E.; Lopa, S.; de Girolamo, L.; Stanco, D.; Brini, A.T. Isolation, characterization and osteogenic differentiation of adipose-derived stem cells: From small to large animal models. Cell Tissue Res. 2009, 338, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.D.M.; Alves, A.L.G.; Golim, M.A.; Moroz, A.; Hussni, C.A.; Oliveira, P.G.G.D.; Deffune, E. Isolation and immunophenotypic characterization of mesenchymal stem cells derived from equine species adipose tissue. Vet. Immunol. Immunopathol. 2009, 132, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Wang, F.; Xiang, B.; Deng, J.; Liu, S.; Lin, H.-Y.; Natarajan, K.; Li, G.; Wang, J.; Lin, F.; et al. Adipose-Derived Stem Cells From Both Visceral and Subcutaneous Fat Deposits Significantly Improve Contractile Function of Infarcted Rat Hearts. Cell Transplant. 2015, 24, 2337–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna, A.C.L.; Madeira, M.E.P.; Conceição, T.O.; Moreira, J.A.L.C.; Laiso, R.A.N.; Maria, D.A. Characterization of adipose-derived stem cells of anatomical region from mice. BMC Res. Notes 2014, 7, 552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panasophonkul, S.; Samart, P.; Kongon, K.; Sathanawongs, A. Phenotypic characteristics of feline adipose-derived stem cells affected by cell passage number. Pol. J. Vet. Sci. 2017, 20, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.F.; El-Sayed, A.K.; Chen, H.; Zhao, R.; Yusuf, M.S.; Zuo, Q.; Zhang, Y.; Li, B. Comparison between curcumin and all-trans retinoic acid in the osteogenic differentiation of mouse bone marrow mesenchymal stem cells. Exp. Ther. Med. 2019, 17, 4154–4166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.; Napoli, J.L. All-trans-retinoic acid stimulates translation and induces spine formation in hippocampal neurons through a membrane-associated RARalpha. FASEB J. 2008, 22, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Kirkham, G.R.; Cartmell, S.H. Genes and Proteins Involved in the Regulation of Osteogenesis. Top. Tissue Eng. 2007, 3, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Runx2, A Multifunctional Transcription Factor in Skeletal Development. J. Cell. Biochem. 2002, 87, 1–8. [Google Scholar] [CrossRef]
- Huang, W.; Yang, S.; Shao, J.; Li, Y.P. Signaling and transcriptional regulation in osteoblast commitment and differentiation. Front. Biosci. A J. Virtual Libr. 2007, 12, 3068–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C. Transcriptional regulation of bone formation by the osteoblast-specific transcription factor Osx. J. Orthop. Surg. Res. 2010, 5, 37. [Google Scholar] [CrossRef] [PubMed]
- Langenbach, F.; Handschel, J. Effects of dexamethasone, ascorbic acid and β-glycerophosphate on the osteogenic differentiation of stem cells in vitro. Stem Cell Res. Ther. 2013, 4, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loebel, C.; Czekanska, E.M.; Bruderer, M.; Salzmann, G.; Alini, M.; Stoddart, M.J. In vitro osteogenic potential of human mesenchymal stem cells is predicted by Runx2/Sox9 ratio. Tissue Eng. Part A 2015, 21, 115–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Bella, E.; Buetti-Dinh, A.; Licandro, G.; Ahmad, P.; Basoli, V.; Alini, M.; Stoddart, M.J. Dexamethasone Induces Changes in Osteogenic Differentiation of Human Mesenchymal Stromal Cells via SOX9 and PPARG, but Not RUNX2. Int. J. Mol. Sci. 2021, 22, 4785. [Google Scholar] [CrossRef]
- Stöckl, S.; Göttl, C.; Grifka, J.; Grässel, S. Sox9 Modulates Proliferation and Expression of Osteogenic Markers of Adipose-Derived Stem Cells (ASC). Cell. Physiol. Biochem. 2013, 31, 703–717. [Google Scholar] [CrossRef]
- Arnhold, S.; Elashry, M.I.; Klymiuk, M.C.; Geburek, F. Investigation of stemness and multipotency of equine adipose-derived mesenchymal stem cells (ASCs) from different fat sources in comparison with lipoma. Stem Cell Res. Ther. 2019, 10, 309. [Google Scholar] [CrossRef]
- Rashid, U.; Yousaf, A.; Yaqoob, M.; Saba, E.; Moaeen-ud-Din, M.; Waseem, S.; Becker, S.K.; Sponder, G.; Aschenbach, J.R.; Sandhu, M.A. Characterization and differentiation potential of mesenchymal stem cells isolated from multiple canine adipose tissue sources. BMC Vet. Res. 2021, 17, 388. [Google Scholar] [CrossRef]
- Chen, M.; Huang, H.-Z.; Wang, M.; Wang, A.-X. Retinoic acid inhibits osteogenic differentiation of mouse embryonic palate mesenchymal cells. Birth Defects Res. Part A Clin. Mol. Teratol. 2010, 88, 965–970. [Google Scholar] [CrossRef]
- Zhang, W.; Deng, Z.-L.; Chen, L.; Zuo, G.-W.; Luo, Q.; Shi, Q.; Zhang, B.-Q.; Wagner, E.R.; Rastegar, F.; Kim, S.H.; et al. Retinoic Acids Potentiate BMP9-Induced Osteogenic Differentiation of Mesenchymal Progenitor Cells. PLoS ONE 2010, 5, e11917. [Google Scholar] [CrossRef] [Green Version]
- Asserson, D.B.; Orbay, H.; Sahar, D.E. Review of the Pathways Involved in the Osteogenic Differentiation of Adipose-Derived Stem Cells. J. Craniofacial Surg. 2019, 30, 703–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) signaling in development and human diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Gui, C.; Parson, J.; Meyer, G.A. Harnessing adipose stem cell diversity in regenerative medicine. APL Bioeng. 2021, 5, 021501. [Google Scholar] [CrossRef] [PubMed]
- Pförringer, D.; Aitzetmüller, M.M.; Brett, E.A.; Houschyar, K.S.; Schäfer, R.; van Griensven, M.; Duscher, D. Single-Cell Gene Expression Analysis and Evaluation of the Therapeutic Function of Murine Adipose-Derived Stromal Cells (ASCs) from the Subcutaneous and Visceral Compartment. Stem Cells Int. 2018, 2018, 2183736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, A.; Friemel, A.; Roth, S.; Kreis, N.N.; Hoock, S.C.; Safdar, B.K.; Fischer, K.; Möllmann, C.; Solbach, C.; Louwen, F.; et al. Subcutaneous and Visceral Adipose-Derived Mesenchymal Stem Cells: Commonality and Diversity. Cells 2019, 8, 1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Zhang, T.; Hu, S.; Tang, Q.; Long, X.; Yang, X.; Gun, S.; Chen, L. Chromatin accessibility landscape of stromal subpopulations reveals distinct metabolic and inflammatory features of porcine subcutaneous and visceral adipose tissue. PeerJ Biochem. Biophys. Mol. Biol. 2022, 10, e13250. [Google Scholar] [CrossRef]
- Vohl, M.-C.; Sladek, R.; Robitaille, J.; Gurd, S.; Marceau, P.; Richard, D.; Hudson, T.J.; Tchernof, A. A survey of genes differentially expressed in subcutaneous and visceral adipose tissue in men. Obes. Res. 2004, 12, 1217–1222. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Assay ID |
|---|---|
| Beta-actin (ACTB) | qCfaCED0037901 |
| SRY-Box transcription factor 9 (SOX9) | qCfaCED0025675 |
| RUNX family transcription factor 2 (RUNX2) | qCfaCED0033695 |
| Collagen type I alpha 1 chain (COL1A1) | qCfaCED0027854 |
| Osterix (SP7) | qCfaCED0032017 |
| Osteocalcin (BGLAP) | qCfaCED0031563 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira-Baptista, C.; Queirós, A.; Ferreira, R.; Fernandes, M.H.; Colaço, B.; Gomes, P.S. The Osteogenic Potential of Falciform Ligament-Derived Stromal Cells—A Comparative Analysis between Two Osteogenic Induction Programs. Bioengineering 2022, 9, 810. https://doi.org/10.3390/bioengineering9120810
Ferreira-Baptista C, Queirós A, Ferreira R, Fernandes MH, Colaço B, Gomes PS. The Osteogenic Potential of Falciform Ligament-Derived Stromal Cells—A Comparative Analysis between Two Osteogenic Induction Programs. Bioengineering. 2022; 9(12):810. https://doi.org/10.3390/bioengineering9120810
Chicago/Turabian StyleFerreira-Baptista, Carla, André Queirós, Rita Ferreira, Maria Helena Fernandes, Bruno Colaço, and Pedro Sousa Gomes. 2022. "The Osteogenic Potential of Falciform Ligament-Derived Stromal Cells—A Comparative Analysis between Two Osteogenic Induction Programs" Bioengineering 9, no. 12: 810. https://doi.org/10.3390/bioengineering9120810