
Effect of Hypoxia on Branching Characteristics and Cell Subpopulations during Kidney Organ Culture
 ,
, 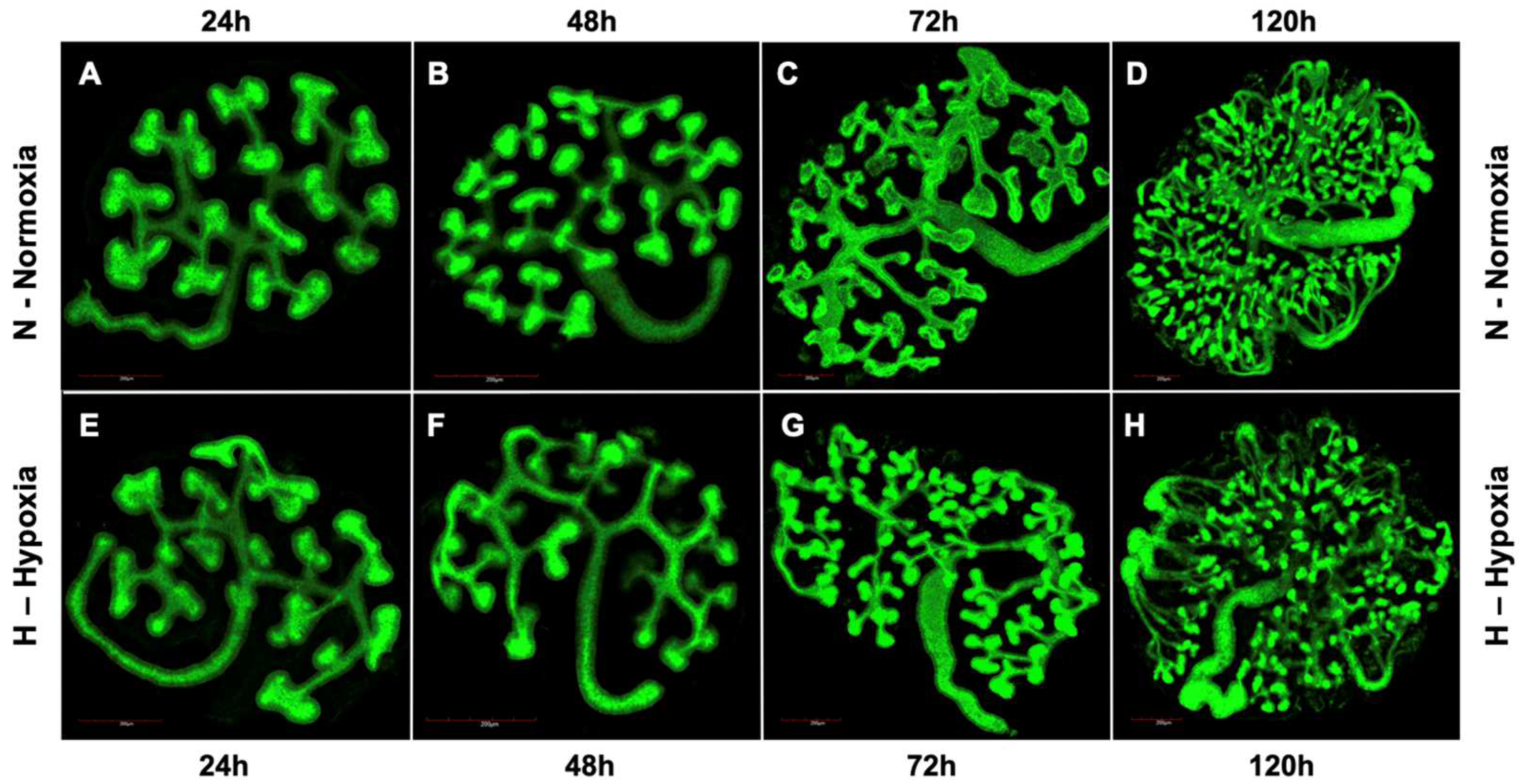{kind=link}
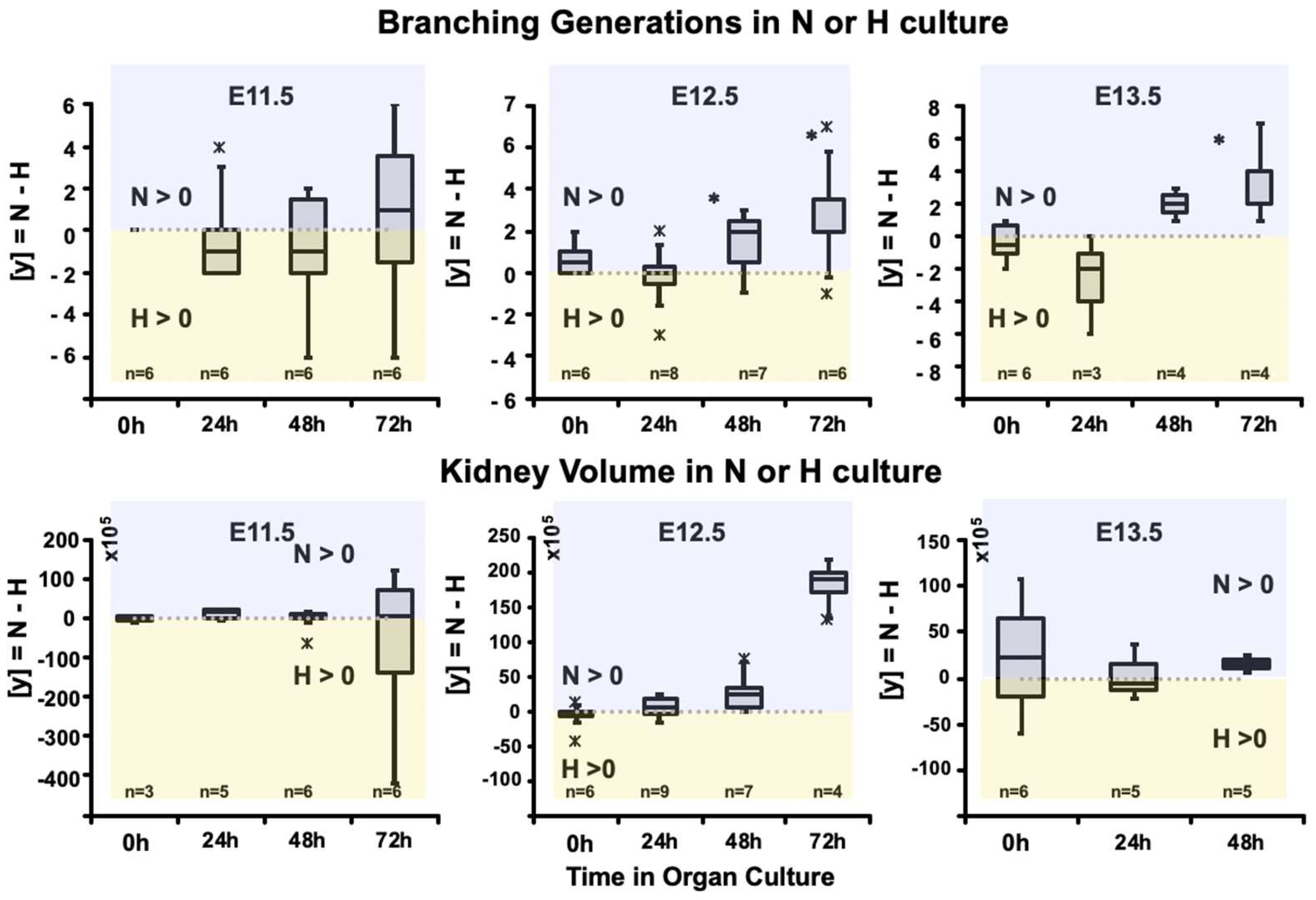{kind=link}
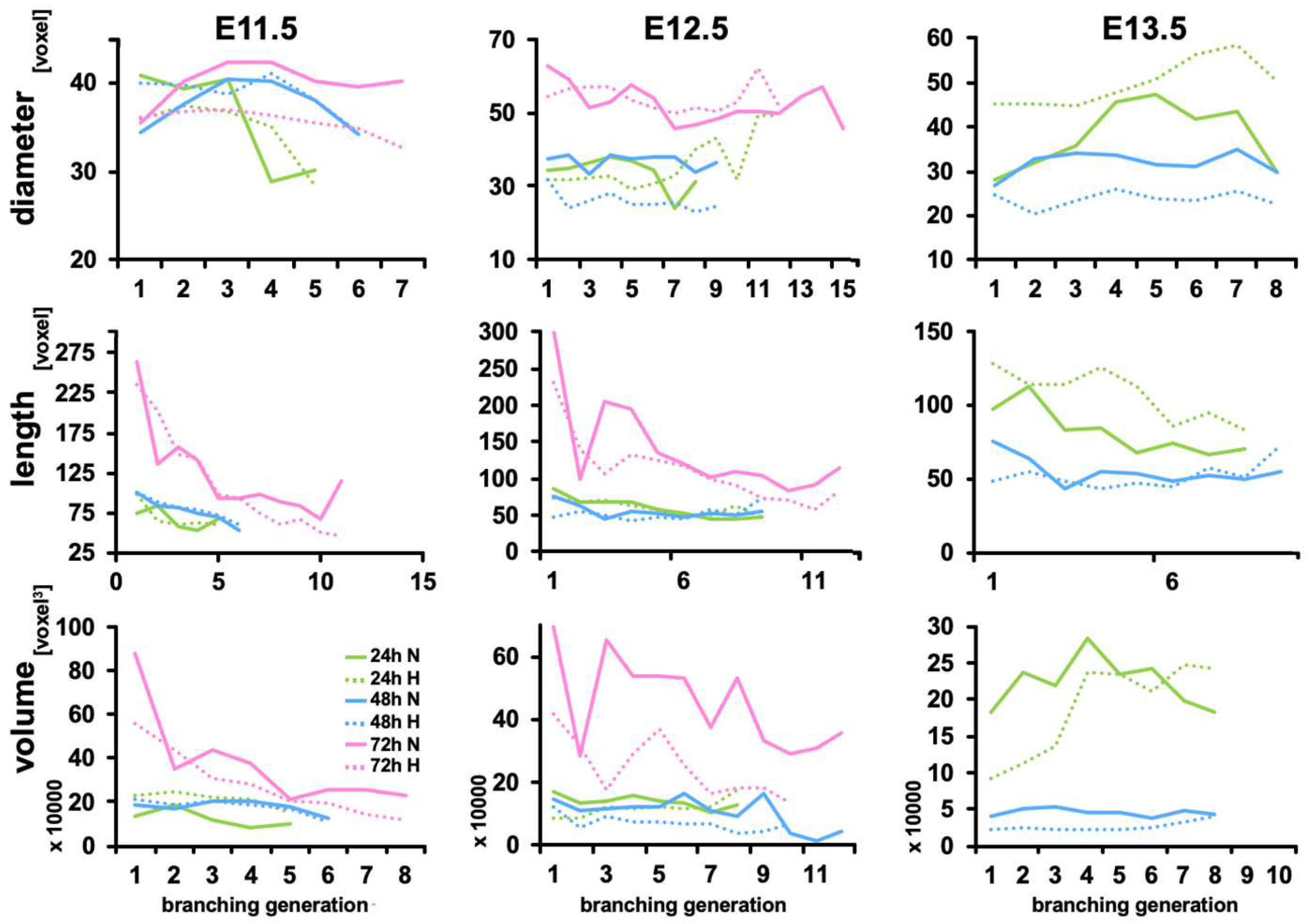{kind=link}
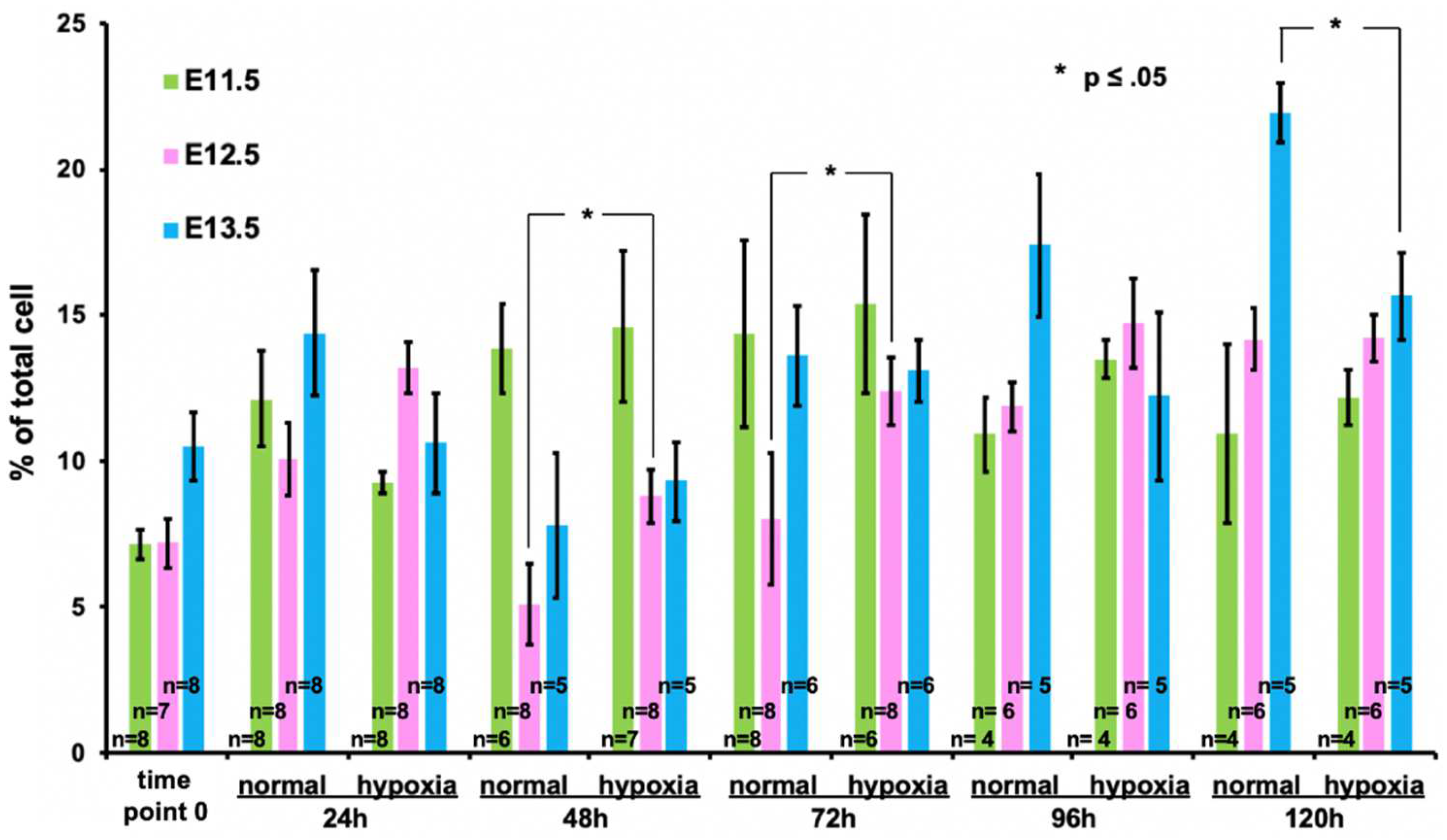{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Isolation of Embryonic Kidneys
2.3. Confocal Microscopy and Analysis of Branching Characteristics
2.4. Statistical Analysis
2.5. RNA Extraction and Gene Expression Analysis
2.6. Hoxb7 Assay
3. Results
3.1. UB Branching Morphogenesis
3.2. Calculated Branching Morphogenesis
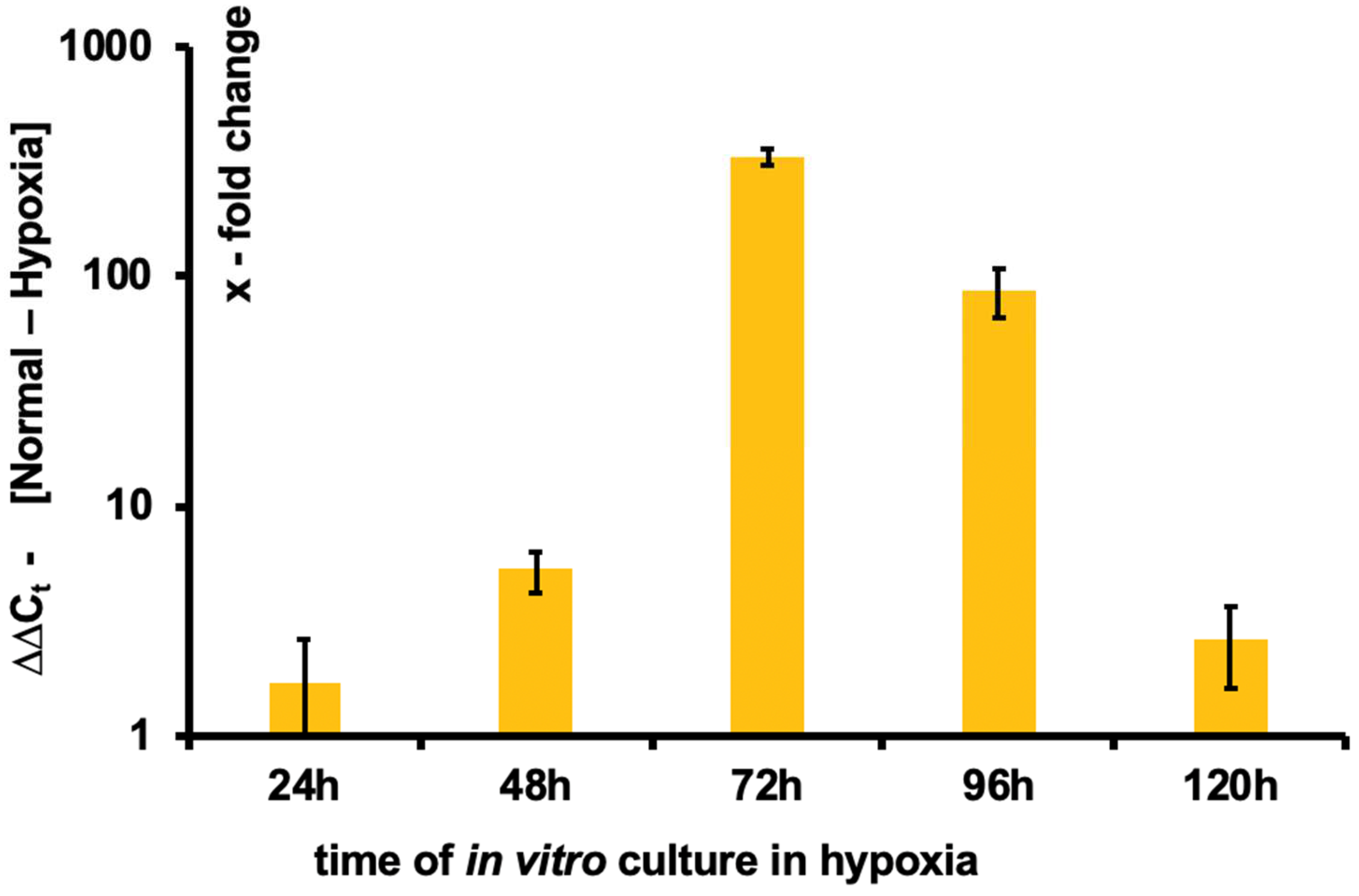
3.3. Expansion of hoxb7+ Cells in Hypoxia
3.4. Induction of HIF1α
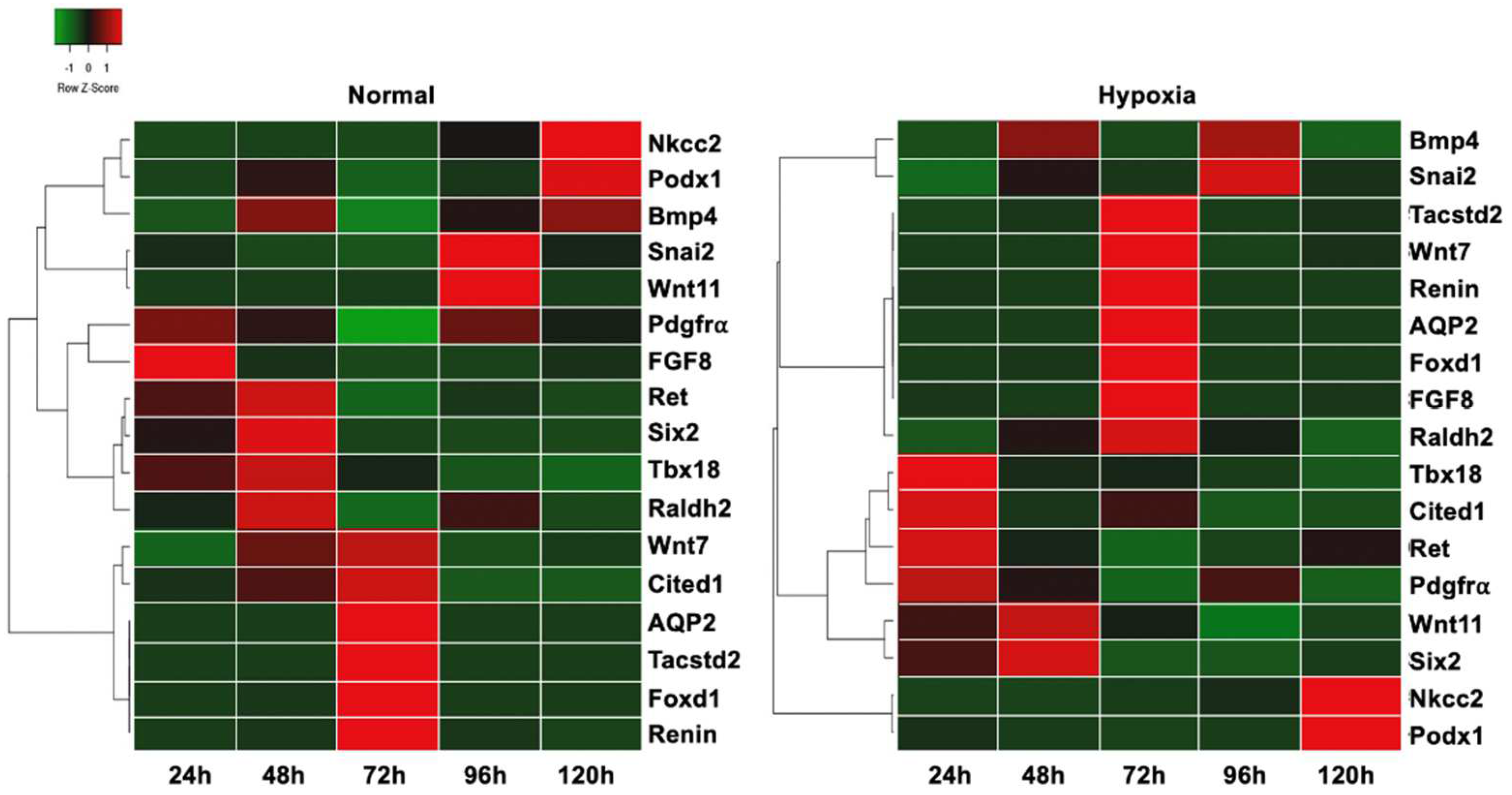
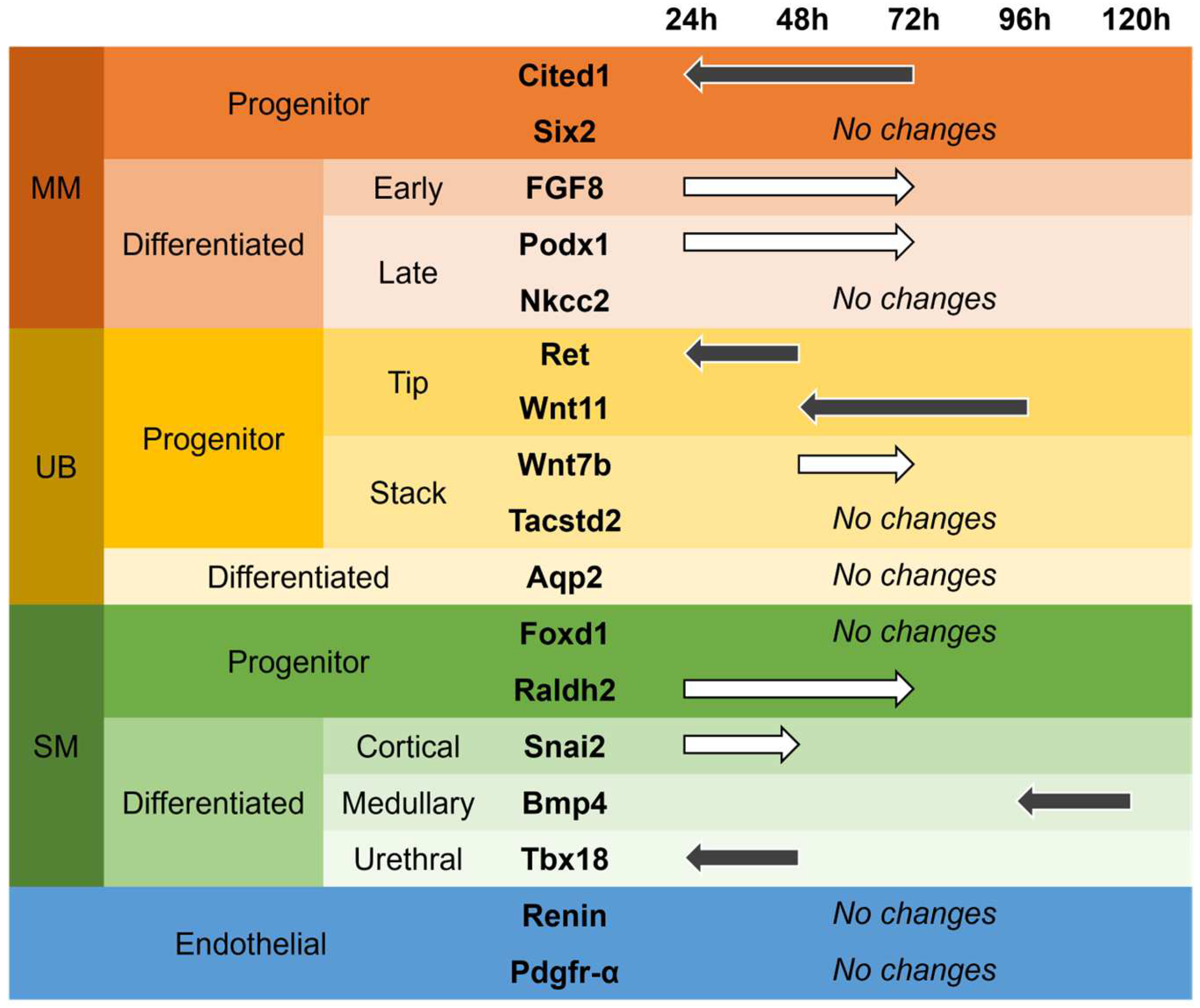
3.5. Differentiation Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fathollahipour, S.; Patil, P.S.; Leipzig, N.D. Oxygen Regulation in Development: Lessons from Embryogenesis towards Tissue Engineering. Cells Tissues Organs 2018, 205, 350–371. [Google Scholar] [CrossRef] [PubMed]
- Michiels, C. Physiological and pathological responses to hypoxia. Am. J. Pathol. 2004, 164, 1875–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdollahi, H.; Harris, L.J.; Zhang, P.; McIlhenny, S.; Srinivas, V.; Tulenko, T.; DiMuzio, P.J. The role of hypoxia in stem cell differentiation and therapeutics. J. Surg. Res. 2011, 165, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forristal, C.E.; Wright, K.L.; Hanley, N.A.; Oreffo, R.O.; Houghton, F.D. Hypoxia inducible factors regulate pluripotency and proliferation in human embryonic stem cells cultured at reduced oxygen tensions. Reproduction 2010, 139, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dressler, G.R. The cellular basis of kidney development. Annu. Rev. Cell Dev. Biol. 2006, 22, 509–529. [Google Scholar] [CrossRef]
- Basu, J.; Ludlow, J.W. Developmental engineering the kidney: Leveraging principles of morphogenesis for renal regeneration. Birth Defects Res. Part C Embryo Today Rev. 2012, 96, 30–38. [Google Scholar] [CrossRef]
- Hauser, P.V.; Nishikawa, M.; Kimura, H.; Fujii, T.; Yanagawa, N. Controlled tubulogenesis from dispersed ureteric bud-derived cells using a micropatterned gel. J. Tissue Eng. Regen. Med. 2016, 10, 762–771. [Google Scholar] [CrossRef] [Green Version]
- Lenas, P.; Moos, M.; Luyten, F.P. Developmental engineering: A new paradigm for the design and manufacturing of cell-based products. Part I: From three-dimensional cell growth to biomimetics of in vivo development. Tissue Eng. Part B Rev. 2009, 15, 381–394. [Google Scholar] [CrossRef] [Green Version]
- Grobstein, C. Inductive epitheliomesenchymal interaction in cultured organ rudiments of the mouse. Science 1953, 118, 52–55. [Google Scholar] [CrossRef]
- Grobstein, C. Trans-filter induction of tubules in mouse metanephrogenic mesenchyme. Exp. Cell Res. 1956, 10, 424–440. [Google Scholar] [CrossRef]
- Trowell, O.A. A modified technique for organ culture in vitro. Exp. Cell Res. 1954, 6, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Rak-Raszewska, A.; Hauser, P.V.; Vaino, S. Organ In Vitro Culture: What Have We Learned about Early Kidney Development? Stem Cells Int. 2015, 2015, 959807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganeva, V.; Unbekandt, M.; Davies, J.A. An improved kidney dissociation and reaggregation culture system results in nephrons arranged organotypically around a single collecting duct system. Organogenesis 2011, 7, 83–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebinger, D.D.R.; Unbekandt, M.; Davies, J.A. A novel, low-volume method for organ culture of embryonic kidneys that allows development of cortico-medullary anatomical organization. PLoS ONE 2010, 5, e10550. [Google Scholar] [CrossRef] [Green Version]
- Short, K.; Hodson, M.; Smyth, I. Spatial mapping and quantification of developmental branching morphogenesis. Dev. Camb. Engl. 2013, 140, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Jeyaseelan, K.; Ma, D.; Armugam, A. Real-time detection of gene promoter activity: Quantitation of toxin gene transcription. Nucleic Acids Res. 2001, 29, E58. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Gunaratnam, L.; Bonventre, J.V. HIF in Kidney Disease and Development. J. Am. Soc. Nephrol. 2009, 20, 1877. [Google Scholar] [CrossRef] [Green Version]
- Grieshammer, U.; Cebrian, C.; Cebrian, C.; Ilagan, R.; Meyers, E.; Herzlinger, D.; Martin, G.R. FGF8 is required for cell survival at distinct stages of nephrogenesis and for regulation of gene expression in nascent nephrons. Development 2005, 132, 3847–3857. [Google Scholar] [CrossRef] [Green Version]
- Boyle, S.; Shioda, T.; Perantoni, A.O.; de Caestecker, M. Cited1 and Cited2 are differentially expressed in the developing kidney but are not required for nephrogenesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2007, 236, 2321–2330. [Google Scholar] [CrossRef]
- Self, M.; Lagutin, O.V.; Bowling, B.; Hendrix, J.; Chai, Y.; Dressler, G.R.; Oliver, G. Six2 is required for suppression of nephrogenesis and progenitor renewal in the developing kidney. EMBO J. 2006, 25, 5214–5228. [Google Scholar] [CrossRef]
- Krause, M.; Rak-Raszewska, A.; Pietilä, I.; Quaggin, S.E.; Vainio, S. Signaling during Kidney Development. Cells 2015, 4, 112–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. Oxygen homeostasis. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 336–361. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.K.; Shah, M.M. How Does the Ureteric Bud Branch? J. Am. Soc. Nephrol. 2009, 20, 1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerychova, R.; Pavlinkova, G. HIF-1, Metabolism, and Diabetes in the Embryonic and Adult Heart. Front. Endocrinol. 2018, 9, 460. [Google Scholar] [CrossRef] [Green Version]
- Minet, E.; Michel, G.; Remacle, J.; Michiels, C. Role of HIF-1 as a transcription factor involved in embryonic development, cancer progression and apoptosis (review). Int. J. Mol. Med. 2000, 5, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Dunwoodie, S.L. The Role of Hypoxia in Development of the Mammalian Embryo. Dev. Cell. 2009, 17, 755–773. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, K.; Kitamura, S.; Makino, H. Hypoxia-inducible factor 1α regulates branching morphogenesis during kidney development. Biochem. Biophys. Res. Commun. 2014, 447, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Schley, G.; Scholz, H.; Kraus, A.; Hackenbeck, T.; Klanke, B.; Willam, C.; Wiesener, M.S.; Heinze, E.; Burzlaff, N.; Eckardt, K.U.; et al. Hypoxia inhibits nephrogenesis through paracrine Vegfa despite the ability to enhance tubulogenesis. Kidney Int. 2015, 88, 1283–1292. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, W.M.; Schmitt, R.; Rosenberger, C.; Münchenhagen, P.M.; Gröne, H.J.; Frei, U.; Warnecke, C.; Bachmann, S.; Wiesener, M.S.; Willam, C.; et al. Expression of hypoxia-inducible transcription factors in developing human and rat kidneys. Kidney Int. 2006, 69, 114–122. [Google Scholar] [CrossRef]
- Brown, A.C.; Muthukrishnan, S.D.; Guay, J.A.; Adams, D.C.; Schafer, D.A.; Fetting, J.L.; Oxburgh, L. Role for compartmentalization in nephron progenitor differentiation. Proc. Natl. Acad. Sci. USA 2013, 110, 4640–4645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtzeborn, K.; Cebrian, C.; Kuure, S. Regulation of Renal Differentiation by Trophic Factors. Front. Physiol. 2018, 9, 1588. [Google Scholar] [CrossRef] [PubMed]
- Haigl, B.; Mayer, C.-E.; Mayer, C.E.; Siegwart, G.; Sutterluety, H. Sprouty4 levels are increased under hypoxic conditions by enhanced mRNA stability and transcription. Biol. Chem. 2010, 391, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Chi, L.; Zhang, S.; Lin, Y.; Prunskaite-Hyyryläinen, R.; Vuolteenaho, R.; Itäranta, P.; Vainio, S. Sprouty proteins regulate ureteric branching by coordinating reciprocal epithelial Wnt11, mesenchymal Gdnf and stromal Fgf7 signaling during kidney development. Dev. Camb. Engl. 2004, 131, 3345–3356. [Google Scholar] [CrossRef] [Green Version]
- Mae, S.-I.; Ryosaka, M.; Sakamoto, S.; Matsuse, K.; Nozaki, A.; Igami, M.; Kabai, R.; Watanabe, A.; Osafune, K. Expansion of Human iPSC-Derived Ureteric Bud Organoids with Repeated Branching Potential. Cell Rep. 2020, 32, 107963. [Google Scholar] [CrossRef]
- Cheng, H.-T.; Kim, M.; Valerius, M.T.; Surendran, K.; Schuster-Gossler, K.; Gossler, A.; McMahon, A.P.; Kopan, R. Notch2, but not Notch1, is required for proximal fate acquisition in the mammalian nephron. Dev. Camb. Engl. 2007, 134, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, S.; Jiang, Q.; Kobayashi, C.; Nishinakamura, R. Notch2 activation in the embryonic kidney depletes nephron progenitors. J. Am. Soc. Nephrol. 2010, 21, 803–810. [Google Scholar] [CrossRef] [Green Version]
- Karner, C.M.; Das, A.; Ma, Z.; Self, M.; Chen, C.; Lum, L.; Oliver, G.; Carroll, T.J. Canonical Wnt9b signaling balances progenitor cell expansion and differentiation during kidney development. Dev. Camb. Engl. 2011, 138, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- Niederreither, K.; McCaffery, P.; Draeger, U.C.; Chambon, P.; Dolle, P. Restricted expression and retinoic acid-induced downregulation of the retinaldehyde dehydrogenase type 2 (RALDH-2) gene during mouse development. Mech. Dev. 1997, 62, 67–78. [Google Scholar] [CrossRef]
- Rosselot, C.; Spraggon, L.; Chia, I.; Batourina, E.; Riccio, P.; Lu, B.; Niederreither, K.; Dolle, P.; Duester, G.; Chambon, P.; et al. Non-cell-autonomous retinoid signaling is crucial for renal development. Development 2010, 137, 283–292. [Google Scholar] [CrossRef]
- Sauter, A.; Machura, K.; Neubauer, B.; Kurtz, A.; Wagner, C. Development of renin expression in the mouse kidney. Kidney Int. 2008, 73, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostendorf, T.; Boor, P.; van Roeyen, C.R.; Floege, J. Platelet-derived growth factors (PDGFs) in glomerular and tubulointerstitial fibrosis. Kidney Int. Suppl. 2014, 4, 65–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sequeira-Lopez, M.L.S.; Lin, E.E.; Li, M.; Hu, Y.; Sigmund, C.D.; Gomez, R.A. The earliest metanephric arteriolar progenitors and their role in kidney vascular development. Am. J. Physiol Regul Integr Comp. Physiol. 2015, 308, R138–R149. [Google Scholar] [CrossRef] [Green Version]
- Gomez, I.G.; Duffield, J.S. The FOXD1 lineage of kidney perivascular cells and myofibroblasts: Functions and responses to injury. Kidney Int. Suppl. 2014, 4, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera Millar, V.R.; Mangiavini, L.; Polito, U.; Canciani, B.; Nguyen, V.T.; Cirillo, F.; Anastasia, L.; Peretti, G.M.; Modina, S.C.; Di Giancamillo, A. Hypoxia as a Stimulus for the Maturation of Meniscal Cells: Highway to Novel Tissue Engineering Strategies? Int. J. Mol. Sci. 2021, 22, 6905. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.; Hanada, S.; Fuji, T.; Sakai, Y. Direct oxygen supply with polydimethylsiloxane (PDMS) membranes induces a spontaneous organization of thick heterogeneous liver tissues from rat fetal liver cells in vitro. Cell Transplant. 2012, 21, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Taheem, D.K.; Jell, G.; Gentleman, E. Hypoxia Inducible Factor-1α in Osteochondral Tissue Engineering. Tissue Eng. Part B Rev. 2020, 26, 105–115. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamon, M.; Cheng, H.-M.; Johnson, M.; Yanagawa, N.; Hauser, P.V. Effect of Hypoxia on Branching Characteristics and Cell Subpopulations during Kidney Organ Culture. Bioengineering 2022, 9, 801. https://doi.org/10.3390/bioengineering9120801
Hamon M, Cheng H-M, Johnson M, Yanagawa N, Hauser PV. Effect of Hypoxia on Branching Characteristics and Cell Subpopulations during Kidney Organ Culture. Bioengineering. 2022; 9(12):801. https://doi.org/10.3390/bioengineering9120801
Chicago/Turabian StyleHamon, Morgan, Hsiao-Min Cheng, Ming Johnson, Norimoto Yanagawa, and Peter V. Hauser. 2022. "Effect of Hypoxia on Branching Characteristics and Cell Subpopulations during Kidney Organ Culture" Bioengineering 9, no. 12: 801. https://doi.org/10.3390/bioengineering9120801