
Construction of HER2-Specific HIV-1-Based VLPs
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
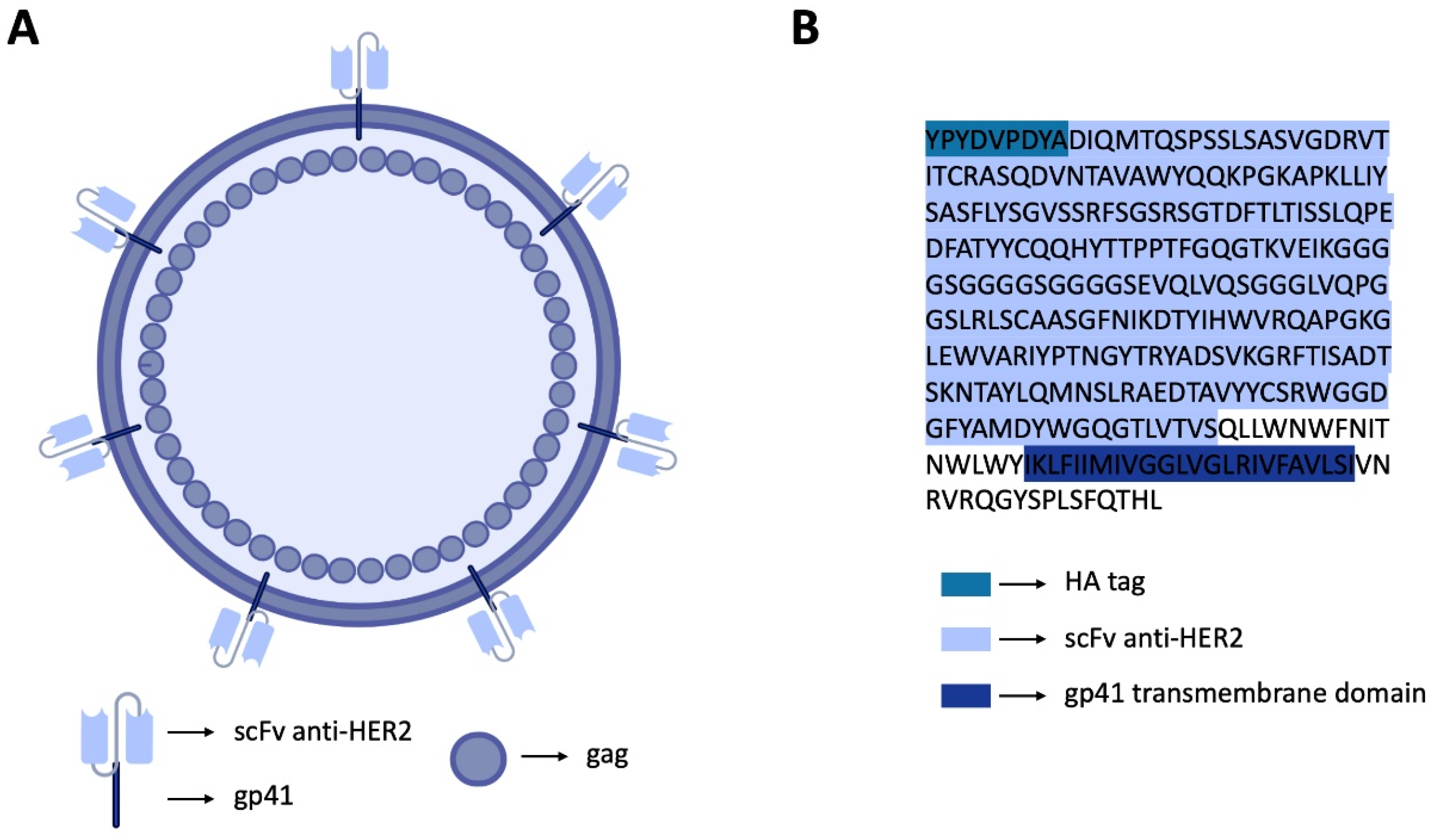
2.1. Plasmid Construction and Transformation
2.2. Cell Culture
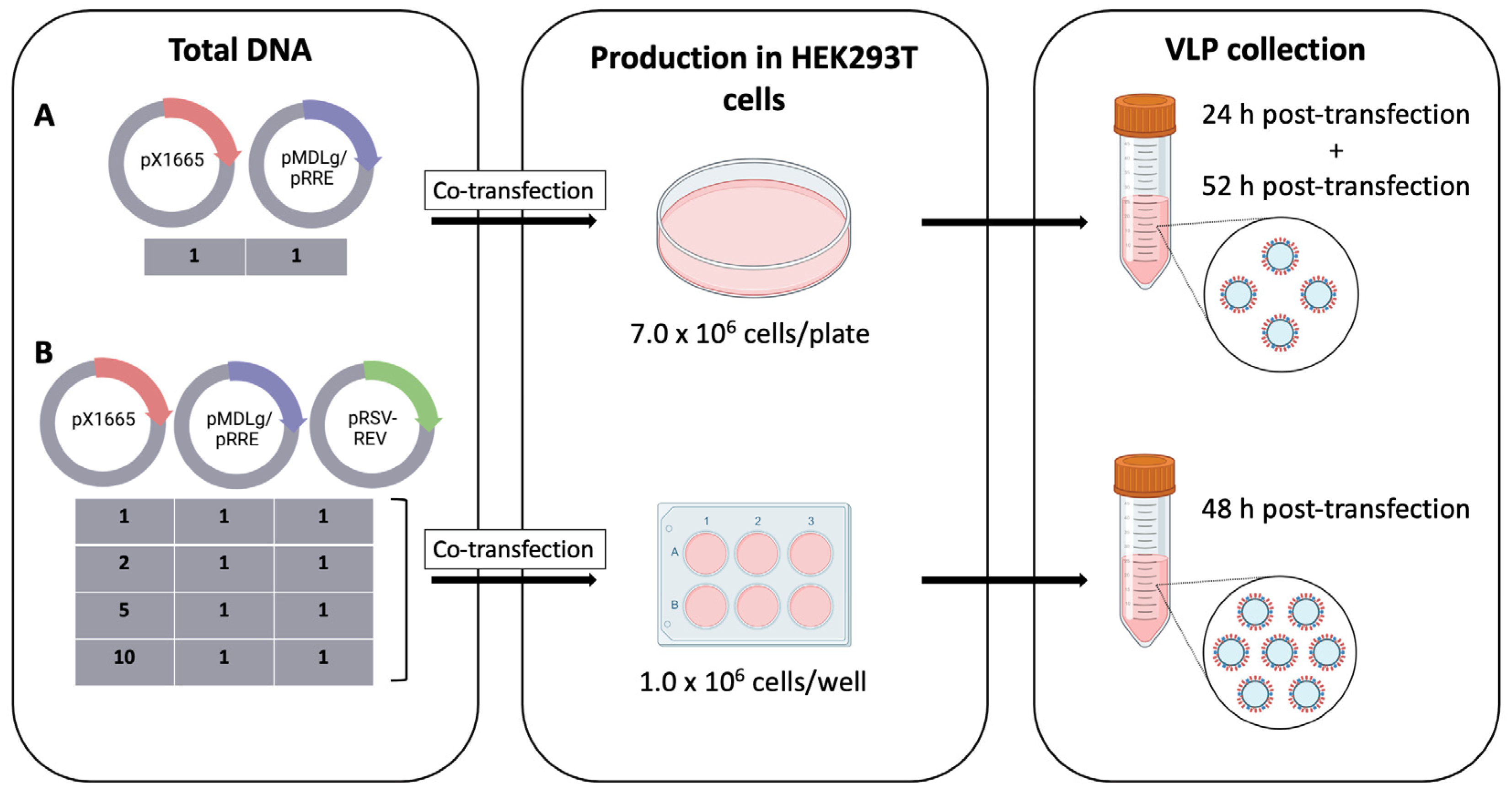
2.3. Transfection
2.4. Enzyme-Linked Immunosorbent Assay
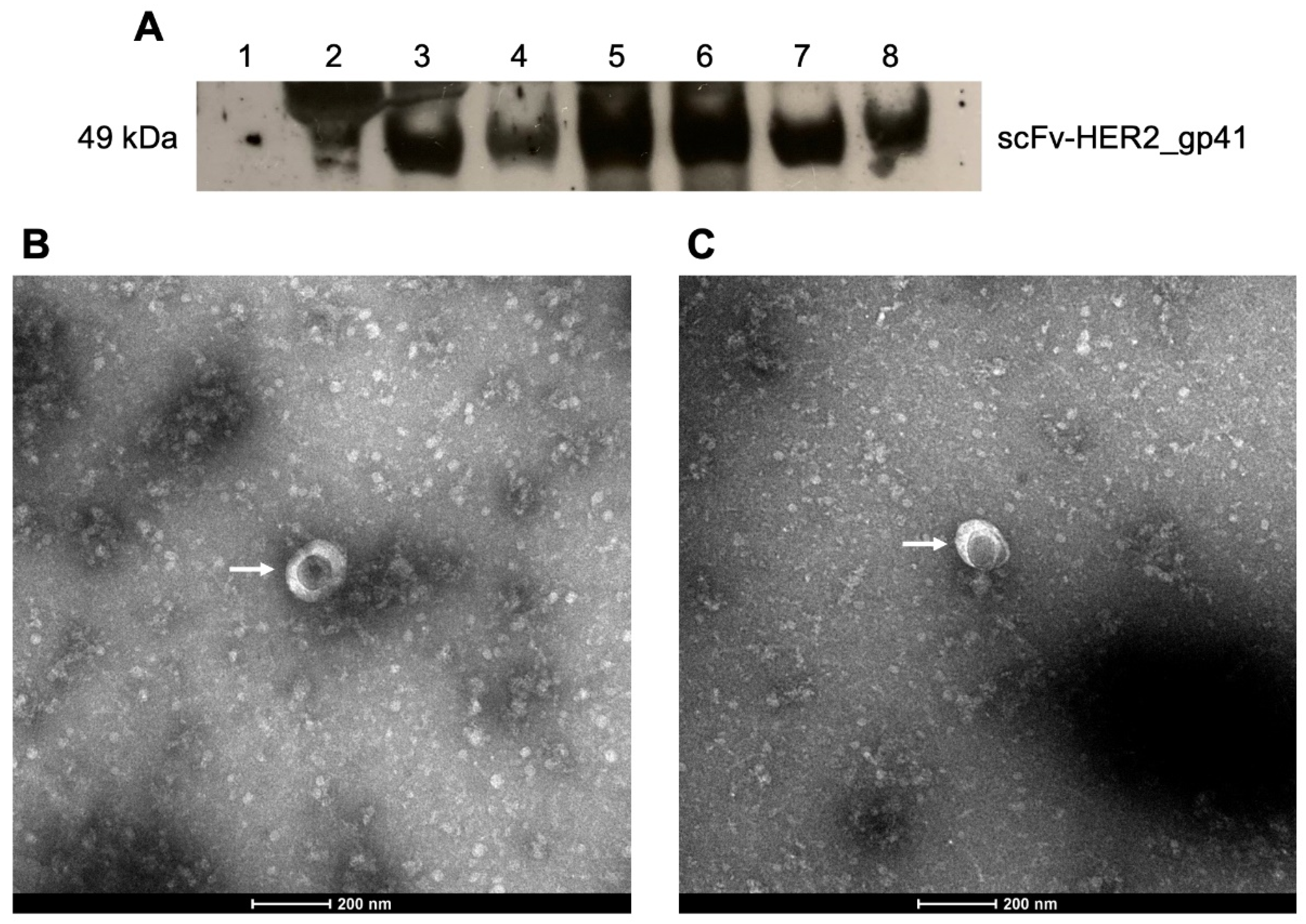
2.5. Western Blot
2.6. Transmission Electron Microscopy
2.7. Cytotoxicity Assay
3. Results
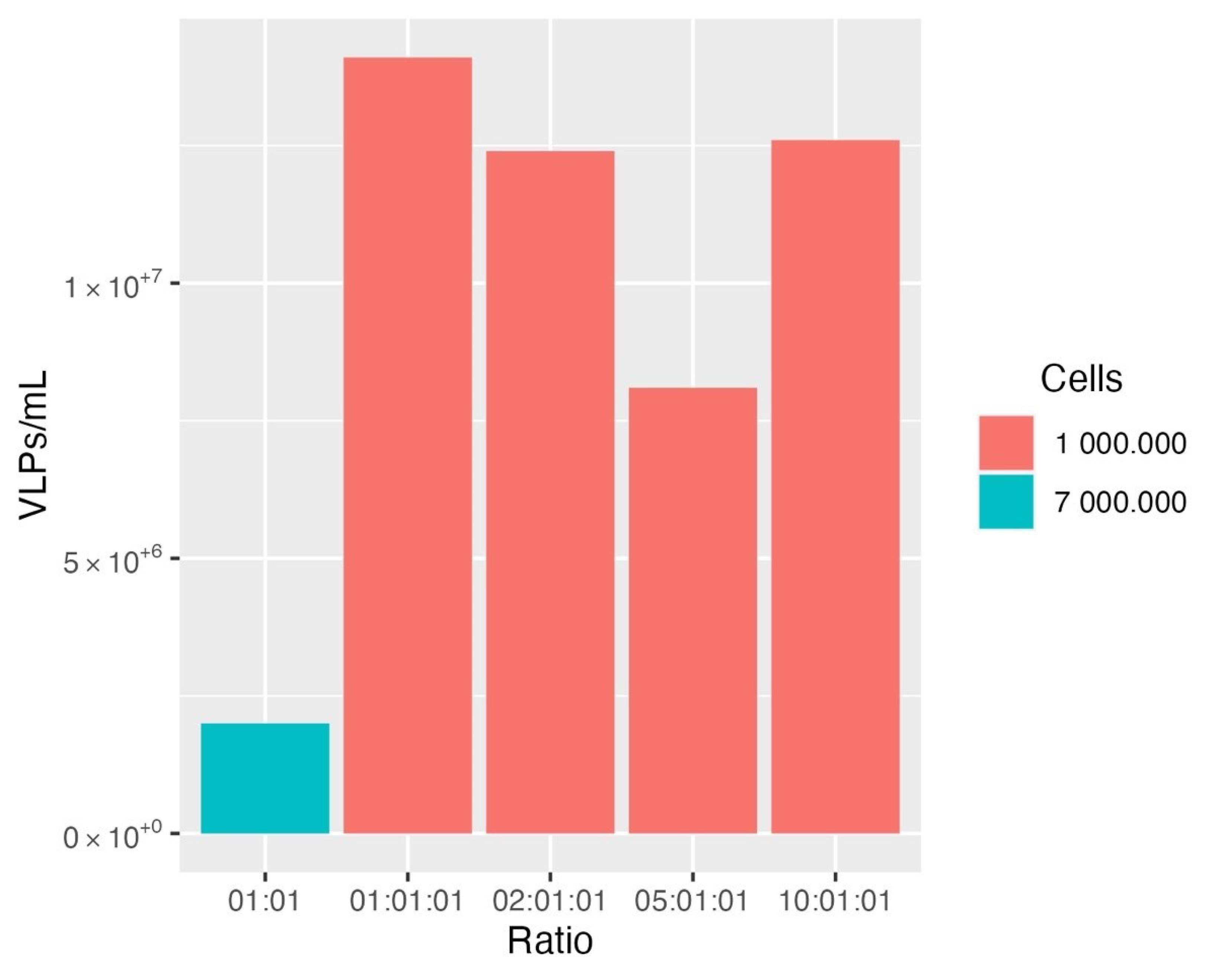
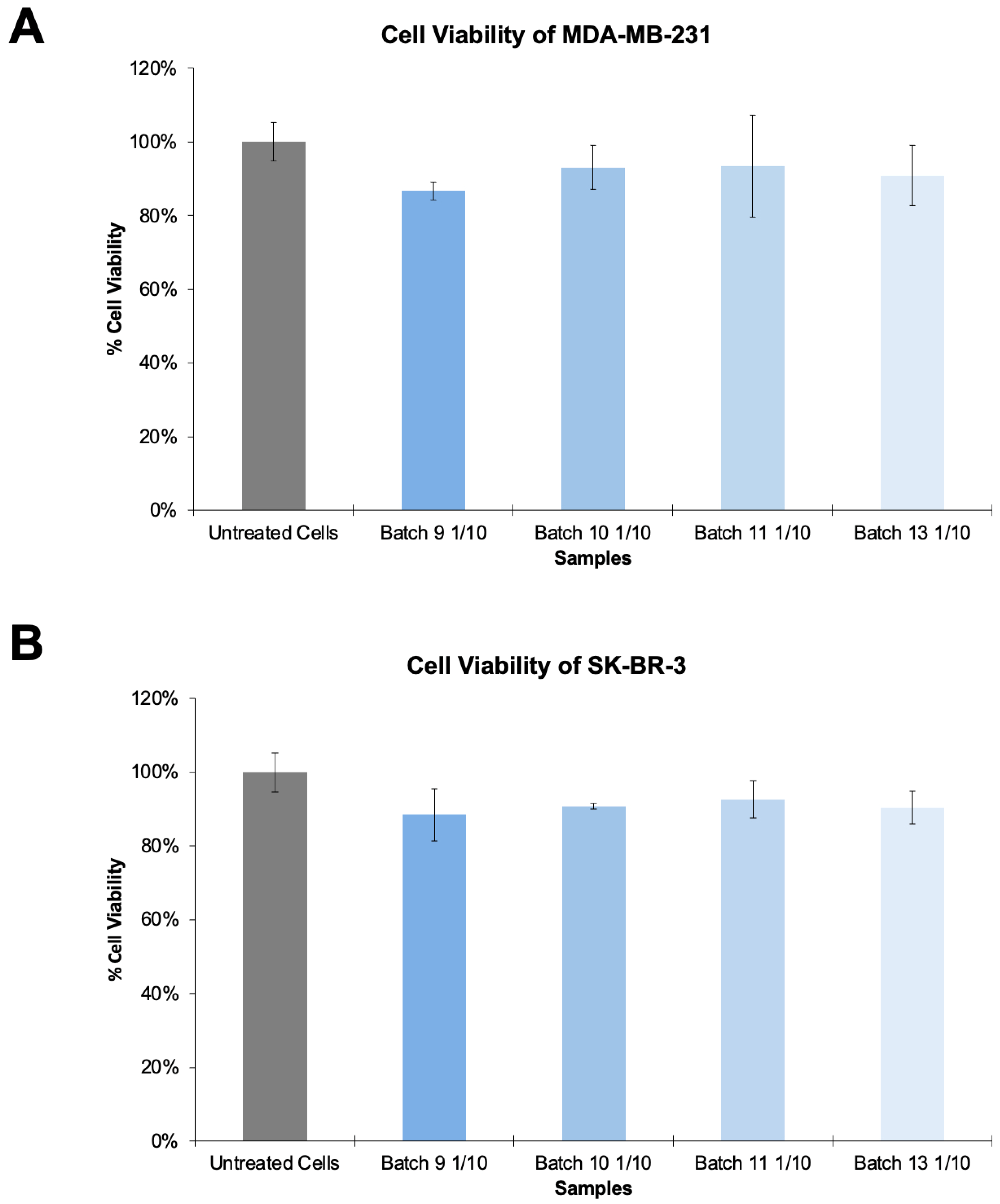
3.1. Production and Optimization of HER2-Specific HIV-1-Based VLPs
3.2. Production and Optimization of HER2-Specific HIV-1-Based VLPs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le, D.T.; Muller, K.M. In Vitro Assembly of Virus-Like Particles and Their Applications. Life 2021, 11, 334. [Google Scholar] [CrossRef]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-Like Particles: Preparation, Immunogenicity and Their Roles as Nanovaccines and Drug Nanocarriers. J. Nanobiotechnology 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Koudelka, K.J.; Pitek, A.S.; Manchester, M.; Steinmetz, N.F. Virus-Based Nanoparticles as Versatile Nanomachines. Annu. Rev. Virol. 2015, 2, 379–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateu, M.G. Virus Engineering: Functionalization and Stabilization. Protein Eng. Des. Sel. 2011, 24, 53–63. [Google Scholar] [CrossRef]
- Zackova Suchanova, J.; Hejtmankova, A.; Neburkova, J.; Cigler, P.; Forstova, J.; Spanielova, H. The Protein Corona Does Not Influence Receptor-Mediated Targeting of Virus-like Particles. Bioconjug. Chem. 2020, 31, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Rohovie, M.J.; Nagasawa, M.; Swartz, J.R. Virus-Like Particles: Next-Generation Nanoparticles for Targeted Therapeutic Delivery. Bioeng. Transl. Med. 2017, 2, 43–57. [Google Scholar] [CrossRef]
- Chung, Y.H.; Cai, H.; Steinmetz, N.F. Viral Nanoparticles for Drug Delivery, Imaging, Immunotherapy, and Theranostic Applications. Adv. Drug Deliv. Rev. 2020, 156, 214–235. [Google Scholar] [CrossRef]
- Piddock, L.J. Multidrug-Resistance Efflux Pumps—Not Just for Resistance. Nat. Rev. Microbiol. 2006, 4, 629–636. [Google Scholar] [CrossRef]
- Franco, D.; Liu, W.; Gardiner, D.F.; Hahn, B.H.; Ho, D.D. CD40L-Containing Virus-Like Particle as a Candidate HIV-1 Vaccine Targeting Dendritic Cells. J. Acquir. Immune Defic. Syndr. 2011, 56, 393–400. [Google Scholar] [CrossRef]
- Beltran-Pavez, C.; Bontjer, I.; Gonzalez, N.; Pernas, M.; Merino-Mansilla, A.; Olvera, A.; Miro, J.M.; Brander, C.; Alcami, J.; Sanders, R.W.; et al. Potent Induction of Envelope-Specific Antibody Responses by Virus-Like Particle Immunogens Based on HIV-1 Envelopes from Patients with Early Broadly Neutralizing Responses. J. Virol. 2022, 96, e0134321. [Google Scholar] [CrossRef]
- Bednarska, J.; Pelchen-Matthews, A.; Novak, P.; Burden, J.J.; Summers, P.A.; Kuimova, M.K.; Korchev, Y.; Marsh, M.; Shevchuk, A. Rapid Formation of Human Immunodeficiency Virus-Like Particles. Proc. Natl. Acad. Sci. USA 2020, 117, 21637–21646. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.A.; Santos, J.; Silva, R.D.; Rosa, C.; Cabo Verde, S.; Galamba Correia, J.D.; Melo, R. How Promising Are HIV-1-Based Virus-Like Particles for Medical Applications? Front. Cell. Infect. Microbiol. 2022, 12, 1526. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.; Cardoso, M.; Moreira, I.S.; Goncalves, J.; Correia, J.D.G.; Verde, S.C.; Melo, R. Integrated In Silico and Experimental Approach Towards the Design of A Novel Recombinant Protein Containing an Anti-HER2 scFv. Int. J. Mol. Sci. 2021, 22, 3547. [Google Scholar] [CrossRef] [PubMed]
- Cervera, L.; Gòdia, F.; Tarrés-Freixas, F.; Aguilar-Gurrieri, C.; Carrillo, J.; Blanco, J.; Gutiérrez-Granados, S. Production of HIV-1-Based Virus-Like Particles for Vaccination: Achievements and Limits. Appl. Microbiol. Biotechnol. 2019, 103, 7367–7384. [Google Scholar] [CrossRef] [PubMed]
- Scientific, T. Improve Lentiviral Production Using Lipofectamine 3000 Reagent. Available online: https://www.thermofisher.com/pt/en/home/life-science/cell-culture/cell-culture-learning-center/cell-culture-resource-library/cell-culture-transfection-application-notes/improve-lentiviral-production-using-lipofectamine-3000-reagent.html (accessed on 10 November 2022).
- Li, W.Y.; Yin, S.; Huang, S.W.; Yang, M.H.; Chen, P.M.; Wu, S.R.; Welsher, K.; Yang, H.; Arthur Chen, Y.M. The Trajectory Patterns of Single HIV-1 Virus-Like Particle in Live CD4 Cells: A Real Time Three-Dimensional Multi-Resolution Microscopy Study Using Encapsulated Nonblinking Giant Quantum Dot. J. Microbiol. Immunol. Infect. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Fuenmayor, J.; Godia, F.; Cervera, L. Production of Virus-Like Particles for Vaccines. New Biotechnol. 2017, 39, 174–180. [Google Scholar] [CrossRef]
- Lessard, J.C. Transformation of E. coli Via electroporation. Methods Enzymol. 2013, 529, 321–327. [Google Scholar] [CrossRef]
- Briggs, J.A.; Simon, M.N.; Gross, I.; Krausslich, H.G.; Fuller, S.D.; Vogt, V.M.; Johnson, M.C. The Stoichiometry of Gag Protein in HIV-1. Nat. Struct. Mol. Biol. 2004, 11, 672–675. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Steppert, P.; Burgstaller, D.; Klausberger, M.; Berger, E.; Aguilar, P.P.; Schneider, T.A.; Kramberger, P.; Tover, A.; Nobauer, K.; Razzazi-Fazeli, E.; et al. Purification of HIV-1 Gag Virus-Like Particles and Separation of Other Extracellular Particles. J. Chromatogr. A 2016, 1455, 93–101. [Google Scholar] [CrossRef]
- Gonzalez-Dominguez, I.; Puente-Massaguer, E.; Cervera, L.; Godia, F. Quality Assessment of Virus-Like Particles at Single Particle Level: A Comparative Study. Viruses 2020, 12, 223. [Google Scholar] [CrossRef] [Green Version]
- Hanne, J.; Zila, V.; Heilemann, M.; Muller, B.; Krausslich, H.G. Super-Resolved Insights into Human Immunodeficiency Virus Biology. FEBS Lett. 2016, 590, 1858–1876. [Google Scholar] [CrossRef] [PubMed]
- Roldao, A.; Mellado, M.C.; Castilho, L.R.; Carrondo, M.J.; Alves, P.M. Virus-Like Particles in Vaccine Development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef] [PubMed]
- Boix-Besora, A.; Lorenzo, E.; Lavado-Garcia, J.; Godia, F.; Cervera, L. Optimization, Production, Purification and Characterization of HIV-1 GAG-Based Virus-like Particles Functionalized with SARS-CoV-2. Vaccines 2022, 10, 250. [Google Scholar] [CrossRef] [PubMed]
- Fontana, D.; Garay, E.; Cervera, L.; Kratje, R.; Prieto, C.; Godia, F. Chimeric VLPs Based on HIV-1 Gag and a Fusion Rabies Glycoprotein Induce Specific Antibodies against Rabies and Foot-and-Mouth Disease Virus. Vaccines 2021, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.A.; Lytvyn, V.; Deforet, F.; Gilbert, R.; Gaillet, B. Virus-Like Particles Derived from HIV-1 for Delivery of Nuclear Proteins: Improvement of Production and Activity by Protein Engineering. Mol. Biotechnol. 2017, 59, 9–23. [Google Scholar] [CrossRef]
- Lyu, P.; Javidi-Parsijani, P.; Atala, A.; Lu, B. Delivering Cas9/sgRNA Ribonucleoprotein (RNP) by Lentiviral Capsid-Based Bionanoparticles for Efficient ‘Hit-and-Run’ Genome Editing. Nucleic Acids Res. 2019, 47, e99. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.R.; Tsuchida, C.A.; Nguyen, D.N.; Shy, B.R.; McGarrigle, E.R.; Sandoval Espinoza, C.R.; Carr, D.; Blaeschke, F.; Marson, A.; Doudna, J.A. Targeted Delivery of CRISPR-Cas9 and Transgenes Enables Complex Immune Cell Engineering. Cell Rep. 2021, 35, 109207. [Google Scholar] [CrossRef]
- Chojnacki, J.; Staudt, T.; Glass, B.; Bingen, P.; Engelhardt, J.; Anders, M.; Schneider, J.; Muller, B.; Hell, S.W.; Krausslich, H.G. Maturation-Dependent HIV-1 Surface Protein Redistribution Revealed by Fluorescence Nanoscopy. Science 2012, 338, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Steppert, P.; Burgstaller, D.; Klausberger, M.; Kramberger, P.; Tover, A.; Berger, E.; Nobauer, K.; Razzazi-Fazeli, E.; Jungbauer, A. Separation of HIV-1 Gag Virus-Like Particles from Vesicular Particles Impurities by Hydroxyl-Functionalized Monoliths. J. Sep. Sci. 2017, 40, 979–990. [Google Scholar] [CrossRef]
- Gupta, S.K.; Shukla, P. Glycosylation Control Technologies for Recombinant Therapeutic Proteins. Appl. Microbiol. Biotechnol. 2018, 102, 10457–10468. [Google Scholar] [CrossRef] [PubMed]
- Loibl, S.; Gianni, L. HER2-Positive Breast Cancer. Lancet 2017, 389, 2415–2429. [Google Scholar] [CrossRef]
- Boku, N. HER2-Positive Gastric Cancer. Gastric Cancer 2014, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuenmayor, J.; Cervera, L.; Gutiérrez-Granados, S.; Gòdia, F. Transient Gene Expression Optimization and Expression Vector Comparison to Improve HIV-1 VLP Production in HEK293 Cell Lines. Appl. Microbiol. Biotechnol. 2018, 102, 165–174. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pX1665: pMDLg/pRRE: pRSV-REV Ratio | Mass (μg) | ||
|---|---|---|---|
| pX1665 | pMDLg/pRRE | pRSV-REV | |
| 1:1:1 | 1.02 | 1.35 | 0.63 |
| 2:1:1 | 1.52 | 1.01 | 0.47 |
| 5:1:1 | 2.16 | 0.57 | 0.27 |
| 10:1:1 | 2.51 | 0.33 | 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, S.A.; Santos, J.; Cabo Verde, S.; Correia, J.D.G.; Melo, R. Construction of HER2-Specific HIV-1-Based VLPs. Bioengineering 2022, 9, 713. https://doi.org/10.3390/bioengineering9110713
Martins SA, Santos J, Cabo Verde S, Correia JDG, Melo R. Construction of HER2-Specific HIV-1-Based VLPs. Bioengineering. 2022; 9(11):713. https://doi.org/10.3390/bioengineering9110713
Chicago/Turabian StyleMartins, Sofia A., Joana Santos, Sandra Cabo Verde, João D. G. Correia, and Rita Melo. 2022. "Construction of HER2-Specific HIV-1-Based VLPs" Bioengineering 9, no. 11: 713. https://doi.org/10.3390/bioengineering9110713