
Adhesion Forces of Oral Bacteria to Titanium and the Correlation with Biophysical Cellular Characteristics
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
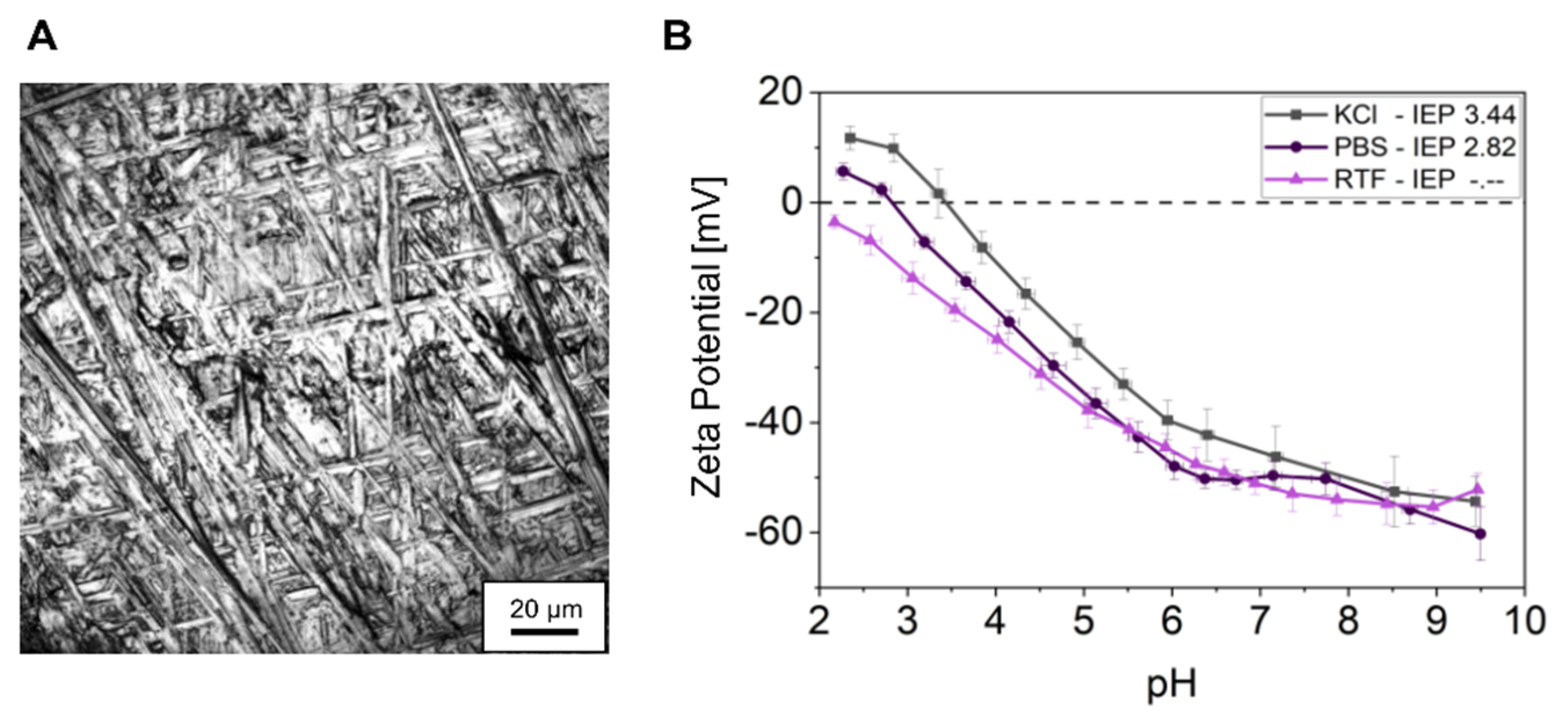
2.1. Titanium Specimen Characterization
2.2. Bacterial Strains and Culture Conditions
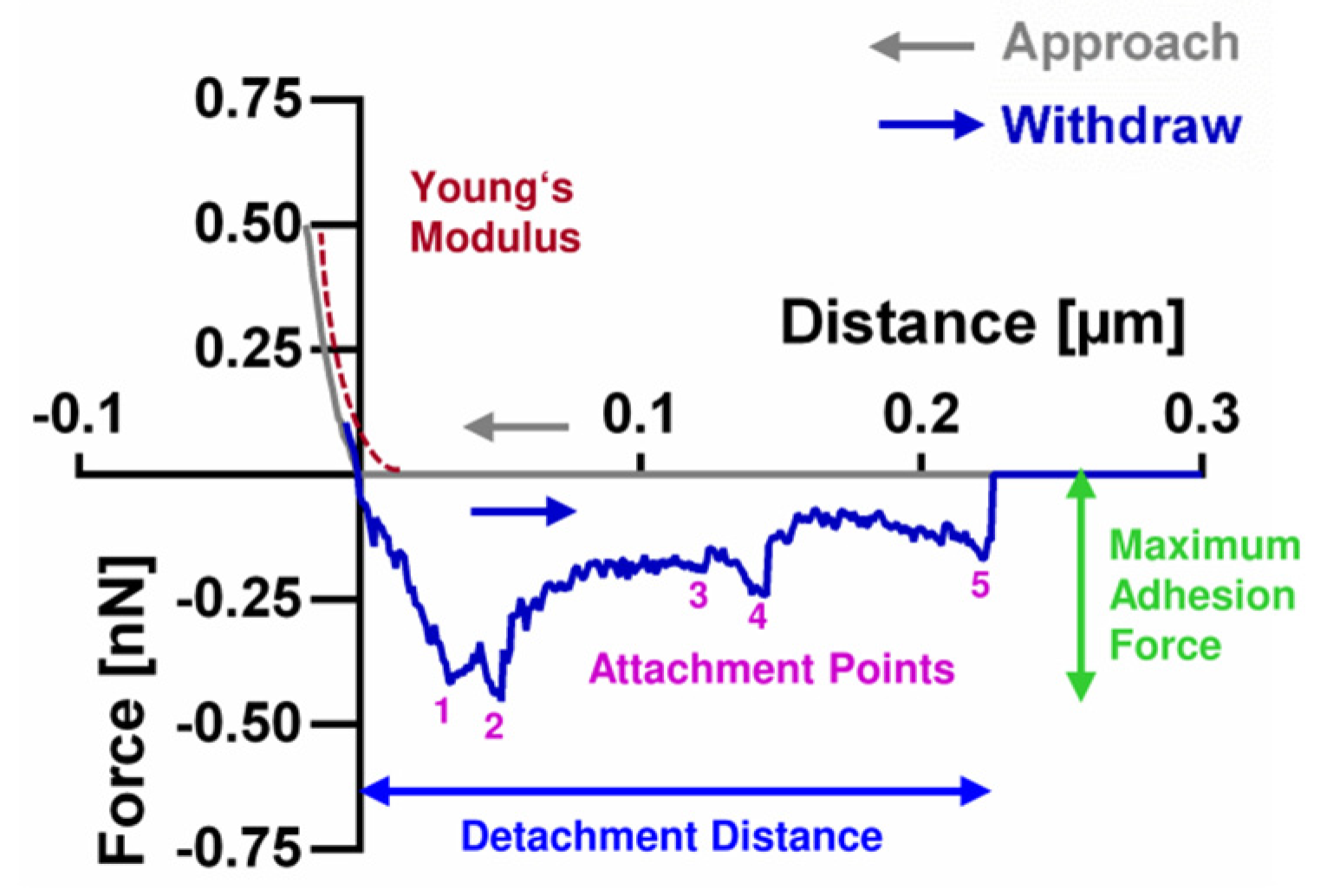
2.3. Bacterial Single-Cell Force Spectroscopy (SCFS)
2.4. Bacterial Zeta Potential Measurement
2.5. Fluorescence Staining and Confocal Laser-Scanning Microscopy
2.6. BacTiter-GloTM Assay
2.7. Statistical Analysis
3. Results
3.1. Titanium Specimen Characteristics
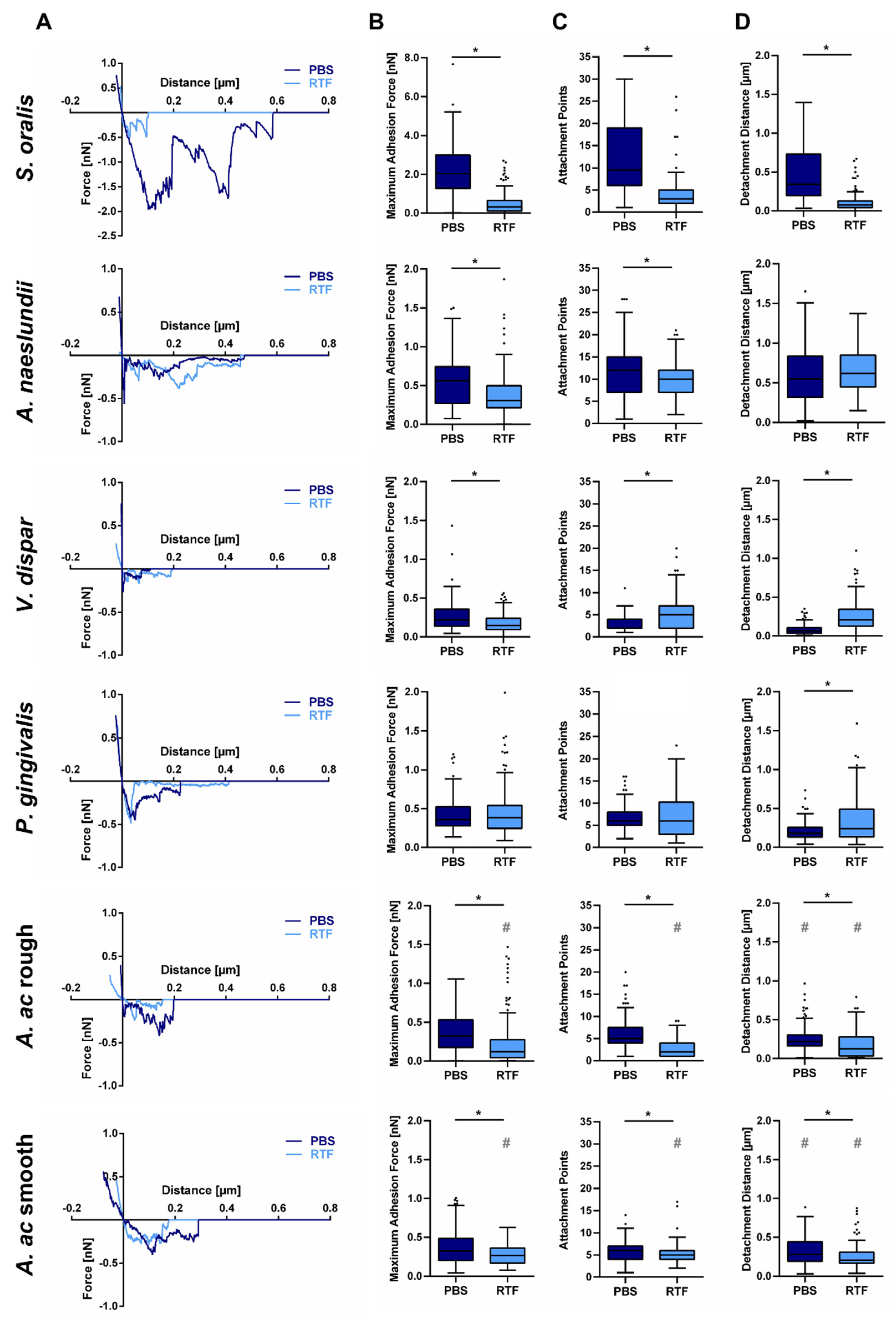
3.2. Strain- and Buffer-Dependent Adhesion Forces of Oral Bacteria to Titanium
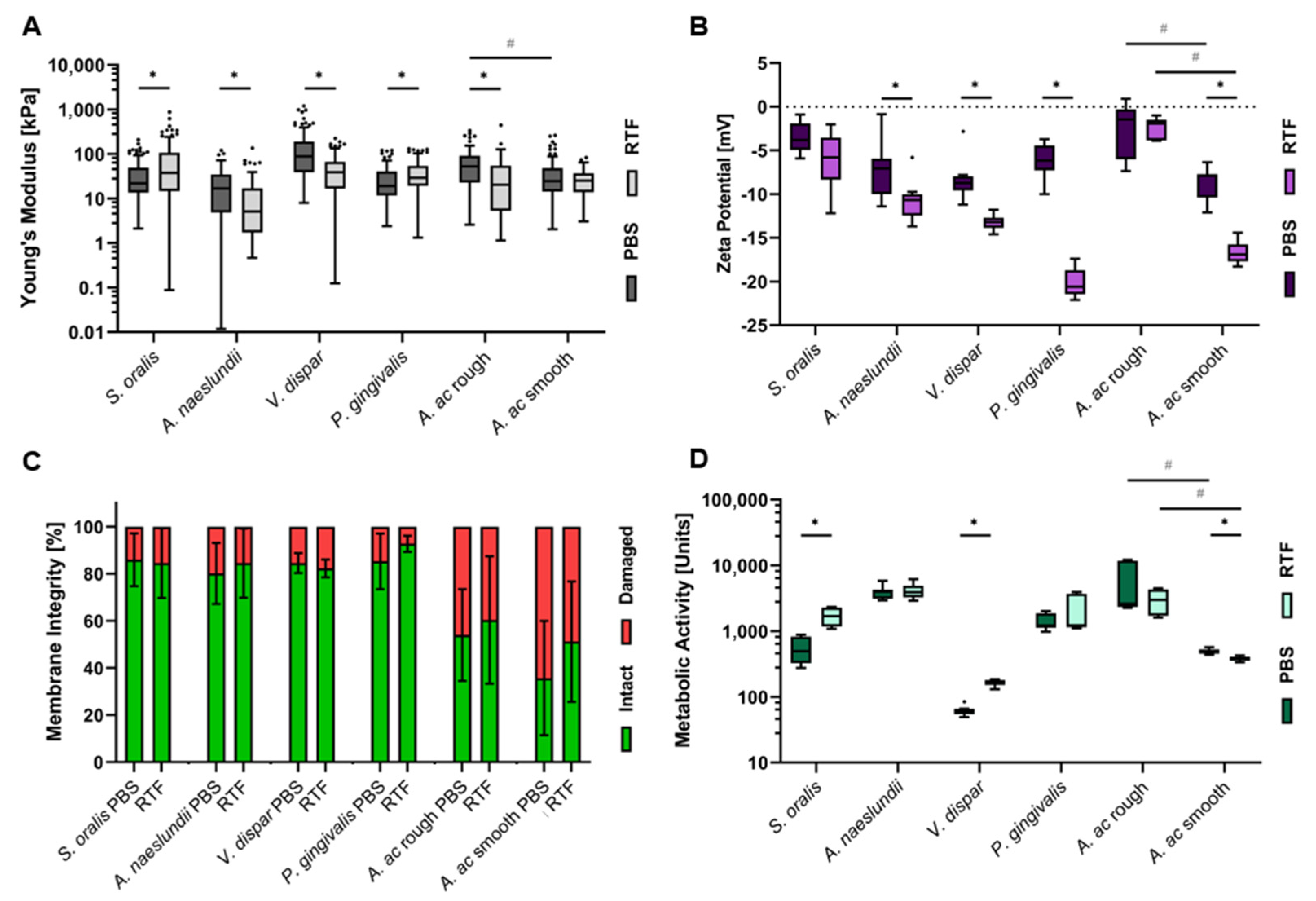
3.3. Strain- and Buffer-Dependent Biophysical Cellular Characteristics
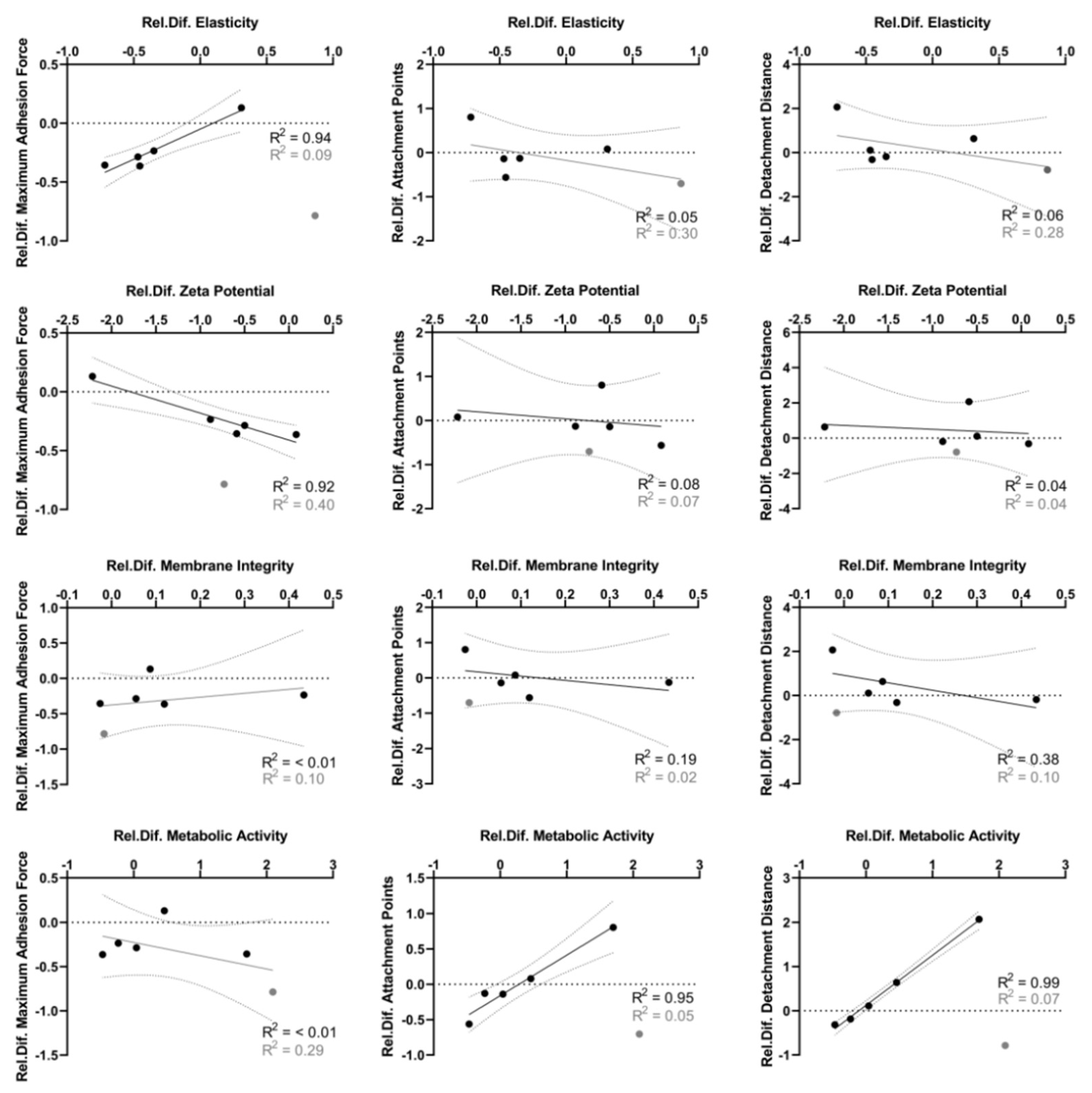
3.4. Distinct Correlation between Adhesion Force Parameters and Biophysical Cellular Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C.; Wingender, J. Biofilme—Die bevorzugte Lebensform der Bakterien. Biol. Unserer Zeit 2001, 31, 169–180. [Google Scholar] [CrossRef]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef] [Green Version]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Huang, R.; Li, M.; Gregory, R.L. Bacterial interactions in dental biofilm. Virulence 2011, 2, 435–444. [Google Scholar] [CrossRef]
- Kolenbrander, P.E.; Palmer, R.J., Jr.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell–cell distance. Nat. Rev. Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, C.; Chen, Z.; Allan, E.; van der Mei, H.C.; Busscher, H.J. Emergent heterogeneous microenvironments in biofilms: Substratum surface heterogeneity and bacterial adhesion force-sensing. FEMS Microbiol. Rev. 2018, 42, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Dufrêne, Y.F. Sticky microbes: Forces in microbial cell adhesion. Trends Microbiol. 2015, 23, 376–382. [Google Scholar] [CrossRef]
- Meister, A.; Gabi, M.; Behr, P.; Studer, P.; Voros, J.; Niedermann, P.; Bitterli, J.; Polesel-Maris, J.; Liley, M.; Heinzelmann, H.; et al. FluidFM: Combining atomic force micros-copy and nanofluidics in a universal liquid delivery system for single cell applications and beyond. Nano Lett. 2009, 9, 2501–2507. [Google Scholar] [CrossRef]
- Guillaume-Gentil, O.; Potthoff, E.; Ossola, D.; Franz, C.M.; Zambelli, T.; Vorholt, J.A. Force-controlled manipulation of single cells: From AFM to FluidFM. Trends Biotechnol. 2014, 32, 381–388. [Google Scholar] [CrossRef]
- Potthoff, E.; Ossola, D.; Zambelli, T.; Vorholt, J.A. Bacterial adhesion force quantification by fluidic force microscopy. Nanoscale 2015, 7, 4070–4079. [Google Scholar] [CrossRef]
- Hofherr, L.; Müller-Renno, C.; Ziegler, C. FluidFM as a tool to study adhesion forces of bacteria—Optimization of parameters and comparison to conventional bacterial probe Scanning Force Spectroscopy. PLoS ONE 2020, 15, e0227395. [Google Scholar] [CrossRef]
- Pattem, J.; Davrandi, M.; Aguayo, S.; Allan, E.; Spratt, D.; Bozec, L. A Multi-scale Biophysical Approach to Develop Structure-Property Relationships in Oral Biofilms. Sci. Rep. 2018, 8, 5691. [Google Scholar] [CrossRef] [Green Version]
- Viljoen, A.; Mignolet, J.; Viela, F.; Mathelié-Guinlet, M.; Dufrêne, Y.F. How Microbes Use Force To Control Adhesion. J. Bacteriol. 2020, 202, e00125-20. [Google Scholar] [CrossRef]
- Loskill, P.; Zeitz, C.; Grandthyll, S.; Thewes, N.; Müller, F.; Bischoff, M.; Herrmann, M.; Jacobs, K. Reduced Adhesion of Oral Bacteria on Hydroxyapatite by Fluoride Treatment. Langmuir 2013, 29, 5528–5533. [Google Scholar] [CrossRef]
- Mei, L.; Busscher, H.J.; Van Der Mei, H.C.; Chen, Y.; De Vries, J.; Ren, Y. Oral bacterial adhesion forces to biomaterial surfaces constituting the bracket-adhesive-enamel junction in orthodontic treatment. Eur. J. Oral Sci. 2009, 117, 419–426. [Google Scholar] [CrossRef]
- Wessel, S.; Chen, Y.; Maitra, A.; van den Heuvel, E.; Slomp, A.M.; Busscher, H.J.; van der Mei, H.C. Adhesion Forces and Composition of Planktonic and Adhering Oral Microbiomes. J. Dent. Res. 2014, 93, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Van Der Mei, H.C.; Busscher, H.J.; Ren, Y. Streptococcus mutans adhesion force sensing in multi-species oral biofilms. npj Biofilms Microbiomes 2020, 6, 25. [Google Scholar] [CrossRef]
- Elbourne, A.; Chapman, J.; Gelmi, A.; Cozzolino, D.; Crawford, R.J.; Truong, V.K. Bacterial-nanostructure interactions: The role of cell elasticity and adhesion forces. J. Colloid Interface Sci. 2019, 546, 192–210. [Google Scholar] [CrossRef]
- Song, F.; Koo, H.; Ren, D. Effects of Material Properties on Bacterial Adhesion and Biofilm Formation. J. Dent. Res. 2015, 94, 1027–1034. [Google Scholar] [CrossRef]
- Maillard, A.P.F.; Espeche, J.C.; Maturana, P.; Cutro, A.C.; Hollmann, A. Zeta potential beyond materials science: Applications to bacterial systems and to the development of novel antimicrobials. Biochim. Biophys. Acta BBA—Biomembr. 2021, 1863, 183597. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.P.; Truong, V.K.; Webb, H.K.; Baulin, V.A.; Wang, J.Y.; Mohammodi, N.; Wang, F.; Fluke, C.; Crawford, R.J. Differential attraction and repulsion of Staphylococcus aureus and Pseudomonas aeruginosa on molecularly smooth titanium films. Sci. Rep. 2011, 1, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.H.; Friedman, R.J. Concise review of mechanisms of bacterial adhesion to biomaterial surfaces. J. Biomed. Mater. Res. 1998, 43, 338–348. [Google Scholar] [CrossRef]
- Syed, S.A.; Loesche, W.J. Survival of human dental plaque flora in various transport media. Appl. Microbiol. 1972, 24, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Ingendoh-Tsakmakidis, A.; Mikolai, C.; Winkel, A.; Szafrański, S.P.; Falk, C.S.; Rossi, A.; Walles, H.; Stiesch, M. Commensal and pathogenic biofilms differently modulate peri-implant oral mucosa in an organotypic model. Cell. Microbiol. 2019, 21, e13078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobbs, A.H.; Lamont, R.J.; Jenkinson, H.F. Streptococcus Adherence and Colonization. Microbiol. Mol. Biol. Rev. 2009, 73, 407–450. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, C.J.; Klier, C.M.; Kolenbrander, P.E. Mechanisms of adhesion by oral bacteria. Annu. Rev. Microbiol. 1996, 50, 513–552. [Google Scholar] [CrossRef]
- Mayrand, D.; Holt, S.C. Biology of asaccharolytic black-pigmented bacteroides species. Microbiol. Rev. 1988, 52, 134–152. [Google Scholar] [CrossRef]
- Inoue, T.; Shingaki, R.; Sogawa, N.; Sogawa, C.A.; Asaumi, J.; Kokeguchi, S.; Fukui, K. Biofilm formation by a fimbriae-deficient mutant of actinobacillus actinomycetemcomitans. Microbiol. Immunol. 2003, 47, 877–881. [Google Scholar] [CrossRef]
- Sader, J.E.; Chon, J.W.M.; Mulvaney, P. Calibration of rectangular atomic force microscope cantilevers. Rev. Sci. Instrum. 1999, 70, 3967–3969. [Google Scholar] [CrossRef]
- Hermanowicz, P.; Sarna, M.; Burda, K.; Gabryś, H. AtomicJ: An open source software for analysis of force curves. Rev. Sci. Instrum. 2014, 85, 063703. [Google Scholar] [CrossRef] [PubMed]
- Doll, K.; Yang, I.; Fadeeva, E.; Kommerein, N.; Szafrański, S.P.; der Wieden, G.B.; Greuling, A.; Winkel, A.; Chichkov, B.N.; Stumpp, N.S.; et al. Liquid-Infused Structured Titanium Surfaces: Antiadhesive Mechanism to Repel Streptococcus oralis Biofilms. ACS Appl. Mater. Interfaces 2019, 11, 23026–23038. [Google Scholar] [CrossRef]
- Doll, K.; Jongsthaphongpun, K.L.; Stumpp, N.S.; Winkel, A.; Stiesch, M. Quantifying implant-associated biofilms: Comparison of microscopic, microbiologic and biochemical methods. J. Microbiol. Methods 2016, 130, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Doll, K.; Fadeeva, E.; Schaeske, J.; Ehmke, T.; Winkel, A.; Heisterkamp, A.; Chichkov, B.; Stiesch, M.; Stumpp, N.S. Development of Laser-Structured Liquid-Infused Titanium with Strong Biofilm-Repellent Properties. ACS Appl. Mater. Interfaces 2017, 9, 9359–9368. [Google Scholar] [CrossRef] [PubMed]
- Kommerein, N.; Doll, K.; Stumpp, N.S.; Stiesch, M. Development and characterization of an oral multispecies biofilm implant flow chamber model. PLoS ONE 2018, 13, e0196967. [Google Scholar] [CrossRef] [Green Version]
- Subh, L.; Correa, W.; Pinkvos, T.J.; Behrens, P.; Brandenburg, K.; Gutsmann, T.; Stiesch, M.; Doll, K.; Winkel, A. Synthetic anti-endotoxin peptides interfere with gram-positive and gram-negative bacteria, their adhesion and biofilm formation on titanium. J. Appl. Microbiol. 2020, 129, 1272–1286. [Google Scholar] [CrossRef]
- Rath, H.; Stumpp, S.N.; Stiesch, M. Development of a flow chamber system for the reproducible in vitro analysis of biofilm formation on implant materials. PLoS ONE 2017, 12, e0172095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desch, A.; Von Maltzahn, N.F.; Stumpp, N.; Dalton, M.; Yang, I.; Stiesch, M. Biofilm formation on zirconia and titanium over time—An in vivo model study. Clin. Oral Implant. Res. 2020, 31, 865–880. [Google Scholar] [CrossRef]
- Von Maltzahn, N.F.; Stumpp, N.S.; Stiesch, M. Antibacterial effect of cupral((R)) on oral biofilms—An in-vitro study. Eur. Endod. J. 2020, 5, 40–45. [Google Scholar]
- McCafferty, E.; Wightman, J.P. Determination of the concentration of surface hydroxyl groups on metal oxide films by a quantitative XPS method. Surf. Interface Anal. 1998, 26, 549–564. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, L.; Ding, X.; Gao, Y.; Liu, X. Biological Effect of Ultraviolet Photocatalysis on Nanoscale Titanium with a Focus on Physicochemical Mechanism. Langmuir 2015, 31, 10037–10046. [Google Scholar] [CrossRef]
- Li, B.; Logan, B.E. Bacterial adhesion to glass and metal-oxide surfaces. Colloids Surf. B Biointerfaces 2004, 36, 81–90. [Google Scholar] [CrossRef]
- Mei, L.; Ren, Y.; Busscher, H.J.; Chen, Y.; Van Der Mei, H.C. Poisson Analysis of Streptococcal Bond-strengthening on Saliva-coated Enamel. J. Dent. Res. 2009, 88, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Busscher, H.J.; van der Mei, H.C.; Ren, Y. Influence of surface roughness on streptococcal adhesion forces to composite resins. Dent. Mater. 2011, 27, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Aguayo, S.; Donos, N.; Spratt, D.; Bozec, L. Nanoadhesion of Staphylococcus aureus onto titanium implant surfaces. J. Dent. Res. 2015, 94, 1078–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Busscher, H.J.; van der Mei, H.C.; Norde, W. Statistical analysis of long- and short-range forces involved in bacterial adhesion to substratum surfaces as meas-ured using atomic force microscopy. Appl. Environ. Microbiol. 2011, 77, 5065–5070. [Google Scholar] [CrossRef] [Green Version]
- Bozec, L.; Donos, N.; Spratt, D.; Aguayo, S. Probing the nanoadhesion of Streptococcus sanguinis to titanium implant surfaces by atomic force microscopy. Int. J. Nanomed. 2016, 11, 1443–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermansson, M. The DLVO theory in microbial adhesion. Colloids Surf. B Biointerfaces 1999, 14, 105–119. [Google Scholar] [CrossRef]
- Spengler, C.; Thewes, N.; Nolle, F.; Faidt, T.; Umanskaya, N.; Hannig, M.; Bischoff, M.; Jacobs, K. Enhanced adhesion of Streptococcus mutans to hydroxyapatite after exposure to saliva. J. Mol. Recognit. 2017, 30, e2615. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, Y.; Zheng, S.; Xue, J.; Zhou, J.; Tang, Y.; Jiang, L.; Li, W. Effect of enamel morphology on nanoscale adhesion forces of streptococcal bacteria: An AFM study. Scanning 2015, 37, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Rahim, M.I.; Doll, K.; Stumpp, N.S.; Eisenburger, M.; Stiesch, M. Multilayered adsorption of commensal microflora on implant surfaces: An unconventional and innovative method to prevent bacterial infections associated with biomaterials. Adv. Mater. Interfaces 2021, 8, 2101410. [Google Scholar] [CrossRef]
- Kristensen, M.F.; Zeng, G.; Neu, T.R.; Meyer, R.L.; Baelum, V.; Schlafer, S. Osteopontin adsorption to Gram-positive cells reduces adhesion forces and attachment to surfaces under flow. J. Oral Microbiol. 2017, 9, 1379826. [Google Scholar] [CrossRef]
- Postollec, F.; Norde, W.; De Vries, J.; Busscher, H.J.; Van Der Mei, H.C. Interactive Forces between Co-aggregating and Non-co-aggregating Oral Bacterial Pairs. J. Dent. Res. 2006, 85, 231–234. [Google Scholar] [CrossRef]
- van der Mei, H.C.; Rustema-Abbing, M.; de Vries, J.; Busscher, H.J. Bond strengthening in oral bacterial adhesion to salivary conditioning films. Appl. Environ. Microbiol. 2008, 74, 5511–5515. [Google Scholar] [CrossRef] [Green Version]
- Bank, T.L.; Dosen, A.; Giese, R.F.; Haase, E.M.; Sojar, H.T. Atomic Force Spectroscopy Evidence of Non-Specific Adhesion of Aggregatibacter actinomycetemcomitans. J. Nanosci. Nanotechnol. 2011, 11, 8450–8456. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, A.; Chen, C. Genetic Basis for Conversion of Rough-to-Smooth Colony Morphology in Actinobacillus actinomycetemcomitans. Infect. Immun. 2005, 73, 3749–3753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, J.B.; Ragunath, C.; Ramasubbu, N.; Fine, D.H. Detachment of Actinobacillus actinomycetemcomitans biofilm cells by an endogenous beta-hexosaminidase activity. J. Bacteriol. 2003, 185, 4693–4698. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.; Ting, Y.P.; Pehkonen, S.O. The influence of ionic strength, nutrients and pH on bacterial adhesion to metals. J. Colloid Interface Sci. 2008, 321, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Vanzieleghem, T.; Couniot, N.; Herman-Bausier, P.; Flandre, D.; Dufrêne, Y.F.; Mahillon, J. Role of Ionic Strength in Staphylococcal Cell Aggregation. Langmuir 2016, 32, 7277–7283. [Google Scholar] [CrossRef]
- Tamayo, L.; Melo, F.; Caballero, L.; Hamm, E.; Diaz, M.; Leal, M.S.; Guiliani, N.; Urzua, M.D. Does bacterial elasticity affect adhesion to polymer fibers? ACS Appl. Mater. Interfaces 2020, 12, 14507–14517. [Google Scholar] [CrossRef]
- Doll, P.W.; Doll, K.; Winkel, A.; Thelen, R.; Ahrens, R.; Stiesch, M.; Guber, A.E. Influence of the Available Surface Area and Cell Elasticity on Bacterial Adhesion Forces on Highly Ordered Silicon Nanopillars. ACS Omega 2022, 7, 17620–17631. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; van der Mei, H.C.; Busscher, H.J.; Norde, W. Viscous Nature of the Bond between Adhering Bacteria and Substratum Surfaces Probed by Atomic Force Microscopy. Langmuir 2014, 30, 3165–3169. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Harapanahalli, A.K.; Busscher, H.J.; Norde, W.; van der Mei, H.C. Nanoscale Cell Wall Deformation Impacts Long-Range Bacterial Adhesion Forces on Surfaces. Appl. Environ. Microbiol. 2013, 80, 637–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bundeleva, I.A.; Shirokova, L.S.; Bénézeth, P.; Pokrovsky, O.S.; Kompantseva, E.I.; Balor, S. Zeta potential of anoxygenic phototrophic bacteria and Ca adsorption at the cell surface: Possible implications for cell protection from CaCO3 precipitation in alkaline solutions. J. Colloid Interface Sci. 2011, 360, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Brown, D.G. Cell surface acid-base properties of Escherichia coli and Bacillus brevis and variation as a function of growth phase, nitrogen source and C:N ratio. Colloids Surf. B Biointerfaces 2006, 50, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Poortinga, A.T.; Bos, R.; Norde, W.; Busscher, H.J. Electric double layer interactions in bacterial adhesion to surfaces. Surf. Sci. Rep. 2002, 47, 1–32. [Google Scholar] [CrossRef]
- Kreve, S.; Candido dos Reis, A. Influence of the electrostatic condition of the titanium surface on bacterial adhesion: A systematic review. J. Prosthet. Dent. 2021, 125, 416–420. [Google Scholar] [CrossRef]
- Netuschil, L.; Auschill, T.M.; Sculean, A.; Arweiler, N.B. Confusion over live/dead stainings for the detection of vital microorganisms in oral biofilms—Which stain is suitable? BMC Oral Health 2014, 14, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abelho, M. Extraction and quantification of ATP as a measure of microbial biomass. In Methods to Study Litter Decomposition: A Practical Guide; Springer: Dordrecht, The Netherlands, 2005; pp. 223–229. [Google Scholar]
- Sule, P.; Wadhawan, T.; Carr, N.J.; Horne, S.M.; Wolfe, A.J.; Pruss, B.M. A combination of assays reveals biomass differences in biofilms formed by Escherichia coli mutants. Lett. Appl. Microbiol. 2009, 49, 299–304. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Shape | Size [µm] | Metabolism | Cell Wall | Adhesion Structures | Characteristic in Oral Biofilm |
|---|---|---|---|---|---|---|
| S. oralis | coccoid | 0.75 × 0.75 | facultative anaerobe | Gram+ | fibrils adhesins [26] | commensal bacterium initial colonizer |
| A. naeslundii | bacillus | 3 × 0.75 | facultative anaerobe | Gram+ | fimbriae [27] | commensal bacterium initial colonizer |
| V. dispar | coccoid | 1.2 × 1.2 | anaerobe | Gram− | fimbriae adhesins [27] | commensal bacterium secondary colonizer |
| P. gingivalis | coccobacillus | 1 × 0.75 | anaerobe | Gram− | fimbriae adhesins [28] | oral pathogen late colonizer |
| A. ac rough | coccobacillus | 1 × 0.75 | facultative anaerobe | Gram− | fimbriae adhesins [29] | oral pathogen late colonizer |
| A. ac smooth | coccobacillus | 1 × 0.75 | facultative anaerobe | Gram− | adhesins [29] | oral pathogen late colonizer |
| Species | Buffer | Maximum Adhesion Force [nN] | Attach. Points | Detach. Distance [µm] | Elasticity [kPa] | Zeta Potential [mV] | Relative Membrane Integrity [%] | Metabolic Activity [Units] |
|---|---|---|---|---|---|---|---|---|
| S. oralis | PBS | 2.19 ± 1.34 | 13 ± 8 | 0.46 ± 0.31 | 38.5 ± 39.9 | −3.6 ± 1.8 | 86.0 ± 11.2 | 551 ± 238 |
| RTF | 0.47 ± 0.51 | 4 ± 3 | 0.10 ± 0.10 | 71.9 ± 86.1 | −6.2 ± 3.2 | 84.5 ± 14.8 | 1705 ± 502 | |
| A. naeslundii | PBS | 0.56 ± 0.31 | 12 ± 6 | 0.60 ± 0.36 | 24.3 ± 25.4 | −7.2 ± 3.2 | 80.2 ± 13.0 | 3936 ± 885 |
| RTF | 0.40 ± 0.28 | 10 ± 4 | 0.66 ± 0.30 | 12.9 ± 18.9 | −10.8 ± 2.3 | 84.5 ± 14.7 | 4102 ± 1121 | |
| V. dispar | PBS | 0.28 ± 0.20 | 3 ± 2 | 0.08 ± 0.07 | 178.2 ± 234.3 | −8.4 ± 2.3 | 84.5 ± 4.2 | 61 ± 11 |
| RTF | 0.18 ± 0.12 | 5 ± 4 | 0.26 ± 0.19 | 50.4 ± 42.4 | −13.3 ± 0.8 | 82.3 ± 3.8 | 165 ± 18 | |
| P. gingivalis | PBS | 0.45 ± 0.33 | 7 ± 3 | 0.21 ± 0.12 | 31.9 ± 27.3 | −6.2 ± 2.0 | 85.3 ± 11.8 | 1404 ± 394 |
| RTF | 0.51 ± 0.44 | 7 ± 6 | 0.34 ± 0.30 | 41.8 ± 29.6 | −20.0 ± 1.6 | 92.7 ± 3.4 | 2052 ± 1303 | |
| A. ac rough | PBS | 0.37 ± 0.23 | 6 ± 4 | 0.26 ± 0.16 | 68.3 ± 64.6 | −2.6 ± 3.1 | 54.0 ± 19.5 | 5632 ± 4735 |
| RTF | 0.24 ± 0.30 | 3 ± 2 | 0.18 ± 0.17 | 37.3 ± 52.3 | −2.4 ± 1.2 | 60.4 ± 27.1 | 2996 ± 1178 | |
| A. ac smooth | PBS | 0.37 ± 0.23 | 6 ± 2 | 0.32 ± 0.17 | 41.0 ± 47.7 | −8.9 ± 1.8 | 35.7 ± 24.2 | 496 ± 44 |
| RTF | 0.28 ± 0.13 | 5 ± 2 | 0.26 ± 0.16 | 26.8 ± 16.7 | −16.7 ± 1.3 | 51.2 ± 25.6 | 380 ± 31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doll-Nikutta, K.; Winkel, A.; Yang, I.; Grote, A.J.; Meier, N.; Habib, M.; Menzel, H.; Behrens, P.; Stiesch, M. Adhesion Forces of Oral Bacteria to Titanium and the Correlation with Biophysical Cellular Characteristics. Bioengineering 2022, 9, 567. https://doi.org/10.3390/bioengineering9100567
Doll-Nikutta K, Winkel A, Yang I, Grote AJ, Meier N, Habib M, Menzel H, Behrens P, Stiesch M. Adhesion Forces of Oral Bacteria to Titanium and the Correlation with Biophysical Cellular Characteristics. Bioengineering. 2022; 9(10):567. https://doi.org/10.3390/bioengineering9100567
Chicago/Turabian StyleDoll-Nikutta, Katharina, Andreas Winkel, Ines Yang, Anna Josefine Grote, Nils Meier, Mosaieb Habib, Henning Menzel, Peter Behrens, and Meike Stiesch. 2022. "Adhesion Forces of Oral Bacteria to Titanium and the Correlation with Biophysical Cellular Characteristics" Bioengineering 9, no. 10: 567. https://doi.org/10.3390/bioengineering9100567