
Biofabrication of Artificial Stem Cell Niches in the Anterior Ocular Segment

 and
and
Abstract
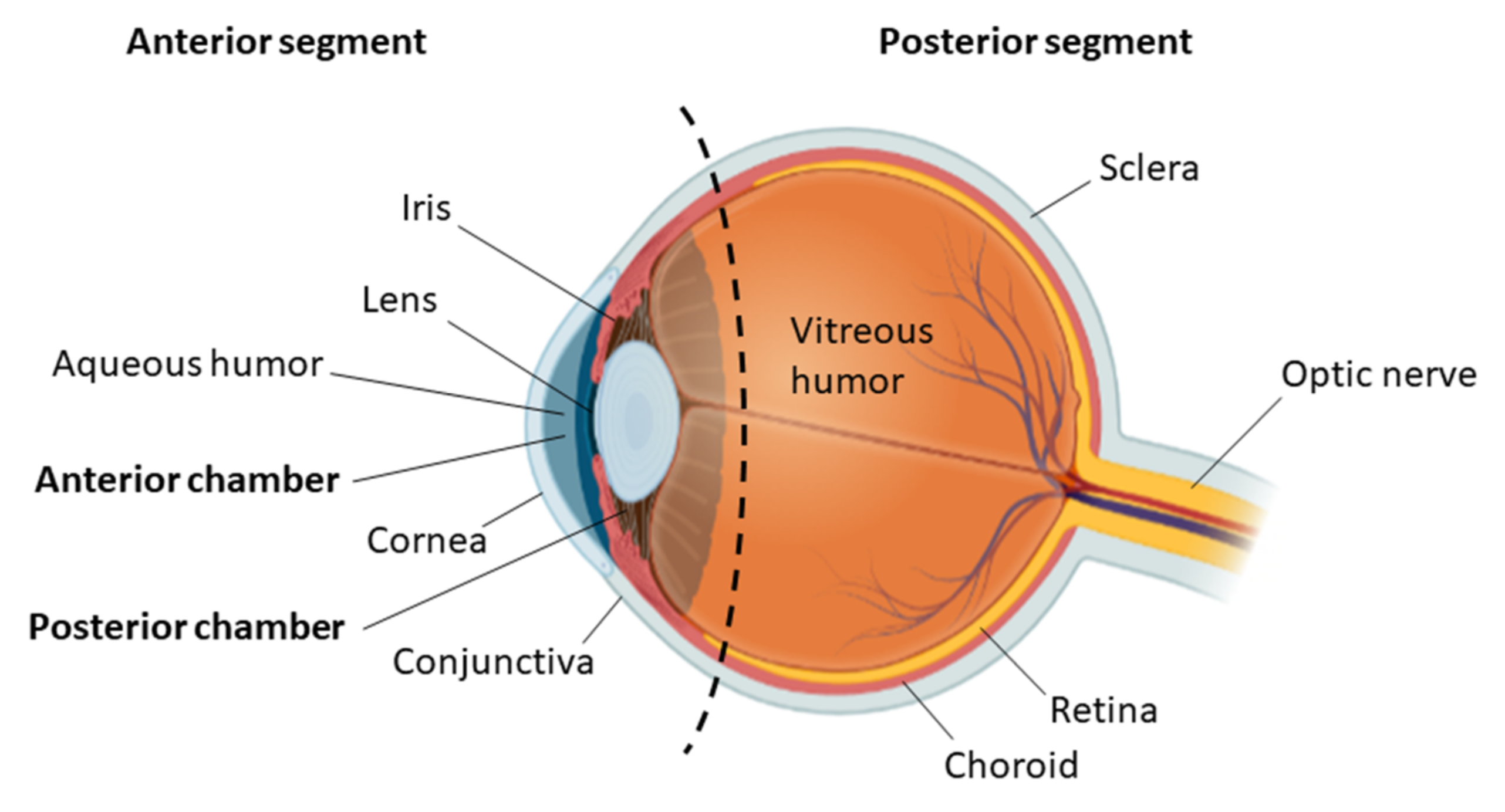
:1. Introduction
2. Ocular Disease and Stem Cell Therapy
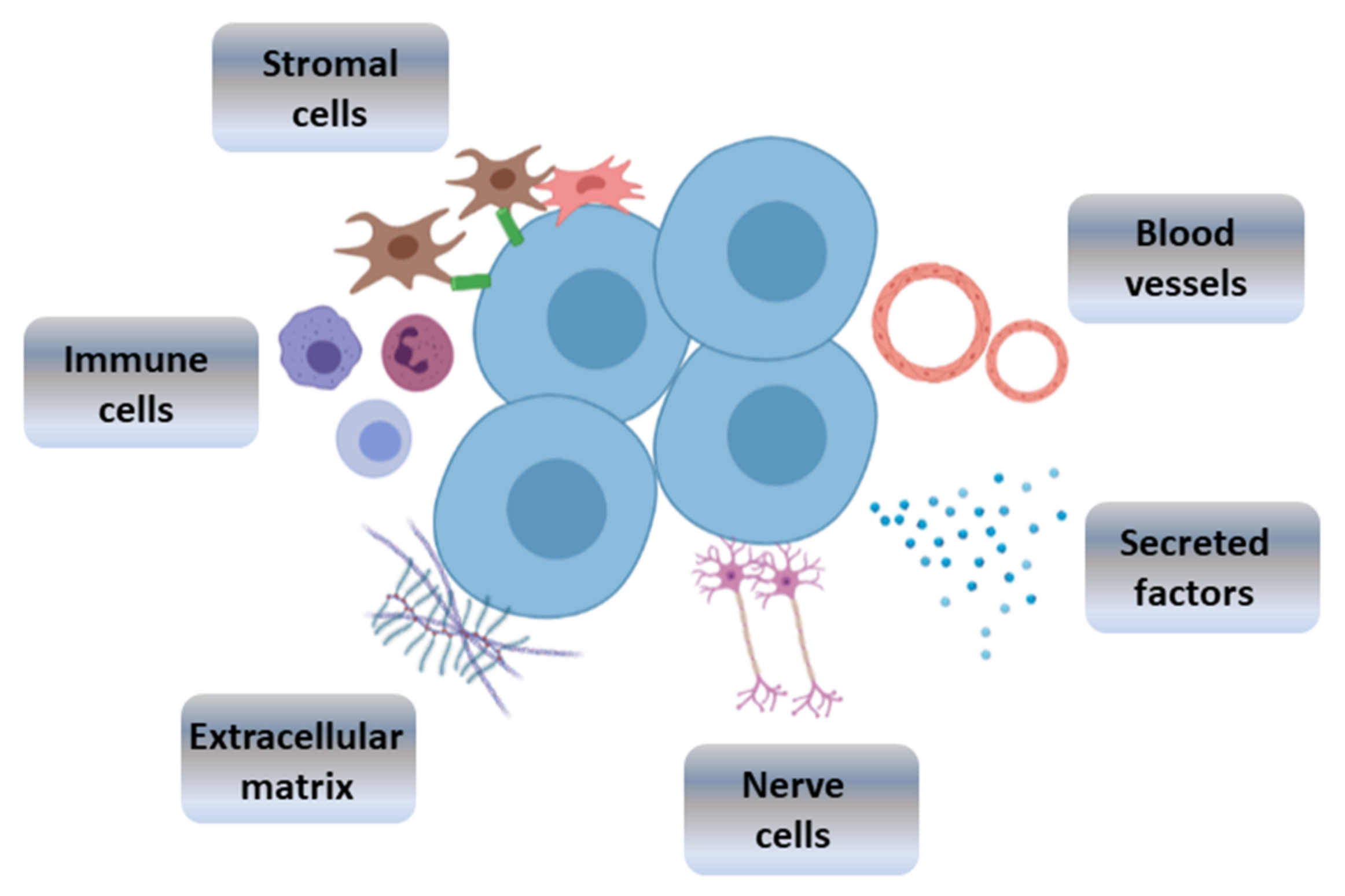
3. Bioengineered Stem Cell Niches
3.1. Photolithography
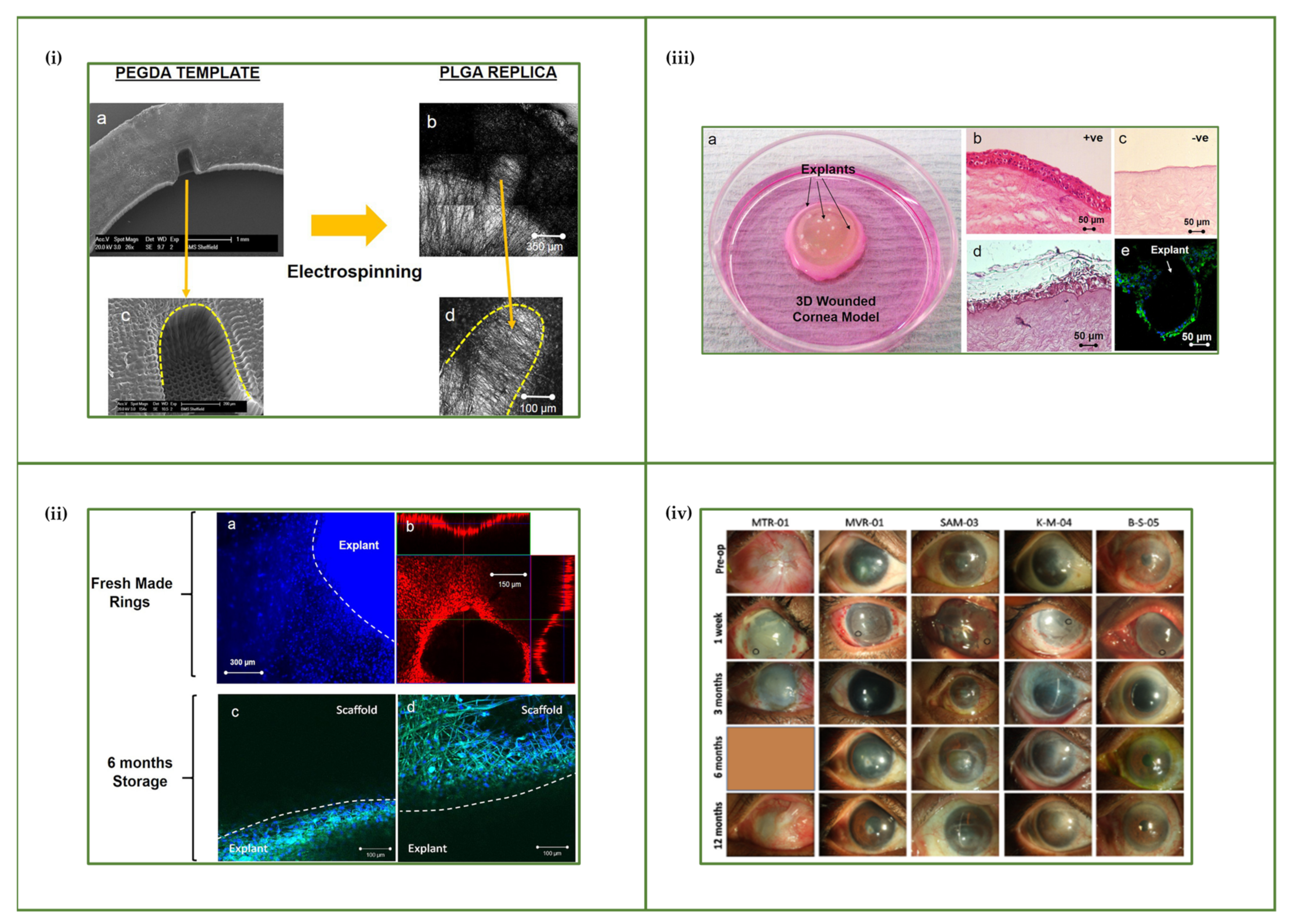
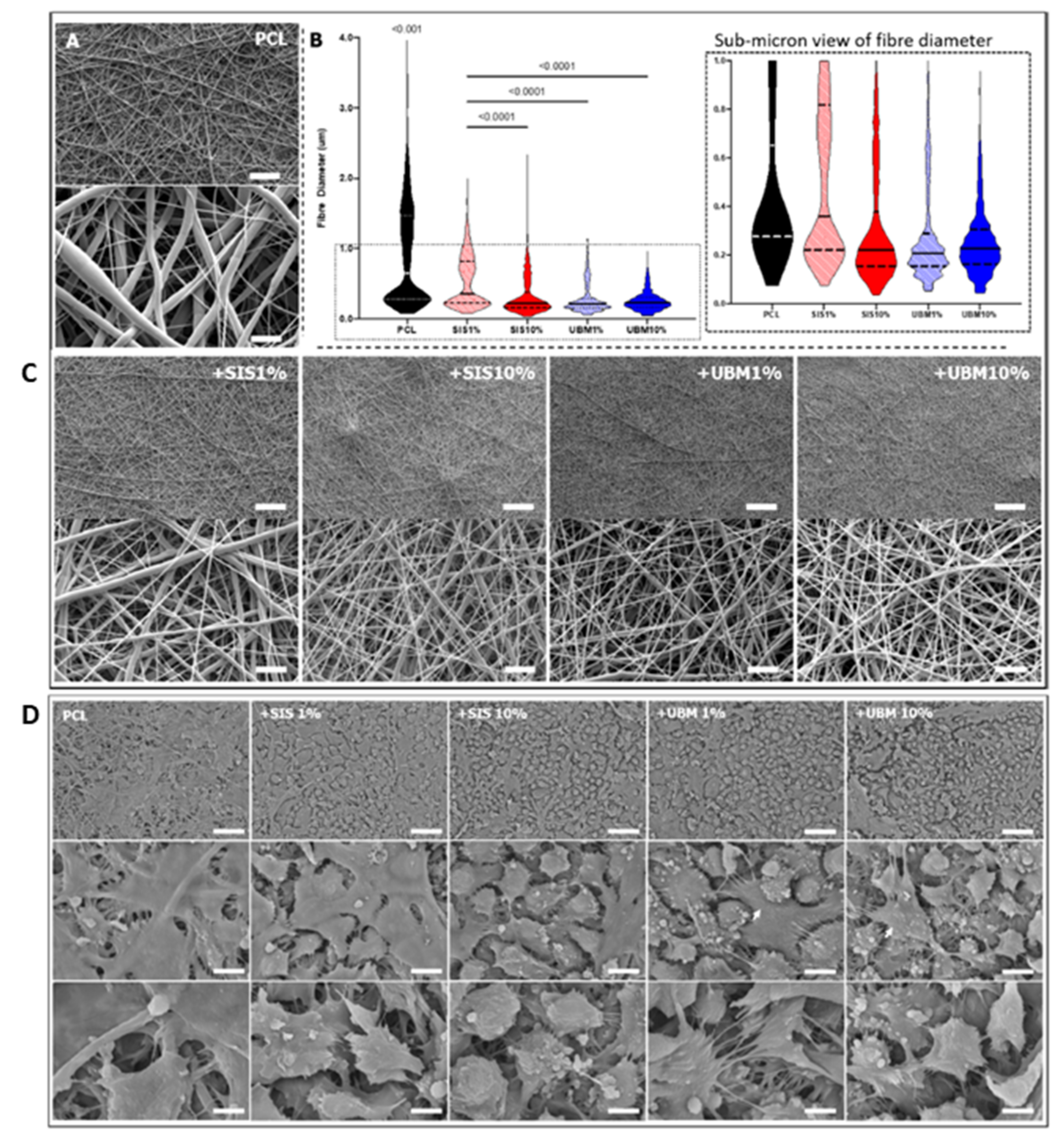
3.2. Electrospinning
3.3. 3D Printing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technique | Principle | Advantages | Disadvantages | Ref. |
|---|---|---|---|---|
| 3D printing/ bioprinting | Production of 3D structures by depositing materials layer by layer | Production of hydrated polymeric structures; production of custom-built structures; easy integration in the clinic; improved mimicry of anatomical structures; higher resolution and reproducibility | High cost of some types of 3DP equipment; it can take a long time, which is very detrimental when producing large constructs; lower dimensional accuracy with some 3DP techniques; difficulties encountered in the development of printable biomaterials | [54,61,62] |
| Electrospinning | Production of nanofibers by applying a high voltage to a polymer solution as it passes through a needle | High-scale production; good mechanical properties; Possible surface modification; large surface-volume ratio; nanotopographical control of cells; high versatility in materials (polymers, metals, ceramic) | Difficult to produce scaffolds with a high volume; non-uniform cellular distribution and poor cell infiltration; limited to polymers; inferior macroscopic mechanical properties in comparison with other techniques; possible organic solvent residue | [54,55,63,64] |
| Photolithography | Use of light to transfer a geometric pattern to a photosensitive material by using a photomask | Uniform cell encapsulation; reduced heat production; controllable reaction kinetics | Potential toxicity of the photoinitiator molecules present in photocurable resins | [53] |
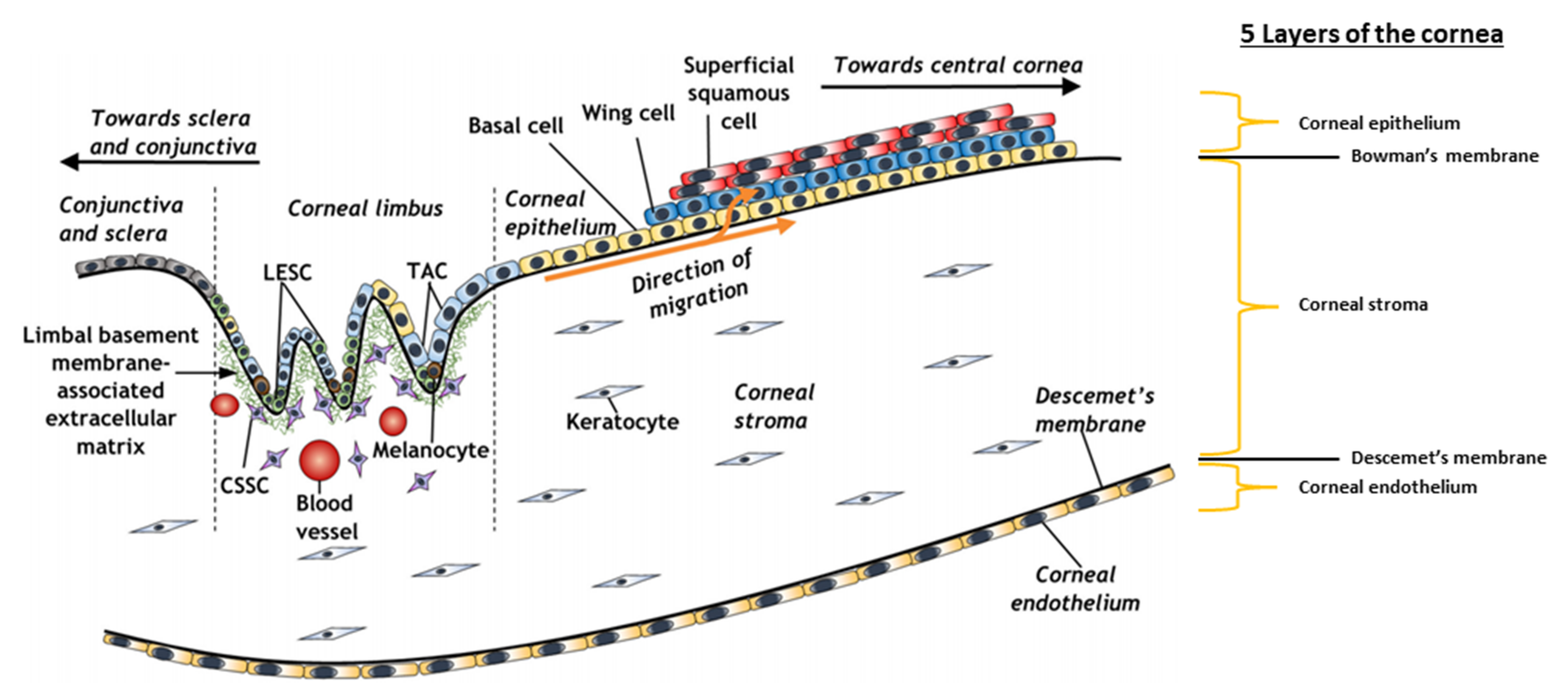
4. Cornea
| Cornea Layer | Function | Composition | Cell layers | Types of Cells | Thickness (µm) | Regenerates | Ref. |
|---|---|---|---|---|---|---|---|
| Epithelium |
| Stratified squamous epithelium | 5 to 7 |
| 50 | Yes (every 7 to 10 days) | [71,72,73,74] |
| Bowman’s Layer |
| Compact layer of unorganised collagen fibres | Monolayer | Acellular (collagens (Type I, V), proteoglycans) | 12 | No | [71,74,75,76] |
| Stroma |
| Orderly arrangement of collagen lamellae with keratocytes | 200–250 distinct lamellae |
| 500 | Yes (slow process over several years) | [71,73,74,76] |
| Descemet’s Membrane |
| Basement membrane materials | Monolayer | Acellular (collagen (Type IV), laminin) | 3–10 | Yes | [71,74,75,76] |
| Endothelium |
| Single layer of simple squamous epithelium | Monolayer | Cellular | 5 | No | [73,77,78] |
4.1. The Corneal Stem Cell Niche

4.2. Bioengineering the Corneal Stem Cell Niche
4.2.1. Corneal Epithelium
4.2.2. Stroma
4.2.3. Endothelium
4.2.4. Corneal Innervation
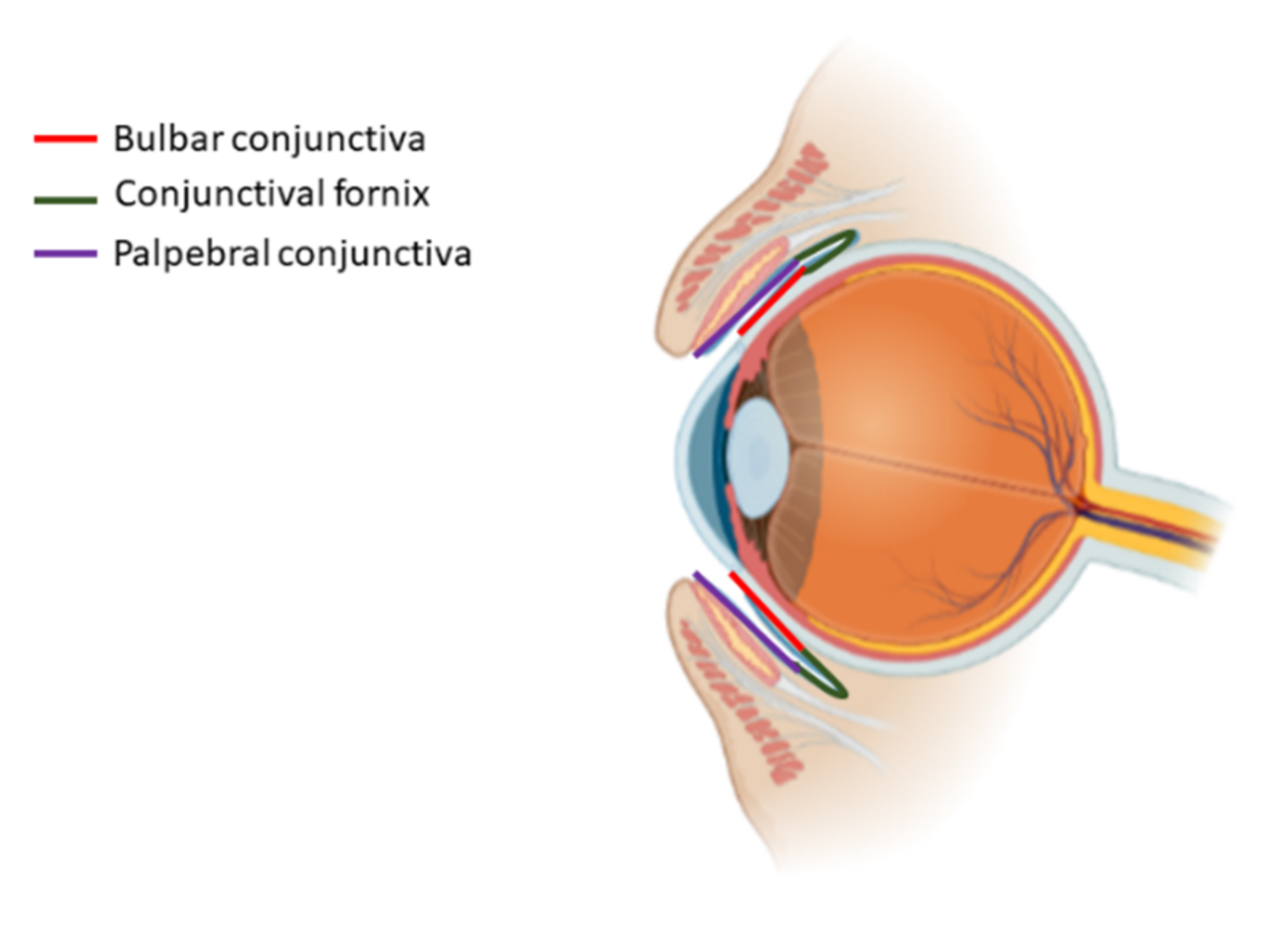
5. Conjunctiva
5.1. The Conjunctival Stem Cell Niche
5.2. Bioengineering the Conjunctival Stem Cell Niche
6. Iris
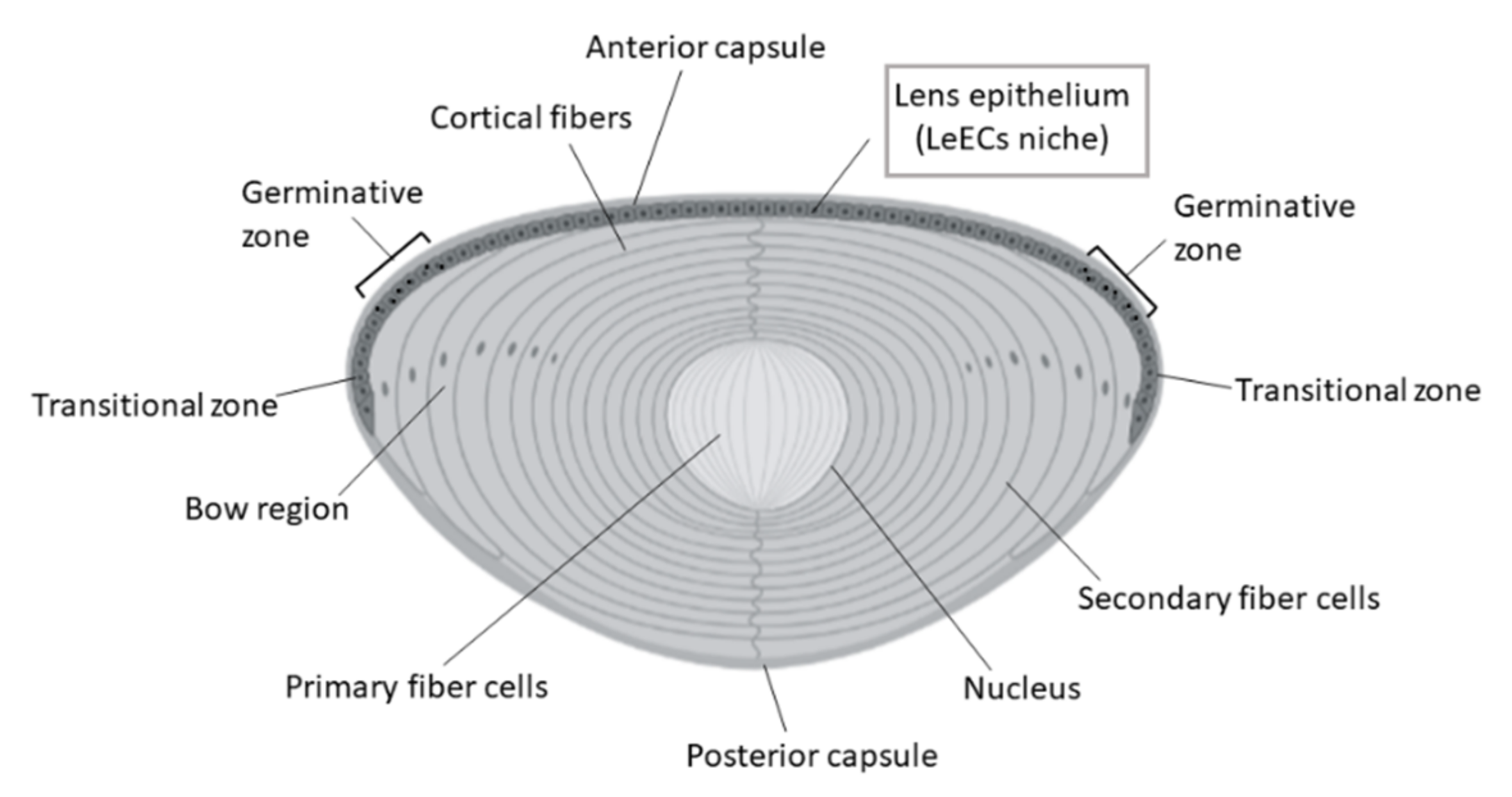
7. Lens
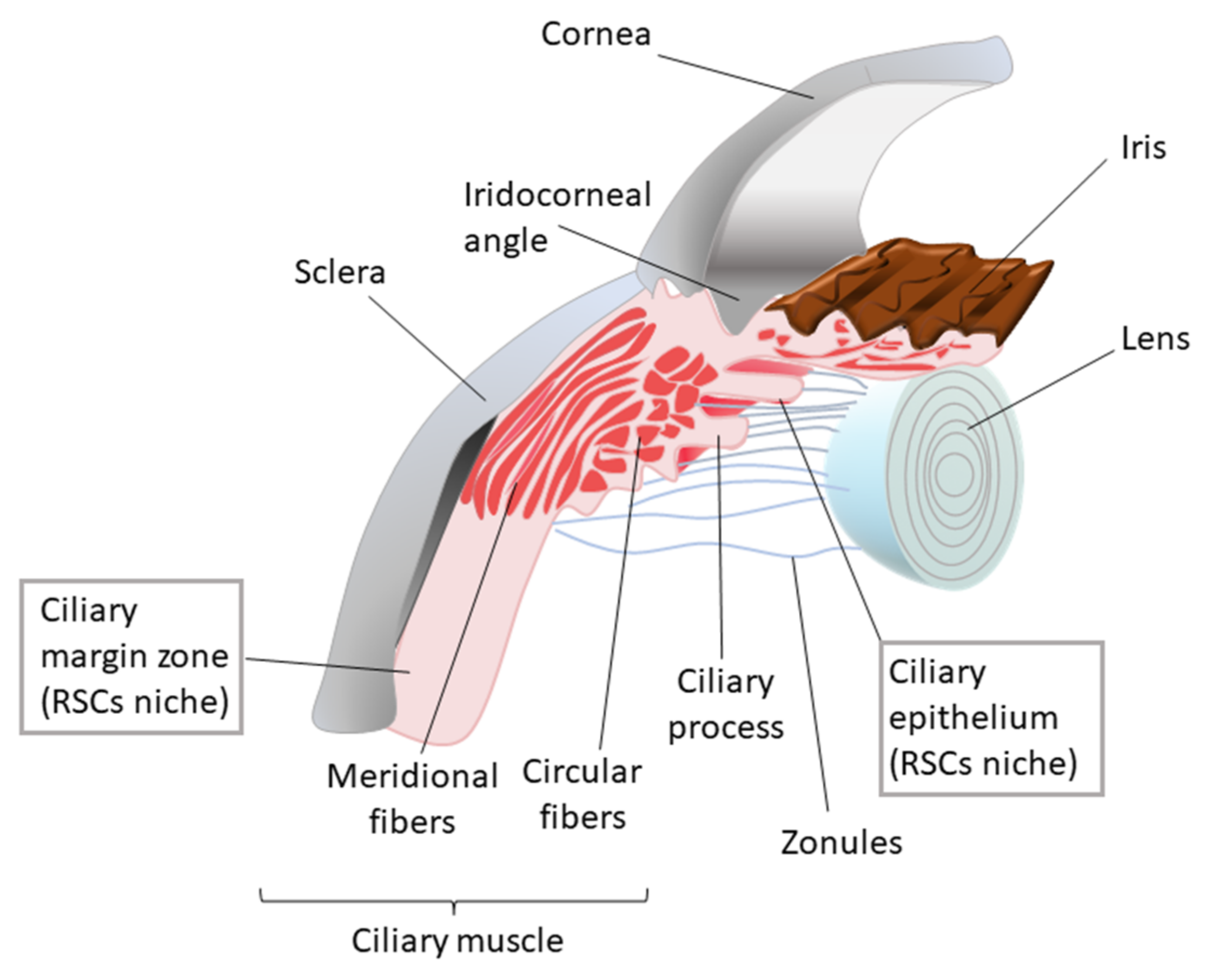
8. Ciliary Body

8.1. The Ciliary Body Stem Cell Niche
8.2. Bioengineering the Ciliary Body Stem Cell Niche
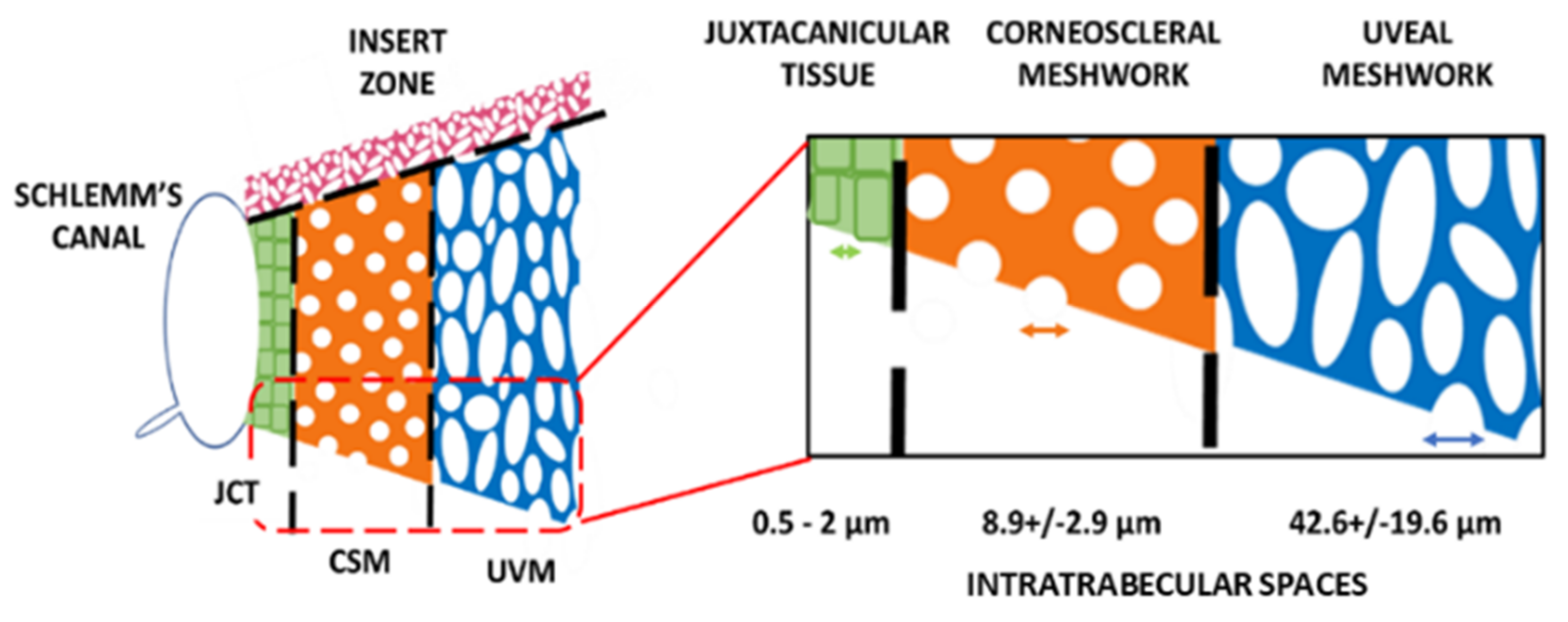
9. Trabecular Meshwork
9.1. The Trabecular Meshwork Stem Cell Niche
9.2. Bioengineering the Trabecular Meshwork Stem Cell Niche
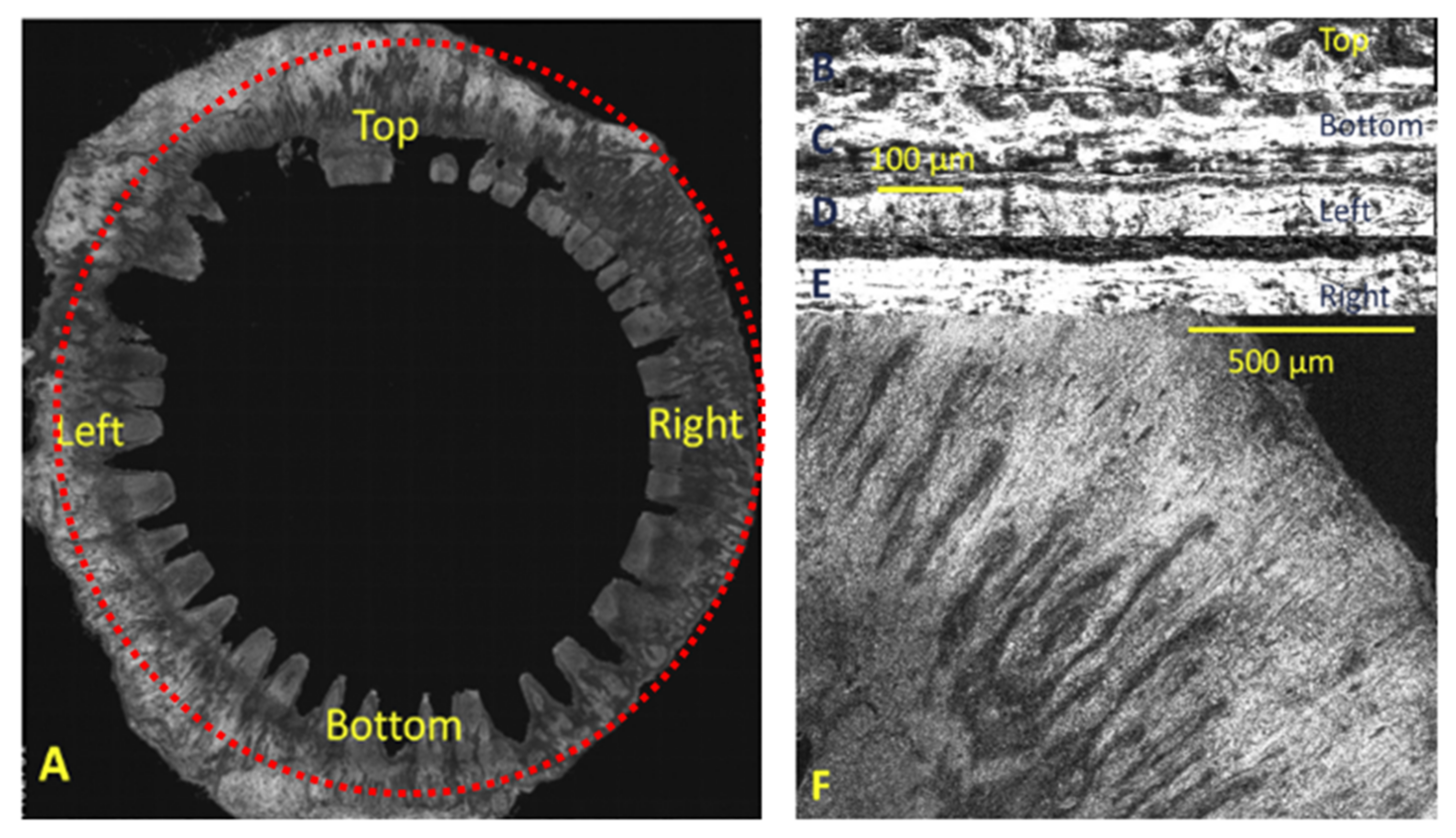
10. Sclera
11. Future Perspectives and Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naisberg, Y. Biophysical vision model and learning paradigms about vision: Review. Med. Hypotheses 2001, 57, 409–418. [Google Scholar] [CrossRef]
- Palczewski, K. Chemistry and biology of vision. J. Biol. Chem. 2012, 287, 1612–1619. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, B.J.; Jenkins, M.W.; Rollins, A.M.; Dupps, W.J. A review of structural and biomechanical changes in the cornea in aging, disease, and photochemical crosslinking. Front. Bioeng. Biotechnol. 2019, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Kels, B.D.; Grzybowski, A.; Grant-Kels, J.M. Human ocular anatomy. Clin. Dermatol. 2015, 33, 140–146. [Google Scholar] [CrossRef]
- Hampson, K.M. Adaptive optics and vision. J. Mod. Opt. 2008, 55, 3425–3467. [Google Scholar] [CrossRef]
- Banh, A.; Bantseev, V.; Choh, V.; Moran, K.L.; Sivak, J.G. The lens of the eye as a focusing device and its response to stress. Prog. Retin. Eye Res. 2006, 25, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Deeb, S.S.; Motulsky, A.G. Molecular Genetics of Human Colour Vision. Behav. Genet. 1996, 26, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Hasrod, N.; Rubin, A. Colour vision: A review of the Cambridge Colour Test and other colour testing methods. Afr. Vis. Eye Health 2015, 74, 7. [Google Scholar] [CrossRef]
- Sung, C.H.; Chuang, J.Z. The cell biology of vision. J. Cell Biol. 2010, 190, 953–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Inatomi, T.; Sotozono, C.; Koizumi, N.; Kinoshita, S. Ocular surface reconstruction using stem cell and tissue engineering. Prog. Retin. Eye Res. 2016, 51, 187–207. [Google Scholar] [CrossRef]
- Khoshzaban, A.; Jafari, E.; Soleimani, M.; Sa, T.; Nekoozadeh, S.; Aaz, E. Introduction of Stem Cells in Ophthalmology. J. Stem Cells Clin. Pract. 2018, 1, 1–8. [Google Scholar]
- Aharony, I.; Michowiz, S.; Goldenberg-Cohen, N. The promise of stem cell-based therapeutics in ophthalmology. Neural Regen. Res. 2017, 12, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Ota, K.I. Stem Cells: Past, Present and Future. IEEE J. Trans. Fundam. Mater. 2008, 128, 329–332. [Google Scholar] [CrossRef]
- Kolios, G.; Moodley, Y. Introduction to stem cells and regenerative medicine. Respiration 2012, 85, 3–10. [Google Scholar] [CrossRef]
- Wagers, A.J.; Weissman, I.L. Plasticity of adult stem cells. Cell 2004, 116, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, F.; Lo Celso, C.; Scadden, D. Adult stem cells and their niches. Adv. Exp. Med. Biol. 2010, 695, 155–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhamodaran, K.; Subramani, M.; Ponnalagu, M.; Shetty, R.; Das, D. Ocular stem cells: A status update! Stem Cell Res. Ther. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Kolli, S.; Lako, M.; Figueiredo, F.; Daniels, J.T. Stem cell therapies for ocular surface disease. Drug Discov. Today 2010, 15, 306–313. [Google Scholar] [CrossRef]
- Peerani, R.; Zandstra, P.W. Enabling stem cell therapies through synthetic stem cell-niche engineering. J. Clin. Investig. 2010, 120, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Barker, N. Engineering the niche for stem cells. Growth Factors 2013, 31, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Lei, L.; Shi, S.; Li, X. Stimulus-Responsive Hydrogel for Ophthalmic Drug Delivery. Macromol. Biosci. 2019, 19, e1900001. [Google Scholar] [CrossRef]
- Sivan, P.P.; Syed, S.; Mok, P.L.; Higuchi, A.; Murugan, K.; Alarfaj, A.A.; Munusamy, M.A.; Hamat, R.A.; Umezawa, A.; Kumar, S. Stem Cell Therapy for Treatment of Ocular Disorders. Stem Cells Int. 2016, 2016, 8304879. [Google Scholar] [CrossRef] [PubMed]
- Bobba, S.; Di Girolamo, N.; Munsie, M.; Chen, F.; Pébay, A.; Harkin, D.; Hewitt, A.; O’Connor, M.; McLenachan, S.; Shadforth, A.M.A.; et al. The current state of stem cell therapy for ocular disease. Exp. Eye Res. 2018, 177, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, H.Y.; Liang, H.L.; Wang, Y.S.; Zhang, Z.X.; Wang, B.R.; Shi, Y.Y.; Dong, X.; Cai, Y. A therapeutic strategy for choroidal neovascularization based on recruitment of mesenchymal stem cells to the sites of lesions. Mol. Ther. 2010, 18, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- Stahl, A.; Smith, L.E.H. An eye for discovery. J. Clin. Investig. 2010, 120, 3008–3011. [Google Scholar] [CrossRef]
- Blenkinsop, T.A.; Corneo, B.; Temple, S.; Stern, J.H. Ophthalmologic stem cell transplantation therapies. Regen. Med. 2012, 7, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Boulton, M.; Albon, J. Stem cells in the eye. Int. J. Biochem. Cell Biol. 2004, 36, 643–657. [Google Scholar] [CrossRef]
- Javid, D.; Zhang, J.; Alamdari, D.H. Hot Topics in Limbal Stem Cells Culture: Mini Review. J. Ophthalmol. Sci. 2021, 3, 1–5. [Google Scholar]
- Atallah, M.R.; Palioura, S.; Perez, V.L.; Amescua, G. Limbal stem cell transplantation: Current perspectives. Clin. Ophthalmol. 2016, 10, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Hayashida, Y.; Chen, Y.-T.; Tseng, S.C. Niche regulation of corneal epithelial stem cells at the limbus. Cell Res. 2007, 17, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Dziasko, M.A.; Daniels, J.T. Anatomical Features and Cell-Cell Interactions in the Human Limbal Epithelial Stem Cell Niche. Ocul. Surf. 2016, 14, 322–330. [Google Scholar] [CrossRef]
- Pinnamaneni, N.; Funderburgh, J.L. Concise review: Stem cells in the corneal stroma. Stem Cells 2012, 30, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Funderburgh, M.L.; Mann, M.M.; SundarRaj, N.; Funderburgh, J.L. Multipotent stem cells in human corneal stroma. Stem Cells 2005, 23, 1266–1275. [Google Scholar] [CrossRef] [Green Version]
- Weng, L.; Funderburgh, J.L.; Khandaker, I.; Geary, M.L.; Yang, T.; Basu, R.; Funderburgh, M.L.; Du, Y.; Yam, G.H. The anti-scarring effect of corneal stromal stem cell therapy is mediated by transforming growth factor β3. Eye Vis. 2020, 7, 52. [Google Scholar] [CrossRef]
- Stewart, R.M.K.; Sheridan, C.M.; Hiscott, P.S.; Czanner, G.; Kaye, S.B. Human Conjunctival Stem Cells are Predominantly Located in the Medial Canthal and Inferior Forniceal Areas. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2021–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gipson, I.K. Goblet cells of the conjunctiva: A review of recent findings. Prog. Retin. Eye Res. 2016, 54, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, H.; Zhou, Y.; Wills, A.; Du, Y. Stem cells in the trabecular meshwork for regulating intraocular pressure. J. Ocul. Pharmacol. Ther. 2016, 32, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.Y.; Sheridan, C.; Grierson, I.; Mason, S.; Kearns, V.; Lo, A.C.Y.; Wong, D. Progenitors for the corneal endothelium and trabecular meshwork: A potential source for personalized stem cell therapy in corneal endothelial diseases and glaucoma. J. Biomed. Biotechnol. 2011, 2011, 412743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Ouyang, H.; Zhu, J.; Huang, S.; Liu, Z.; Chen, S.; Cao, G.; Li, G.; Signer, R.A.J.; Xu, Y.; et al. Lens regeneration using endogenous stem cells with gain of visual function. Nature 2016, 531, 323–328. [Google Scholar] [CrossRef]
- Saxby, L.; Rosen, E.; Boulton, M. Lens epithelial cell proliferation, migration, and metaplasia following capsulorhexis. Br. J. Ophthalmol. 1998, 82, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Mao, Y.; Bouaziz, M.; Yu, H.; Qu, X.; Wang, F.; Feng, G.S.; Shawber, C.; Zhang, X. Lens differentiation is controlled by the balance between PDGF and FGF signaling. PLoS Biol. 2019, 17, e3000133. [Google Scholar] [CrossRef] [Green Version]
- Asami, M.; Sun, G.; Yamaguchi, M.; Kosaka, M. Multipotent cells from mammalian iris pigment epithelium. Dev. Biol. 2007, 304, 433–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Asami, M.; Ohta, H.; Kosaka, J.; Kosaka, M. Retinal stem/progenitor properties of iris pigment epithelial cells. Dev. Biol. 2006, 289, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, T.; Kagawa, T.; Fukushima, M.; Shimizu, T.; Yoshinaga, Y.; Takada, S.; Tanihara, H.; Taga, T. Activation of Canonical Wnt Pathway Promotes Proliferation of Retinal Stem Cells Derived from Adult Mouse Ciliary Margin. Stem Cells 2006, 24, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Kubo, F.; Takeichi, M.; Nakagawa, S. Wnt2b controls retinal cell differentiation at the ciliary marginal zone. Development 2003, 130, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Song, W.T.; Zhang, X.Y.; Xia, X.B. Atoh7 promotes the differentiation of retinal stem cells derived from Müller cells into retinal ganglion cells by inhibiting Notch signaling. Stem Cell Res. Ther. 2013, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.J.; Spradling, A.C. Stem Cells and Niches: Mechanisms That Promote Stem Cell Maintenance throughout Life. Cell 2008, 132, 598–611. [Google Scholar] [CrossRef] [Green Version]
- Mitsiadis, T.A.; Barrandon, O.; Rochat, A.; Barrandon, Y.; De Bari, C. Stem cell niches in mammals. Exp. Cell Res. 2007, 313, 3377–3385. [Google Scholar] [CrossRef]
- Rezza, A.; Sennett, R.; Rendl, M. Adult stem cell niches: Cellular and molecular components. Curr. Top. Dev. Biol. 2014, 107, 333–372. [Google Scholar] [CrossRef]
- Raymond, K.; Deugnier, M.A.; Faraldo, M.M.; Glukhova, M.A. Adhesion within the stem cell niches. Curr. Opin. Cell Biol. 2009, 21, 623–629. [Google Scholar] [CrossRef]
- Donnelly, H.; Salmeron-Sanchez, M.; Dalby, M.J. Designing stem cell niches for differentiation and self-renewal. J. R. Soc. Interface 2018, 15, 20180388. [Google Scholar] [CrossRef]
- Pennings, S.; Liu, K.J.; Qian, H. The Stem Cell Niche: Interactions between Stem Cells and Their Environment. Stem Cells Int. 2018, 2018, 4879379. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, P.; Schweller, R.M.; Khademhosseini, A.; West, J.L.; Bashir, R. 3D biofabrication strategies for tissue engineering and regenerative medicine. Annu. Rev. Biomed. Eng. 2014, 16, 247–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.S.; Lee, S.H.; Park, W.J.; Lee, J.E.; Kim, B.; Han, D.W. Advanced techniques for skeletal muscle tissue engineering and regeneration. Bioengineering 2020, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Jeong, S.I.; Bae, M.S.; Yang, D.H.; Heo, D.N.; Kim, C.H.; Alsberg, E.; Kwon, I.K. Highly porous electrospun nanofibers enhanced by ultrasonication for improved cellular infiltration. Tissue Eng. Part. A 2011, 17, 2695–2702. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.; Huang, Y.; Agarwal, S.; Lannutti, J. Improved cellular infiltration in electrospun fiber via engineered porosity. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Pham, Q.P.; Sharma, U.; Mikos, A.G. Electrospun Poly(ε-caprolactone) Microfiber and Multilayer Nanofiber/Microfiber Scaffolds: Characterization of Scaffolds and Measurement of Cellular Infiltration. Biomacromolecules 2006, 7, 2796–2805. [Google Scholar] [CrossRef]
- Prasad, L.K.; Smyth, H. 3D Printing technologies for drug delivery: A review. Drug Dev. Ind. Pharm. 2016, 42, 1019–1031. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Z.; Li, J.; Liu, C.; Lao, C.; Fu, Y.; Liu, C.; Li, Y.; Wang, P.; He, Y. 3D printing of ceramics: A review. J. Eur. Ceram. Soc. 2019, 39, 661–687. [Google Scholar] [CrossRef]
- Serpooshan, V.; Mahmoudi, M.; Hu, D.A.; Hu, J.B.; Wu, S.M. Bioengineering cardiac constructs using 3D printing. J. 3d Print. Med. 2017, 1, 123–139. [Google Scholar] [CrossRef]
- Ali, M.P.R.; Anil, K.; Lee, S.J.; Jackson, J.D. Three-dimensional bioprinting for organ bioengineering: Promise and pitfalls. Curr. Opin. Organ. Transplant. 2018, 23, 649–656. [Google Scholar] [CrossRef]
- Martelli, N.; Serrano, C.; Van Den Brink, H.; Pineau, J.; Prognon, P.; Borget, I.; El Batti, S. Advantages and disadvantages of 3-dimensional printing in surgery: A systematic review. Surgery 2016, 159, 1485–1500. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Kundu, S.C. Electrospinning: A fascinating fiber fabrication technique. Biotechnol. Adv. 2010, 28, 325–347. [Google Scholar] [CrossRef]
- Khorshidi, S.; Solouk, A.; Mirzadeh, H.; Mazinani, S.; Lagaron, J.M.; Sharifi, S.; Ramakrishna, S. A review of key challenges of electrospun scaffolds for tissue-engineering applications. J. Tissue Eng. Regen. Med. 2016, 10, 715–738. [Google Scholar] [CrossRef]
- Meek, K.M.; Knupp, C. Corneal structure and transparency. Prog. Retin. Eye Res. 2015, 49, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ahearne, M.; Fernández-Pérez, J.; Masterton, S.; Madden, P.W.; Bhattacharjee, P. Designing Scaffolds for Corneal Regeneration. Adv. Funct. Mater. 2020, 30, 1908996. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Chow, J.; Liu, J. Corneal Innervation and Sensation: The Eye and Beyond. Yale J. Biol. Med. 2018, 91, 13–21. [Google Scholar] [PubMed]
- Chen, Z.; You, J.; Liu, X.; Cooper, S.; Hodge, C.; Sutton, G.; Crook, J.M.; Wallace, G.G. Biomaterials for corneal bioengineering. Biomed. Mater. 2018, 13, 032002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Stewart, R.; Yung, S.; Kolli, S.; Armstrong, L.; Stojkovic, M.; Figueiredo, F.; Lako, M. Differentiation of Human Embryonic Stem Cells into Corneal Epithelial-Like Cells by In Vitro Replication of the Corneal Epithelial Stem Cell Niche. Stem Cells 2007, 25, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, R.M.; Lepert, G.; Gupta, S.; Mohan, R.R.; Paterson, C.; Connon, C.J. Assessment of corneal substrate biomechanics and its effect on epithelial stem cell maintenance and differentiation. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Notara, M.; Lentzsch, A.; Coroneo, M.; Cursiefen, C. The role of limbal epithelial stem cells in regulating corneal (Lymph)angiogenic Privilege and the micromilieu of the limbal niche following UV Exposure. Stem. Cells Int. 2018, 2018, 8620172. [Google Scholar] [CrossRef] [Green Version]
- Willoughby, C.E.; Ponzin, D.; Ferrari, S.; Lobo, A.; Landau, K.; Omidi, Y. Anatomy and physiology of the human eye: Effects of mucopolysaccharidoses disease on structure and function—A review. Clin. Exp. Ophthalmol. 2010, 38, 2–11. [Google Scholar] [CrossRef]
- McCabe, K.L.; Lanza, R. Corneal Replacement Tissue. Princ. Tissue Eng. Fourth Ed. 2013, 1413–1425. [Google Scholar] [CrossRef]
- Sridhar, M.S. Anatomy of cornea and ocular surface. Indian J. Ophthalmol. 2018, 66, 190–194. [Google Scholar] [CrossRef]
- Lagali, N.; Germundsson, J.; Fagerholm, P.; Germundsson, J.; Fagerholm, P. The role of Bowman’s layer in corneal regeneration after phototherapeutic keratectomy: A prospective study using in vivo confocal microscopy. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4192–4198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoubay, D.; Borderie, M.; Grieve, K.; Martos, R.; Bocheux, R.; Nguyen, T.-M.; Callard, P.; Chédotal, A.; Borderie, V.M. Corneal stromal stem cells restore transparency after N2 injury in mice. Stem Cells Transl. Med. 2020, 9, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Hatou, S.; Shimmura, S. Review: Corneal endothelial cell derivation methods from ES/iPS cells. Inflamm. Regen. 2019, 39, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalili, M.; Asadi, M.; Kahroba, H.; Soleyman, M.R.; Andre, H.; Alizadeh, E. Corneal endothelium tissue engineering: An evolution of signaling molecules, cells, and scaffolds toward 3D bioprinting and cell sheets. J. Cell. Physiol. 2021, 236, 3275–3303. [Google Scholar] [CrossRef]
- Shortt, A.J.; Tuft, S.J.; Daniels, J.T. Corneal stem cells in the eye clinic. Br. Med. Bull. 2011, 100, 209–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.J.; Ismail, S.; Sherwin, T. Limbal stem cells: Central concepts of corneal epithelial homeostasis. World J. Stem Cells 2014, 6, 391. [Google Scholar] [CrossRef]
- Sherwin, T. Defining the Limbal Stem Cell Niche. J. Cell Signal. 2016, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Notara, M.; Shortt, A.J.; Galatowicz, G.; Calder, V.; Daniels, J.T. IL6 and the human limbal stem cell niche: A mediator of epithelial-stromal interaction. Stem Cell Res. 2010, 5, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Shanmuganathan, V.A.; Foster, T.; Kulkarni, B.B.; Hopkinson, A.; Gray, T.; Powe, D.G.; Lowe, J.; Dua, H.S. Morphological characteristics of the limbal epithelial crypt. Br. J. Ophthalmol. 2007, 91, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziasko, M.A.; Armer, H.E.; Levis, H.J.; Shortt, A.J.; Tuft, S.; Daniels, J.T. Localisation of epithelial cells capable of holoclone formation in vitro and direct interaction with stromal cells in the native human limbal crypt. PLoS ONE 2014, 9, e94283. [Google Scholar] [CrossRef]
- Kureshi, A.K.; Dziasko, M.; Funderburgh, J.L.; Daniels, J.T. Human corneal stromal stem cells support limbal epithelial cells cultured on RAFT tissue equivalents. Sci. Rep. 2015, 5, 16186. [Google Scholar] [CrossRef]
- Duan, C.Y.; Xie, H.T.; Zhao, X.Y.; Zhang, M.C. Limbal niche cells: A novel feeder cell for autologous cultivated oral mucosal epithelial transplantation. Regen. Med. 2019, 14, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Mariappan, I.; Maddileti, S.; Savy, S.; Tiwari, S.; Gaddipati, S.; Fatima, A.; Sangwan, V.S.; Balasubramanian, D.; Vemuganti, G.K. In vitro culture and expansion of human limbal epithelial cells. Nat. Protoc. 2010, 5, 1470–1479. [Google Scholar] [CrossRef]
- Barui, A.; Chowdhury, F.; Pandit, A.; Datta, P. Rerouting mesenchymal stem cell trajectory towards epithelial lineage by engineering cellular niche. Biomaterials 2018, 156, 28–44. [Google Scholar] [CrossRef]
- Kafarnik, C.; McClellan, A.; Dziasko, M.; Daniels, J.T.; Guest, D.J. Canine Corneal Stromal Cells Have Multipotent Mesenchymal Stromal Cell Properties in Vitro. Stem Cells Dev. 2020, 29, 425–439. [Google Scholar] [CrossRef]
- Feizi, S. Corneal endothelial cell dysfunction: Etiologies and management. Ther. Adv. Ophthalmol. 2018, 10, 251584141881580. [Google Scholar] [CrossRef] [Green Version]
- Van den Bogerd, B.; Dhubhghaill, S.N.; Koppen, C.; Tassignon, M.J.; Zakaria, N. A review of the evidence for in vivo corneal endothelial regeneration. Surv. Ophthalmol. 2018, 63, 149–165. [Google Scholar] [CrossRef] [Green Version]
- Fuest, M.; Hin-Fai Yam, G.; Swee-Lim Peh, G.; Mehta, J.S. Advances in corneal cell therapy. Regen. Med. 2016, 11, 601–615. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Zeng, X.; Hu, J.; Chen, Q. Characterization of Nestin, a Selective Marker for Bone Marrow Derived Mesenchymal Stem Cells. Stem Cells Int. 2015, 2015, 762098. [Google Scholar] [CrossRef] [Green Version]
- Takács, L.; Tóth, E.; Berta, A.; Vereb, G. Stem cells of the adult cornea: From cytometric markers to therapeutic applications. Cytom. Part A 2009, 75, 54–66. [Google Scholar] [CrossRef]
- Ming Sie, N.; Hin-Fai Yam, G.; Qiang Soh, Y.; Lovatt, M.; Dhaliwal, D.; Kocaba, V.; Mehta, J.S. Regenerative capacity of the corneal transition zone for endothelial cell therapy. Stem Cell Res. Ther. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Abib, F.; Hida, R.; Martins, R.; Santos, D. Corneal Endothelium: Histology, Physiology and In-vivo Examination with Specular Microscope. JSM Ophthalmol. 2017, 5, 1063. [Google Scholar]
- Ashworth, S.; Harrington, J.; Hammond, G.M.; Bains, K.K.; Koudouna, E.; Hayes, A.J.; Ralphs, J.R.; Regini, J.W.; Young, R.D.; Hayashi, R.; et al. Chondroitin Sulfate as a Potential Modulator of the Stem Cell Niche in Cornea. Front. Cell Dev. Biol. 2021, 8, 567358. [Google Scholar] [CrossRef] [PubMed]
- Yokoo, S.; Yamagami, S.; Yanagi, Y.; Uchida, S.; Mimura, T.; Usui, T.; Amano, S. Human Corneal Endothelial Cell Precursors Isolated by Sphere-Forming Assay. Invest. Ophthalmol. Vis. Sci. 2005, 46, 1626–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimura, T.; Yamagami, S.; Amano, S. Corneal endothelial regeneration and tissue engineering. Prog. Retin. Eye Res. 2013, 35, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, S.; Yokoo, S.; Mimura, T.; Takato, T.; Araie, M.; Amano, S. Distribution of Precursors in Human Corneal Stromal Cells and Endothelial Cells. Ophthalmology 2007, 114, 433–439. [Google Scholar] [CrossRef]
- Hirata-Tominaga, K.; Nakamura, T.; Okumura, N.; Kawasaki, S.; Kay, E.P.; Barrandon, Y.; Koizumi, N.; Kinoshita, S. Corneal endothelial cell fate is maintained by LGR5 through the regulation of hedgehog and Wnt pathway. Stem Cells 2013, 31, 1396–1407. [Google Scholar] [CrossRef]
- Hara, S.; Hayashi, R.; Soma, T.; Kageyama, T.; Duncan, T.; Tsujikawa, M.; Nishida, K. Identification and potential application of human corneal endothelial progenitor cells. Stem Cells Dev. 2014, 23, 2190–2201. [Google Scholar] [CrossRef]
- Katikireddy, K.R.; Schmedt, T.; Price, M.O.; Price, F.W.; Jurkunas, U.V. Existence of Neural Crest–Derived Progenitor Cells in Normal and Fuchs Endothelial Dystrophy Corneal Endothelium. Am. J. Pathol. 2016, 186, 2736–2750. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ahmad, A.M.; Ng, H.; Shi, J.; McGhee, C.N.J.; Patel, D.V. Successful culture of human transition zone cells. Clin. Exp. Ophthalmol. 2020, 48, 689–700. [Google Scholar] [CrossRef]
- Repp, A.C.; Barton, L.E.; Gottlieb, J. Naturalistic studies of institutionalized profoundly or severely mentally retarded persons: The relationship of density and behavior. Am. J. Ment. Defic. 1983, 87, 441–447. [Google Scholar] [PubMed]
- World Health Organization. World report on vision. World Health Organ. 2019, 214, 1–160. [Google Scholar]
- Yu, T.; Rajendran, V.; Griffith, M.; Forrester, J.V.; Kuffová, L. High-risk corneal allografts: A therapeutic challenge. World J. Transplant. 2016, 6, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Ang, M.; Moriyama, A.; Colby, K.; Sutton, G.; Liang, L.; Sharma, N.; Hjortdal, J.; Shun Chiu Lam, D.; Williams, G.P.; Armitage, J.; et al. Corneal transplantation in the aftermath of the COVID-19 pandemic: An international perspective. Br. J. Ophthalmol. 2020, 104, 1477. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, C.E.; Rnjak-Kovacina, J.; Kaplan, D.L. Corneal Tissue Engineering: Recent Advances and Future Perspectives. Tissue Eng. Part. B Rev. 2015, 21, 278–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdavi, S.S.; Abdekhodaie, M.J.; Mashayekhan, S.; Baradaran-Rafii, A.; Djalilian, A.R. Bioengineering Approaches for Corneal Regenerative Medicine. Tissue Eng. Regen. Med. 2020, 17, 567–593. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, K.; Shetty, R.; Ghosh, A. Corneal cell therapy: With iPSCs, it is no more a far-sight. Stem Cell Res. Ther. 2018, 9, 287. [Google Scholar] [CrossRef] [Green Version]
- Schwab, I.R.; Reyes, M.; Isseroff, R.R. Successful transplantation of bioengineered tissue replacements in patients with ocular surface disease. Cornea 2000, 19, 421–426. [Google Scholar] [CrossRef]
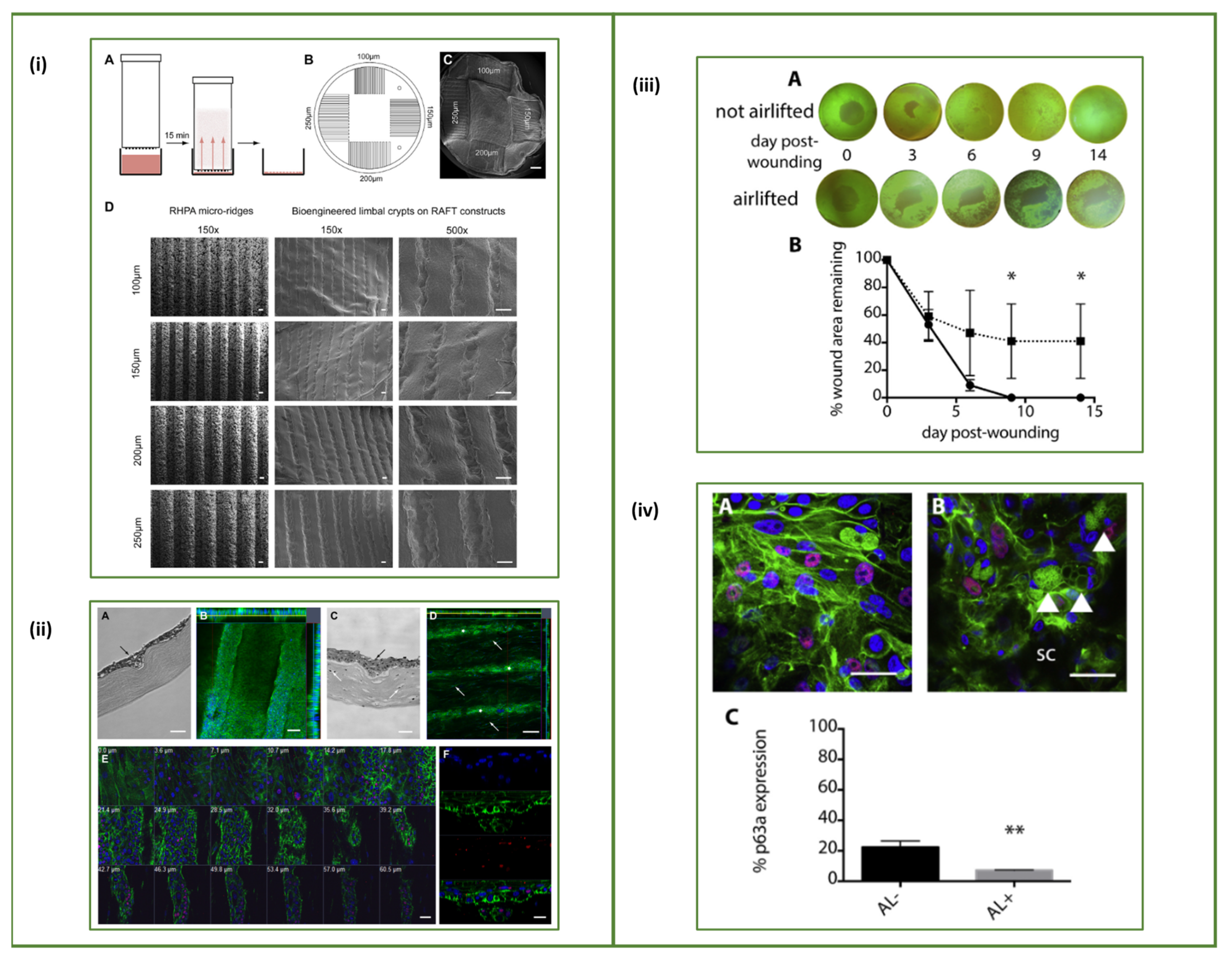
- Ramos-Rodriguez, D.H.; MacNeil, S.; Claeyssens, F.; Asencio, I.O. The Use of Microfabrication Techniques for the Design and Manufacture of Artificial Stem Cell Microenvironments for Tissue Regeneration. Bioengineering 2021, 8, 50. [Google Scholar] [CrossRef]
- Ortega, Í.; Sefat, F.; Deshpande, P.; Paterson, T.; Ramachandran, C.; Ryan, A.J.; Macneil, S.; Claeyssens, F. Combination of microstereolithography and electrospinning to produce membranes equipped with niches for corneal regeneration. J. Vis. Exp. 2014, 12, 51826. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, C.; Sangwan, V.S.; Ortega, I.; Bhatnagar, U.; Mulla, S.M.A.; McKean, R.; MacNeil, S. Synthetic biodegradable alternatives to the use of the amniotic membrane for corneal regeneration: Assessment of local and systemic toxicity in rabbits. Br. J. Ophthalmol. 2019, 103, 286–292. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, C.; Deshpande, P.; Ortega, I.; Sefat, F.; Mckean, R.; Srivastava, M.; Macneil, S.; Basu, S.; Sangwan, V.S. Proof-of-concept study of electrospun PLGA membrane in the treatment of limbal stem cell deficiency. BMJ Open Ophthalmol. 2021, 6, 762. [Google Scholar] [CrossRef]
- Paterson, T.E.; Dhowre, H.S.; Villanueva, D.; Holland, J.W.; Reddy Kethiri, A.; Singh, V.; Claeyssens, F.; MacNeil, S.; Ortega Asencio, I. Tuning Electrospun Substrate Stiffness for the Fabrication of a Biomimetic Amniotic Membrane Substitute for Corneal Healing. ACS Appl. Bio Mater. 2021, 4, 5638–5649. [Google Scholar] [CrossRef]
- Levis, H.J.; Massie, I.; Dziasko, M.A.; Kaasi, A.; Daniels, J.T. Rapid tissue engineering of biomimetic human corneal limbal crypts with 3D niche architecture. Biomaterials 2013, 34, 8860–8868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massie, I.; Levis, H.J.; Daniels, J.T. Response of human limbal epithelial cells to wounding on 3D RAFT tissue equivalents: Effect of airlifting and human limbal fibroblasts. Exp. Eye Res. 2014, 127, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Dziasko, M.A.; Tuft, S.J.; Daniels, J.T. Limbal melanocytes support limbal epithelial stem cells in 2D and 3D microenvironments. Exp. Eye Res. 2015, 138, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Levis, H.J.; Daniels, J.T. Recreating the Human Limbal Epithelial Stem Cell Niche with Bioengineered Limbal Crypts. Curr. Eye Res. 2016, 41, 1153–1160. [Google Scholar] [CrossRef]
- Cortina, M.S.; de la Cruz, J. Keratoprostheses and Artificial Corneas—Fundamentals and Surgical Applications, 1st ed.; Cortina, M.S., de la Cruz, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Matthyssen, S.; Van den Bogerd, B.; Dhubhghaill, S.N.; Koppen, C.; Zakaria, N. Corneal regeneration: A review of stromal replacements. Acta Biomater. 2018, 69, 31–41. [Google Scholar] [CrossRef]
- Lagali, N. Corneal Stromal Regeneration: Current Status and Future Therapeutic Potential. Curr. Eye Res. 2020, 45, 278–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorkio, A.; Koch, L.; Koivusalo, L.; Deiwick, A.; Miettinen, S.; Chichkov, B.; Skottman, H. Human stem cell based corneal tissue mimicking structures using laser-assisted 3D bioprinting and functional bioinks. Biomaterials 2018, 171, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Parekh, M.; Romano, V.; Hassanin, K.; Testa, V.; Wongvisavavit, R.; Ferrari, S.; Haneef, A.; Willoughby, C.; Ponzin, D.; Jhanji, V.; et al. Biomaterials for corneal endothelial cell culture and tissue engineering. J. Tissue Eng. 2021, 12, 2041731421990536. [Google Scholar] [CrossRef] [PubMed]
- Peh, G.S.L.; Ong, H.S.; Adnan, K.; Ang, H.P.; Lwin, C.N.; Seah, X.Y.; Lin, S.J.; Mehta, J.S. Functional Evaluation of Two Corneal Endothelial Cell-Based Therapies: Tissue-Engineered Construct and Cell Injection. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formisano, N.; Sahin, G.; Català, P.; Truckenmüller, R.; Nuijts, R.M.M.A.; Dickman, M.M.; LaPointe, V.L.S.; Giselbrecht, S. Nanoscale Topographies for Corneal Endothelial Regeneration. Appl. Sci. 2021, 11, 827. [Google Scholar] [CrossRef]
- Gutermuth, A.; Maassen, J.; Harnisch, E.; Kuhlen, D.; Sauer-Budge, A.; Skazik-Voogt, C.; Engelmann, K. Descemet’s Membrane Biomimetic Microtopography Differentiates Human Mesenchymal Stem Cells into Corneal Endothelial-Like Cells. Cornea 2019, 38, 110. [Google Scholar] [CrossRef]
- Sharif, R.; Priyadarsini, S.; Rowsey, T.G.; Ma, J.X.; Karamichos, D. Corneal tissue engineering: An in vitro model of the stromal-nerve interactions of the human cornea. J. Vis. Exp. 2018, 131, 56308. [Google Scholar] [CrossRef]
- Wang, S.; Ghezzi, C.E.; Gomes, R.; Pollard, R.E.; Funderburgh, J.L.; Kaplan, D.L. In vitro 3D corneal tissue model with epithelium, stroma, and innervation. Biomaterials 2017, 112, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Efron, N.; Al-Dossari, M.; Pritchard, N. In vivo confocal microscopy of the bulbar conjunctiva. Clin. Exp. Ophthalmol. 2009, 37, 335–344. [Google Scholar] [CrossRef]
- Schrader, S.; Notara, M.; Beaconsfield, M.; Tuft, S.; Geerling, G.; Daniels, J.T. Conjunctival epithelial cells maintain stem cell properties after long-term culture and cryopreservation. Regen. Med. 2009, 4, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Alkatan, H.; AlZahim, T. Histopathology, Srivastava S; Intechopen: London, UK, 2018; Histopathology of the Ocular Surface. [Google Scholar]
- Hosoya, K.I.; Lee, V.H.L.; Kim, K.J. Roles of the conjunctiva in ocular drug delivery: A review of conjunctival transport mechanisms and their regulation. Eur. J. Pharm. Biopharm. 2005, 60, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Eidet, J.; Dartt, D.; Utheim, T. Concise Review: Comparison of Culture Membranes Used for Tissue Engineered Conjunctival Epithelial Equivalents. J. Funct. Biomater. 2015, 6, 1064–1084. [Google Scholar] [CrossRef] [PubMed]
- Meller, D.; Dabul, V.; Tseng, S.C.G. Expansion of conjunctival epithelial progenitor cells on amniotic membrane. Exp. Eye Res. 2002, 74, 537–545. [Google Scholar] [CrossRef]
- Budak, M.T.; Alpdogan, O.S.; Zhou, M.; Lavker, R.M.; Akinci, M.A.M.; Wolosin, J.M. Ocular surface epithelia contain ABCG2-dependent side population cells exhibiting features associated with stem cells. J. Cell Sci. 2005, 118, 1715–1724. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, G.; Golisano, O.; Paterna, P.; Lambiase, A.; Bonini, S.; Rama, P.; De Luca, M. Location and clonal analysis of stem cells and their differentiated progeny in the human ocular surface. J. Cell Biol. 1999, 145, 769–782. [Google Scholar] [CrossRef]
- Chen, W.; Ishikawa, M.; Yamaki, K.; Sakuragi, S. Wistar rat palpebral conjunctiva contains more slow-cycling stem cells that have larger proliferative capacity: Implication for conjunctival epithelial homeostasis. Jpn. J. Ophthalmol. 2003, 47, 119–128. [Google Scholar] [CrossRef]
- Pe’er, J.; Zajicek, G.; Greifner, H.; Kogan, M. Streaming conjunctiva. Anat. Rec. 1996, 245, 36–40. [Google Scholar] [CrossRef]
- Wirtschafter, J.D.; Ketcham, J.M.; Weinstock, R.J.; Tabesh, T.; McLoon, L.K. Mucocutaneous junction as the major source of replacement palpebral conjunctival epithelial cells. Investig. Ophthalmol. Vis. Sci. 1999, 40, 3138–3146. [Google Scholar]
- Ramos, T.; Scott, D.; Ahmad, S. An Update on Ocular Surface Epithelial Stem Cells: Cornea and Conjunctiva. Stem Cells Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.G.; Wu, R.L.; Lavker, R.M.; Sun, T.T. In vitro growth and differentiation of rabbit bulbar, fornix, and palpebral conjunctival epithelia: Implications on conjunctival epithelial transdifferentiation and stem cells. Investig. Ophthalmol. Vis. Sci. 1993, 34, 1814–1828. [Google Scholar]
- Wei, Z.G.; Cotsarelis, G.; Sun, T.T.; Lavker, R.M. Label-retaining cells are preferentially located in fornical epithelium: Implications on conjunctival epithelial homeostasis. Investig. Ophthalmol. Vis. Sci. 1995, 36, 236–246. [Google Scholar]
- Wei, Z.G.; Lin, T.; Sun, T.T.; Lavker, R.M. Clonal analysis of the in vivo differentiation potential of keratinocytes. Investig. Ophthalmol. Vis. Sci. 1997, 38, 753–761. [Google Scholar]
- Clark, J.N.; Marchok, A.C. The Effect of Vitamin A on Cellular Differentiation and Mucous Glycoprotein Synthesis in Long-Term Rat Tracheal Organ Cultures. Differentiation 1979, 14, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Tsai, R.J.F.; Tseng, S.C.G. Substrate modulation of cultured rabbit conjunctival epithelial cell differentiation and morphology. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1565–1576. [Google Scholar]
- Tsai, R.J.F.; Tseng, S.C.G.; Chen, J.K. Conjunctival epithelial cells in culture-growth and goblet cell differentiation. Prog. Retin. Eye Res. 1997, 16, 227–241. [Google Scholar] [CrossRef]
- Ruiz-Alonso, S.; Villate-Beitia, I.; Gallego, I.; Lafuente-Merchan, M.; Puras, G.; Saenz-del-Burgo, L.; Pedraz, J.L. Current insights into 3D bioprinting: An advanced approach for eye tissue regeneration. Pharmaceutics 2021, 13, 308. [Google Scholar] [CrossRef]
- Yao, Q.; Hu, Y.; Yu, F.; Zhang, W.; Fu, Y. A novel application of electrospun silk fibroin/poly(l-lactic acid-: Co -ϵ-caprolactone) scaffolds for conjunctiva reconstruction. RSC Adv. 2018, 8, 18372–18380. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Li, X.; Gao, W.; Fu, X.; Fang, R.H.; Zhang, L.; Zhang, K. Tissue repair and regeneration with endogenous stem cells. Nat. Rev. Mater. 2018, 3, 174–193. [Google Scholar] [CrossRef]
- Lane, S.W.; Williams, D.A.; Watt, F.M. Modulating the stem cell niche for tissue regeneration. Nat. Biotechnol. 2014, 32, 795–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vining, K.H.; Mooney, D.J. Mechanical forces direct stem cell behaviour in development and regeneration. Nat. Rev. Mol. Cell Biol. 2017, 18, 728–742. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Deng, X.; Wang, P.; Yu, C.; Kiratitanaporn, W.; Wu, X.; Schimelman, J.; Tang, M.; Balayan, A.; Yao, E.; et al. Rapid bioprinting of conjunctival stem cell micro-constructs for subconjunctival ocular injection. Biomaterials 2021, 267, 120462. [Google Scholar] [CrossRef]
- Kasbekar, S.; Kaye, S.B.; Williams, R.L.; Stewart, R.M.K.; Leow-Dyke, S.; Rooney, P. Development of decellularized conjunctiva as a substrate for the ex vivo expansion of conjunctival epithelium. J. Tissue Eng. Regen. Med. 2018, 12, e973–e982. [Google Scholar] [CrossRef]
- Dehghani, S.; Rasoulianboroujeni, M.; Ghasemi, H.; Keshel, S.H.; Nozarian, Z.; Hashemian, M.N.; Zarei-Ghanavati, M.; Latifi, G.; Ghaffari, R.; Cui, Z.; et al. 3D-Printed membrane as an alternative to amniotic membrane for ocular surface/conjunctival defect reconstruction: An in vitro & in vivo study. Biomaterials 2018, 174, 95–112. [Google Scholar] [CrossRef]
- Bosworth, L.A.; Doherty, K.G.; Hsuan, J.D.; Cray, S.P.; D’sa, R.A.; Molina, C.P.; Badylak, S.F.; Williams, R.L. Material characterisation and stratification of conjunctival epithelial cells on electrospun poly(ε-caprolactone) fibres loaded with decellularised tissue matrices. Pharmaceutics 2021, 13, 318. [Google Scholar] [CrossRef]
- Hong, S.; Kim, G. Electrospun micro/nanofibrous conduits composed of poly(ε- caprolactone) and small intestine submucosa powder for nerve tissue regeneration. J. Biomed. Mater. Res. Part. B Appl. Biomater. 2010, 94, 421–428. [Google Scholar] [CrossRef]
- Schrader, S.; Notara, M.; Tuft, S.J.; Beaconsfield, M.; Geerling, G.; Daniels, J.T. Simulation of an in vitro niche environment that preserves conjunctival progenitor cells. Regen. Med. 2010, 5, 877–889. [Google Scholar] [CrossRef]
- Schrader, S.; Notara, M.; Beaconsfield, M.; Tuft, S.J.; Daniels, J.T.; Geerling, G. Tissue engineering for conjunctival reconstruction: Established methods and future outlooks. Curr. Eye Res. 2009, 34, 913–924. [Google Scholar] [CrossRef]
- Tsai, R.J.F.; Ho, Y.S.; Chen, J.K. The effects of fibroblasts on the growth and differentiation of human bulbar conjunctival epithelial cells in an in vitro conjunctival equivalent. Investig. Ophthalmol. Vis. Sci. 1994, 35, 2865–2875. [Google Scholar]
- Moazed, K.T. The Iris: Understanding the Essentials; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Addo, R.T. Ocular Drug Delivery: Advances, Challenges and Applications; Springer: Cham, Switzerland, 2016; pp. 53–74. [Google Scholar] [CrossRef]
- Davis-Silberman, N.; Ashery-Padan, R. Iris development in vertebrates; genetic and molecular considerations. Brain Res. 2008, 1192, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhang, E.; Del Rio-Tsonis, K. Eye Anatomy. In eLS; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Thumann, G. Development and cellular functions of the iris pigment epithelium. Surv. Ophthalmol. 2001, 45, 345–354. [Google Scholar] [CrossRef]
- Malhotra, A.; Minja, F.J.; Crum, A.; Burrowes, D. Ocular Anatomy and Cross-Sectional Imaging of the Eye. Semin. Ultrasound CT MRI 2011, 32, 2–13. [Google Scholar] [CrossRef]
- Kosaka, M.; Kodama, R.; Eguchi, G. In vitro culture system for iris-pigmented epithelial cells for molecular analysis of transdifferentiation. Exp. Cell Res. 1998, 245, 245–251. [Google Scholar] [CrossRef]
- Tsonis, P.A.; Jang, W.; Del Rio-Tsonis, K.; Eguchi, G. A unique aged human retinal pigmented epithelial cell line useful for studying lens differentiation in vitro. Int. J. Dev. Biol. 2001, 45, 753–758. [Google Scholar] [CrossRef]
- Akagi, T.; Mandai, M.; Ooto, S.; Hirami, Y.; Osakada, F.; Kageyama, R.; Yoshimura, N.; Takahashi, M. Otx2 homeobox gene induces photoreceptor-specific phenotypes in cells derived from adult iris and ciliary tissue. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4570–4575. [Google Scholar] [CrossRef] [Green Version]
- Akagi, T.; Akita, J.; Haruta, M.; Suzuki, T.; Honda, Y.; Inoue, T.; Yoshiura, S.; Kageyama, R.; Yatsu, T.; Yamada, M.; et al. Iris-derived cells from adult rodents and primates adopt photoreceptor-specific phenotypes. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3411–3419. [Google Scholar] [CrossRef]
- Haruta, M.; Kosaka, M.; Kanegae, Y.; Saito, I.; Inoue, T.; Kageyama, R.; Nishida, A.; Honda, Y.; Takahashi, M. Induction of photoreceptor-specific phenotypes in adult mammalian iris tissue. Nat. Neurosci. 2001, 4, 1163–1164. [Google Scholar] [CrossRef]
- Tropepe, V.; Coles, B.L.K.; Chiasson, B.J.; Horsford, D.J.; Elia, A.J.; McInnes, R.R.; Van Der Kooy, D.D. Retinal stem cells in the adult mammalian eye. Science 2000, 287, 2032–2036. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Tang, L.; Pham, H. Identification of neural progenitors in the adult mammalian eye. Biochem. Biophys. Res. Commun. 2000, 270, 517–521. [Google Scholar] [CrossRef]
- Nian, S.; Kearns, V.R.; Wong, D.S.H.; Bachhuka, A.; Vasilev, K.; Williams, R.L.; Lai, W.W.; Lo, A.; Sheridan, C.M. Plasma polymer surface modified expanded polytetrafluoroethylene promotes epithelial monolayer formation in vitro and can be transplanted into the dystrophic rat subretinal space. J. Tissue Eng. Regen. Med. 2021, 15, 49–62. [Google Scholar] [CrossRef]
- Thompson, J.; Lakhani, N. Cataracts. Prim. Care Clin. Off. Pract. 2015, 42, 409–423. [Google Scholar] [CrossRef]
- Abdelkader, H.; Alany, R.G.; Pierscionek, B. Age-related cataract and drug therapy: Opportunities and challenges for topical antioxidant delivery to the lens. J. Pharm. Pharmacol. 2015, 67, 537–550. [Google Scholar] [CrossRef]
- Borchman, D.; Yappert, M.C. Lipids and the ocular lens. J. Lipid Res. 2010, 51, 2473–2488. [Google Scholar] [CrossRef] [Green Version]
- Bozukova, D.; Pagnoulle, C.; Jérôme, R.; Jérôme, C. Polymers in modern ophthalmic implants—Historical background and recent advances. Mater. Sci. Eng. R Rep. 2010, 69, 63–83. [Google Scholar] [CrossRef]
- Hejtmancik, J.F.; Shiels, A. Overview of the Lens. Prog. Mol. Biol. Transl. Sci. 2015, 134, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Danysh, B.P.; Duncan, M.K. The lens capsule. Exp. Eye Res. 2009, 88, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Remington, S.G.; Meyer, R.A. Lens stem cells may reside outside the lens capsule: An hypothesis. Theor. Biol. Med. Model. 2007, 4, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekin, K.; Ozdamar Erol, Y.; Inanc, M.; Sargon, M.F.; Cakar Ozdal, P.; Berker, N. Ultrastructural Analysis of the Anterior Lens Epithelium in Cataracts Associated with Uveitis. Ophthalmic Res. 2020, 63, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Martinez, G.; De Iongh, R.U. The lens epithelium in ocular health and disease. Int. J. Biochem. Cell Biol. 2010, 42, 1945–1963. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.K.; Santhoshkumar, P. Lens aging: Effects of crystallins. Biochim. Biophys. Acta 2009, 1790, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Garcia, C.M.; Shui, Y.B.; Beebe, D.C. Expression and regulation of α-, β-, and γ-crystallins in mammalian lens epithelial cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3608–3619. [Google Scholar] [CrossRef]
- Vaghefi, E.; Kim, A.; Donaldson, P.J. Active maintenance of the gradient of refractive index is required to sustain the optical properties of the lens. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7195–7208. [Google Scholar] [CrossRef] [Green Version]
- Derham, B.K.; Harding, J.J. α-Crystallin as a molecular chaperone. Prog. Retin. Eye Res. 1999, 18, 463–509. [Google Scholar] [CrossRef]
- Kumar, P.A.; Reddy, G.B. Modulation of α-crystallin chaperone activity: A target to prevent or delay cataract? IUBMB Life 2009, 61, 485–495. [Google Scholar] [CrossRef]
- Yamamoto, N.; Majima, K.; Marunouchi, T. A study of the proliferating activity in lens epithelium and the identification of tissue-type stem cells. Med. Mol. Morphol. 2008, 41, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Raji, B.; Dansault, A.; Leemput, J.; de la Houssaye, G.; Vieira, V.; Kobetz, A.; Arbogast, L.; Masson, C.; Menasche, M.; Abitbol, M. The RNA-binding protein Musashi-1 is produced in the developing and adult mouse eye. Mol. Vis. 2007, 13, 1412–1427. [Google Scholar]
- Van Schoore, G.; Mendive, F.; Pochet, R.; Vassart, G. Expression pattern of the orphan receptor LGR4/GPR48 gene in the mouse. Histochem. Cell Biol. 2005, 124, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Wormstone, I.M.; Eldred, J.A. Experimental models for posterior capsule opacification research. Exp. Eye Res. 2016, 142, 2–12. [Google Scholar] [CrossRef]
- Wormstone, I.M. Posterior capsule opacification: A cell biological perspective. Exp. Eye Res. 2002, 74, 337–347. [Google Scholar] [CrossRef]
- Alió, J.L.; Alió del Barrio, J.L.; Vega-Estrada, A. Accommodative intraocular lenses: Where are we and where we are going. Eye Vis. 2017, 4, 1–12. [Google Scholar] [CrossRef]
- Wang, K.; Han, Z. Injectable hydrogels for ophthalmic applications. J. Control. Release 2017, 268, 212–224. [Google Scholar] [CrossRef]
- De Groot, J.H.; Van Beijma, F.J.; Haitjema, H.J.; Dillingham, K.A.; Hodd, K.A.; Koopmans, S.A.; Norrby, S. Injectable intraocular lens materials based upon hydrogels. Biomacromolecules 2001, 2, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Coca-Prados, M.; Escribano, J. New perspectives in aqueous humor secretion and in glaucoma: The ciliary body as a multifunctional neuroendocrine gland. Prog. Retin. Eye Res. 2007, 26, 239–262. [Google Scholar] [CrossRef] [PubMed]
- Delamere, N.A. Ciliary Body and Ciliary Epithelium. Adv. Organ. Biol. 2005, 10, 127–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges- Giampani, A.S.; Giampani, J. Anatomy of Ciliary Body, Ciliary Processes, Anterior Chamber Angle and Collector Vessels. Glaucoma Basic Clin. Asp. 2013, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Soni, V.; Pandey, V.; Tiwari, R.; Asati, S.; Tekade, R.K. Design and evaluation of ophthalmic delivery formulations. Basic Fundam. Drug Deliv. 2018, 473–538. [Google Scholar] [CrossRef]
- Dhamodaran, K.; Shetty, R.; Subramani, M.; Das, D. Ocular stem cells: An overview. In Recent Advances in Ophthalmology Research; Nova: Hauppauge, NY, USA, 2013; pp. 103–135. [Google Scholar]
- Gelatt, K.N.; Wilkie, D.A. Surgical procedures of the anterior chamber and anterior uvea. In Veterinary Ophthalmic Surgery; Elsevier: Amsterdam, The Netherlands, 2011; pp. 237–262. [Google Scholar] [CrossRef]
- Napier, H.R.L.; Kidson, S.H. Molecular events in early development of the ciliary body: A question of folding. Exp. Eye Res. 2007, 84, 615–625. [Google Scholar] [CrossRef]
- Attar, M.; Brassard, J.A.; Kim, A.S.; Matsumoto, S.; Ramos, M.; Vangyi, C. Safety evaluation of ocular drugs. In A Comprehensive Guide to Toxicology in Preclinical Drug Development; Elsevier: Amsterdam, The Netherlands, 2013; pp. 567–617. [Google Scholar] [CrossRef]
- Goel, M.e.a. Aqueous Humor Dynamics: A Review. Open Ophthalmol. J. 2010, 4, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Zeiss, C.J.; Tu, D.C.; Phan, I.; Wong, R.; Treuting, P.M. Special Senses: Eye. Comp. Anat. Histol. 2018, 445–470. [Google Scholar] [CrossRef]
- Tamm, E.R. The role of the ciliary body in aqueous humor dynamics structural aspects. Encycl. Eye 2010, 179–186. [Google Scholar] [CrossRef]
- Fischer, A.J.; Reh, T.A. Growth factors induce neurogenesis in the ciliary body. Dev. Biol. 2003, 259, 225–240. [Google Scholar] [CrossRef] [Green Version]
- Pašta, J. Laser therapy in ophthalmology. Lasers Med. Appl. Diagn. Ther. Surg. 2013, 1960, 395–458. [Google Scholar] [CrossRef]
- Lin, T.-C.; Hsu, C.-C.; Chien, K.-H.; Hung, K.-H.; Peng, C.-H.; Chen, S.-J. Retinal stem cells and potential cell transplantation treatments. J. Chin. Med. Assoc. 2014, 77, 556–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, P.A.; Barthel, L.K.; Bernardos, R.L.; Perkowski, J.J. Molecular characterization of retinal stem cells and their niches in adult zebrafish. BMC Dev. Biol. 2006, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Miyake, A.; Araki, M. Retinal stem/progenitor cells in the ciliary marginal zone complete retinal regeneration: A study of retinal regeneration in a novel animal model. Dev. Neurobiol. 2014, 74, 739–756. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, C.; Ueda, Y.; Okamoto, M.; Araki, M. Neural retinal regeneration in the anuran amphibian Xenopus laevis post-metamorphosis: Transdifferentiation of retinal pigmented epithelium regenerates the neural retina. Dev. Biol. 2007, 303, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Gao, J.; Jia, X.; Zhao, W.; Zhang, Y.; Pan, W.; He, J. Bipotent progenitors as embryonic origin of retinal stem cells. J. Cell Biol. 2017, 216, 1833–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikelle, L.; Al-Ubaidi, M.R.; Naash, M.I. Pluripotent Stem Cells for the Treatment of Retinal Degeneration: Current Strategies and Future Directions. Front. Cell Dev. Biol. 2020, 8, 743. [Google Scholar] [CrossRef] [PubMed]
- Marcucci, F.; Murcia-Belmonte, V.; Wang, Q.; Coca, Y.; Ferreiro-Galve, S.; Kuwajima, T.; Khalid, S.; Ross, M.E.; Mason, C.; Herrera, E. The Ciliary Margin Zone of the Mammalian Retina Generates Retinal Ganglion Cells. Cell Rep. 2016, 17, 3153–3164. [Google Scholar] [CrossRef]
- Del Debbio, C.B.; Peng, X.; Xiong, H.; Ahmad, I. Adult ciliary epithelial stem cells generate functional neurons and differentiate into both early and late born retinal neurons under non-cell autonomous influences. BMC Neurosci. 2013, 14, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballios, B.G.; Clarke, L.; Coles, B.L.K.; Shoichet, M.S.; Van Der Kooy, D. The adult retinal stem cell is a rare cell in the ciliary epithelium whose progeny can differentiate into photoreceptors. Biol. Open 2012, 1, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cicero, S.A.; Johnson, D.; Reyntjens, S.; Frase, S.; Connell, S.; Chow, L.M.L.; Baker, S.J.; Sorrentino, B.P.; Dyer, M.A. Cells previously identified as retinal stem cells are pigmented ciliary epithelial cells. Proc. Natl. Acad. Sci. USA 2009, 106, 6685–6690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualdoni, S.; Baron, M.; Lakowski, J.; Decembrini, S.; Smith, A.J.; Pearson, R.A.; Ali, R.R.; Sowden, J.C. Adult ciliary epithelial cells, previously identified as retinal stem cells with potential for retinal repair, fail to differentiate into new rod photoreceptors. Stem Cells (Dayt. Ohio) 2010, 28, 1048–1059. [Google Scholar] [CrossRef] [PubMed]
- Coles, B.L.K.; Angénieux, B.; Inoue, T.; Del Rio-Tsonis, K.; Spence, J.R.; McInnes, R.R.; Arsenijevic, Y.; Van Der Kooy, D. Facile isolation and the characterization of human retinal stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 15772–15777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Iglesia, D.; Kielczewski, J.; Valenta, D.; Pease, M.E.; Zack, D.; Quigley, H. Characteristics of Progenitor Cells Derived from Adult Ciliary Body in Mouse, Rat, and Human Eyes. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1674–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frøen, R.; Johnsen, E.O.; Nicolaissen, B.; Facskó, A.; Petrovski, G.; Moe, M.C. Does the adult human ciliary body epithelium contain “true” retinal stem cells? BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef]
- Kuwahara, A.; Ozone, C.; Nakano, T.; Saito, K.; Eiraku, M.; Sasai, Y. Generation of a ciliary margin-like stem cell niche from self-organizing human retinal tissue. Nat. Commun. 2015, 6, 6286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, B.; Singhal, S.; Jayaram, H.; Khaw, P.T.; Limb, G.A. Adult retinal stem cells revisited. Open Ophthalmol. J. 2010, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Larsen, K.B.; Lutterodt, M.C.; Møllgård, K.; Møller, M. Expression of the Homeobox Genes OTX2 and OTX1 in the Early Developing Human Brain. J. Histochem. Cytochem. 2010, 58, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, H.; Suzuma, K.; Kaneko, J.; Mandai, M.; Kitaoka, T.; Takahashi, M. Induction of functional 3D ciliary epithelium–like structure from mouse induced pluripotent stem cells. Investig. Ophthalmol. Vis. Sci. 2016, 57, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llobet, A.; Gasull, X.; Gual, A. Understanding Trabecular Meshwork Physiology: A Key to the Control of Intraocular Pressure? Physiology 2003, 18, 205–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tun, T.A.; Baskaran, M.; Zheng, C.; Sakata, L.M.; Perera, S.A.; Chan, A.S.; Friedman, D.S.; Cheung, C.Y.; Aung, T. Assessment of trabecular meshwork width using swept source optical coherence tomography. Graefes Arch. Clin. Exp. Ophthalmol. 2013, 251, 1587–1592. [Google Scholar] [CrossRef] [PubMed]
- Abu-Hassan, D.W.; Acott, T.S.; Kelley, M.J. The Trabecular Meshwork: A Basic Review of Form and Function. J. Ocul. Biol. 2014, 2. [Google Scholar] [CrossRef]
- Tamm, E.R. The trabecular meshwork outflow pathways: Structural and functional aspects. Exp. Eye Res. 2009, 88, 648–655. [Google Scholar] [CrossRef]
- King, B.J.; Burns, S.A.; Sapoznik, K.A.; Luo, T.; Gast, T.J. High-resolution, adaptive optics imaging of the human trabecular meshwork in vivo. Transl. Vis. Sci. Technol. 2019, 8, 5. [Google Scholar] [CrossRef]
- Stamer, W.D.; Clark, A.F. The many faces of the trabecular meshwork cell. Exp. Eye Res. 2017, 158, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Overby, D.R.; Stamer, W.D.; Johnson, M. The changing paradigm of outflow resistance generation: Towards synergistic models of the JCT and inner wall endothelium. Exp. Eye Res. 2009, 88, 656–670. [Google Scholar] [CrossRef] [Green Version]
- Buller, C.; Johnson, D.H.; Tschumper, R.C. Human trabecular meshwork phagocytosis: Observations in an organ culture system. Investig. Ophthalmol. Vis. Sci. 1990, 31, 2156–2163. [Google Scholar]
- Acott, T.S.; Samples, J.R.; Bradley, J.M.B.; Bacon, D.R.; Bylsma, S.S.; Van Buskirk, E.M. Trabecular repopulation by anterior trabecular meshwork cells after laser trabeculoplasty. Am. J. Ophthalmol. 1989, 107, 1–6. [Google Scholar] [CrossRef]
- Kelley, M.J.; Rose, A.Y.; Keller, K.E.; Hessle, H.; Samples, J.R.; Acott, T.S. Stem cells in the trabecular meshwork: Present and future promises. Exp. Eye Res. 2009, 88, 747–751. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.Y.; Grierson, I.; Sheridan, C.; Lo, A.C.Y.; Wong, D.S.H. Bovine posterior limbus: An evaluation of an alternative source for corneal endothelial and trabecular meshwork stem/progenitor cells. Stem Cells Dev. 2015, 24, 624–639. [Google Scholar] [CrossRef] [Green Version]
- Crouch, D.J.; Sheridan, C.M.; D’Sa, R.A.; Willoughby, C.E.; Bosworth, L.A. Exploiting biomaterial approaches to manufacture an artificial trabecular meshwork: A progress report. Biomater. Biosyst. 2021, 1, 100011. [Google Scholar] [CrossRef]
- Keller, K.E.; Bhattacharya, S.K.; Borrás, T.; Brunner, T.M.; Chansangpetch, S.; Clark, A.F.; Dismuke, W.M.; Du, Y.; Elliott, M.H.; Ethier, C.R.; et al. Consensus recommendations for trabecular meshwork cell isolation, characterization and culture. Exp. Eye Res. 2018, 171, 164–173. [Google Scholar] [CrossRef]
- Reina-Torres, E.; De Ieso, M.L.; Pasquale, L.R.; Madekurozwa, M.; van Batenburg-Sherwood, J.; Overby, D.R.; Stamer, W.D. The vital role for nitric oxide in intraocular pressure homeostasis. Prog. Retin. Eye Res. 2021, 83, 100922. [Google Scholar] [CrossRef]
- Kumar, A.; Xu, Y.; Du, Y. Stem Cells from Human Trabecular Meshwork Hold the Potential to Develop into Ocular and Non-Ocular Lineages After Long-Term Storage. Stem Cells Dev. 2020, 29, 49–61. [Google Scholar] [CrossRef]
- Whikehart, D.R.; Parikh, C.H.; Vaughn, A.V.; Mishler, K.; Edelhauser, H.F. Evidence suggesting the existence of stem cells for the human corneal epithelium. Mol. Vis. 2005, 816–824. [Google Scholar]
- McGowan, S.; Edelhauser, H.; Pfister, R.; Whikehart, D.R. Stem cell markers in the human posterior limbus and corneal endothelium of unwounded and wounded corneas. Mol. Vis. 2007, 13, 1984–2000. [Google Scholar] [PubMed]
- Du, Y.; Yun, H.; Yang, E.; Schuman, J.S. Stem cells from trabecular meshwork home to TM tissue in vivo. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Bilir, E.K.; Kingston, O.A.; Oldershaw, R.A.; Kearns, V.R.; Willoughby, C.E.; Sheridan, C.M. Replacement of the Trabecular Meshwork Cells—A Way Ahead in IOP Control? Biomolecules 2021, 11, 1371. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; McNally, S.; Kilpatrick, J.I.; Jarvis, S.P.; O’Brien, C.J. Aging and ocular tissue stiffness in glaucoma. Surv. Ophthalmol. 2018, 63, 56–74. [Google Scholar] [CrossRef]
- Chowdhury, U.R.; Hann, C.R.; Stamer, W.D.; Fautsch, M.P. Aqueous humor outflow: Dynamics and disease. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2993–3003. [Google Scholar] [CrossRef] [Green Version]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. JAMA J. Am. Med. Assoc. 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Grierson, I.; Howes, R.C. Age-related depletion of the cell population in the human trabecular meshwork. Eye (Basingstoke) 1987, 1, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Tektas, O.Y.; Lütjen-Drecoll, E. Structural changes of the trabecular meshwork in different kinds of glaucoma. Exp. Eye Res. 2009, 88, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, J.; Murphy, C.; Juster, R. Trabecular Meshwork Cellularity in Primary Open-angle Glaucoma and Nonglaucomatous Normals. Ophthalmology 1984, 91, 564–579. [Google Scholar] [CrossRef]
- Last, J.A.; Pan, T.; Ding, Y.; Reilly, C.M.; Keller, K.; Acott, T.S.; Fautsch, M.P.; Murphy, C.J.; Russell, P. Elastic modulus determination of normal and glaucomatous human trabecular meshwork. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2147–2152. [Google Scholar] [CrossRef] [PubMed]
- Lütjen-Drecoll, E.; Rittig, M.; Rauterberg, J.; Jander, R.; Mollenhauer, J. Immunomicroscopical study of type VI collagen in the trabecular meshwork of normal and glaucomatous eyes. Exp. Eye Res. 1989, 48, 139–147. [Google Scholar] [CrossRef]
- Medina-Ortiz, W.E.; Belmares, R.; Neubauer, S.; Wordinger, R.J.; Clark, A.F. Cellular fibronectin expression in human trabecular meshwork and induction by transforming growth factor-β2. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6779–6788. [Google Scholar] [CrossRef] [PubMed]
- Kasetti, R.B.; Maddineni, P.; Millar, J.C.; Clark, A.F.; Zode, G.S. Increased synthesis and deposition of extracellular matrix proteins leads to endoplasmic reticulum stress in the trabecular meshwork. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Trappmann, B.; Chen, C.S. How cells sense extracellular matrix stiffness: A material’s perspective. Curr. Opin. Biotechnol. 2013, 24, 948–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghunathan, V.K.; Benoit, J.; Kasetti, R.; Zode, G.; Salemi, M.; Phinney, B.S.; Keller, K.E.; Staverosky, J.A.; Murphy, C.J.; Acott, T.; et al. Glaucomatous cell derived matrices differentially modulate non-glaucomatous trabecular meshwork cellular behavior. Acta Biomater. 2018, 71, 444–459. [Google Scholar] [CrossRef] [PubMed]
- Dautriche, C.N.; Xie, Y.; Sharfstein, S.T. Walking through trabecular meshwork biology: Toward engineering design of outflow physiology. Biotechnol. Adv. 2014, 32, 971–983. [Google Scholar] [CrossRef]
- Waduthanthri, K.D.; He, Y.; Montemagno, C.; Cetinel, S. An injectable peptide hydrogel for reconstruction of the human trabecular meshwork. Acta Biomater. 2019, 100, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Izagirre, M.; González, E.; Esquisabel, L.; Aldazabal, J.; Montañés, J.; Paredes, J. Characterization of Polycaprolactone Based Electrospun Scaffold Towards in Vitro Human Trabecular Meshwork Model. 2020. Available online: https://www.researchgate.net/publication/346491866_Characterization_of_polycaprolactone_based_electrospun_scaffold_towards_in_vitro_human_trabecular_meshwork_model?_sg=ewv6Wf42MAAbJGflTCLBVExGMUI6vLystoDbcd6XiM0jn1rrIEt108iloUwGaeAqozQvukdHN08_b1tnh2Apz8q8hrE (accessed on 31 August 2021).
- Berkowski, K.L.; Plunkett, K.N.; Yu, Q.; Moore, J.S. Introduction to photolithography: Preparation of microscale polymer silhouettes. J. Chem. Educ. 2005, 82, 1365–1369. [Google Scholar] [CrossRef]
- Torrejon, K.Y.; Pu, D.; Bergkvist, M.; Danias, J.; Sharfstein, S.T.; Xie, Y. Recreating a human trabecular meshwork outflow system on microfabricated porous structures. Biotechnol. Bioeng. 2013, 110, 3205–3218. [Google Scholar] [CrossRef]
- Wu, H.; Chen, W.; Zhao, F.; Zhou, Q.; Reinach, P.S.; Deng, L.; Ma, L.; Luo, S.; Srinivasalu, N.; Pan, M.; et al. Scleral hypoxia is a target for myopia control. Proc. Natl. Acad. Sci. USA 2018, 115, E7091–E7100. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Zhang, D.; Zhou, Q.; Zhao, F.; He, M.; Yang, Z.; Su, Y.; Zhai, Y.; Yan, J.; Zhang, G.; et al. Scleral HIF-1α is a prominent regulatory candidate for genetic and environmental interactions in human myopia pathogenesis. EBioMedicine 2020, 57. [Google Scholar] [CrossRef]
- Holden, B.A.; Fricke, T.R.; Wilson, D.A.; Jong, M.; Naidoo, K.S.; Sankaridurg, P.; Wong, T.Y.; Naduvilath, T.J.; Resnikoff, S. Global Prevalence of Myopia and High Myopia and Temporal Trends from 2000 through 2050. Ophthalmology 2016, 123, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Ohno-Matsui, K. Sclera-targeted therapies for pathologic myopia. In Pathologic Myopia; Spaide, R., Ohno-Matsui, K., Yannuzzi, L., Eds.; Springer: New York, NY, USA, 2014; pp. 353–360. [Google Scholar] [CrossRef]
- Summers Rada, J.A.; Shelton, S.; Norton, T.T. The sclera and myopia. Exp. Eye Res. 2006, 82, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.L.; Wu, P.C.; Elizabeth Fini, M.; Shi, S. Identification of multipotent stem/progenitor cells in murine sclera. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5481–5487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.E.; Kwan, J.L.; Goh, L.K.; Beuerman, R.W.; Barathi, V.A. Evaluation of gene expression profiles and pathways underlying postnatal development in mouse sclera. Mol. Vis. 2012, 18, 1436–1448. [Google Scholar] [PubMed]
- Carriel, V.; Vizcaíno-López, G.; Chato-Astrain, J.; Durand-Herrera, D.; Alaminos, M.; Campos, A.; Sánchez-Montesinos, I.; Campos, F. Scleral surgical repair through the use of nanostructured fibrin/agarose-based films in rabbits. Exp. Eye Res. 2019, 186, 107717. [Google Scholar] [CrossRef]
- Watson, P.G.; Young, R.D. Scleral structure, organisation and disease. A review. Exp. Eye Res. 2004, 78, 609–623. [Google Scholar] [CrossRef]
- Treuting, P.M.; Wong, R.; Tu, D.C.; Phan, I. Special Senses: Eye. Comp. Anat. Histol. 2012, 395–418. [Google Scholar] [CrossRef]
- Seko, Y.; Azuma, N.; Yokoi, T.; Kami, D.; Ishii, R.; Nishina, S.; Toyoda, M.; Shimokawa, H.; Umezawa, A. Anteroposterior Patterning of Gene Expression in the Human Infant Sclera: Chondrogenic Potential and Wnt Signaling. Curr. Eye Res. 2017, 42, 145–154. [Google Scholar] [CrossRef]
- Boote, C.; Sigal, I.A.; Grytz, R.; Hua, Y.; Nguyen, T.D.; Girard, M.J.A. Scleral structure and biomechanics HHS Public Access. Prog. Retin. Eye Res. 2020, 74, 100773. [Google Scholar] [CrossRef]
- Petrea, C.E.; Crăiţoiu; Vrapciu, A.D.; Mănoiu, V.S.; Rusu, M.C. The telopode- and filopode-projecting heterogeneous stromal cells of the human sclera niche. Ann. Anat. 2018, 218, 129–140. [Google Scholar] [CrossRef]
- Eberwein, P.; Reinhard, T. Concise reviews: The role of biomechanics in the limbal stem cell niche: New insights for our understanding of this structure. Stem Cells 2015, 33, 916–924. [Google Scholar] [CrossRef]
- Grieve, K.; Ghoubay, D.; Georgeon, C.; Thouvenin, O.; Bouheraoua, N.; Paques, M.; Borderie, V.M. Three-dimensional structure of the mammalian limbal stem cell niche. Exp. Eye Res. 2015, 140, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Tena, M.A.; Alvarez-Peregrina, C.; Santos-Arias, F.; Villa-Collar, C. Application of 3D Printing Technology in Scleral Cover Shell Prosthesis. J. Med. Syst. 2019, 43, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.B.; Jha, A.K.; Healy, K.E.; Wildsoet, C.F. A bioengineering approach to myopia control tested in a Guinea pig model. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1875–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limb, G.A.; Daniels, J.T. Ocular regeneration by stem cells: Present status and future prospects. Br. Med. Bull. 2008, 85, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Gattazzo, F.; Urciuolo, A.; Bonaldo, P. Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 2506–2519. [Google Scholar] [CrossRef]









| Niche | Stem Cell Type | Potency | Regulatory Signalling Pathways | Markers | Ref. |
|---|---|---|---|---|---|
| Cornea (limbus) | Limbal stem cells | Unipotent | Notch, Wnt, Stat3, IL6, Shh | Undifferentiated: TP63, ABCG2, ABCB5, K14, K15, K19, K3/K12, ΔNp63α Differentiated: K3, involucrin, connexin 43, KRT12, KRT24, AREG | [28,29,30,31] |
| Stromal stem cells | Multipotent | Undifferentiated: ABCG2, Nestin, Pax6, BMi1, CD73, CD90, CD166, cKIT, Notch1, Six2 Differentiated: ALDH3A1, CXADR, PTDGS, PDK4, CD34 Insignificant expression of fibroblast related genes: αSMA and TCC | [32,33,34] | ||
| Conjunctiva (epithelium) | Conjunctival stem cells | Bipotent-goblet and non-goblet cells | NGF-p75-SALL2, Notch and Wnt | Undifferentiated: CK19, ABCG2, p63, ΔNp63, Hsp70, KRT15 Differentiated: MUC5AC, claudin 10, claudin 2 | [35,36] |
| Trabecular meshwork (insert zone) | Trabecular meshwork stem cells | Multipotent-adipocytes, osteocytes and chondrocytes | - | Undifferentiated: ABCG2, Notch1, OCT-3/4, ankyrin G, mucin 1, CD73, CD90 and CD105 Differentiated: AQP1, MGP, CHI3L1, TIMP3 | [37,38] |
| Lens (anterior capsule) | Lens epithelial/stem cells | Unipotent | FGF, MAPK | Undifferentiated: PAX6, C-MAF, E-cadherin, Sox2, vimentin Differentiated: Filensin, CP49, CRYBA2 | [39,40,41] |
| Iris (epithelium) | Iris pigmented epithelium cells | They can form lentoids and neurospheres | bFGF | Undifferentiated: Nestin, Msi1 Differentiated (neural): Map2, TuJ, Gfap, O4 oligodendrocyte marker | [42,43] |
| Ciliary body (ciliary epithelium and CMZ) | Ciliary epithelium stem cells/Retinal stem cells | Possible differentiation into neurons, RGCs and photoreceptors | Wnt, FGF | Undifferentiated: Nestin, CHX 10 Differentiated RGCs: Thy1.1, Brn-3b Photoreceptors: rhodopsin Neural: HPC-1 (amacrine cells), calbindin (horizontal cells) | [44,45,46] |
| Possible Location of Endothelial Progenitors | Methods of Identification | Markers | Remarks | Ref |
|---|---|---|---|---|
| Not specific corneal endothelium | Sphere-forming assay | Nestin, GFAP, β3-tubulin, αSMA | Dissociated sphere cells showed hexagonal shape and pumping activity; no p75NTR expression. | [98] |
| Peripheral endothelium (PE) | BrdU labelling and immunostaining | Alkaline phosphatase, Telomerase | Located at the corneal endothelium/TM junction | [99] |
| Sphere-forming assay | Nil | PE had a significantly higher percentage of sphere formation, representing precursor density | [100] | |
| Immunostaining and flow cytometry | Lgr5, Hedgehog pathway markers (SHH, Gli1, Gli2) | Lgr5+ cells were proliferative. Generation of differentiated corneal endothelium and functional assay was not demonstrated | [101] | |
| Central and peripheral endothelium; progenitor-enriched at CE-TM transition region | Immunostaining and flow cytometry | p75NTR, Sox9, FoxC2 | Expressed partial properties of neural crest and periocular mesenchyme; differentiated cell sheet had pumping activity by using chamber system and in vivo transplantation to rabbit corneas. | [102] |
| Whole corneal endothelium of normal and FECD corneas | Colony-forming populations; >80 passages | Pax3, Nestin, Sox9, AP-2β, p75NTR, Sox2, Lgr5, p63, Oct4 | Adult corneal endothelium harboured neural crest-derived progenitors capable of perpetual proliferation and formation of endothelial layer exhibiting trans-endothelial resistance. | [103] |
| Trabecular meshwork (TM) | 3D Matrigel culture to activate BMP signalling | AQP1, MGP, CHI3L1, AnkG, Oct4, Sox2, Nanog, ABCG2, p75NTR, FOXD3, Sox9, Sox10, MSX1 | TM progenitors differentiated into corneal endothelial cells, adipocytes and chondrocytes. | [104] |
| TM and transition zone between TM and corneal periphery | Corneal wound model and immunostaining | Alkaline phosphatase, Nestin, Telomerase, Oct3/4, Pax6, Wnt1, Sox2 | Wounding activated Oct3/4 and Wnt1 expression as a response to initiate the endothelial repair process. | [38] |
| Transition zone (inner TZ) | Immunostaining, cell culture | Lgr5, Telomerase, Nestin, Sox2, p75NTR, Pitx2, HNK1 | Progenitors projected as multicellular clusters into the adjacent PE. Porcine TZ progenitors differentiated into an endothelial monolayer expressing ZO-1 and ATPase. | [105] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hidalgo-Alvarez, V.; Dhowre, H.S.; Kingston, O.A.; Sheridan, C.M.; Levis, H.J. Biofabrication of Artificial Stem Cell Niches in the Anterior Ocular Segment. Bioengineering 2021, 8, 135. https://doi.org/10.3390/bioengineering8100135
Hidalgo-Alvarez V, Dhowre HS, Kingston OA, Sheridan CM, Levis HJ. Biofabrication of Artificial Stem Cell Niches in the Anterior Ocular Segment. Bioengineering. 2021; 8(10):135. https://doi.org/10.3390/bioengineering8100135
Chicago/Turabian StyleHidalgo-Alvarez, Veronica, Hala S. Dhowre, Olivia A. Kingston, Carl M. Sheridan, and Hannah J. Levis. 2021. "Biofabrication of Artificial Stem Cell Niches in the Anterior Ocular Segment" Bioengineering 8, no. 10: 135. https://doi.org/10.3390/bioengineering8100135