Three Decades of Research on Recombinant Collagens: Reinventing the Wheel or Developing New Biomedical Products?


Abstract
:1. Introduction
2. Collagens: The Cornerstones of Tissue Architecture
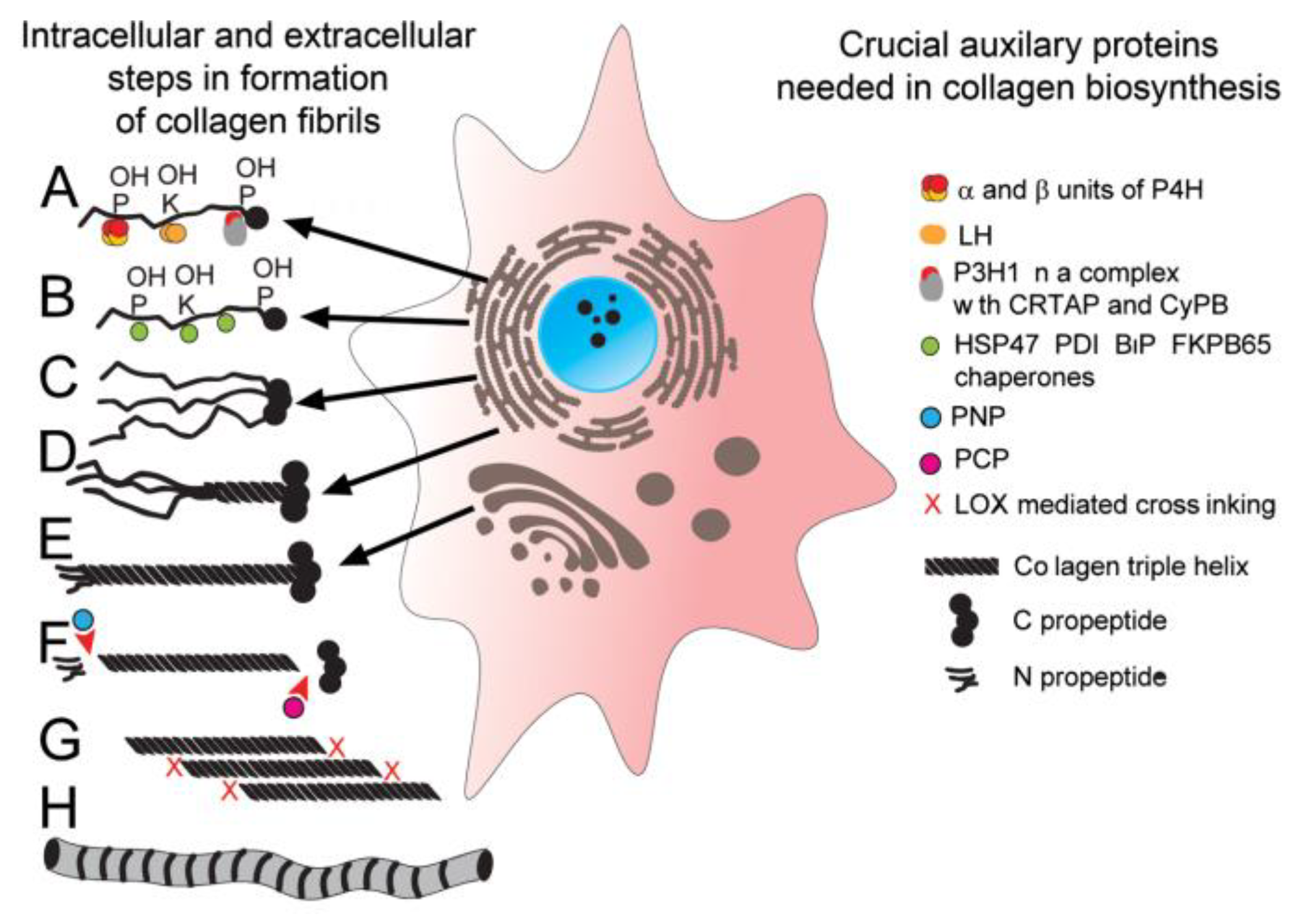
2.1. Biosynthesis of Triple-Helical Collagen Molecules
2.2. Diverse Architectures of Collagen-Rich Matrices
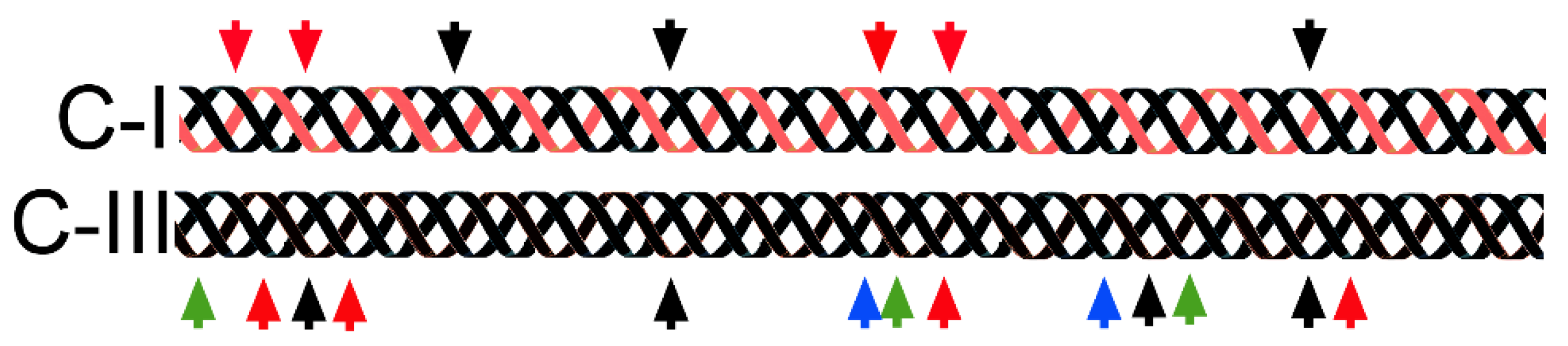
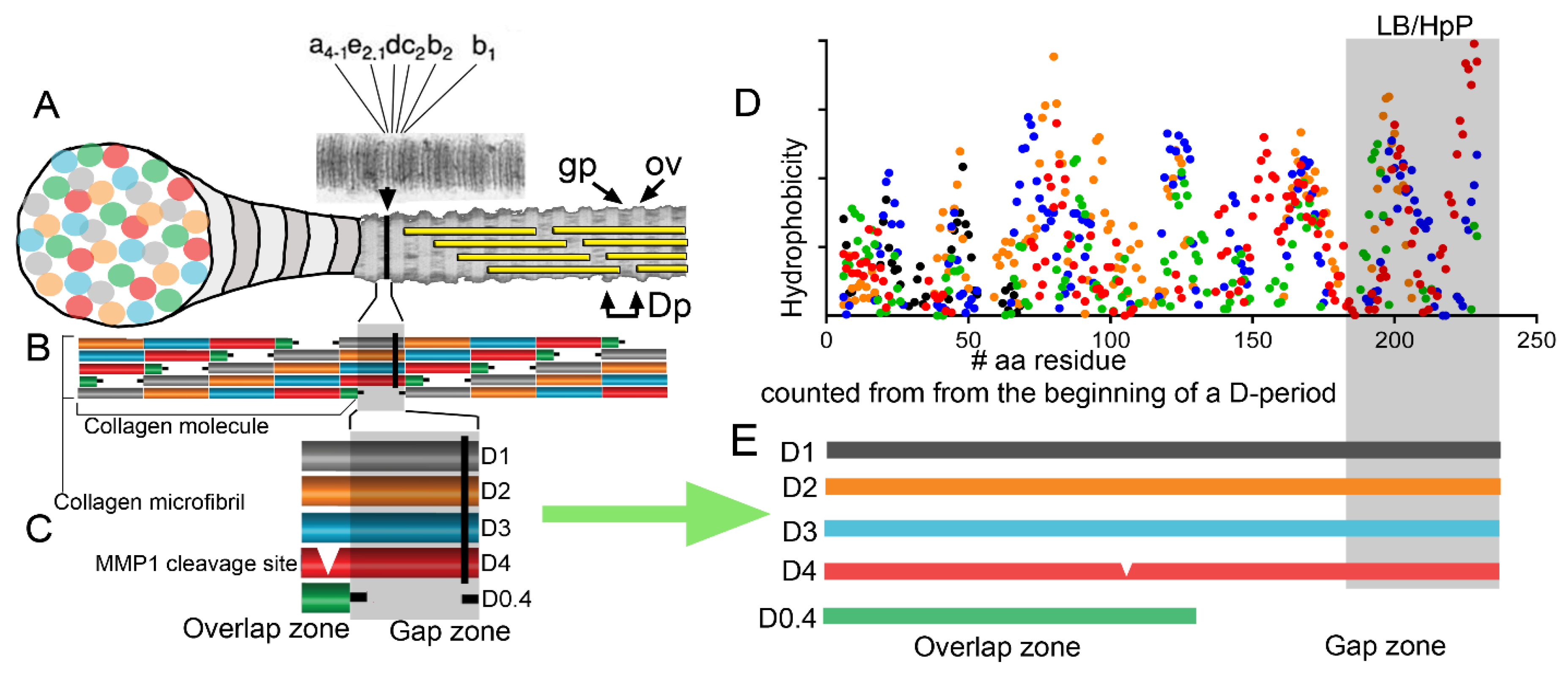
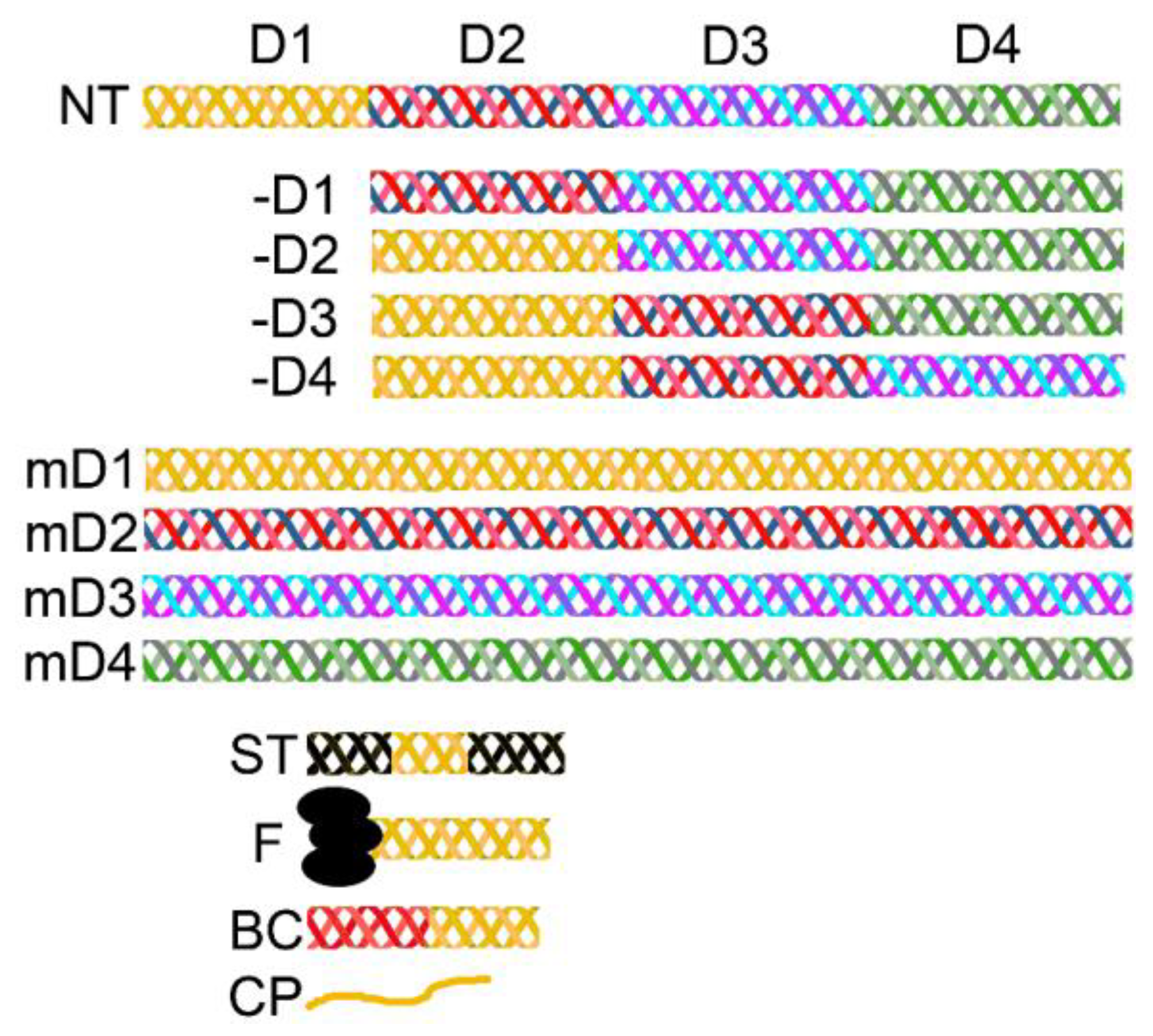
2.3. Self-Assembly of Collagens: Paradigms of Collagen I Fibrils, Collagen VII Anchoring Fibrils, and Collagen IV Networks
2.4. Collagens as Signaling Molecules
3. Practical Utilization of Collagens
3.1. Applications of Collagens
3.2. Potential Limitations of Collagens Isolated from the Natural Sources
4. Recombinant Collagens
4.1. Prerequisites for Engineering Mammalian Recombinant Collagens
4.2. Collagen Expression Systems
4.3. Recombinant Collagen Variants
4.4. Proposed Biomedical Applications of Recombinant Collagen Constructs
5. Recombinant Collagens for Protein Replacement Therapies
6. Constraints on Implementing Recombinant Collagen Technology in Clinical Applications
- There is no clear consensus on a system for large-scale recombinant collagen production that would be accepted by the regulatory agencies responsible for approving biologics for commercial clinical use. The production systems encompass different organisms, including bacteria, mammalian cells, insect cells, yeast, transgenic animals, and transgenic plants.
- There is no consensus on the most relevant form of recombinant collagens needed in the market. While some studies consider production and application of the native-like collagens, others propose to manufacture and use collagen-derived synthetic linear peptides, triple-helical fragments, and genetically engineered collagen-inspired constructs. For instance, it is not clear whether recombinant gelatin will be manufactured from the full-length recombinant collagens or selected recombinant fragments.
- Because of the wide span of potential biomedical applications of recombinant collagens, ranging from drug delivery, tissue engineering, wound healing, and protein replacement therapies, there is no identifiable leading product that could attract the attention of the market. Many different collagen types are needed for medical applications in distinct tissues and organs, amplifying this problem.
- Although some concerns about the safety of animal-derived collagen materials exist, pharmaceutical, cosmetic, and food industries continue to use them. Furthermore, because these materials are readily available from tissues of isolated animal herds, they are likely less expensive than recombinant collagens whose production requires advanced technologies.
- Even with a few companies’ early interest in producing and delivering large amounts of recombinant collagens and gelatins, no commercial products are widely available on the market. This situation may indicate that the market’s needs differed from companies’ expectations about recombinant collagens’ commercial potential.
- Tissue engineering is a crucial proposed use for recombinant collagen variants. Despite the promising preclinical results of many tissue-engineered medical products, only a few have had success in the clinic thus far [169]. Consequently, it is likely that the primary potential beneficiary of recombinant collagen technology, i.e., the tissue engineering industry, does not clamor for novel recombinant collagen-based materials in any significant way.
Funding
Acknowledgments
Conflicts of Interest
References
- Kesik-Brodacka, M. Progress in biopharmaceutical development. Biotechnol. Appl. Biochem. 2018, 65, 306–322. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.M.; Chaudhry, P.; Zo, S.M.; Han, S.S. Advances in protein-based materials: From origin to novel biomaterials. Adv. Exp. Med. Biol. 2018, 1078, 161–210. [Google Scholar]
- Girotti, A.; Orbanic, D.; Ibanez-Fonseca, A.; Gonzalez-Obeso, C.; Rodriguez-Cabello, J.C. Recombinant technology in the development of materials and systems for soft-tissue repair. Adv. Healthc. Mater. 2015, 4, 2423–2455. [Google Scholar] [CrossRef]
- Brodsky, B.; Ramshaw, J.A.M. Bioengineered collagens. In Fibrous Proteins: Structures and Mechanisms. Subcellular Biochemistry; Parry, D.A.D., Squire, J.M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; Volume 82, pp. 601–629. [Google Scholar]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [Green Version]
- Hintze, V.; Steplewski, A.; Ito, H.; Jensen, D.A.; Rodeck, U.; Fertala, A. Cells expressing partially unfolded R789C/p.R989C type II procollagen mutant associated with spondyloepiphyseal dysplasia undergo apoptosis. Hum. Mutat. 2008, 29, 841–851. [Google Scholar] [CrossRef]
- Silver, F.H.; Freeman, J.W.; Seehra, G.P. Collagen self-assembly and the development of tendon mechanical properties. J. Biomech. 2003, 36, 1529–1553. [Google Scholar] [CrossRef]
- Prockop, D.J.; Fertala, A. The collagen fibril: The almost crystalline structure. J. Struct. Biol. 1998, 122, 111–118. [Google Scholar] [CrossRef]
- Silver, D.; Miller, J.; Harrison, R.; Prockop, D.J. Helical model of nucleation and propagation to account for the growth of type I collagen fibrils from symmetrical pointed tips: A special example of self-assembly of rod-like monomers. Proc. Natl. Acad. Sci. USA 1992, 89, 9860–9864. [Google Scholar] [CrossRef] [Green Version]
- Sundaramoorthy, M.; Meiyappan, M.; Todd, P.; Hudson, B.G. Crystal structure of NC1 domains. Structural basis for type IV collagen assembly in basement membranes. J. Biol. Chem. 2002, 277, 31142–31153. [Google Scholar] [CrossRef] [Green Version]
- Kadler, K.E.; Hojima, Y.; Prockop, D.J. Assembly of collagen fibrils de novo by cleavage of the type I pC-collagen with procollagen C-proteinase. Assay of critical concentration demonstrates that collagen self-assembly is a classical example of an entropy-driven process. J. Biol. Chem. 1987, 262, 15696–15701. [Google Scholar]
- Shayegan, M.; Altindal, T.; Kiefl, E.; Forde, N.R. Intact telopeptides enhance interactions between collagens. Biophys. J. 2016, 111, 2404–2416. [Google Scholar] [CrossRef] [Green Version]
- Prockop, D.J.; Fertala, A. Inhibition of the self-assembly of collagen I into fibrils with synthetic peptides. Demonstration that assembly is driven by specific binding sites on the monomers. J. Biol. Chem. 1998, 273, 15598–15604. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yang, A.; Jia, J.; Popov, Y.V.; Schuppan, D.; You, H. Lysyl oxidase (LOX) family members: Rationale and their potential as therapeutic targets for liver fibrosis. Hepatology 2020, 72, 729–741. [Google Scholar] [CrossRef]
- Khoshnoodi, J.; Sigmundsson, K.; Cartailler, J.P.; Bondar, O.; Sundaramoorthy, M.; Hudson, B.G. Mechanism of chain selection in the assembly of collagen IV: A prominent role for the α2 chain. J. Biol. Chem. 2006, 281, 6058–6069. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Brittingham, R.J.; Klement, J.F.; Majsterek, I.; Birk, D.E.; Uitto, J.; Fertala, A. Procollagen VII self-assembly depends on site-specific interactions and is promoted by cleavage of the NC2 domain with procollagen C-proteinase. Biochemistry 2003, 42, 11434–11442. [Google Scholar] [CrossRef]
- Royce, P.M.; Steinmann, B. Connective Tissue and Its Heritable Disorders: Molecular, Genetic, and Medical Aspects, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2002; p. 1201. [Google Scholar]
- Sweeney, S.M.; Orgel, J.P.; Fertala, A.; McAuliffe, J.D.; Turner, K.R.; Di Lullo, G.A.; Chen, S.; Antipova, O.; Perumal, S.; Ala-Kokko, L.; et al. Candidate cell and matrix interaction domains on the collagen fibril, the predominant protein of vertebrates. J. Biol. Chem. 2008, 283, 21187–21197. [Google Scholar] [CrossRef] [Green Version]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.; Zhang, D.; Macedo, M.H.; Cui, W.; Sarmento, B.; Shen, G. Advanced Collagen-Based Biomaterials for Regenerative Biomedicine. Adv. Funct. Mater. 2019, 29. [Google Scholar] [CrossRef]
- Sorushanova, A.; Delgado, L.M.; Wu, Z.; Shologu, N.; Kshirsagar, A.; Raghunath, R.; Mullen, A.M.; Bayon, Y.; Pandit, A.; Raghunath, M.; et al. The collagen suprafamily: From biosynthesis to advanced biomaterial development. Adv. Mater. 2019, 31. [Google Scholar] [CrossRef] [Green Version]
- Bogdanovic, J.; Halsey, N.A.; Wood, R.A.; Hamilton, R.G. Bovine and porcine gelatin sensitivity in children sensitized to milk and meat. J. Allergy Clin. Immunol. 2009, 124, 1108–1110. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, F.M.C.; Petrovajova, D.; Hornakova, T. Viral vaccine stabilizers: Status and trends. Acta Virol. 2017, 61, 231–239. [Google Scholar] [CrossRef]
- Olsen, D.; Yang, C.; Bodo, M.; Chang, R.; Leigh, S.; Baez, J.; Carmichael, D.; Perala, M.; Hamalainen, E.R.; Jarvinen, M.; et al. Recombinant collagen and gelatin for drug delivery. Adv. Drug Deliv. Rev. 2003, 55, 1547–1567. [Google Scholar] [CrossRef]
- Brey Gil, V.S.; Brey Gil, C.S.; Goulart, G.A.C.; Oréfice, R.L. Multi-drug hybrid delivery systems with distinct release profiles based on gelatin/collagen containing vesicles derived from block copolymers. Int. J. Biol. Macromol. 2019, 139, 967–974. [Google Scholar] [CrossRef]
- Yu, J.; Lai, T.-C.; Tsai, W.-B. Fabrication of gelatin- and collagen-based hydrogels for controlled drug release. Front. Bioeng. Biotechnol. 2016, 4. [Google Scholar] [CrossRef]
- Liu, D.; Nikoo, M.; Boran, G.; Zhou, P.; Regenstein, J.M. Collagen and gelatin. Annu. Rev. Food Sci. Technol. 2015, 6, 527–557. [Google Scholar] [CrossRef]
- Avila Rodriguez, M.I.; Rodriguez Barroso, L.G.; Sanchez, M.L. Collagen: A review on its sources and potential cosmetic applications. J. Cosmet. Dermatol. 2018, 17, 20–26. [Google Scholar] [CrossRef]
- Senadheera, T.R.L.; Dave, D.; Shahidi, F. Sea cucumber derived type I collagen: A comprehensive review. Mar. Drugs 2020, 18, 471. [Google Scholar] [CrossRef]
- Sionkowska, A.; Adamiak, K.; Musial, K.; Gadomska, M. Collagen based materials in cosmetic applications: A review. Materials 2020, 13, 4217. [Google Scholar] [CrossRef]
- Davison-Kotler, E.; Marshall, W.S.; García-Gareta, E. Sources of collagen for biomaterials in skin wound healing. Bioengineering 2019, 6, 56. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.M.; Marques, A.P.; Silva, T.J.Q.L.H.; Reis, R.L. Evaluation of the potential of collagen from codfish skin as a biomaterial for biomedical applications. Mar. Drugs 2018, 16, 495. [Google Scholar] [CrossRef] [Green Version]
- Mullins, R.J.; Richards, C.; Walker, T. Allergic reactions to oral, surgical and topical bovine collagen: Anaphylactic risk for surgeons. Aust. N. Z. J. Ophthalmol. 1996, 24, 257–260. [Google Scholar] [CrossRef]
- Keefe, J.; Wauk, L.; Chu, S.; DeLustro, F. Clinical use of injectable bovine collagen: A decade of experience. Clin. Mater. 1992, 9, 155–162. [Google Scholar] [CrossRef]
- García-Domingo, M.I.; Alijotas-Reig, J.; Cisteró-Bahima, A.; Tresserra, F.; Enrique, E. Disseminated and recurrent sarcoid-like granulomatous panniculitis due to bovine collagen injection. J. Investig. Allergol. Clin. Immunol. 2000, 10, 107–109. [Google Scholar]
- Collinge, J. Prion diseases of humans and animals: Their causes and molecular basis. Annu. Rev. Neurosci. 2001, 24, 519–550. [Google Scholar] [CrossRef] [Green Version]
- Richt, J.A.; Kasinathan, P.; Hamir, A.N.; Castilla, J.; Sathiyaseelan, T.; Vargas, F.; Sathiyaseelan, J.; Wu, H.; Matsushita, H.; Koster, J.; et al. Production of cattle lacking prion protein. Nat. Biotechnol. 2007, 25, 132–138. [Google Scholar] [CrossRef]
- Lynn, A.K.; Yannas, I.V.; Bonfield, W. Antigenicity and immunogenicity of collagen. J. Biomed. Mater. Res. B 2004, 71, 343–354. [Google Scholar] [CrossRef]
- Charriere, G.; Bejot, M.; Schnitzler, L.; Ville, G.; Hartmann, D.J. Reactions to a bovine collagen implant. Clinical and immunologic study in 705 patients. J. Am. Acad. Dermatol. 1989, 21, 1203–1208. [Google Scholar] [CrossRef]
- Eaglstein, W.H.; Alvarez, O.M.; Auletta, M.; Leffel, D.; Rogers, G.S.; Zitelli, J.A.; Norris, J.E.C.; Thomas, I.; Irondo, M.; Fewkes, J.; et al. Acute excisional wounds treated with a tissue-engineered skin (Apligraf). Dermatol. Surg. 1999, 25, 195–201. [Google Scholar] [CrossRef]
- Requena, L.; Requena, C.; Christensen, L.; Zimmermann, U.S.; Kutzner, H.; Cerroni, L. Adverse reactions to injectable soft tissue fillers. J. Am. Acad. Dermatol. 2011, 64, 1–34. [Google Scholar] [CrossRef]
- Zeng, W.; Osterman, M.; Stern, P.J. Inflammatory reactions to xenogenic nerve wraps: A report of three cases. J. Bone Jt. Surg. Am. 2019, 9, e0302. [Google Scholar] [CrossRef]
- Budinger, L.; Hertl, M. Immunologic mechanisms in hypersensitivity reactions to metal ions: An overview. Allergy 2000, 55, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Flier, M.; Burg, C.; Pos, O.; Plassche-Boers, E.; Bruynzeel, D.P.; Garotta, G.; Scheper, R.J. In vitro studies in nickel allergy: Diagnostic value of a dual parameter analysis. J. Investig. Dermatol. 1987, 88, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Meyer, K.K.; Beezhold, D.H. Latex allergy: How safe are your gloves? Bull. Am. Coll. Surg. 1997, 82, 72. [Google Scholar]
- Giordano, P.; Sileri, P.; Buntzen, S.; Stuto, A.; Nunoo-Mensah, J.; Lenisa, L.; Singh, B.; Thorlacius-Ussing, O.; Griffiths, B.; Ziyaie, D. Final results of a European, multi-centre, prospective, observational study of Permacol™ collagen paste injection for the treatment of anal fistula. Colorectal. Dis. 2018, 20, 243–251. [Google Scholar] [CrossRef]
- Miyoshi, M.; Liu, S. Collagen-induced arthritis models. Methods Mol. Biol. 2018, 1868, 3–7. [Google Scholar]
- Fertala, A.; Shah, M.; Hoffman, R.; Arnold, W.V. Designing recombinant collagens for biomedical applications. Curr. Tissue Eng. 2016, 5, 73–84. [Google Scholar] [CrossRef]
- Schnieke, A.; Dziadek, M.; Bateman, J.; Mascara, T.; Harbers, K.; Gelinas, R.; Jaenisch, R. Introduction of the human pro α1(I) collagen gene into pro α1(I)-deficient Mov-13 mouse cells leads to formation of functional mouse-human hybrid type I collagen. Proc. Natl. Acad. Sci. USA 1987, 84, 764–768. [Google Scholar] [CrossRef] [Green Version]
- Olsen, A.S.; Geddis, A.E.; Prockop, D.J. High levels of expression of a minigene version of the human pro α1 (I) collagen gene in stably transfected mouse fibroblasts. Effects of deleting putative regulatory sequences in the first intron. J. Biol. Chem. 1991, 266, 1117–1121. [Google Scholar]
- Specks, U.; Mayer, U.; Nischt, R.; Spissinger, T.; Mann, K.; Timpl, R.; Engel, J.; Chu, M.L. Structure of recombinant N-terminal globule of type VI collagen α3 chain and its binding to heparin and hyaluronan. EMBO J. 1992, 11, 4281–4290. [Google Scholar] [CrossRef]
- Mazzorana, M.; Gruffat, H.; Sergeant, A.; Rest, M. Mechanisms of collagen trimer formation. Construction and expression of a recombinant minigene in HeLa cells reveals a direct effect of prolyl hydroxylation on chain assembly of type XII collagen. J. Biol. Chem. 1993, 268, 3029–3032. [Google Scholar]
- Fertala, A.; Sieron, A.L.; Ganguly, A.; Li, S.W.; Ala-Kokko, L.; Anumula, K.R.; Prockop, D.J. Synthesis of recombinant human procollagen II in a stably transfected tumour cell line (HT1080). Biochem. J. 1994, 298, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulleid, N.J.; John, D.C.; Kadler, K.E. Recombinant expression systems for the production of collagen. Biochem. Soc. Trans. 2000, 28, 350–353. [Google Scholar] [CrossRef]
- Lukomski, S.; Nakashima, K.; Abdi, I.; Cipriano, V.J.; Ireland, R.M.; Reid, S.D.; Adams, G.G.; Musser, J.M. Identification and characterization of the scl gene encoding a group A Streptococcus extracellular protein virulence factor with similarity to human collagen. Infect. Immun. 2000, 68, 6542–6553. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, T.D.; Peng, Y.Y.; Trueman, H.E.; Weisman, S.; Okada, S.; Walker, A.A.; Sriskantha, A.; White, J.F.; Huson, M.G.; Werkmeister, J.A.; et al. A new class of animal collagen masquerading as an insect silk. Sci. Rep. 2013, 3, 2864. [Google Scholar] [CrossRef] [Green Version]
- Fertala, A.; Sieron, A.L.; Hojima, Y.; Ganguly, A.; Prockop, D.J. Self-assembly into fibrils of collagen II by enzymic cleavage of recombinant procollagen II. Lag period, critical concentration, and morphology of fibrils differ from collagen I. J. Biol. Chem. 1994, 269, 11584–11589. [Google Scholar]
- Ruggiero, F.; Exposito, J.Y.; Bournat, P.; Gruber, V.; Perret, S.; Comte, J.; Olagnier, B.; Garrone, R.; Theisen, M. Triple helix assembly and processing of human collagen produced in transgenic tobacco plants. FEBS Lett. 2000, 469, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Gan, Q.; Clough, R.C.; Pappu, K.M.; Howard, J.A.; Baez, J.A.; Wang, K. Hydroxylation of recombinant human collagen type I α1 in transgenic maize co-expressed with a recombinant human prolyl 4-hydroxylase. BMC Biotechnol. 2011, 11, 69. [Google Scholar] [CrossRef] [Green Version]
- Gellermann, P.; Schneider-Barthold, C.; Bolten, S.N.; Overfelt, E.; Scheper, T.; Pepelanova, I. Production of a recombinant non-hydroxylated gelatin mimetic in Pichia pastoris for biomedical applications. J. Funct. Biomater. 2019, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Rutschmann, C.; Baumann, S.; Cabalzar, J.; Luther, K.B.; Hennet, T. Recombinant expression of hydroxylated human collagen in Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 4445–4455. [Google Scholar] [CrossRef] [Green Version]
- John, D.C.; Watson, R.; Kind, A.J.; Scott, A.R.; Kadler, K.E.; Bulleid, N.J. Expression of an engineered form of recombinant procollagen in mouse milk. Nat. Biotechnol. 1999, 17, 385–389. [Google Scholar] [CrossRef]
- Berg, R.A. Human Recombinant Collagen in the Milk of Transgenic Animals. Application Number 183,648. United States Patent US 5,667,839, 16 September 1997. [Google Scholar]
- Harvey, A.J.; Rapp, J.C. Method of Using a Transgenic Chicken to Produce Exogenous Proteins in Its Eggs. Application Number 15/181,987. United States Patent US 10,182,561, 22 January 2019. [Google Scholar]
- Brodsky, B.; Ramshaw, J.A. Bioengineered collagens. Subcell. Biochem. 2017, 82, 601–629. [Google Scholar]
- Jacobsen, M.K.; Andresen, A.D.K.; Jespersen, A.B.; Støttrup, C.; Carreon, L.Y.; Overgaard, S.; Andersen, M. Randomized double blind clinical trial of ABM/P-15 versus allograft in noninstrumented lumbar fusion surgery. Spine J. 2020, 20, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Mohanram, Y.; Zhang, J.; Tsiridis, E.; Yang, X.B. Comparing bone tissue engineering efficacy of HDPSCs, HBMSCs on 3D biomimetic ABM-P-15 scaffolds in vitro and in vivo. Cytotechnology 2020, 72, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Weber, S.C.; Herz, A.H. Method for Recombinant Yeast Expression and Isolation of Water-Soluble Collagen-Type Polypeptides. Application Number 383,748. United States Patent US 5,710,252, 20 January 1998. [Google Scholar]
- Mashiko, T.; Takada, H.; Wu, S.H.; Kanayama, K.; Feng, J.; Tashiro, K.; Asahi, R.; Sunaga, A.; Hoshi, K.; Kurisaki, A.; et al. Therapeutic effects of a recombinant human collagen peptide bioscaffold with human adipose-derived stem cells on impaired wound healing after radiotherapy. J. Tissue Eng. Regen. Med. 2018, 12, 1186–1194. [Google Scholar] [CrossRef]
- Shi, J.; Ma, X.; Gao, Y.; Fan, D.; Zhu, C.; Mi, Y.; Xue, W. Hydroxylation of human type III collagen α chain by recombinant coexpression with a viral prolyl 4-hydroxylase in Escherichia coli. Protein J. 2017, 36, 322–331. [Google Scholar] [CrossRef]
- Vuorela, A.; Myllyharju, J.; Nissi, R.; Pihlajaniemi, T.; Kivirikko, K.I. Assembly of human prolyl 4-hydroxylase and type III collagen in the yeast Pichia pastoris: Formation of a stable enzyme tetramer requires coexpression with collagen and assembly of a stable collagen requires coexpression with prolyl 4-hydroxylase. EMBO J. 1997, 16, 6702–6712. [Google Scholar] [CrossRef] [Green Version]
- Baez, J.; Olsen, D.; Polarek, J.W. Recombinant microbial systems for the production of human collagen and gelatin. Appl. Microbiol. Biotechnol. 2005, 69, 245–252. [Google Scholar] [CrossRef]
- Stein, H.; Wilensky, M.; Tsafrir, Y.; Rosenthal, M.; Amir, R.; Avraham, T.; Ofir, K.; Dgany, O.; Yayon, A.; Shoseyov, O. Production of bioactive, post-translationally modified, heterotrimeric, human recombinant type-I collagen in transgenic tobacco. Biomacromolecules 2009, 10, 2640–2645. [Google Scholar] [CrossRef]
- McTiernan, C.D.; Simpson, F.C.; Haagdorens, M.; Samarawickrama, C.; Hunter, D.; Buznyk, O.; Fagerholm, P.; Ljunggren, M.K.; Lewis, P.; Pintelon, I.; et al. LiQD Cornea: Pro-regeneration collagen mimetics as patches and alternatives to corneal transplantation. Sci. Adv. 2020, 6. [Google Scholar] [CrossRef]
- McLaughlin, S.; McNeill, B.; Podrebarac, J.; Hosoyama, K.; Sedlakova, V.; Cron, G.; Smyth, D.; Seymour, R.; Goel, K.; Liang, W.; et al. Injectable human recombinant collagen matrices limit adverse remodeling and improve cardiac function after myocardial infarction. Nat. Commun. 2019, 10, 4866. [Google Scholar] [CrossRef] [Green Version]
- Farkash, U.; Avisar, E.; Volk, I.; Slevin, O.; Shohat, N.; El Haj, M.; Dolev, E.; Ashraf, E.; Luria, S. First clinical experience with a new injectable recombinant human collagen scaffold combined with autologous platelet-rich plasma for the treatment of lateral epicondylar tendinopathy (tennis elbow). J. Shoulder Elbow. Surg. 2019, 28, 503–509. [Google Scholar] [CrossRef]
- Ritala, A.; Wahlstrom, E.H.; Holkeri, H.; Hafren, A.; Makelainen, K.; Baez, J.; Makinen, K.; Nuutila, A.M. Production of a recombinant industrial protein using barley cell cultures. Protein Expr. Purif. 2008, 59, 274–281. [Google Scholar] [CrossRef]
- Steplewski, A.; Hintze, V.; Fertala, A. Molecular basis of organization of collagen fibrils. J. Struct. Biol. 2007, 157, 297–307. [Google Scholar] [CrossRef]
- Perumal, S.; Antipova, O.; Orgel, J.P. Collagen fibril architecture, domain organization, and triple-helical conformation govern its proteolysis. Proc. Natl. Acad. Sci. USA 2008, 105, 2824–2829. [Google Scholar] [CrossRef] [Green Version]
- Haywood, J.; Qi, J.; Chen, C.C.; Lu, G.; Liu, Y.; Yan, J.; Shi, Y.; Gao, G.F. Structural basis of collagen recognition by human osteoclast-associated receptor and design of osteoclastogenesis inhibitors. Proc. Natl. Acad. Sci. USA 2016, 113, 1038–1043. [Google Scholar] [CrossRef] [Green Version]
- Boraschi-Diaz, I.; Wang, J.; Mort, J.S.; Komarova, S.V. Collagen type I as a ligand for receptor-mediated signaling. Front. Phys. 2017, 5. [Google Scholar] [CrossRef]
- Zhu, J.; Madhurapantula, R.S.; Kalyanasundaram, A.; Sabharwal, T.; Antipova, O.; Bishnoi, S.W.; Orgel, J. Ultrastructural location and interactions of the immunoglobulin receptor binding sequence within fibrillar type I collagen. Int. J. Mol. Sci. 2020, 21, 4166. [Google Scholar] [CrossRef]
- Brittingham, R.; Colombo, M.; Ito, H.; Steplewski, A.; Birk, D.E.; Uitto, J.; Fertala, A. Single amino acid substitutions in procollagen vii affect early stages of assembly of anchoring fibrils. J. Biol. Chem. 2005, 280, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Brittingham, R.; Uitto, J.; Fertala, A. High-affinity binding of the NC1 domain of collagen VII to laminin 5 and collagen IV. Biochem. Biophys. Res. Commun. 2006, 343, 692–699. [Google Scholar] [CrossRef] [Green Version]
- Geddis, A.E.; Prockop, D.J. Expression of human COL1A1 gene in stably transfected HT1080 cells: The production of a thermostable homotrimer of type I collagen in a recombinant system. Matrix 1993, 13, 399–405. [Google Scholar] [CrossRef]
- Arnold, W.V.; Bachinger, H.P.; Sieron, A.; Fertala, A.; Mechling, D.; Prockop, D.J. Circular dichroism analysis of novel recombinant type II procollagens. Matrix Biol. 1996, 15, 153. [Google Scholar] [CrossRef]
- Colombatti, A.; Mucignat, M.T.; Bonaldo, P. Secretion and matrix assembly of recombinant type VI collagen. J. Biol. Chem. 1995, 270, 13105–13111. [Google Scholar] [CrossRef] [Green Version]
- Marinkovich, M.P.; Lane, A.T.; Rajadas, J. Production and Delivery of a Stable Collagen. Application Number 14/112,444, 26 May 2015. [Google Scholar]
- Myllyharju, J.; Lamberg, A.; Notbohm, H.; Fietzek, P.P.; Pihlajaniemi, T.; Kivirikko, K.I. Expression of wild-type and modified proα chains of human type I procollagen in insect cells leads to the formation of stable [α1(I)]2α2(I) collagen heterotrimers and [α1(I)]3 homotrimers but not [α2(I)]3 homotrimers. J. Biol. Chem. 1997, 272, 21824–21830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamberg, A.; Helaakoski, T.; Myllyharju, J.; Peltonen, S.; Notbohm, H.; Pihlajaniemi, T.; Kivirikko, K.I. Characterization of human type III collagen expressed in a baculovirus system. Production of a protein with a stable triple helix requires coexpression with the two types of recombinant prolyl 4-hydroxylase subunit. J. Biol. Chem. 1996, 271, 11988–11995. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.Y.; Yoshizumi, A.; Danon, S.J.; Glattauer, V.; Prokopenko, O.; Mirochnitchenko, O.; Yu, Z.; Inouye, M.; Werkmeister, J.A.; Brodsky, B.; et al. A Streptococcus pyogenes derived collagen-like protein as a non-cytotoxic and non-immunogenic cross-linkable biomaterial. Biomaterials 2010, 31, 2755–2761. [Google Scholar] [CrossRef] [Green Version]
- Ramshaw, J.A.; Werkmeister, J.A.; Dumsday, G.J. Bioengineered collagens: Emerging directions for biomedical materials. Bioengineered 2014, 5, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Nokelainen, M.; Tu, H.; Vuorela, A.; Notbohm, H.; Kivirikko, K.I.; Myllyharju, J. High-level production of human type I collagen in the yeast Pichia Pastoris. Yeast 2001, 18, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Baez, J.; Pappu, K.M.; Glatz, C.E. Purification and characterization of a transgenic corn grain-derived recombinant collagen type I α1. Biotechnol. Prog. 2009, 25, 1660–1668. [Google Scholar]
- Shoseyov, O.; Posen, Y.; Grynspan, F. Human collagen produced in plants: More than just another molecule. Bioengineered 2014, 5, 49–52. [Google Scholar] [CrossRef] [Green Version]
- Shilo, S.; Roth, S.; Amzel, T.; Harel-Adar, T.; Tamir, E.; Grynspan, F.; Shoseyov, O. Cutaneous wound healing after treatment with plant-derived human recombinant collagen flowable gel. Tissue Eng. Part A 2013, 19, 1519–1526. [Google Scholar] [CrossRef]
- Pihlajamaa, T.; Perala, M.; Vuoristo, M.M.; Nokelainen, M.; Bodo, M.; Schulthess, T.; Vuorio, E.; Timpl, R.; Engel, J.; Ala-Kokko, L. Characterization of recombinant human type IX collagen. Association of α chains into homotrimeric and heterotrimeric molecules. J. Biol. Chem. 1999, 274, 22464–22468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Raynal, N.; Stathopoulos, S.; Myllyharju, J.; Farndale, R.W.; Leitinger, B. Collagen binding specificity of the discoidin domain receptors: Binding sites on collagens II and III and molecular determinants for collagen IV recognition by DDR1. Matrix Biol. 2011, 30, 16–26. [Google Scholar] [CrossRef]
- He, Y.; Hou, Z.; Wang, J.; Wang, Z.; Li, X.; Liu, J.; Yang, X.; Liang, Q.; Zhao, J. Assessment of biological properties of recombinant collagen-hyaluronic acid composite scaffolds. Int. J. Biol. Macromol. 2020, 149, 1275–1284. [Google Scholar] [CrossRef]
- Tytgat, L.; Markovic, M.; Qazi, T.H.; Vagenende, M.; Bray, F.; Martins, J.C.; Rolando, C.; Thienpont, H.; Ottevaere, H.; Ovsianikov, A.; et al. Photo-crosslinkable recombinant collagen mimics for tissue engineering applications. J. Mater. Chem. B 2019, 7, 3100–3108. [Google Scholar] [CrossRef] [Green Version]
- An, B.; Lin, Y.S.; Brodsky, B. Collagen interactions: Drug design and delivery. Adv. Drug Deliv. Rev. 2016, 97, 69–84. [Google Scholar] [CrossRef]
- An, B.; Abbonante, V.; Yigit, S.; Balduini, A.; Kaplan, D.L.; Brodsky, B. Definition of the native and denatured type II collagen binding site for fibronectin using a recombinant collagen system. J. Biol. Chem. 2014, 289, 4941–4951. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, T.D.; Huson, M.G.; Rapson, T.D. Rational design of new materials using recombinant structural proteins: Current state and future challenges. J. Struct. Biol. 2018, 201, 76–83. [Google Scholar] [CrossRef]
- Arnold, W.V.; Fertala, A.; Sieron, A.L.; Hattori, H.; Mechling, D.; Bachinger, H.P.; Prockop, D.J. Recombinant procollagen II: Deletion of D period segments identifies sequences that are required for helix stabilization and generates a temperature-sensitive N-proteinase cleavage site. J. Biol. Chem. 1998, 273, 31822–31828. [Google Scholar] [CrossRef] [Green Version]
- Fertala, A.; Han, W.B.; Ko, F.K. Mapping critical sites in collagen II for rational design of gene- engineered proteins for cell-supporting materials. J. Biomed. Mater. Res. 2001, 57, 48–58. [Google Scholar] [CrossRef]
- Sieron, A.L.; Louneva, N.; Fertala, A. Site-specific interaction of bone morphogenetic protein 2 with procollagen II. Cytokine 2002, 18, 214–221. [Google Scholar] [CrossRef]
- Majsterek, I.; McAdams, E.; Adachi, E.; Dhume, S.T.; Fertala, A. Prospects and limitations of the rational engineering of fibrillar collagens. Protein Sci. 2003, 12, 2063–2072. [Google Scholar] [CrossRef] [Green Version]
- Leitinger, B.; Steplewski, A.; Fertala, A. The D2 period of collagen II contains a specific binding site for the human discoidin domain receptor, DDR2. J. Mol. Biol. 2004, 344, 993–1003. [Google Scholar] [CrossRef]
- Ito, H.; Steplewski, A.; Alabyeva, T.; Fertala, A. Testing the utility of rationally engineered recombinant collagen-like proteins for applications in tissue engineering. J. Biomed. Mater. Res. Part A 2006, 76, 551–560. [Google Scholar] [CrossRef]
- Arnold, P.M.; Sasso, R.C.; Janssen, M.E.; Fehlings, M.G.; Heary, R.F.; Vaccaro, A.R.; Kopjar, B. i-Factor™ bone graft vs autograft in anterior cervical discectomy and fusion: 2-year follow-up of the randomized single-blinded food and drug administration investigational device exemption study. Neurosurgery 2018, 83, 377–384. [Google Scholar] [CrossRef]
- Axelsen, M.G.; Overgaard, S.; Jespersen, S.M.; Ding, M. Comparison of synthetic bone graft ABM/P-15 and allograft on uninstrumented posterior lumbar spine fusion in sheep. J. Orthop. Surg. Res. 2019, 14, 2. [Google Scholar] [CrossRef]
- Bhatnagar, R.S.; Qian, J.J.; Wedrychowska, A.; Sadeghi, M.; Wu, Y.M.; Smith, N. Design of biomimetic habitats for tissue engineering with P-15, a synthetic peptide analogue of collagen. Tissue Eng. 1999, 5, 53–65. [Google Scholar] [CrossRef]
- Umeyama, R.; Yamawaki, T.; Liu, D.; Kanazawa, S.; Takato, T.; Hoshi, K.; Hikita, A. Optimization of culture duration of bone marrow cells before transplantation with a β-tricalcium phosphate/recombinant collagen peptide hybrid scaffold. Regen. Ther. 2020, 14, 284–295. [Google Scholar] [CrossRef]
- Nakamura, K. CellSaic, a cell aggregate-like technology using recombinant peptide pieces for MSC transplantation. Curr. Stem. Cell Res. Ther. 2019, 14, 52–56. [Google Scholar] [CrossRef]
- Confalonieri, D.; La Marca, M.; van Dongen, E.; Walles, H.; Ehlicke, F. An injectable recombinant collagen I peptide-based macroporous microcarrier allows superior expansion of C2C12 and human bone marrow-derived mesenchymal stromal cells and supports deposition of mineralized matrix. Tissue Eng. Part A 2017, 23, 946–957. [Google Scholar] [CrossRef]
- Karimi, F.; McKenzie, T.G.; O’Connor, A.J.; Qiao, G.G.; Heath, D.E. Nano-scale clustering of integrin-binding ligands regulates endothelial cell adhesion, migration, and endothelialization rate: Novel materials for small diameter vascular graft applications. J. Mater. Chem. B 2017, 5, 5942–5953. [Google Scholar] [CrossRef]
- Kapp, T.G.; Rechenmacher, F.; Neubauer, S.; Maltsev, O.V.; Cavalcanti-Adam, E.A.; Zarka, R.; Reuning, U.; Notni, J.; Wester, H.J.; Mas-Moruno, C.; et al. A Comprehensive Evaluation of the activity and selectivity profile of ligands for RGD-binding integrins. Sci. Rep. 2017, 7, 39805. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Yu, Y.; Wang, Q.; Xi, Y.; Liu, Y. Interaction mechanism and clustering among RGD peptides and integrins. Mol. Inform. 2017, 36. [Google Scholar] [CrossRef] [PubMed]
- Persikov, A.V.; Ramshaw, J.A.; Kirkpatrick, A.; Brodsky, B. Triple-helix propensity of hydroxyproline and fluoroproline: Comparison of host-guest and repeating tripeptide collagen models. J. Am. Chem. Soc. 2003, 125, 11500–11501. [Google Scholar] [CrossRef]
- Brodsky, B.; Thiagarajan, G.; Madhan, B.; Kar, K. Triple-helical peptides: An approach to collagen conformation, stability, and self-association. Biopolymers 2008, 89, 345–353. [Google Scholar] [CrossRef]
- Kubyshkin, V. Stabilization of the triple helix in collagen mimicking peptides. Org. Biomol. Chem. 2019, 17, 8031–8047. [Google Scholar] [CrossRef]
- Delsuc, N.; Uchinomiya, S.; Ojida, A.; Hamachi, I. A host-guest system based on collagen-like triple-helix hybridization. Chem. Commun. (Camb.) 2017, 53, 6856–6859. [Google Scholar] [CrossRef] [Green Version]
- Setina, C.M.; Haase, J.P.; Glatz, C.E. Process integration for recovery of recombinant collagen type I α1 from corn seed. Biotechnol. Prog. 2016, 32, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Du, C.; Wang, M.; Liu, J.; Pan, M.; Cai, Y.; Yao, J. Improvement of thermostability of recombinant collagen-like protein by incorporating a foldon sequence. Appl. Microbiol. Biotechnol. 2008, 79, 195–202. [Google Scholar] [CrossRef]
- Humtsoe, J.O.; Kim, J.K.; Xu, Y.; Keene, D.R.; Hook, M.; Lukomski, S.; Wary, K.K. A streptococcal collagen-like protein interacts with the α2β1 integrin and induces intracellular signaling. J. Biol. Chem. 2005, 280, 13848–13857. [Google Scholar] [CrossRef] [Green Version]
- An, B.; Kaplan, D.L.; Brodsky, B. Engineered recombinant bacterial collagen as an alternative collagen-based biomaterial for tissue engineering. Front. Chem. 2014, 2, 40. [Google Scholar] [CrossRef] [Green Version]
- Yoshizumi, A.; Yu, Z.; Silva, T.; Thiagarajan, G.; Ramshaw, J.A.; Inouye, M.; Brodsky, B. Self-association of streptococcus pyogenes collagen-like constructs into higher order structures. Protein Sci. 2009, 18, 1241–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; An, B.; Ramshaw, J.A.; Brodsky, B. Bacterial collagen-like proteins that form triple-helical structures. J. Struct. Biol. 2014, 186, 451–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Que, R.A.; Arulmoli, J.; Da Silva, N.A.; Flanagan, L.A.; Wang, S.W. Recombinant collagen scaffolds as substrates for human neural stem/progenitor cells. J. Biomed. Mater. Res. Part A 2018, 106, 1363–1372. [Google Scholar] [CrossRef]
- Deng, A.; Yang, Y.; Du, S.; Yang, S. Electrospinning of in situ crosslinked recombinant human collagen peptide/chitosan nanofibers for wound healing. Biomater. Sci. 2018, 6, 2197–2208. [Google Scholar] [CrossRef]
- Builles, N.; Janin-Manificat, H.; Malbouyres, M.; Justin, V.; Rovere, M.R.; Pellegrini, G.; Torbet, J.; Hulmes, D.J.; Burillon, C.; Damour, O.; et al. Use of magnetically oriented orthogonal collagen scaffolds for hemi-corneal reconstruction and regeneration. Biomaterials 2010, 31, 8313–8322. [Google Scholar] [CrossRef]
- Tytgat, L.; Dobos, A.; Markovic, M.; Van Damme, L.; Van Hoorick, J.; Bray, F.; Thienpont, H.; Ottevaere, H.; Dubruel, P.; Ovsianikov, A.; et al. High-resolution 3D bioprinting of photo-cross-linkable recombinant collagen to serve tissue engineering applications. Biomacromolecules 2020, 21, 3997–4007. [Google Scholar] [CrossRef]
- Hu, K.; Hu, M.; Xiao, Y.; Cui, Y.; Yan, J.; Yang, G.; Zhang, F.; Lin, G.; Yi, H.; Han, L.; et al. Preparation recombination human-like collagen/fibroin scaffold and promoting the cell compatibility with osteoblasts. J. Biomed. Mater. Res. Part A 2020. [Google Scholar] [CrossRef]
- Wiser, I.; Tamir, E.; Kaufman, H.; Keren, E.; Avshalom, S.; Klein, D.; Heller, L.; Shapira, E. A novel recombinant human collagen-based flowable matrix for chronic lower limb wound management: First results of a clinical trial. Wounds 2019, 31, 103–107. [Google Scholar]
- IVS. Excipients in Vaccines. Available online: http://www.vaccinesafety.edu/components-Excipients.htm (accessed on 4 September 2020).
- Yang, C.; Hillas, P.; Tang, J.; Balan, J.; Notbohm, H.; Polarek, J. Development of a recombinant human collagen-type III based hemostat. J. Biomed. Mater. Res. Part B Appl. Biomater. 2004, 69, 18–24. [Google Scholar] [CrossRef]
- Fagerholm, P.; Lagali, N.S.; Ong, J.A.; Merrett, K.; Jackson, W.B.; Polarek, J.W.; Suuronen, E.J.; Liu, Y.; Brunette, I.; Griffith, M. Stable corneal regeneration four years after implantation of a cell-free recombinant human collagen scaffold. Biomaterials 2014, 35, 2420–2427. [Google Scholar] [CrossRef] [Green Version]
- Fagerholm, P.; Lagali, N.S.; Merrett, K.; Jackson, W.B.; Munger, R.; Liu, Y.; Polarek, J.W.; Soderqvist, M.; Griffith, M. A biosynthetic alternative to human donor tissue for inducing corneal regeneration: 24-month follow-up of a phase 1 clinical study. Sci. Transl. Med. 2010, 2, 46ra61. [Google Scholar] [CrossRef] [Green Version]
- Que, R.; Mohraz, A.; Da Silva, N.A.; Wang, S.W. Expanding functionality of recombinant human collagen through engineered non-native cysteines. Biomacromolecules 2014, 15, 3540–3549. [Google Scholar] [CrossRef] [Green Version]
- Uematsu, S.S.; Inagaki, A.; Nakamura, Y.; Imura, T.; Igarashi, Y.; Fathi, I.; Miyagi, S.; Ohuchi, N.; Satomi, S.; Goto, M. The optimization of the prevascularization procedures for improving subcutaneous islet engraftment. Transplantation 2018, 102, 387–395. [Google Scholar] [CrossRef]
- Tsukioka, T.; Hiratsuka, T.; Nakamura, M.; Watanabe, T.; Kitamura, Y.; Isobe, K.; Okudera, T.; Okudera, H.; Azuma, A.; Uematsu, K.; et al. An on-site preparable, novel bone-grafting complex consisting of human platelet-rich fibrin and porous particles made of a recombinant collagen-like protein. J. Biomed. Mater. Res. Part B Appl. Biomater. 2019, 107, 1420–1430. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Rodriguez, G.B.; Montesi, M.; Panseri, S.; Sprio, S.; Tampieri, A.; Sandri, M. Biomineralized recombinant collagen-based scaffold mimicking native bone enhances mesenchymal stem cell interaction and differentiation. Tissue Eng. Part A 2017, 23, 1423–1435. [Google Scholar] [CrossRef]
- Pawelec, K.M.; Confalonieri, D.; Ehlicke, F.; van Boxtel, H.A.; Walles, H.; Kluijtmans, S. Osteogenesis and mineralization of mesenchymal stem cells in collagen type I-based recombinant peptide scaffolds. J. Biomed. Mater. Res. Part A 2017, 105, 1856–1866. [Google Scholar] [CrossRef]
- Ramirez-Rodriguez, G.B.; Delgado-Lopez, J.M.; Iafisco, M.; Montesi, M.; Sandri, M.; Sprio, S.; Tampieri, A. Biomimetic mineralization of recombinant collagen type I derived protein to obtain hybrid matrices for bone regeneration. J. Struct. Biol. 2016, 196, 138–146. [Google Scholar] [CrossRef]
- Muhonen, V.; Narcisi, R.; Nystedt, J.; Korhonen, M.; van Osch, G.J.; Kiviranta, I. Recombinant human type II collagen hydrogel provides a xeno-free 3D micro-environment for chondrogenesis of human bone marrow-derived mesenchymal stromal cells. J. Tissue Eng. Regen. Med. 2017, 11, 843–854. [Google Scholar] [CrossRef]
- Ilamaran, M.; Janeena, A.; Valappil, S.; Ramudu, K.N.; Shanmugam, G.; Niraikulam, A. A self-assembly and higher order structure forming triple helical protein as a novel biomaterial for cell proliferation. Biomater. Sci. 2019, 7, 2191–2199. [Google Scholar] [CrossRef]
- An, B.; Abbonante, V.; Xu, H.; Gavriilidou, D.; Yoshizumi, A.; Bihan, D.; Farndale, R.W.; Kaplan, D.L.; Balduini, A.; Leitinger, B.; et al. Recombinant collagen engineered to bind to discoidin domain receptor functions as a receptor inhibitor. J. Biol. Chem. 2016, 291, 4343–4355. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhang, Z.; Ma, X.; Duan, Z.; Hui, J.; Zhu, C.; Zhang, D.; Fan, D.; Shang, L.; Chen, F. Newly designed human-like collagen to maximize sensitive release of BMP-2 for remarkable repairing of bone defects. Biomolecules 2019, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Golser, A.V.; Röber, M.; Börner, H.G.; Scheibel, T. Engineered collagen: A redox switchable framework for tunable assembly and fabrication of biocompatible surfaces. ACS Biomater. Sci. Eng. 2017, 4, 2106–2114. [Google Scholar] [CrossRef]
- Xu, R.; Li, D.; Peng, J.; Fang, J.; Zhang, L.; Liu, L. Cloning, expression and antioxidant activity of a novel collagen from Pelodiscus sinensis. World J. Microbiol. Biotechnol. 2016, 32, 100. [Google Scholar] [CrossRef]
- You, S.; Liu, S.; Dong, X.; Li, H.; Zhu, Y.; Hu, L. Intravaginal administration of human type III collagen-derived biomaterial with high cell-adhesion activity to treat vaginal atrophy in rats. ACS Biomater. Sci. Eng. 2020, 6, 1977–1988. [Google Scholar] [CrossRef]
- Abir, R.; Stav, D.; Taieb, Y.; Gabbay-Benziv, R.; Kirshner, M.; Ben-Haroush, A.; Freud, E.; Ash, S.; Yaniv, I.; Herman-Edelstein, M.; et al. Novel extra cellular-like matrices to improve human ovarian grafting. J. Assist. Reprod. Genet. 2020, 37, 2105–2117. [Google Scholar] [CrossRef]
- Rumian, Ł.; Wolf-Brandstetter, C.; Rößler, S.; Reczyńska, K.; Tiainen, H.; Haugen, H.J.; Scharnweber, D.; Pamuła, E. Sodium alendronate loaded poly(l-lactide- co-glycolide) microparticles immobilized on ceramic scaffolds for local treatment of bone defects. Regen. Biomater. 2020, 7, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Willard, J.J.; Drexler, J.W.; Das, A.; Roy, S.; Shilo, S.; Shoseyov, O.; Powell, H.M. Plant-derived human collagen scaffolds for skin tissue engineering. Tissue Eng. Part A 2013, 19, 1507–1518. [Google Scholar] [CrossRef]
- Yaari, A.; Schilt, Y.; Tamburu, C.; Raviv, U.; Shoseyov, O. Wet spinning and drawing of human recombinant collagen. ACS Biomater. Sci. Eng. 2016, 2, 349–360. [Google Scholar] [CrossRef]
- Ding, M.; Andreasen, C.M.; Dencker, M.L.; Jensen, A.E.; Theilgaard, N.; Overgaard, S. Efficacy of a small cell-binding peptide coated hydroxyapatite substitute on bone formation and implant fixation in sheep. J. Biomed. Mater. Res. Part A 2015, 103, 1357–1365. [Google Scholar] [CrossRef]
- Hou, Y.; Guey, L.T.; Wu, T.; Gao, R.; Cogan, J.; Wang, X.; Hong, E.; Vivian Ning, W.; Keene, D.; Liu, N.; et al. Intravenously administered recombinant human type VII collagen derived from chinese hamster ovary cells reverses the disease phenotype in recessive dystrophic epidermolysis bullosa mice. J. Investig. Dermatol. 2015, 135, 3060–3067. [Google Scholar] [CrossRef] [Green Version]
- Remington, J.; Wang, X.; Hou, Y.; Zhou, H.; Burnett, J.; Muirhead, T.; Uitto, J.; Keene, D.R.; Woodley, D.T.; Chen, M. Injection of recombinant human type VII collagen corrects the disease phenotype in a murine model of dystrophic epidermolysis bullosa. Mol. Ther. 2009, 17, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Hodges, B.L.; Barnes, T.M.; Reilly, P.R.; Kowtoniuk, W.E. Collagen IV replacement. Application Number 15/328,215, 26 July 2018. [Google Scholar]
- Hudson, B.G.; Cummings, C.F.; Pedchenko, V.; Brown, K.; Vanacore, R. Recombinant Collagen IV Surrogates and Uses Thereof. Application Number 15/329,900, 1 March 2018. [Google Scholar]
- Bardhan, A.; Bruckner-Tuderman, L.; Chapple, I.L.C.; Fine, J.D.; Harper, N.; Has, C.; Magin, T.M.; Marinkovich, M.P.; Marshall, J.F.; McGrath, J.A.; et al. Epidermolysis bullosa. Nat. Rev. Dis. Primers 2020, 6, 78. [Google Scholar] [CrossRef]
- Has, C.; South, A.; Uitto, J. Molecular therapeutics in development for epidermolysis bullosa: Update 2020. Mol. Diagn. Ther. 2020, 24, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Supp, D.M.; Hahn, J.M.; Combs, K.A.; McFarland, K.L.; Schwentker, A.; Boissy, R.E.; Boyce, S.T.; Powell, H.M.; Lucky, A.W. Collagen VII expression is required in both keratinocytes and fibroblasts for anchoring fibril formation in bilayer engineered skin substitutes. Cell Transpl. 2019, 28, 1242–1256. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, T.; Mezger, M.; Hausser, I.; Guey, L.T.; Handgretinger, R.; Bruckner-Tuderman, L.; Nystrom, A. Collagen VII Half-Life at the dermal-epidermal junction zone: Implications for mechanisms and therapy of genodermatoses. J. Investig. Dermatol. 2016, 136, 1116–1123. [Google Scholar] [CrossRef] [Green Version]
- Saelman, E.U.; Nieuwenhuis, H.K.; Hese, K.M.; de Groot, P.G.; Heijnen, H.F.; Sage, E.H.; Williams, S.; McKeown, L.; Gralnick, H.R.; Sixma, J.J. Platelet adhesion to collagen types I through VIII under conditions of stasis and flow is mediated by GPIa/IIa (α2 β1-integrin). Blood 1994, 83, 1244–1250. [Google Scholar] [CrossRef] [Green Version]
- Arita, M.; Fertala, J.; Hou, C.; Kostas, J.; Steplewski, A.; Fertala, A. Prospects and limitations of improving skeletal growth in a mouse model of spondyloepiphyseal dysplasia caused by R992C (p.R1192C) substitution in collagen II. PLoS ONE 2017, 12, e0172068. [Google Scholar] [CrossRef]
- Arita, M.; Fertala, J.; Hou, C.; Steplewski, A.; Fertala, A. Mechanisms of aberrant organization of growth plates in conditional transgenic mouse model of spondyloepiphyseal dysplasia associated with the R992C substitution in collagen II. Am. J. Pathol. 2015, 185, 214–229. [Google Scholar] [CrossRef]
- A Phase 1/2 Trial of PTR-01 in Adult Patients with Recessive Dystrophic Epidermolysis Bullosa (RDEB). Available online: https://ClinicalTrials.gov/show/NCT03752905 (accessed on 20 October 2020).
- O’Donnell, B.T.; Ives, C.J.; Mohiuddin, O.A.; Bunnell, B.A. Beyond the present constraints that prevent a wide spread of tissue engineering and regenerative medicine approaches. Front. Bioeng. Biotechnol. 2019, 7, 95. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Expression System | Examples of Collagen Constructs | Requirement for Co-Expression of P4H (N = No, Y = Yes) | Industrial-Scale Production (N = No, Y = Yes) | Commercial Evaluation (N = No, Y = Yes) | References |
|---|---|---|---|---|---|
| Mammalian cells (HT1080, CHO, HEK293, NIH3T3) | Native-like human procollagens, including procollagen I, procollagen II, collagen VI, procollagen VII. Fragments of procollagens, including mini-collagen II, mini-collagen I homotrimer, mini-collagen VII, C-terminal propeptides of procollagen III, and fragments of collagen IV | N | N | N | [4,50,53,83,84,85,86,87,88] |
| Insect cells | Native-like collagens including collagen I, collagen II, collagen III, collagen IX, collagen | N | N | N | [89,90] |
| Mammary glands of transgenic mice | Collagen I homotrimer | N | N | N | [62] |
| Escherichia coli | Human-derived mini-collagen III, collagen fragments, including C propeptide of collagen XVIII, and fragments of collagen I | Y | N | N | [61] |
| Escherichia coli | Collagen fragments stabilized by bacterial collagen-like sequences | N | N | N | [91,92] |
| Yeast cells | Native-like human collagen I, collagen III, gelatin | Y | Y | Y | [28,60,72,93] |
| Transgenic plants | Native-like human collagen I | Y | Y | Y | [59,77,94,95,96] |
| Collagen Construct | Expression System | Proposed Application | Experimental Tests | Applied Products (N = No, Y = Yes) | References |
|---|---|---|---|---|---|
| Full-length native-like collagen II, collagen VII | Mammalian cells HT1080, CHO | Cartilage engineering, protein replacement in patients harboring mutations in collagen VII, research tool | In vitro, mouse | N | [88,105] |
| Truncated and modified collagen II variants, truncated collagen VII | Mammalian cells HT1080, HEK293 | Cartilage engineering, research tool | In vitro, mouse | N | [16,83,109] |
| Full-length native-like human collagen I, collagen III | Yeasts | Fabrication of scaffolds and hydrogels to repair damaged tissues | Mouse | N | [75] |
| Hemostatic materials | Rabbit | N | [136] | ||
| Implants to regenerate cornea | Human | N | [137,138] | ||
| Modified collagen III | Yeasts | Materials with increased thermostability | In vitro | N | [139] |
| Materials with customized collagen III sequences for support of stem cells | In vitro | N | [129] | ||
| Collagen III constructs containing integrin-binding sites from collagen I and laminin | Yeasts | Scaffolds to support neural progenitor cells | In vitro | N | [129] |
| Non-triple helical collagen I fragment | Yeasts | Scaffolds for tissue regeneration | In vitro | N | [99] |
| Scaffolds for transplantation of pancreatic islets | Mouse | N | [140] | ||
| Grafting material for bone regeneration | In vitro | N | [115,141,142,143,144] | ||
| Native-like collagen II | Yeasts | Hydrogel to support chondrogenesis of mesenchymal stromal cells | In vitro | N | [145,146] |
| Collagen I fragment fused with (Pro-Gly-Pro)9 peptides | Yeasts | Gelatin mimetic | In vitro | N | [31] |
| Full-length human collagen III | Bacteria | NA | In vitro | N | [70] |
| Collagen III fragments fused with bacteria-derived collagen-like proteins | Bacteria | Inhibitors of DDR signaling | In vitro | N | [147] |
| Tandem repeats of the (GAPGAPGSQGAPGLQ) fragment | Bacteria | Material to deliver BMP-2 for bone repair | Mouse | N | [148] |
| Tandem repeats of (GPP) fragment | Fabrication of biocompatible surfaces | In vitro | N | [149] | |
| Fragment of turtle-derived collagen | Bacteria | Antioxidant material | In vitro | N | [150] |
| Collagen III-derived fragments | Bacteria | Treatment of vaginal atrophy | Rat | N | [151] |
| Full-length native-like human collagen I | Tobacco | Wound dressing materials | Y | [134] | |
| Matrices for ovarian grafting | Mouse | N | [152] | ||
| Scaffolds for bone and skin repair | In vitro | N | [153,154] | ||
| Electro-spun fibrils for tendon-repair materials | In vitro | N | [155] | ||
| Injectable material combined with platelet-rich plasma for treatment of lateral epicondylar tendinopathy | Human | Y | [76] | ||
| Short collagen-derived linear peptides | Bacteria, yeast (recombinant technology) and chemical synthesis | Osteogenic material | In vitro, human | Y | [68,69,110,115,156] |
| Disease | Collagen Target | Experimental Model | Clinical Tests | Applied Clinically | References |
|---|---|---|---|---|---|
| Dystrophic epidermolysis bullosa | Collagen VII | Intradermal or intravenous delivery of recombinant collagen VII into mice | Y | N | [168] |
| Alport syndrome | Collagen IV | Systemic delivery | N | N | [159,160] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fertala, A. Three Decades of Research on Recombinant Collagens: Reinventing the Wheel or Developing New Biomedical Products? Bioengineering 2020, 7, 155. https://doi.org/10.3390/bioengineering7040155
Fertala A. Three Decades of Research on Recombinant Collagens: Reinventing the Wheel or Developing New Biomedical Products? Bioengineering. 2020; 7(4):155. https://doi.org/10.3390/bioengineering7040155
Chicago/Turabian StyleFertala, Andrzej. 2020. "Three Decades of Research on Recombinant Collagens: Reinventing the Wheel or Developing New Biomedical Products?" Bioengineering 7, no. 4: 155. https://doi.org/10.3390/bioengineering7040155