Proteomic Examination for Gluconeogenesis Pathway-Shift during Polyhydroxyalkanoate Formation in Cupriavidus necator Grown on Glycerol



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and PHA Synthesis
2.2. Analytical Procedures
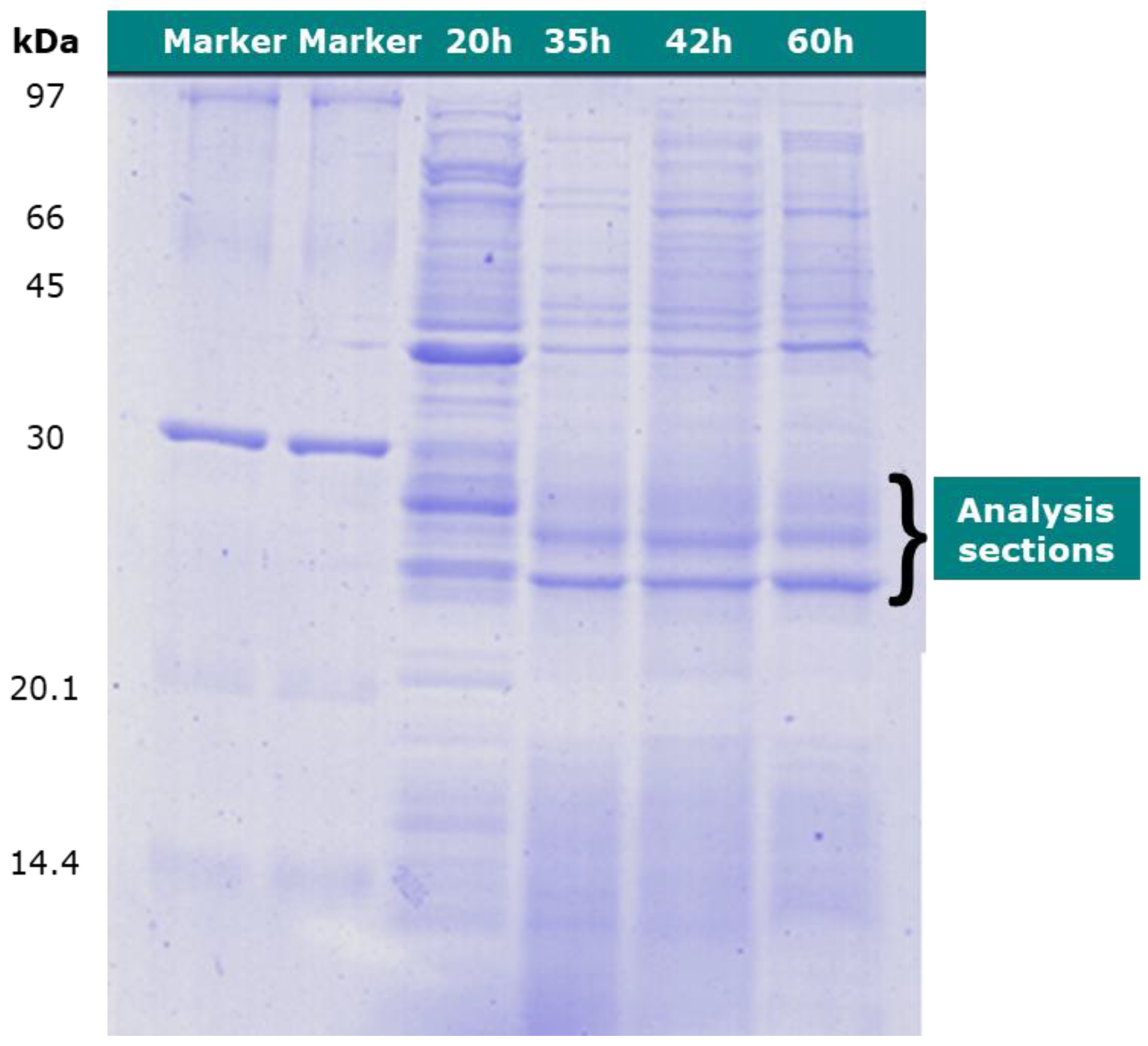
2.3. Protein Assay with 1-D SDS–PAGE
2.4. Analysis of Proteomic Information
2.5. Statistical Analysis
3. Results
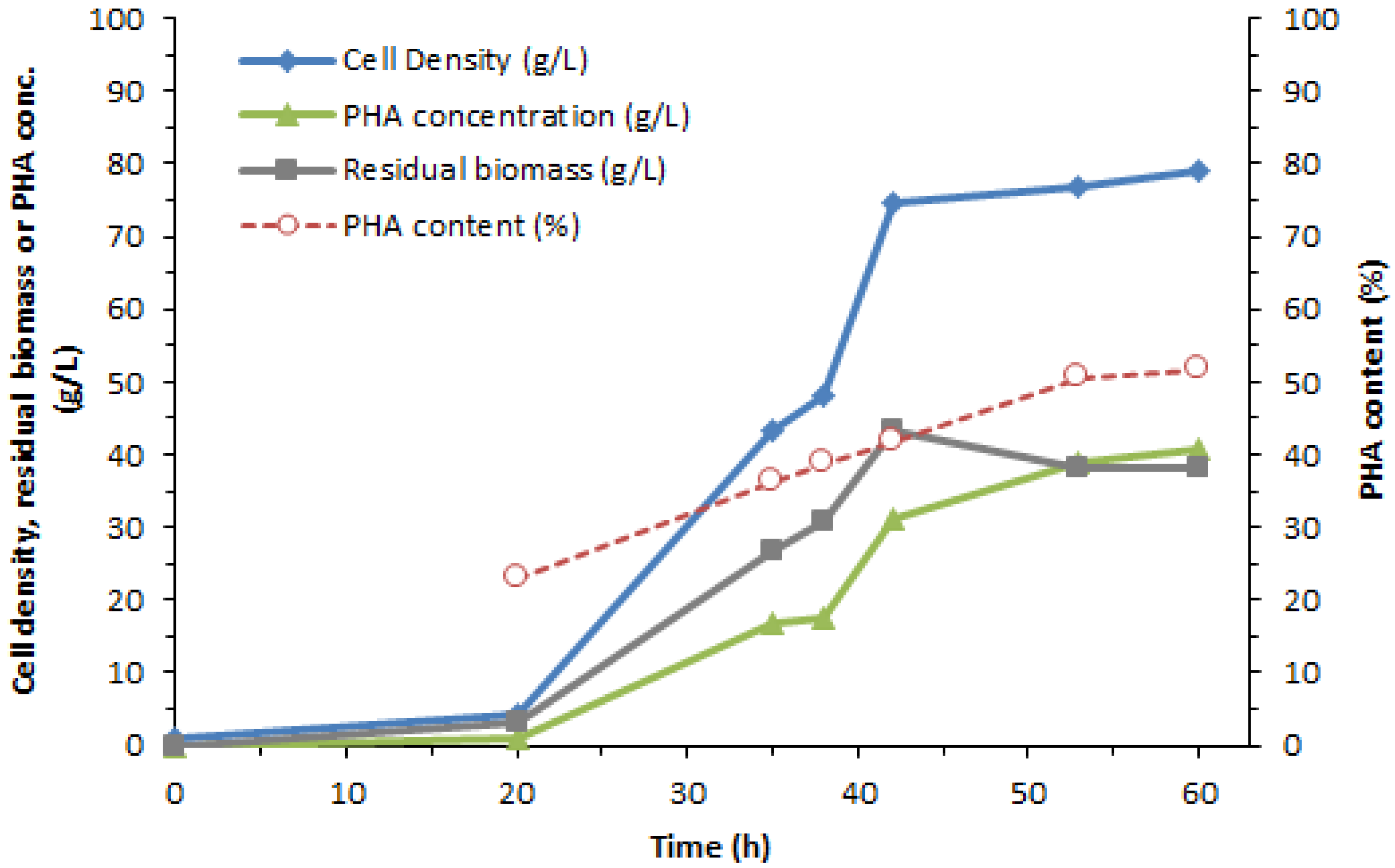
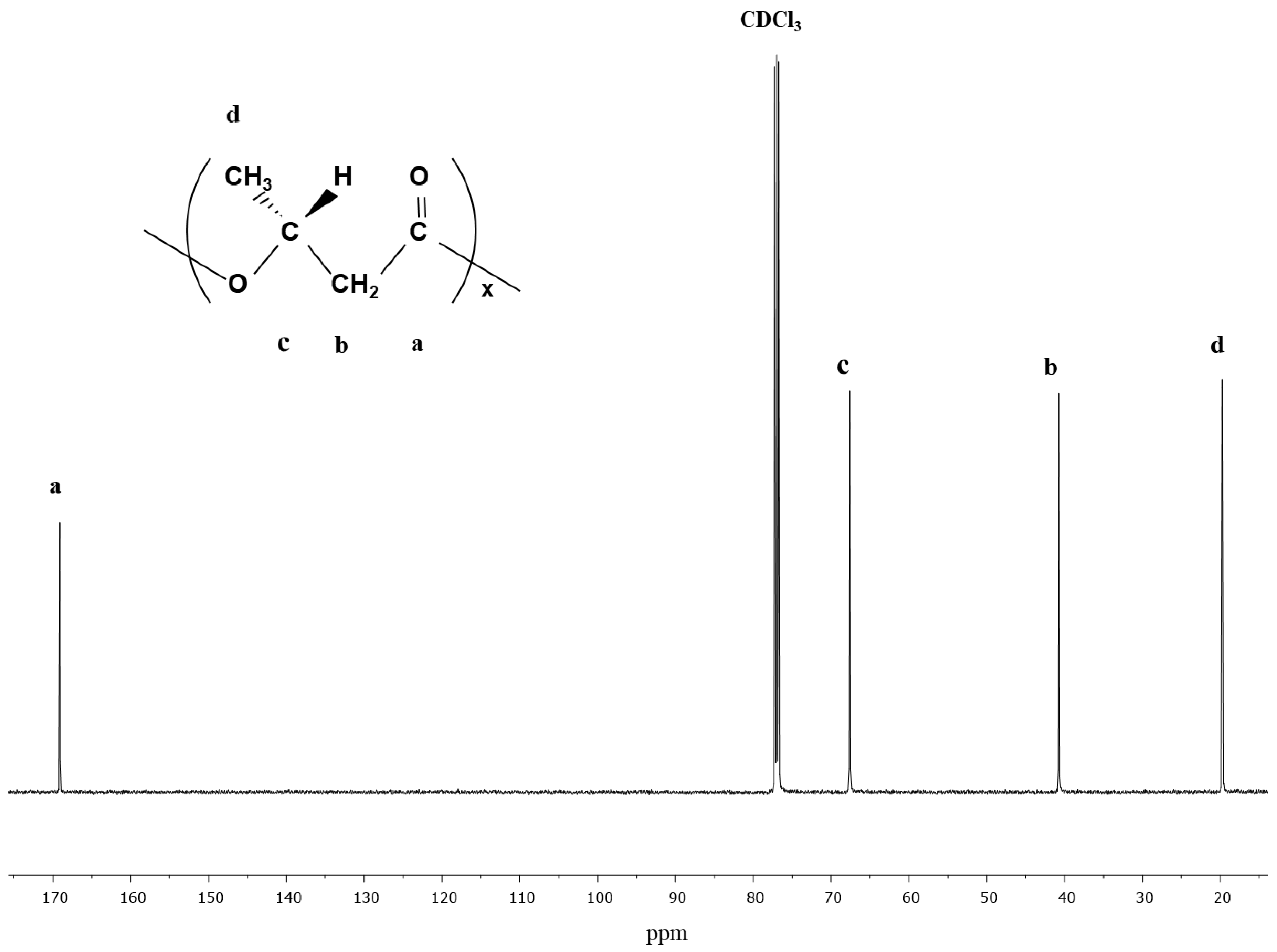
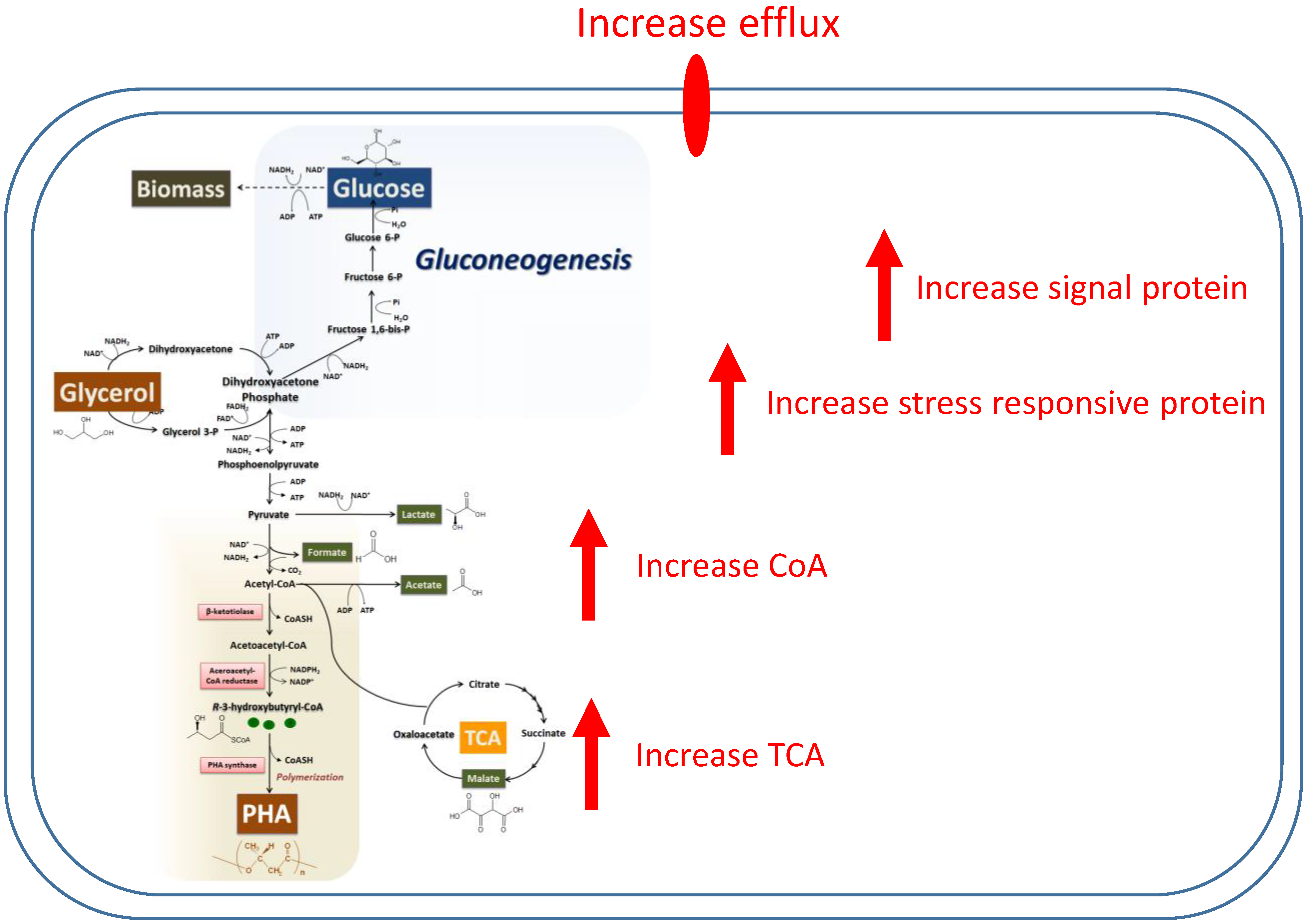
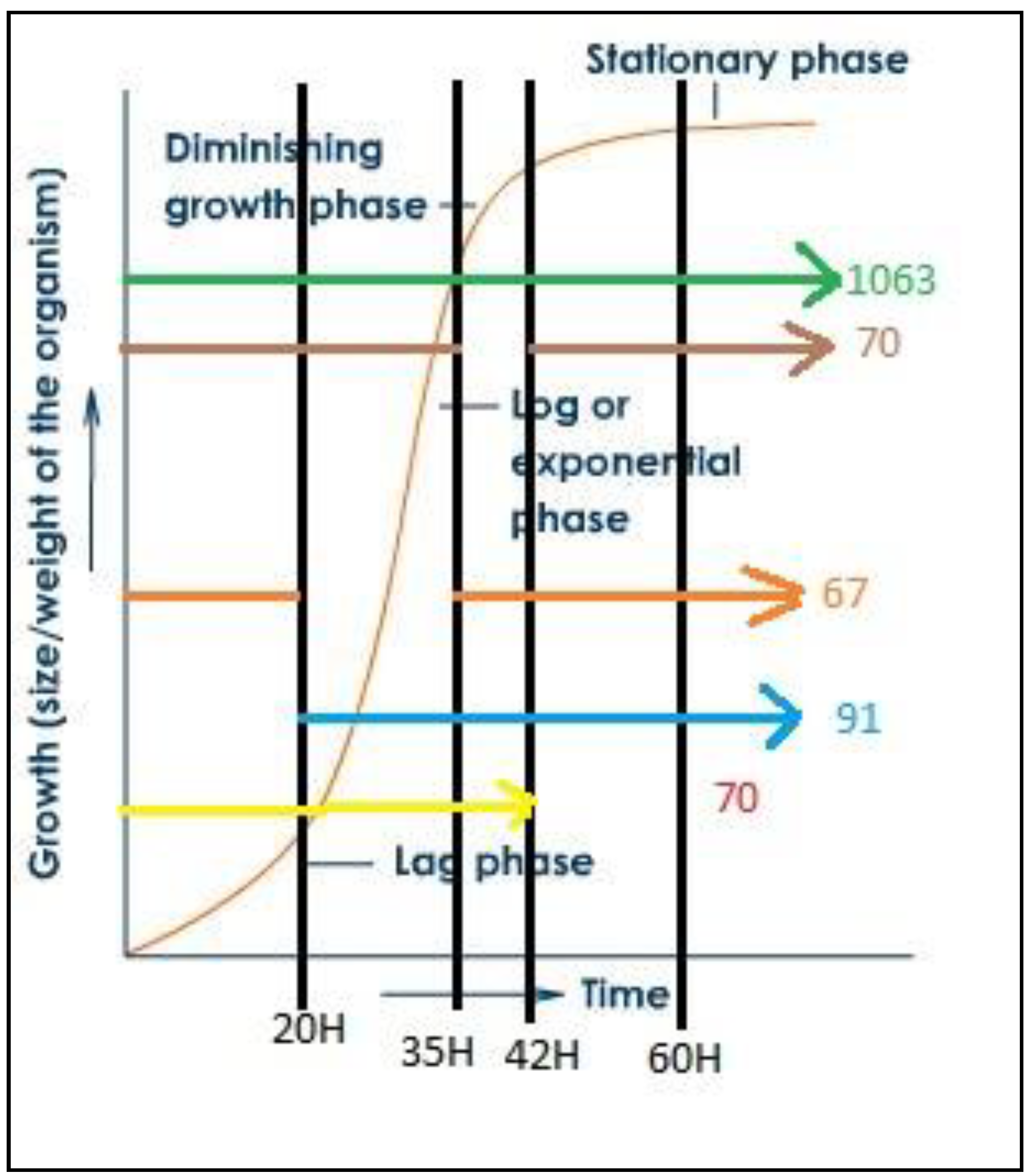
3.1. PHA Synthesis from C. necator with Glycerol as a Carbon Source
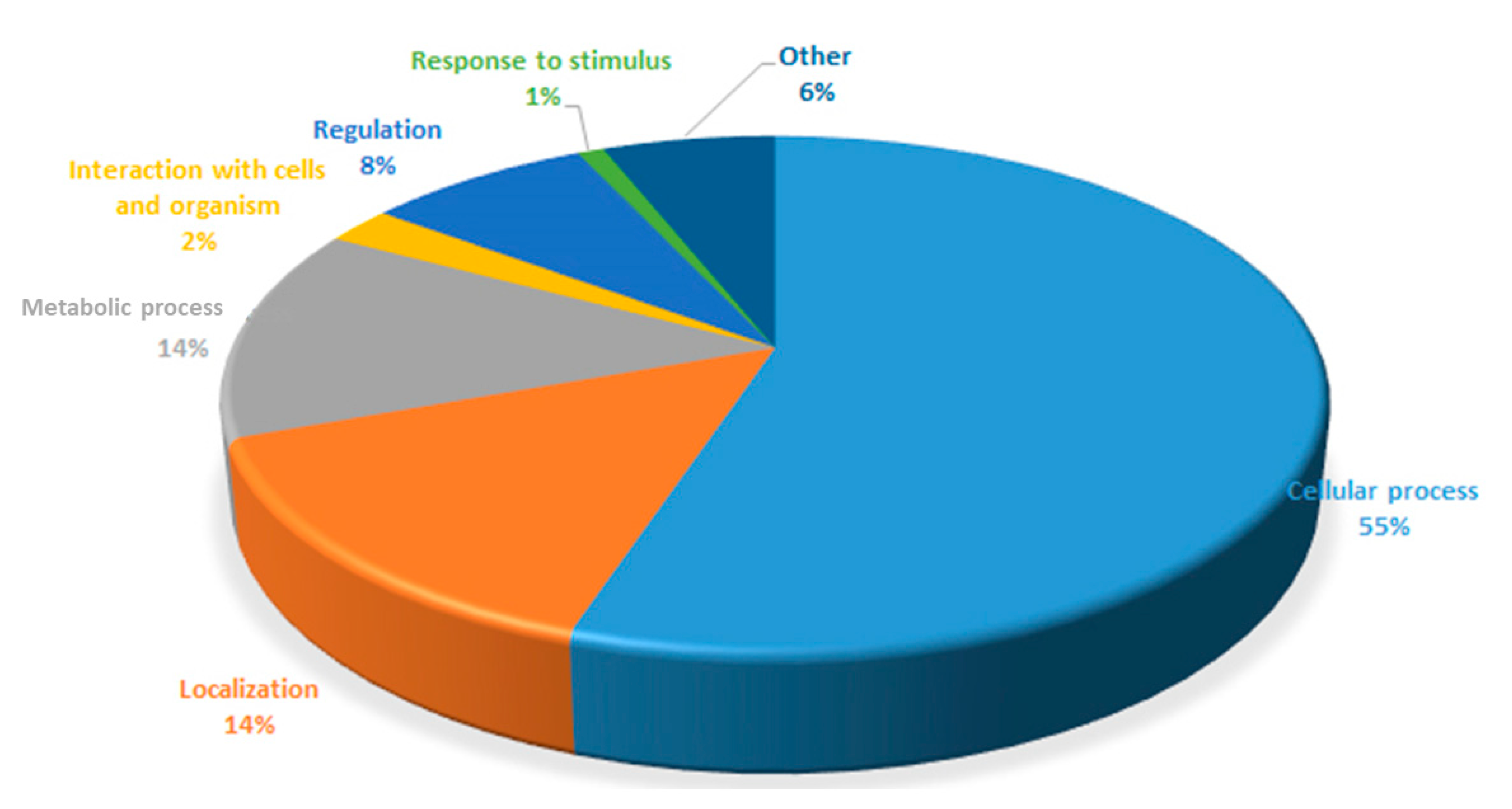
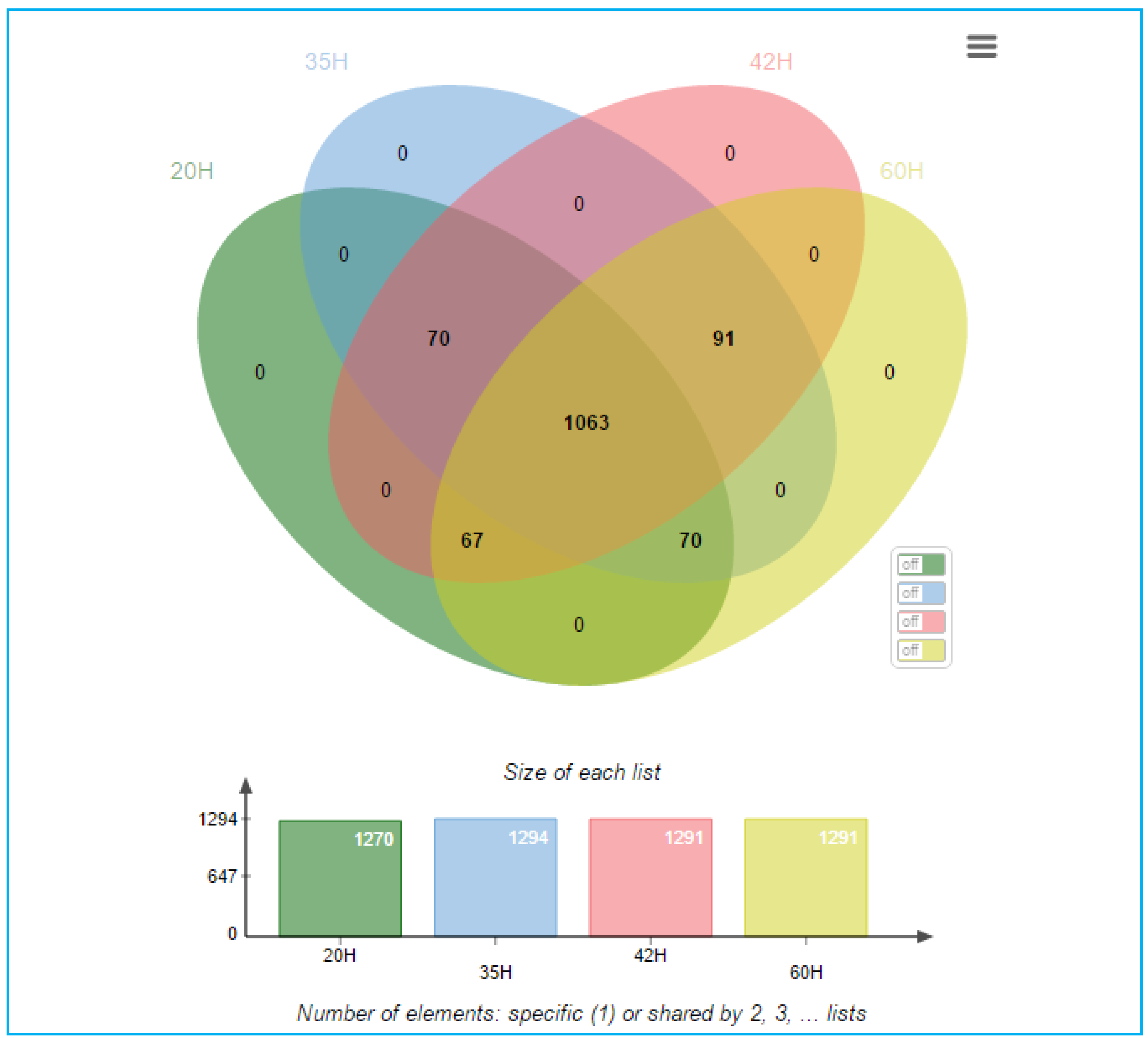
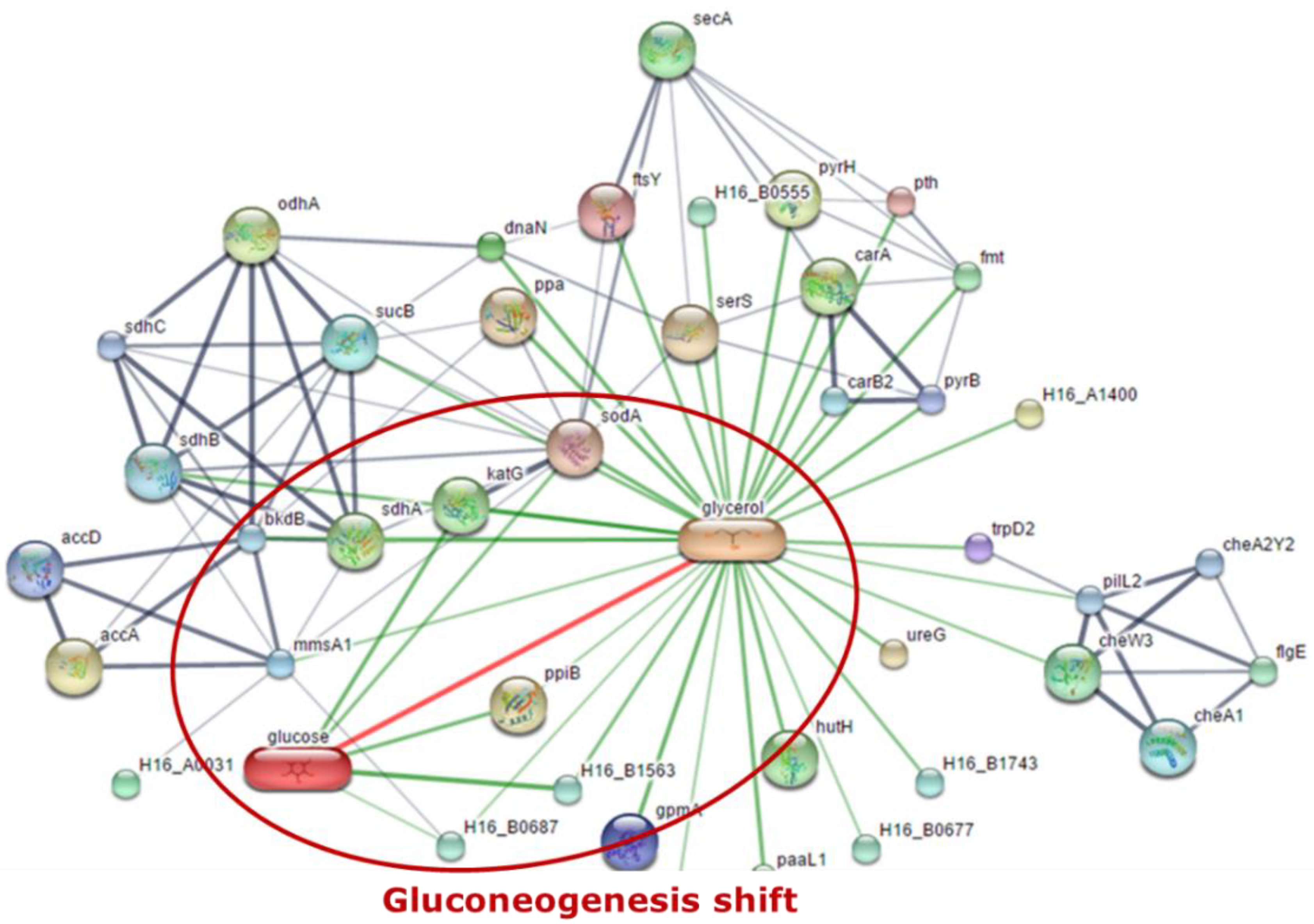
3.2. Proteomic Analysis of C. necator during PHA Synthesis
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, S.Y. Deciphering bioplastic production. Nat. Biotechnol. 2006, 24, 1227–1229. [Google Scholar] [CrossRef] [PubMed]
- Akaraonye, E.; Keshavarz, T.; Roy, I. Production of polyhydroxyalkanoates: The future green materials of choice. J. Chem. Technol. Biotechnol. 2010, 85, 732–743. [Google Scholar] [CrossRef]
- Lee, S.Y. Bacterial polyhydroxyalkanoates. Biotechnol. Bioeng. 1996, 49, 1–4. [Google Scholar] [PubMed]
- Witholt, B.; Kessler, B. Perspectives of medium chain length poly(hydroxyalkanoates), a versatile set of bacterial bioplastics. Curr. Opin. Biotechnol. 1999, 10, 279–285. [Google Scholar]
- Williams, S.F.; Martin, D.P.; Horowitz, D.M.; Peoples, O.P. PHA applications: Addressing the price performance issue: I. Tissue engineering. Int. J. Biol. Macromol. 1999, 25, 111–121. [Google Scholar]
- Zinn, M.; Witholt, B.; Egli, T. Occurrence, synthesis and medical application of bacterial polyhydroxyalkanoate. Adv. Drug Deliv. Rev. 2001, 53, 5–21. [Google Scholar]
- Chen, G.-Q.; Wu, Q. The application of polyhydroxyalkanoates as tissue engineering materials. Biomaterials 2005, 26, 6565–6578. [Google Scholar]
- Dellomonaco, C.; Fava, F.; Gonzalez, R. The path to next generation biofuels: Successes and challenges in the era of synthetic biology. Microb. Cell Factories 2010, 9, 3. [Google Scholar]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef]
- Da Silva, G.P.; Mack, M.; Contiero, J. Glycerol: A promising and abundant carbon source for industrial microbiology. Biotechnol. Adv. 2009, 27, 30–39. [Google Scholar] [CrossRef]
- Choi, W.J. Glycerol-Based Biorefinery for Fuels and Chemicals. Recent Pat. Biotechnol. 2008, 2, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Ashby, R.; Solaiman, D.; Strahan, G. Efficient Utilization of Crude Glycerol as Fermentation Substrate in the Synthesis of Poly(3-hydroxybutyrate) Biopolymers. J. Am. Oil Chem. Soc. 2011, 88, 949–959. [Google Scholar] [CrossRef]
- Zhu, C.; Nomura, C.T.; Perrotta, J.A.; Stipanovic, A.J.; Nakas, J.P. Production and characterization of poly-3-hydroxybutyrate from biodiesel-glycerol by Burkholderia cepacia ATCC 17759. Biotechnol. Prog. 2010, 26, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, J.M.B.T.; De Almeida, M.C.M.D.; Grandfils, C.; Da Fonseca, M.M.R. Poly(3-hydroxybutyrate) production by Cupriavidus necator using waste glycerol. Process. Biochem. 2009, 44, 509–515. [Google Scholar]
- Ibrahim, M.H.A.; Steinbüchel, A. Poly(3-Hydroxybutyrate) Production from Glycerol by Zobellella denitrificans MW1 via High-Cell-Density Fed-Batch Fermentation and Simplified Solvent Extraction. Appl. Environ. Microbiol. 2009, 75, 6222–6231. [Google Scholar] [CrossRef] [Green Version]
- Pohlmann, A.; Fricke, W.F.; Reinecke, F.; Kusian, B.; Liesegang, H.; Cramm, R.; Eitinger, T.; Ewering, C.; Potter, M.; Schwartz, E.; et al. Genome sequence of the bioplastic-producing bacterium Ralstonia eutropha H16. Nat. Biotechnol. 2006, 24, 1257–1262. [Google Scholar]
- Tanadchangsaeng, N.; Yu, J. Microbial synthesis of polyhydroxybutyrate from glycerol: Gluconeogenesis, molecular weight and material properties of biopolyester. Biotechnol. Bioeng. 2012, 109, 2808–2818. [Google Scholar] [CrossRef]
- Schwartz, E.; Voigt, B.; Zühlke, D.; Pohlmann, A.; Lenz, O.; Albrecht, D.; Schwarze, A.; Kohlmann, Y.; Krause, C.; Hecker, M.; et al. A proteomic view of the facultatively chemolithoautotrophic lifestyle of Ralstonia eutropha H16. Proteomics 2009, 9, 5132–5142. [Google Scholar] [CrossRef]
- Chandramouli, K.; Qian, P.-Y. Proteomics: Challenges, Techniques and Possibilities to Overcome Biological Sample Complexity. Hum. Genom. Proteom. 2009, 1. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.E.; Li, Q.X.; Yu, J. Proteomic examination of Ralstonia eutropha in cellular responses to formic acid. Proteomics 2006, 6, 4259–4268. [Google Scholar] [CrossRef]
- Lee, S.E.; Li, Q.X.; Yu, J. Diverse protein regulations on PHA formation in Ralstonia eutropha on short chain organic acids. Int. J. Biol. Sci. 2009, 5, 215–225. [Google Scholar] [PubMed] [Green Version]
- Yu, J.; Chen, L.X.L.; Sato, S. Biopolyester Synthesis and Protein Regulations in Ralstonia eutropha on Levulinic Acid and Its Derivatives from Biomass Refining. J. Biobased Mater. Bioenergy 2009, 3, 113–122. [Google Scholar] [CrossRef]
- Braunegg, G.; Sonnleitner, B.; Lafferty, R.M. A rapid gas chromatographic method for the determination of poly-β-hydroxybutyric acid in microbial biomass. Eur. J. Appl. Microbiol. Biotechnol. 1978, 6, 29–37. [Google Scholar] [CrossRef]
- Davies, G.; Henrissat, B. Structures and mechanisms of glycosyl hydrolases. Structure 1995, 3, 853–859. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivation Time (h) | Cell Dry Mass (g/L) | PHA Content (%) | PHA Concentration (g/L) | PHA Productivity (g/L/h) | Glucose/D2O NMR Area Ratio |
|---|---|---|---|---|---|
| 0 | 0.8 | ND | ND | ND | ND |
| 20 | 4.1 | 23.0 | 0.9 | 0.05 | 0 |
| 35 | 43.5 | 36.3 | 16.8 | 0.48 | 1.13 |
| 38 | 48.2 | 18.7 | 17.5 | 0.46 | ND |
| 42 | 74.7 | 41.8 | 31.2 | 0.74 | 2.26 |
| 53 | 77.0 | 50.6 | 38.9 | 0.73 | ND |
| 60 | 78.9 | 51.6 | 40.7 | 0.68 | 4.55 |
| No. | Protein Name | Accession Number | Function |
|---|---|---|---|
| 1 | methylmalonate-semialdehyde dehydrogenase (Jannaschia sp. CCS1) | gi|89054404 | Amino acid metabolism |
| 2 | acyl-CoA dehydrogenase domain-containing protein (Methylobacterium radiotolerans JCM 2831) | gi|170749664 | Lipid metabolism |
| 3 | succinate dehydrogenase flavoprotein subunit (gamma proteobacterium IMCC1989) | gi|497354190 | Respiration |
| 4 | capsular exopolysaccharide family protein (Thioalkalivibrio sp. K90mix) | gi|289207806 | Signal |
| 5 | integral membrane sensor signal transduction histidine kinase (Burkholderia sp. CCGE1002) | gi|295676684 | Signal |
| 6 | membrane protease subunit stomatin/prohibitin-like protein (Magnetospirillum magneticum AMB-1) | gi|83312588 | Signal |
| 7 | protein YhiI (Pasteurella multocida subsp. multocida str. HN06) | gi|383310937 | Signal |
| 8 | glycosyl transferase family protein (Burkholderia vietnamiensis G4) | gi|134291866 | Signal |
| 9 | short-chain dehydrogenase (marine gamma proteobacterium HTCC2143) | gi|494429549 | Signal |
| 10 | phosphoserine phosphatase SerB (Methylotenera versatilis 301) | gi|297537989 | Stress response |
| 11 | superoxide dismutase (Burkholderia cenocepacia AU 1054) | gi|107023480 | Stress response |
| 12 | beta-lactamase class C-like protein (Shewanella amazonensis SB2B) | gi|119774488 | Stress response |
| 13 | chemotaxis protein (Burkholderia thailandensis E264) | gi|83717148 | Stress response |
| 14 | preprotein translocase subunit SecA (Idiomarina baltica) | gi|494014389 | Transport |
| 15 | ABC-type transporter, auxilary periplasmic component involved in toluene tolerance (Ralstonia eutropha H16) | gi|113869370 | Transport |
| 16 | type II secretion system protein E (Pectobacterium carotovorum subsp. carotovorum PC1) | gi|253687071 | Transport |
| 17 | hypothetical protein (Vibrio mimicus) | gi|445940548 | Unknown |
| 18 | HrpA protein (Neisseria gonorrhoeae NCCP11945) | gi|194099492 | Unknown |
| 19 | hypothetical protein G157_07140 (Campylobacter coli CVM N29710) | gi|543941344 | Unknown |
| 20 | flagellar scaffolding protein FlgD (Loktanella vestfoldensis) | gi|494388140 | Unknown |
| 21 | hypothetical protein (Candidatus Regiella insecticola) | gi|493755517 | Unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanadchangsaeng, N.; Roytrakul, S. Proteomic Examination for Gluconeogenesis Pathway-Shift during Polyhydroxyalkanoate Formation in Cupriavidus necator Grown on Glycerol. Bioengineering 2020, 7, 154. https://doi.org/10.3390/bioengineering7040154
Tanadchangsaeng N, Roytrakul S. Proteomic Examination for Gluconeogenesis Pathway-Shift during Polyhydroxyalkanoate Formation in Cupriavidus necator Grown on Glycerol. Bioengineering. 2020; 7(4):154. https://doi.org/10.3390/bioengineering7040154
Chicago/Turabian StyleTanadchangsaeng, Nuttapol, and Sittiruk Roytrakul. 2020. "Proteomic Examination for Gluconeogenesis Pathway-Shift during Polyhydroxyalkanoate Formation in Cupriavidus necator Grown on Glycerol" Bioengineering 7, no. 4: 154. https://doi.org/10.3390/bioengineering7040154