Bioreactor Parameters for Microcarrier-Based Human MSC Expansion under Xeno-Free Conditions in a Vertical-Wheel System

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. hMSC Planar Culture for Seed Train
2.2. hMSC Bioreactor Culture
2.3. Bioreactor Sampling
2.4. In-Vessel Harvest
2.5. Analysis of Cell Health, Quality Attributes and Functionality
2.5.1. Cell Health Analysis
2.5.2. Tri-Lineage Differentiation
2.5.3. Flow Cytometry for Cell Surface Marker Expression
2.5.4. Immunomodulatory Function
2.5.5. Angiogenic Cytokine Secretion
2.6. Statistical Analysis
3. Results and Discussion
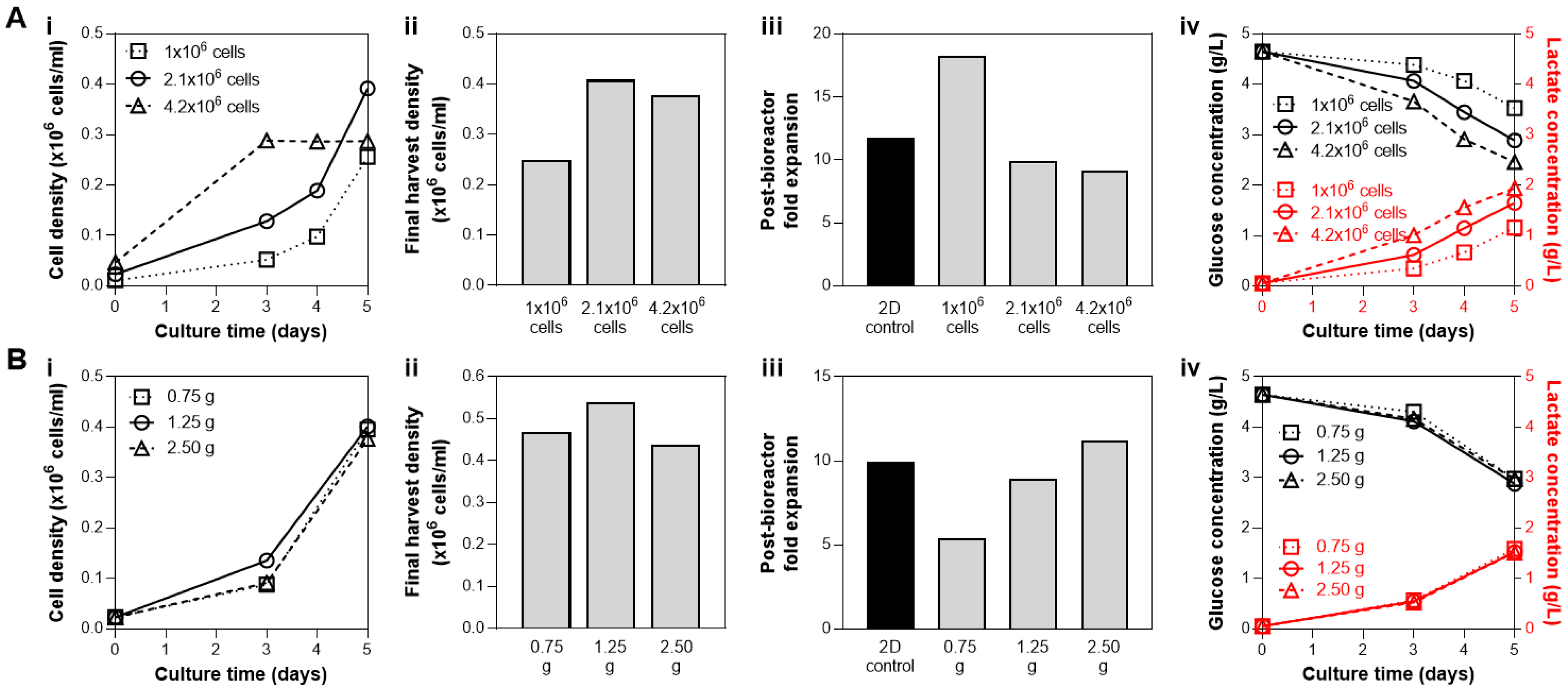
3.1. Cell and Microcarrier Inoculation Concentration in a Fixed Media Volume
3.2. Bioreactor Seeding Strategies: Cell and Microcarrier Concentrations at Inoculation
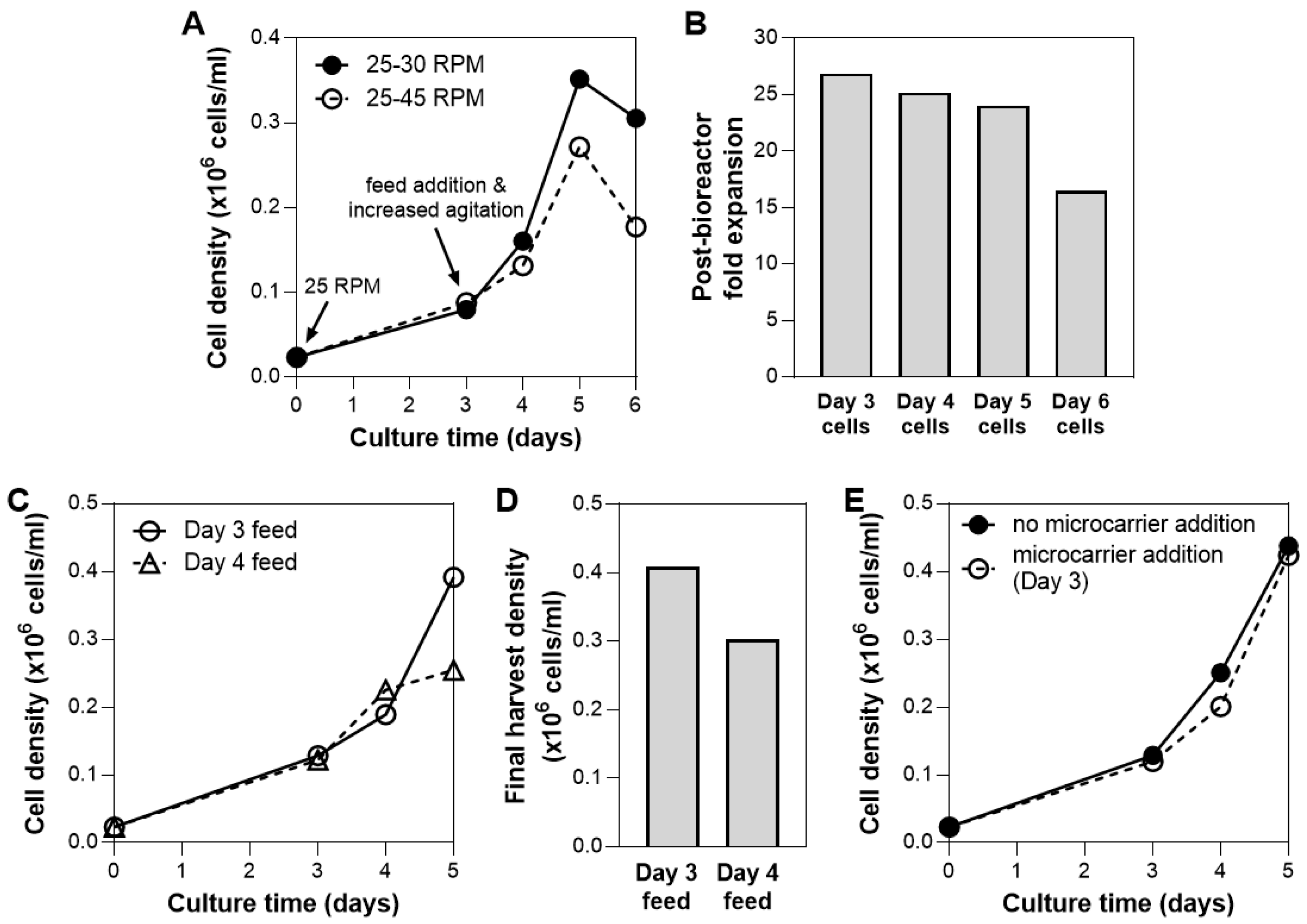
3.3. Bioreactor Culture Parameters: Agitation Speed, Day of Bioreactor Feed, Microcarrier Addition
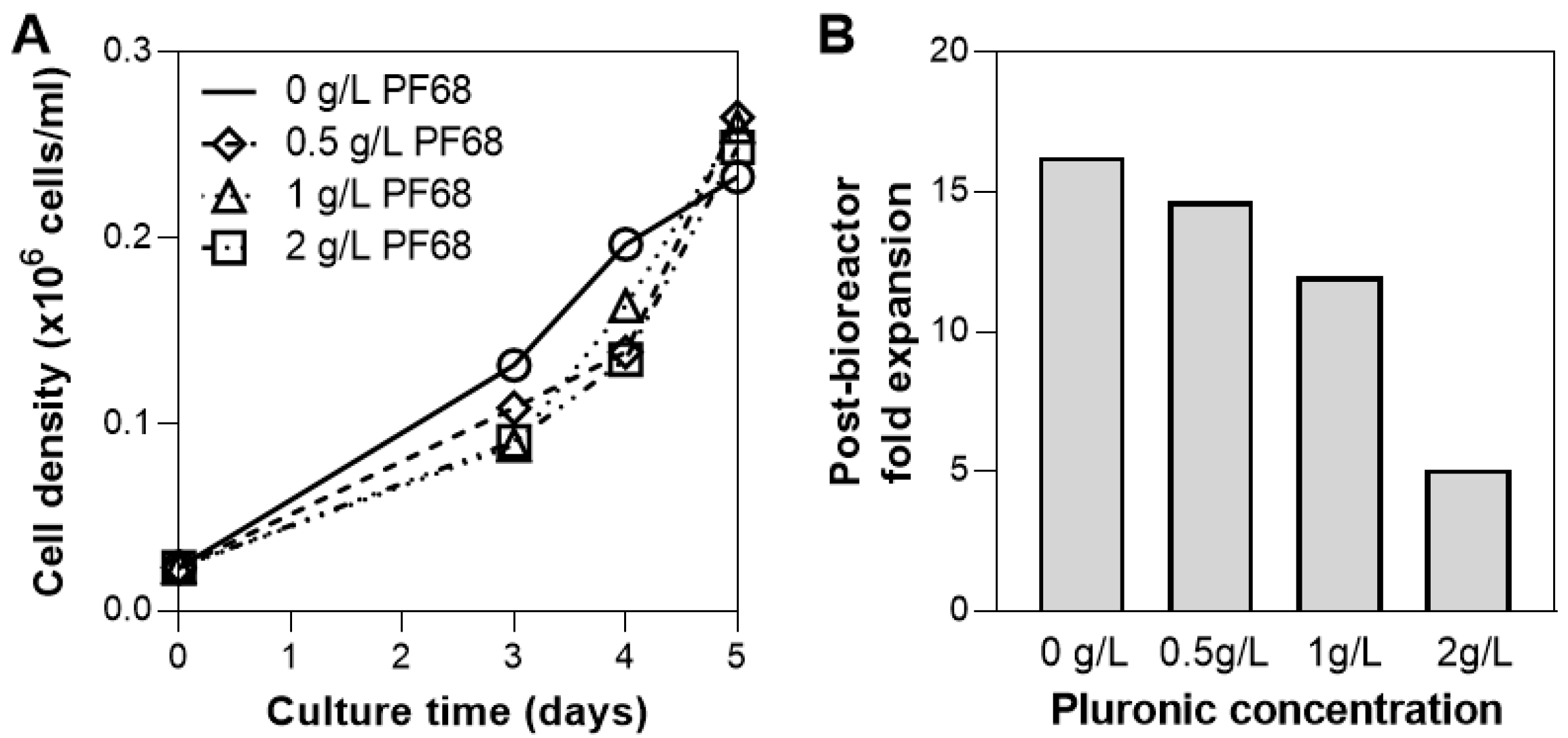
3.4. Addition of Surfactant
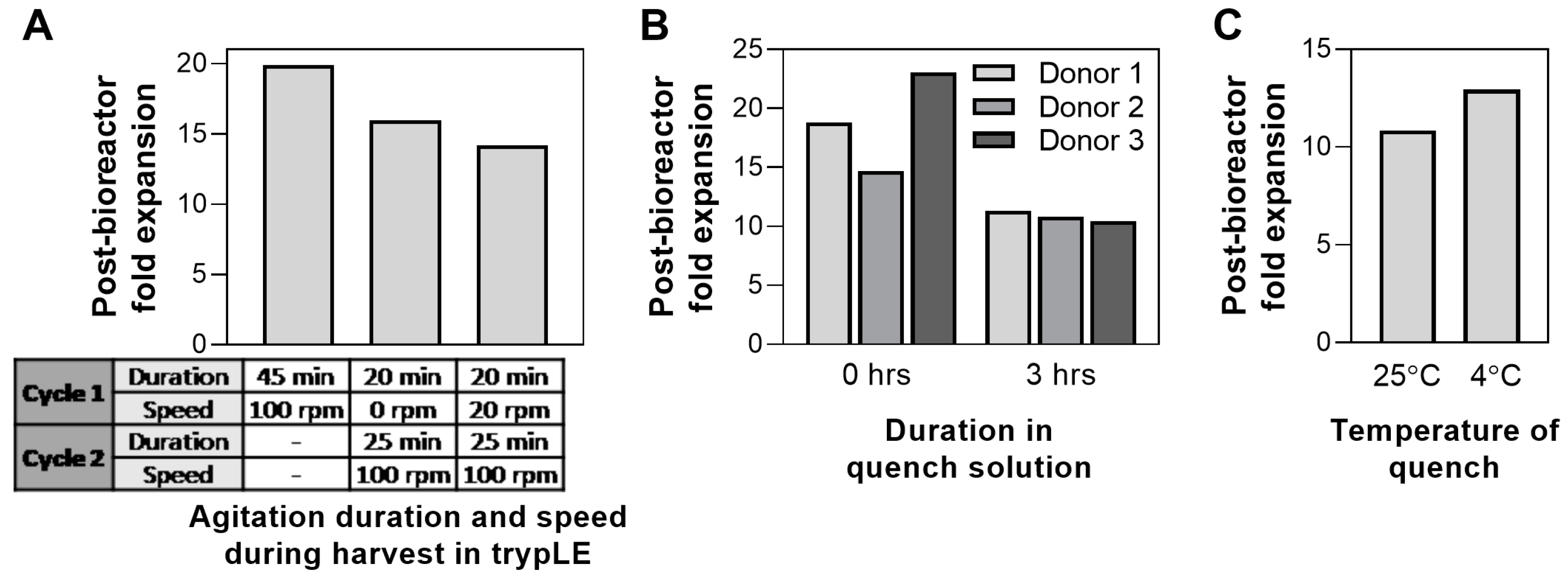
3.5. Bioreactor Harvest Process: Agitation Speed, Quench Hold Time, Quench Solution Temperature
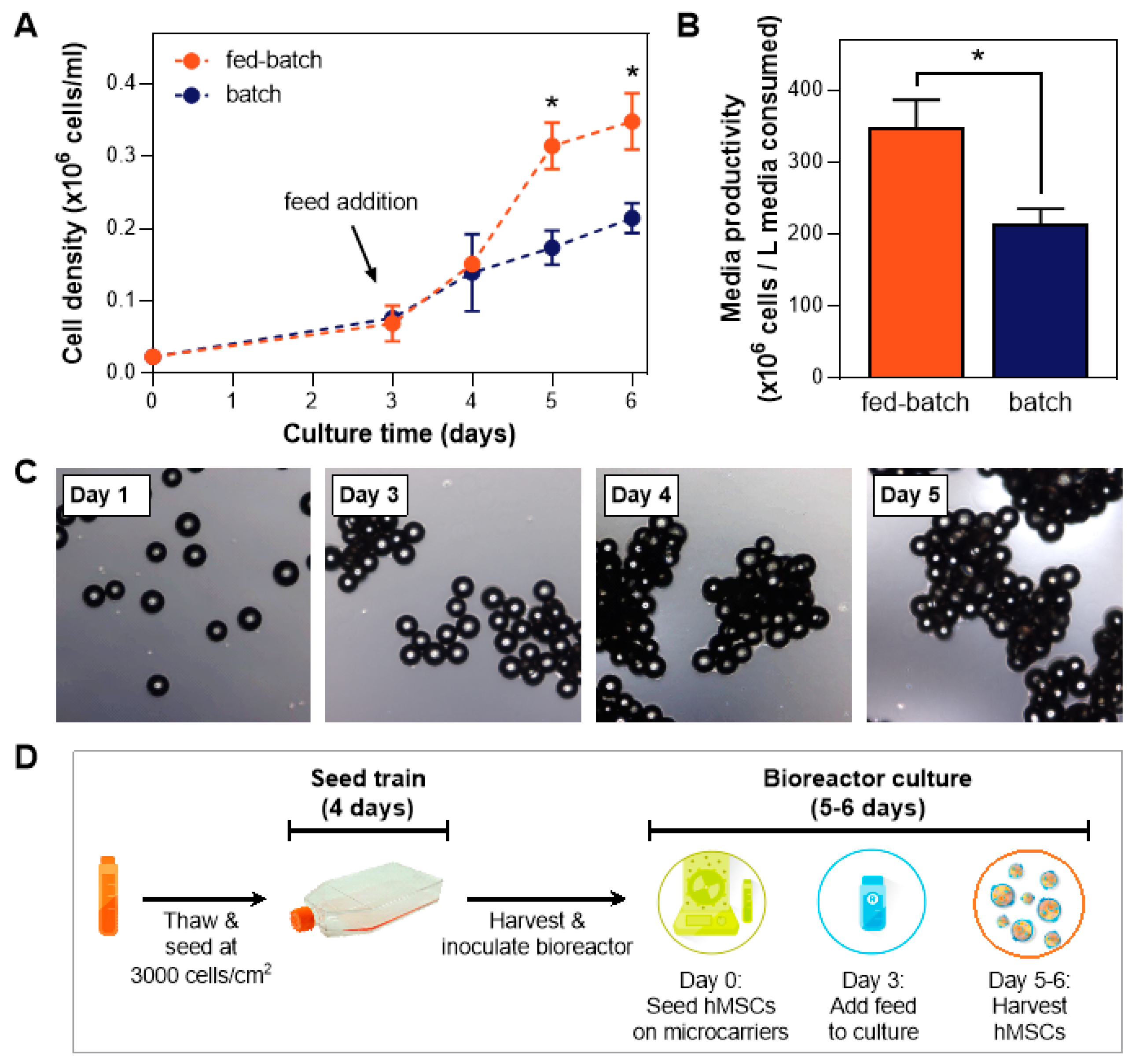
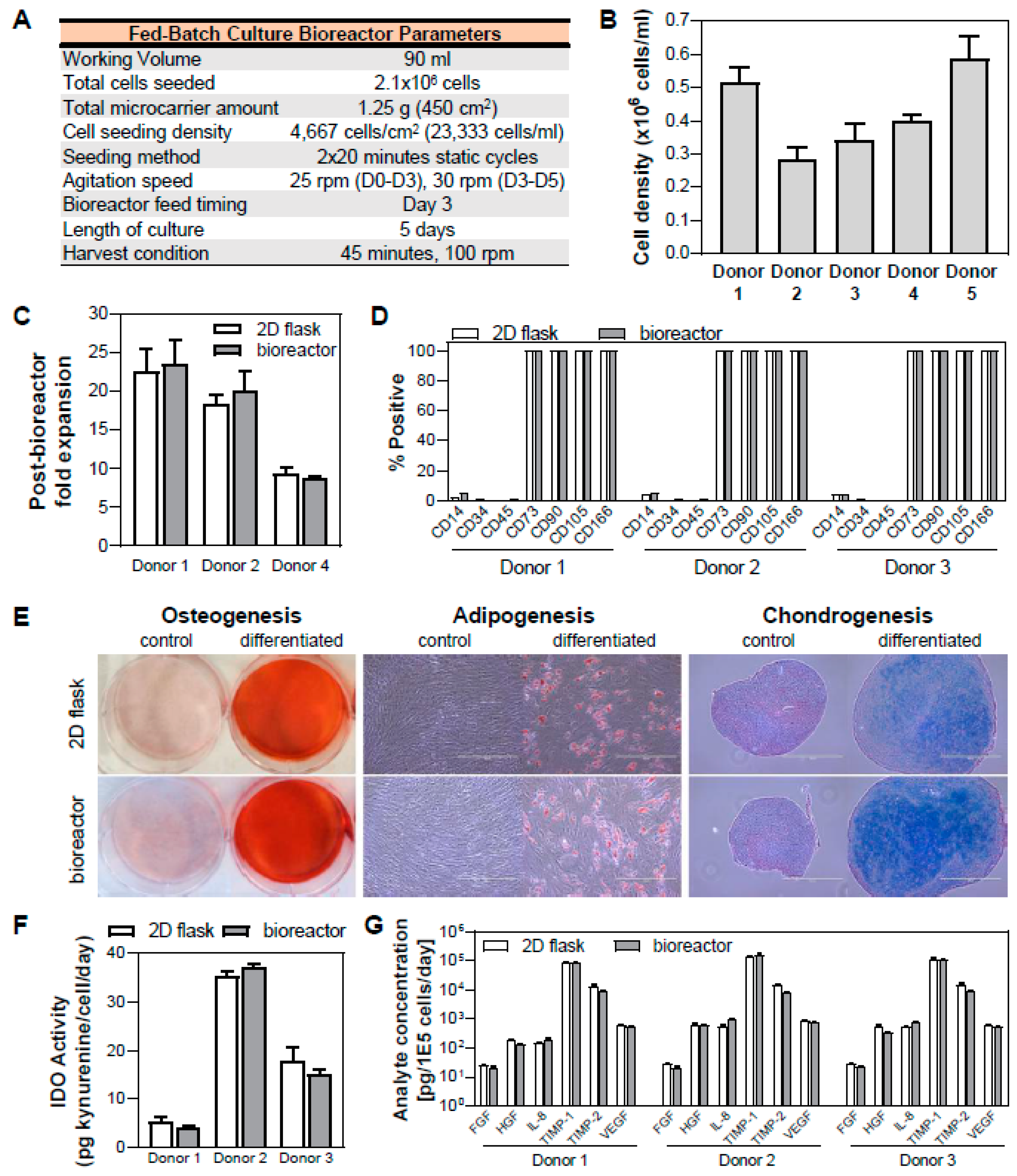
3.6. Validation of hMSC Expansion in Fed-batch Bioreactor Process and Maintenance of Critical Quality Attributes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Olsen, T.R.; Ng, K.S.; Lock, L.T.; Ahsan, T.; Rowley, J.A. Peak MSC-Are We There Yet? Front. Med. (Lausanne) 2018, 5, 178. [Google Scholar] [CrossRef]
- Rowley, J.A. Meeting Lot-Size Challenges of Manufacturing Adherent Cells for Therapy. Bioprocess. Int. 2012, 10, 16–22. [Google Scholar]
- Simaria, A.S.; Hassan, S.; Varadaraju, H.; Rowley, J.; Warren, K.; Vanek, P.; Farid, S.S. Allogeneic cell therapy bioprocess economics and optimization: Single-use cell expansion technologies. Biotechnol. Bioeng. 2014, 111, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.K.-L.; Chew, Y.K.; Tan, H.Y.; Reuveny, S.; Oh, S.K.W. Increasing efficiency of human mesenchymal stromal cell culture by optimization of microcarrier concentration and design of medium feed. Cytotherapy 2015, 17, 163–173. [Google Scholar] [CrossRef]
- Schnitzler, A.C.; Kehoe, D.; Simler, J.; DiLeo, A.; Ball, A. Scale-up of Human Mesenchymal Stem Cells on Microcarriers in Suspension in a Single-use Bioreactor. Bioprocess. Int. 2012, 25, 28–39. [Google Scholar]
- Szczypka, M.; Splan, D.; Woolls, H.; Brandwein, H. Single-Use Bioreactors and Microcarriers. Bioprocess. Int. 2014, 12, 54–64. [Google Scholar]
- Yourek, G.; McCormick, S.M.; Mao, J.J.; Reilly, G.C. Shear stress induces osteogenic differentiation of human mesenchymal stem cells. Regen. Med. 2010, 5, 713–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.-D.; Gu, Y.-Q.; Li, C.-M.; Wang, C.-R.; Feng, Z.-G.; Qiu, R.-X.; Chen, B.; Li, J.-X.; Zhang, S.-W.; Wang, Z.-G.; et al. Response of mesenchymal stem cells to shear stress in tissue-engineered vascular grafts. Acta Pharm. Sin. 2009, 30, 530–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, B.P.; Babalola, O.M.; Bonassar, L.J. Quantitative characterization of mesenchymal stem cell adhesion to the articular cartilage surface. J. Biomed. Mater. Res. A 2013, 101, 3592–3598. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, Q.A.; Brosnan, K.M.; Coopman, K.; Nienow, A.W.; Hewitt, C. Culture of human mesenchymal stem cells on microcarriers in a 5 l stirred-tank bioreactor. Biotechnol. Lett. 2013, 35, 1233–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, T.; Kehoe, D.E.; Schnitzler, A.; Rapiejko, P.J.; Der, K.A.; Philbrick, K.; Punreddy, S.; Rigby, S.; Smith, R.; Feng, Q.; et al. Process development for expansion of human mesenchymal stromal cells in a 50 L single-use stirred tank bioreactor. Biochem. Eng. J. 2017, 120, 49–62. [Google Scholar] [CrossRef]
- Kehoe, D.; Niss, K.; Rook, M.; Punreddy, S.; Murrel, J.; Sunil, N.; Aysola, M.; Jing, D. Growth Kinetics of Human Mesenchymal Stem Cells in a 3-L Single-Use, Stirred-Tank Bioreactor. Biopharm. Int. 2013, 26, 28–38. [Google Scholar]
- Mizukami, A.; Fernandes-Platzgummer, A.; Carmelo, J.G.; Swiech, K.; Covas, D.T.; Cabral, J.M.S.; da Silva, C.L. Stirred tank bioreactor culture combined with serum-/xenogeneic-free culture medium enables an efficient expansion of umbilical cord-derived mesenchymal stem/stromal cells. Biotechnol. J. 2016, 11, 1048–1059. [Google Scholar] [CrossRef] [PubMed]
- Croughan, M.; Giroux, D.; Fang, D.; Lee, B. Novel single-use bioreactors for scale-up of Anchorage-dependent cell manufacturing for cell therapies. In Stem Cell Manufacturing; Cabral, J.M.S., Chase, de Silva, C.L., Diogo, M.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 105–139. [Google Scholar]
- Cabral, J.M.S.; da Silva, C.L. Bioreactors for Stem Cell Expansion and Differentiation, 1st ed.; Atala, A., Almeida-Porada, G., Eds.; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Pinto, D.D.S.; Bandeiras, C.; Fuzeta, M.D.A.; Rodrigues, C.A.V.; Jung, S.; Hashimura, Y.; Tseng, R.; Milligan, W.; Lee, B.; Ferreira, F.C.; et al. Scalable Manufacturing of Human Mesenchymal Stromal Cells in the Vertical-Wheel Bioreactor System: An Experimental and Economic Approach. Biotechnol. J. 2019, 14, 1800716. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Giroux, D.; Hashimura, Y.; Starkweather, N.; Rosello, F.; Wesselschmidt, R.; Croughan, M. New Scalable Manufacturing Platform for Shear-Sensitive Cell Therapy Products. Cytotherapy 2016, 18, S140. [Google Scholar] [CrossRef]
- Sousa, M.; Silva, M.M.; Giroux, D.; Hashimura, Y.; Wesselschmidt, R.; Lee, B.; Roldão, A.; Carrondo, M.J.T.; Alves, P.M.; Serra, M. Production of oncolytic adenovirus and human mesenchymal stem cells in a single-use, Vertical-Wheel bioreactor system: Impact of bioreactor design on performance of microcarrier-based cell culture processes. Biotechnol. Prog. 2015, 31, 1600–1612. [Google Scholar] [CrossRef]
- Shahdadfar, A.; Frønsdal, K.; Haug, T.; Reinholt, F.P.; Brinchmann, J.E. In vitro expansion of human mesenchymal stem cells: Choice of serum is a determinant of cell proliferation, differentiation, gene expression, and transcriptome stability. Stem Cells 2005, 23, 1357–1366. [Google Scholar] [CrossRef]
- Cobo, F.; Stacey, G.N.; Hunt, C.; Cabrera, C.; Nieto, A.; Montes, R.; Cortés, J.L.; Catalina, P.; Barnie, A.; Concha, Á. Microbiological control in stem cell banks: Approaches to standardisation. Appl. Microbiol. Biotechnol. 2005, 68, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, Z.-B.; Song, Y.-P.; Han, Z.C. Safety of mesenchymal stem cells for clinical application. Stem Cells Int. 2012, 2012, 652034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stute, N.; Holtz, K.; Bubenheim, M.; Lange, C.; Blake, F.; Zander, A.R. Autologous serum for isolation and expansion of human mesenchymal stem cells for clinical use. Exp. Hematol. 2004, 32, 1212–1225. [Google Scholar] [CrossRef] [PubMed]
- Capelli, C.; Domenghini, M.; Borleri, G.; Bellavita, P.; Poma, R.; Carobbio, A.; Micò, C.; Rambaldi, A.; Golay, J.; Introna, M.; et al. Human platelet lysate allows expansion and clinical grade production of mesenchymal stromal cells from small samples of bone marrow aspirates or marrow filter washouts. Bone Marrow Transpl. 2007, 40, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Sen, A.; Rosenberg, L.; Behie, L.A. Identification of growth and attachment factors for the serum-free isolation and expansion of human mesenchymal stromal cells. Cytotherapy 2010, 12, 637–657. [Google Scholar] [CrossRef]
- Goh, T.K.-P.; Zhang, Z.; Chen, A.K.-L.; Reuveny, S.; Choolani, M.; Chan, J.; Oh, S.K.-W. Microcarrier culture for efficient expansion and osteogenic differentiation of human fetal mesenchymal stem cells. BioRes. Open Access 2013, 2, 84–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frauenschuh, S.; Reichmann, E.; Ibold, Y.; Goetz, P.M.; Sittinger, M.; Ringe, J. A microcarrier-based cultivation system for expansion of primary mesenchymal stem cells. Biotechnol. Prog. 2007, 23, 187–193. [Google Scholar] [CrossRef]
- Santos, F.; Andrade, P.Z.; Abecasis, M.M.; Gimble, J.M.; Chase, L.G.; Campbell, A.M.; Boucher, S.; Vemuri, M.C.; Da Silva, C.L.; Cabral, J.M.S. Toward a clinical-grade expansion of mesenchymal stem cells from human sources: A microcarrier-based culture system under xeno-free conditions. Tissue Eng. Part C Methods 2011, 17, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Heathman, T.R.J.; Glyn, V.A.M.; Picken, A.; Rafiq, Q.A.; Coopman, K.; Nienow, A.W.; Kara, B.; Hewitt, C. Expansion, harvest and cryopreservation of human mesenchymal stem cells in a serum-free microcarrier process. Biotechnol. Bioeng. 2015, 112, 1696–1707. [Google Scholar] [CrossRef] [Green Version]
- Nienow, A.; Rafiq, Q.A.; Coopman, K.; Hewitt, C. A potentially scalable method for the harvesting of hMSCs from microcarriers. Biochem. Eng. J. 2014, 85, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Rowley, J.A.; Montgomery, S.A. The Need for Adherent Cell Manufacturing: Production Platform and Media Strategies Drive Cell Production Economics. BioProcess Int. 2018, 16, 34–49. [Google Scholar]
- The Glossary for Cell & Gene Therapy and Regenerative Medicine. Regen. Med. 2018, 13, 1–124.
- Gjoka, X.; Gantier, R.; Schofield, M. Going from Fed-Batch to Perfusion. Biopharm. Int. 2017, 30, 32. [Google Scholar]
- Abraham, E.; Gupta, S.; Jung, S.; McAfee, E. Bioreactor for Scale-Up: Process Control, in Mesenchymal Stromal Cells: Translational Pathways to Clinical Adoption; Viswanathan, S., Hematti, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Krampera, M.; Galipeau, J.; Shi, Y.; Tarte, K.; Sensebé, L. Immunological characterization of multipotent mesenchymal stromal cells—The International Society for Cellular Therapy (ISCT) working proposal. Cytotherapy 2013, 15, 1054–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- François, M.; Galipeau, J. New insights on translational development of mesenchymal stromal cells for suppressor therapy. J. Cell. Physiol. 2012, 227, 3535–3538. [Google Scholar] [CrossRef] [PubMed]
- Meisel, R.P.; Zibert, A.; Laryea, M.; Gobel, U.; Daubener, W.; Dilloo, D. Human bone marrow stromal cells inhibit allogeneic T-cell responses by indoleamine 2,3-dioxygenase-mediated tryptophan degradation. Blood 2004, 103, 4619–4621. [Google Scholar] [CrossRef] [Green Version]
- Boomsma, R.A.; Geenen, D.L. Mesenchymal stem cells secrete multiple cytokines that promote angiogenesis and have contrasting effects on chemotaxis and apoptosis. PLoS ONE 2012, 7, e35685. [Google Scholar] [CrossRef] [Green Version]
- Duffy, G.; Ahsan, T.; O’Brien, T.; Barry, F.; Nerem, R.M. Bone marrow-derived mesenchymal stem cells promote angiogenic processes in a time- and dose-dependent manner in vitro. Tissue Eng. Part A 2009, 15, 2459–2470. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.M.; Hur, S.-M.; Park, K.-Y.; Kim, C.; Kim, Y.-M.; Kim, H.-S.; Shin, H.-C.; Won, M.H.; Ha, K.-S.; Kwon, Y.-G.; et al. Multiple paracrine factors secreted by mesenchymal stem cells contribute to angiogenesis. Vasc. Pharm. 2014, 63, 19–28. [Google Scholar] [CrossRef]
- Ferrari, C.; Balandras, F.; Guedon, E.; Olmos, E.; Chevalot, I.; Marc, A. Limiting cell aggregation during mesenchymal stem cell expansion on microcarriers. Biotechnol. Prog. 2012, 28, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Caruso, S.R.; Orellana, M.D.; Mizukami, A.; Fernandes, T.R.; Fontes, A.; Suazo, C.A.T.; Oliveira, V.D.C.; Covas, D.T.; Swiech, K. Growth and functional harvesting of human mesenchymal stromal cells cultured on a microcarrier-based system. Biotechnol. Prog. 2014, 30, 889–895. [Google Scholar] [CrossRef]
- Bartosh, T.J.; Ylostalo, J.; Mohammadipoor, A.; Bazhanov, N.; Coble, K.; Claypool, K.; Lee, R.H.; Choi, H.; Prockop, D.J. Aggregation of human mesenchymal stromal cells (MSCs) into 3D spheroids enhances their antiinflammatory properties. Proc. Natl. Acad. Sci. USA 2010, 107, 13724–13729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jossen, V.; Schirmer, C.; Sindi, D.M.; Eibl, R.; Kraume, M.; Pörtner, R.; Eibl, D. Theoretical and Practical Issues That Are Relevant When Scaling Up hMSC Microcarrier Production Processes. Stem Cells Int. 2016, 2016, 4760414. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.C.F.; Wong, K.T.K.; Goh, L.T.; Heng, C.-K.; Yap, M.G.S. Impact of dynamic online fed-batch strategies on metabolism, productivity and N-glycosylation quality in CHO cell cultures. Biotechnol. Bioeng. 2005, 89, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Ofiteru, I.D.; Lavric, V.; Woinaroschy, A. A sensitivity analysis of the fed-batch animal-cell bioreactor with respect to some control parameters. Biotechnol. Appl. Biochem. 2005, 41 Pt 1, 29–35. [Google Scholar] [CrossRef]
- Ohlson, S.; Branscomb, J.; Nilsson, K. Bead-to-bead transfer of Chinese hamster ovary cells using macroporous microcarriers. Cytotechnology 1994, 14, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ouyang, F. Bead-to-bead transfer of Vero cells in microcarrier culture. Cytotechnology 1999, 31, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Leber, J.; Barekzai, J.; Blumenstock, M.; Pospisil, B.; Salzig, D.; Czermak, P. Microcarrier choice and bead-to-bead transfer for human mesenchymal stem cells in serum-containing and chemically defined media. Process. Biochem. 2017, 59, 255–265. [Google Scholar] [CrossRef]
- Rafiq, Q.A.; Ruck, S.; Hanga, M.P.; Heathman, T.R.; Coopman, K.; Nienow, A.W.; Williams, D.J.; Hewitt, C. Qualitative and quantitative demonstration of bead-to-bead transfer with bone marrow-derived human mesenchymal stem cells on microcarriers: Utilising the phenomenon to improve culture performance. Biochem. Eng. J. 2018, 135, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Panchalingam, K.M.; Jung, S.; Rosenberg, L.; Behie, L.A. Bioprocessing strategies for the large-scale production of human mesenchymal stem cells: A review. Stem Cell Res. Ther. 2015, 6, 225–265. [Google Scholar] [CrossRef] [Green Version]
- Nienow, A.W.; Coopman, K.; Heathman, T.R.J.; Rafiq, Q.A.; Hewitt, C.J. Bioreactor Engineering Fundamentals for Stem Cell Manufacturing. In Stem Cell Manufacturing; Cabral, J.M.S., de Silva, C.L., Chase, L.G., Diogo, M.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 43–75. [Google Scholar]
- Chisti, Y. Animal-cell damage in sparged bioreactors. Trends Biotechnol. 2000, 18, 420–432. [Google Scholar] [CrossRef]
- Zhang, S.; Handa-Corrigan, A.; Spier, R. Foaming and media surfactant effects on the cultivation of animal cells in stirred and sparged bioreactors. J. Biotechnol. 1992, 25, 289–306. [Google Scholar] [CrossRef]
- Leung, H.W.; Chen, A.; Choo, A.; Reuveny, S.; Oh, S.K.-W. Agitation can induce differentiation of human pluripotent stem cells in microcarrier cultures. Tissue Eng. Part C Methods 2011, 17, 165–172. [Google Scholar] [CrossRef]
- Lecina, M.; Ting, S.; Choo, A.; Reuveny, S.; Oh, S. Scalable platform for human embryonic stem cell differentiation to cardiomyocytes in suspended microcarrier cultures. Tissue Eng. Part C Methods 2010, 16, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, C.; Lee, K.; Nienow, A.W.; Thomas, R.; Smith, M.; Thomas, C.R. Expansion of human mesenchymal stem cells on microcarriers. Biotechnol. Lett. 2011, 33, 2325–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, C.; Pohl, S.; Pörtner, R.; Wallrapp, C.; Kassem, M.; Geigle, P.; Czermak, P. Expansion and Harvesting of hMSC-TERT. Open Biomed. Eng. J. 2007, 1, 38–46. [Google Scholar] [CrossRef]
- Heng, B.C.; Cowan, C.M.; Basu, S. Comparison of enzymatic and non-enzymatic means of dissociating adherent monolayers of mesenchymal stem cells. Biol. Proced. Online 2009, 11, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, K.; Ojima, M.; Otabe, K.; Horie, M.; Koga, H.; Sekiya, I.; Muneta, T. Effects of Different Cell-Detaching Methods on the Viability and Cell Surface Antigen Expression of Synovial Mesenchymal Stem Cells. Cell Transpl. 2017, 26, 1089–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potapova, I.A.; Brink, P.R.; Cohen, I.S.; Doronin, S.V. Culturing of human mesenchymal stem cells as three-dimensional aggregates induces functional expression of CXCR4 that regulates adhesion to endothelial cells. J. Biol. Chem. 2008, 283, 13100–13107. [Google Scholar] [CrossRef] [Green Version]
- Pattasseril, J.; Varadaraju, H.; Lock, L.; Rowley, J.A. Downstream technology landscape for large-scale therapeutic cell processing. Bioprocess. Int. 2013, 11, 38–47. [Google Scholar]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.-C.; Jeske, R.; Chen, X.; Yuan, X.; Li, Y. Influence of Microenvironment on Mesenchymal Stem Cell Therapeutic Potency: From Planar Culture to Microcarriers. Front. Bioeng. Biotechnol. 2020, 8, 640. [Google Scholar] [CrossRef]
- Rowley, J. Developing Cell Therapy Biomanufacturing Processes. Chem. Eng. Process. 2010, 106, 50–55. [Google Scholar]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lembong, J.; Kirian, R.; Takacs, J.D.; Olsen, T.R.; Lock, L.T.; Rowley, J.A.; Ahsan, T. Bioreactor Parameters for Microcarrier-Based Human MSC Expansion under Xeno-Free Conditions in a Vertical-Wheel System. Bioengineering 2020, 7, 73. https://doi.org/10.3390/bioengineering7030073
Lembong J, Kirian R, Takacs JD, Olsen TR, Lock LT, Rowley JA, Ahsan T. Bioreactor Parameters for Microcarrier-Based Human MSC Expansion under Xeno-Free Conditions in a Vertical-Wheel System. Bioengineering. 2020; 7(3):73. https://doi.org/10.3390/bioengineering7030073
Chicago/Turabian StyleLembong, Josephine, Robert Kirian, Joseph D. Takacs, Timothy R. Olsen, Lye Theng Lock, Jon A. Rowley, and Tabassum Ahsan. 2020. "Bioreactor Parameters for Microcarrier-Based Human MSC Expansion under Xeno-Free Conditions in a Vertical-Wheel System" Bioengineering 7, no. 3: 73. https://doi.org/10.3390/bioengineering7030073