The Effects of Lactose Induction on a Plasmid-Free E. coli T7 Expression System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Transformation with pET-21 d (+)
2.3. Bioreactor Cultivations
2.4. Feed Control Strategy
- µ specific growth rate on glucose/lactose [h−1]
- qs specific glucose/lactose uptake rate [gx/gs/h]
- V reactor volume [L]
- cx biomass concentration [gx/L]
- t time [h]
- cs concentration of the feed [gs/L]
2.5. Sampling
2.6. Product Analytics
2.7. Leakiness (Alkaline Phosphatase Assay)
2.8. Lysis (Picogreen Assay)
3. Results and Discussion
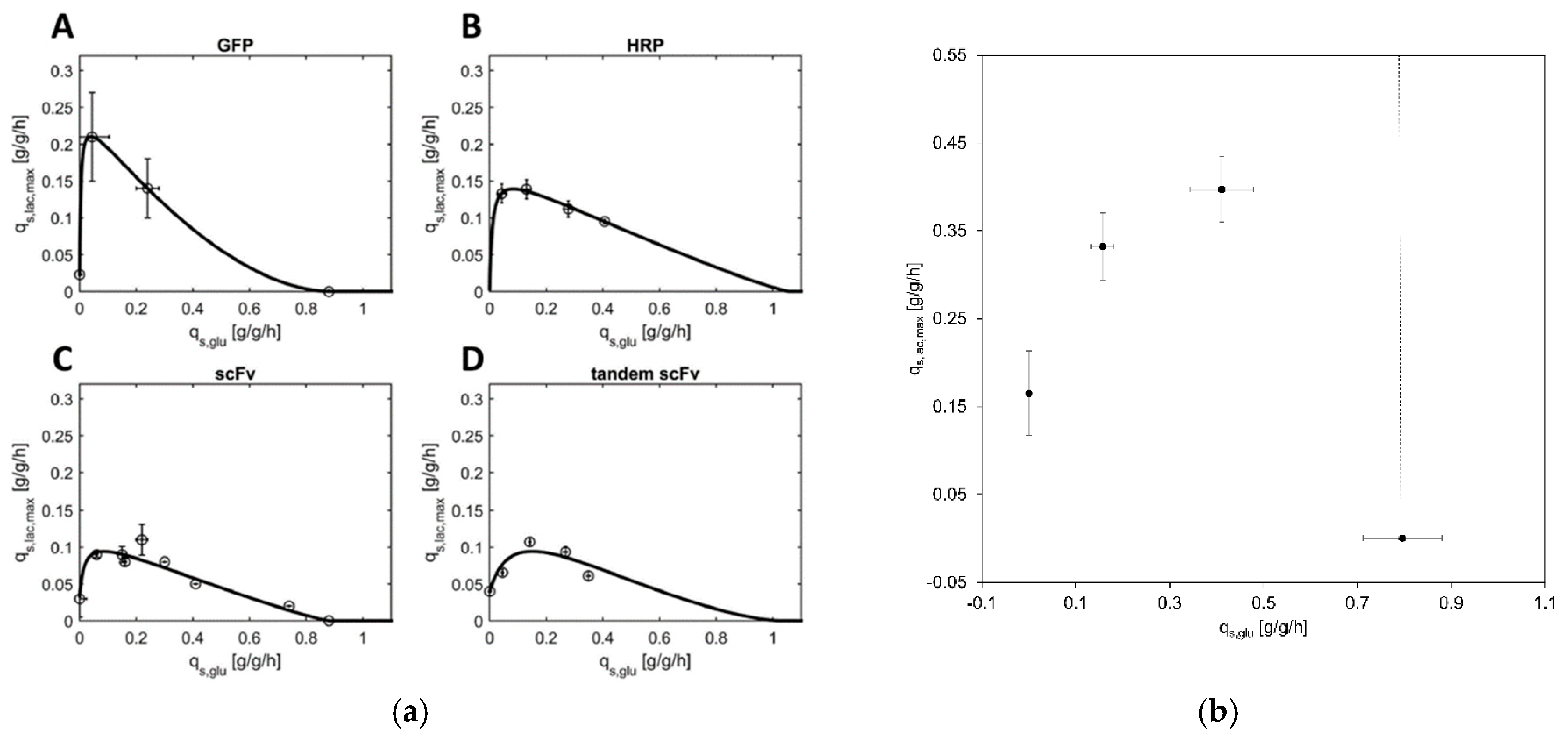
3.1. Correlation between qs,glu and qs,lac,max
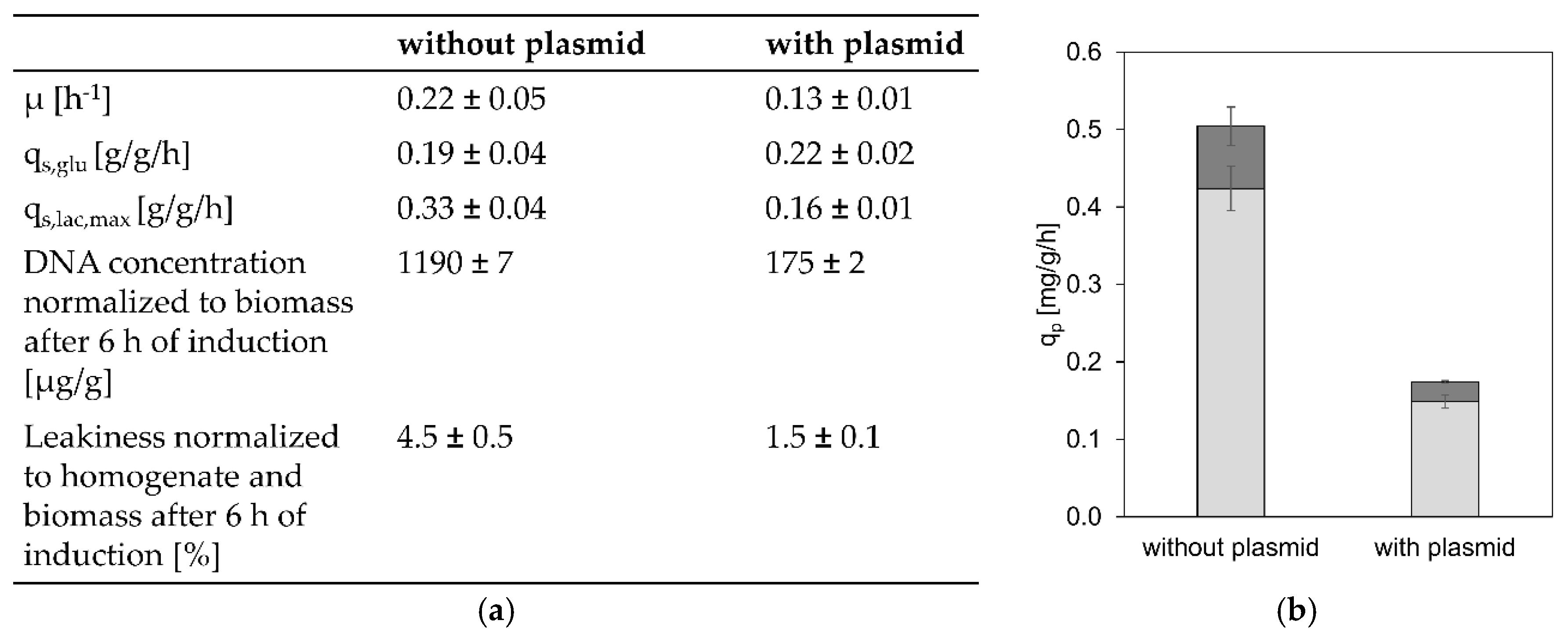
3.2. The Influence of LacI
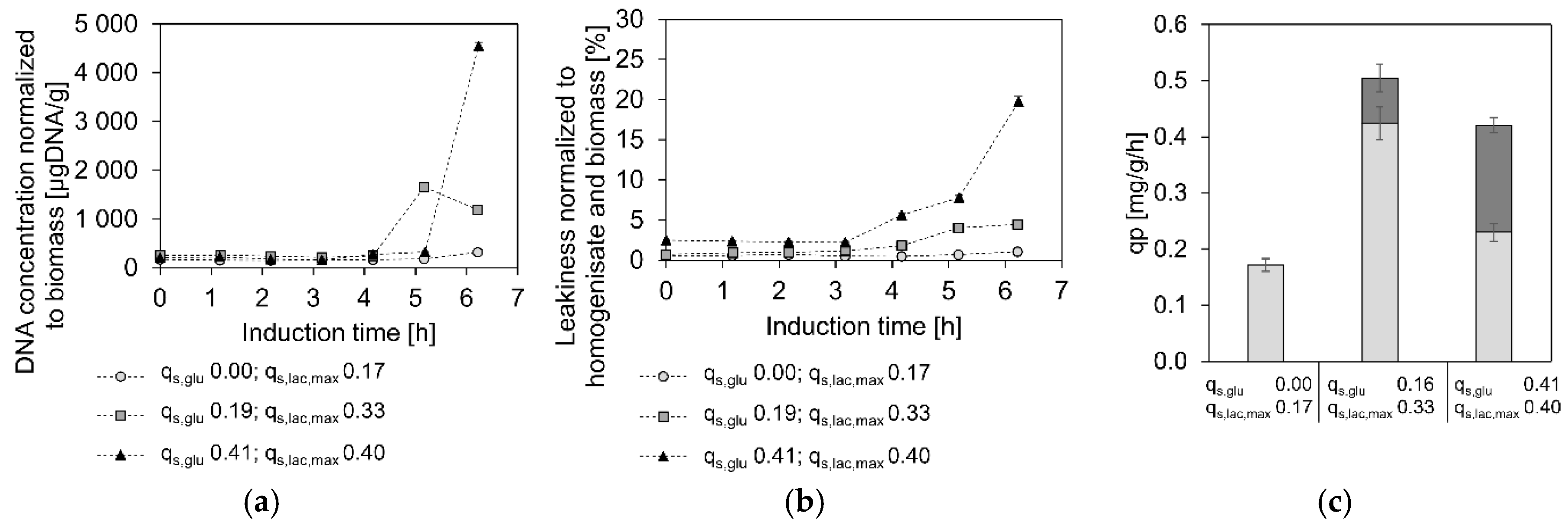
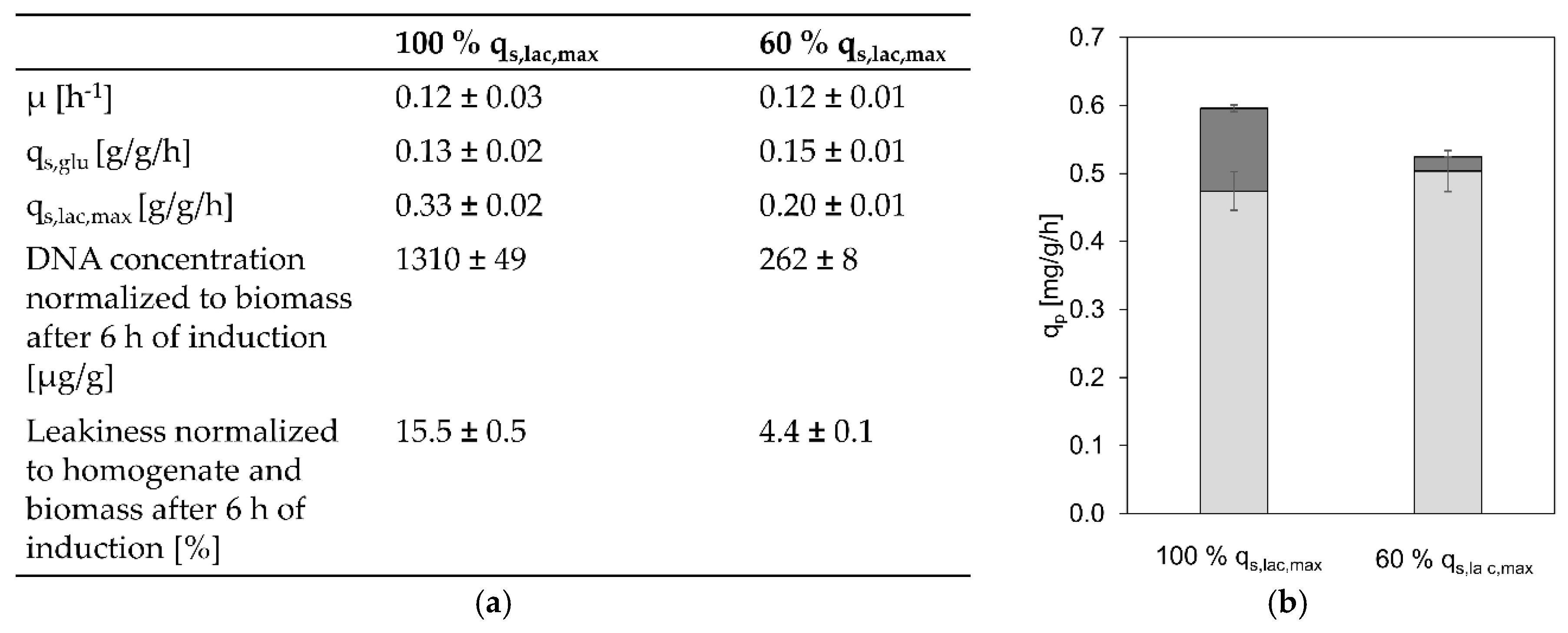
3.3. Toxicity of High Lactose Uptake Rates
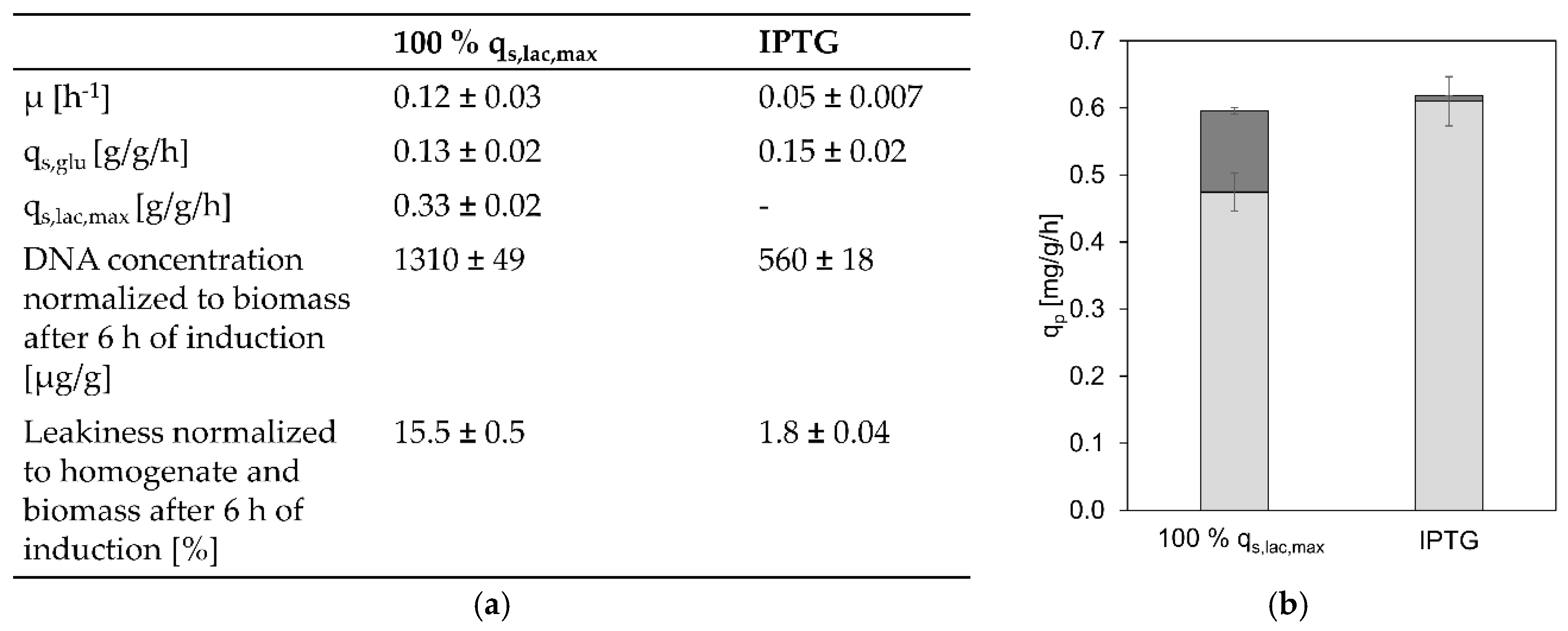
3.4. Comparison between Lactose Induction and IPTG Induction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jia, B.; Jeon, C.O. High-throughput recombinant protein expression in Escherichia coli: Current status and future perspectives. Open Biol. 2016, 6, 160196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørensen, H.P.; Mortensen, K.K. Advanced genetic strategies for recombinant protein expression in Escherichia coli. J. Biotechnol. 2005, 115, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sezonov, G.; Joseleau-Petit, D.; D’Ari, R. Escherichia coli physiology in Luria-Bertani broth. J. Bacteriol. 2007, 189, 8746–8749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daegelen, P.; Studier, F.W.; Lenski, R.E.; Cure, S.; Kim, J.F. Tracing Ancestors and Relatives of Escherichia coli B, and the Derivation of B Strains REL606 and BL21(DE3). J. Mol. Biol. 2009, 394, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Fulcrand, G.; Dages, S.; Zh, X.; Chapagain, P.; Gerstman, B.S.; Dunlap, D.; Leng, F. DNA supercoiling, a critical signal regulating the basal expression of the lac operon in Escherichia coli. Sci. Rep. 2016, 6, 19243. [Google Scholar] [CrossRef] [Green Version]
- Studier, F.W.; Moffatt, B.A. Use of Bacteriophage-T7 Rna-Polymerase to Direct Selective High-Level Expression of Cloned Genes. J. Mol. Biol. 1986, 189, 113–130. [Google Scholar] [CrossRef]
- Studier, F.W. Use of bacteriophage T7 lysozyme to improve an inducible T7 expression system. J. Mol. Biol. 1991, 219, 37–44. [Google Scholar] [CrossRef]
- Dubendorff, J.W.; Studier, F.W. Controlling basal expression in an inducible T7 expression system by blocking the target T7 promoter with lac repressor. J. Mol. Biol. 1991, 219, 45–59. [Google Scholar] [CrossRef]
- Popov, M.; Petrov, S.; Nacheva, G.; Ivanov, I.; Reichl, U. Effects of a recombinant gene expression on ColE1-like plasmid segregation in Escherichia coli. BMC Biotechnol. 2011, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Teich, A.; Lin, H.Y.; Andersson, L.; Meyer, S.; Neubauer, P. Amplification of ColE1 related plasmids in recombinant cultures of Escherichia coli after IPTG induction. J. Biotechnol. 1998, 64, 197–210. [Google Scholar] [CrossRef]
- Juhas, M.; Ajioka, J.W. Lambda Red recombinase-mediated integration of the high molecular weight DNA into the Escherichia coli chromosome. Microb. Cell Fact. 2016, 15, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juhas, M.; Ajioka, J.W. Flagellar region 3b supports strong expression of integrated DNA and the highest chromosomal integration efficiency of the Escherichia coli flagellar regions. Microb. Biotechnol. 2015, 8, 726–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Striedner, G.; Pfaffenzeller, I.; Markus, L.; Nemecek, S.; Grabherr, R.; Bayer, K. Plasmid-free T7-based Escherichia coli expression systems. Biotechnol. Bioeng. 2010, 105, 786–794. [Google Scholar]
- Fink, M.; Vazulka, S.; Egger, E.; Jarmer, J.; Grabherr, R.; Cserjan-Puschmann, M.; Striedner, G. Micro-bioreactor cultivations of Fab producing Escherichia coli reveal genome-integrated systems as suitable for prospective studies on direct Fab expression effects. Biotechnol. J. 2019, 14, e1800637. [Google Scholar] [CrossRef] [Green Version]
- St-Pierre, F.; Cui, L.; Priest, D.G.; Endy, D.; Dodd, I.B.; Shearwin, K.E. One-Step Cloning and Chromosomal Integration of DNA. ACS Synth. Biol. 2013, 2, 537–541. [Google Scholar] [CrossRef]
- Dvorak, P.; Chrast, L.; Nikel, P.I.; Fedr, R.; Soucek, K.; Sedlackova, M.; Chaloupkova, R.; de Lorenzo, V.; Prokop, Z.; Damborsky, J. Exacerbation of substrate toxicity by IPTG in Escherichia coli BL21(DE3) carrying a synthetic metabolic pathway. Microb. Cell Fact. 2015, 14, 201. [Google Scholar] [CrossRef] [Green Version]
- Nausch, H.; Huckauf, J.; Koslowski, R.; Meyer, U.; Broer, I.; Mikschofsky, H. Recombinant production of human interleukin 6 in Escherichia coli. PLoS ONE 2013, 8, e54933. [Google Scholar] [CrossRef] [Green Version]
- Gombert, A.K.; Kilikian, B.V. Recombinant gene expression in Escherichia coli cultivation using lactose as inducer. J. Biotechnol. 1998, 60, 47–54. [Google Scholar] [CrossRef]
- Wurm, D.J.; Quehenberger, J.; Mildner, J.; Eggenreich, B.; Slouka, C.; Schwaighofer, A.; Wieland, K.; Lendl, B.; Rajamanickam, V.; Herwig, C.; et al. Teaching an old pET new tricks: Tuning of inclusion body formation and properties by a mixed feed system in E. coli. Appl. Microbiol. Biotechnol. 2018, 102, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Fruchtl, M.; Sakon, J.; Beitle, R. Expression of a collagen-binding domain fusion protein: Effect of amino acid supplementation, inducer type, and culture conditions. Biotechnol. Prog. 2015, 31, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Bashir, H.; Ahmed, N.; Khan, M.A.; Zafar, A.U.; Tahir, S.; Khan, M.I.; Khan, F.; Husnain, T. Simple procedure applying lactose induction and one-step purification for high-yield production of rhCIFN. Biotechnol. Appl. Biochem. 2016, 63, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Wurm, D.J.; Hausjell, J.; Ulonska, S.; Herwig, C.; Spadiut, O. Mechanistic platform knowledge of concomitant sugar uptake in Escherichia coli BL21(DE3) strains. Sci. Rep. 2017, 7, 45072. [Google Scholar] [CrossRef] [PubMed]
- Wurm, D.J.; Veiter, L.; Ulonska, S.; Eggenreich, B.; Herwig, C.; Spadiut, O. The E. coli pET expression system revisited-mechanistic correlation between glucose and lactose uptake. Appl. Microbiol. Biotechnol. 2016, 100, 8721–8729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurm, D.J.; Marschall, L.; Sagmeister, P.; Herwig, C.; Spadiut, O. Simple monitoring of cell leakiness and viability in Escherichia coli bioprocesses—A case study. Eng. Life Sci. 2017, 17, 598–604. [Google Scholar] [CrossRef] [Green Version]
- Sharan, S.K.; Thomason, L.C.; Kuznetsov, S.G.; Court, D.L. Recombineering: A homologous recombination-based method of genetic engineering. Nat. Protoc. 2009, 4, 206–223. [Google Scholar] [CrossRef] [Green Version]
- DeLisa, M.P.; Li, J.; Rao, G.; Weigand, W.A.; Bentley, W.E. Monitoring GFP-operon fusion protein expression during high cell density cultivation of Escherichia coli using an on-line optical sensor. Biotechnol. Bioeng. 1999, 65, 54–64. [Google Scholar] [CrossRef]
- Janzen, N.H.; Striedner, G.; Jarmer, J.; Voigtmann, M.; Abad, S.; Reinisch, D. Implementation of a Fully Automated Microbial Cultivation Platform for Strain and Process Screening. Biotechnol. J. 2019, 14, e1800625. [Google Scholar] [CrossRef] [Green Version]
- Kremling, A.; Bettenbrock, K.; Laube, B.; Jahreis, K.; Lengeler, J.W.; Gilles, E.D. The organization of metabolic reaction networks. III. Application for diauxic growth on glucose and lactose. Metab. Eng. 2001, 3, 362–379. [Google Scholar] [CrossRef]
- Kremling, A.; Geiselmann, J.; Ropers, D.; de Jong, H. Understanding carbon catabolite repression in Escherichia coli using quantitative models. Trends Microbiol. 2015, 23, 99–109. [Google Scholar] [CrossRef]
- Loomis, W.F.J.; Magasanik, B. Glucose-lactose diauxie in Escherichia coli. J. Bacteriol. 1967, 93, 1397–1401. [Google Scholar] [CrossRef] [Green Version]
- Dykhuizen, D.; Hartl, D. Transport by the lactose permease of Escherichia coli as the basis of lactose killing. J. Bacteriol. 1978, 135, 876–882. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.M.; Putzrath, R.M.; Wilson, T.H. Inhibition of growth of Escherichia coli by lactose and other galactosides. Biochim. Biophys. Acta 1981, 649, 377–384. [Google Scholar] [CrossRef]
- Kopp, J.; Slouka, C.; Ulonska, S.; Kager, J.; Fricke, J.; Spadiut, O.; Herwig, C. Impact of Glycerol as Carbon Source onto Specific Sugar and Inducer Uptake Rates and Inclusion Body Productivity in E. coli BL21(DE3). Bioengineering 2017, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, H.; Barbe, V.; Lee, C.H.; Vallenet, D.; Yu, D.S.; Choi, S.H.; Couloux, A.; Lee, S.W.; Yoon, S.H.; Cattolico, L.; et al. Genome sequences of Escherichia coli B strains REL606 and BL21(DE3). J. Mol. Biol. 2009, 394, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Sina, M.; Farajzadeh, D.; Dastmalchi, S. Effects of Environmental Factors on Soluble Expression of a Humanized Anti-TNF-α scFv Antibody in Escherichia coli. Adv. Pharm. Bull. 2015, 5, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, X.-L.; Wang, Q.-Y.; Li, C.-L.; Qiu, X.-F.; Xie, K.-L.; Huang, L.-F.; Wang, A.-M.; Zeng, Z.-W.; Xie, T. Efficient Production of a Thermophilic 2-Deoxyribose-5-Phosphate Aldolase in Glucose-Limited Fed-Batch Cultivations of Escherichia coli by Continuous Lactose Induction Strategy. Appl. Biochem. Biotechnol. 2011, 165, 416–425. [Google Scholar] [CrossRef]
- Tian, H.; Tang, L.; Wang, Y.; Wang, X.; Guan, L.; Zhang, J.; Wu, X.; Li, X. Lactose Induction Increases Production of Recombinant Keratinocyte Growth Factor-2 in Escherichia coli. Int. J. Pept. Res. Ther. 2011, 17, 123–129. [Google Scholar] [CrossRef]
- Huang, L.; Wang, Q.; Jiang, S.; Zhou, Y.; Zhang, G.; Ma, Y. Improved extracellular expression and high-cell-density fed-batch fermentation of chitosanase from Aspergillus Fumigatus in Escherichia coli. Bioprocess Biosyst. Eng. 2016, 39, 1679–1687. [Google Scholar] [CrossRef]
- Kim, S.; Cheung, L.H.; Zhang, W.; Rosenblum, M.G. Improved expression of a soluble single chain antibody fusion protein containing tumor necrosis factor in Escherichia coli. Appl. Microbiol. Biotechnol. 2007, 77, 99–106. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hausjell, J.; Kutscha, R.; Gesson, J.D.; Reinisch, D.; Spadiut, O. The Effects of Lactose Induction on a Plasmid-Free E. coli T7 Expression System. Bioengineering 2020, 7, 8. https://doi.org/10.3390/bioengineering7010008
Hausjell J, Kutscha R, Gesson JD, Reinisch D, Spadiut O. The Effects of Lactose Induction on a Plasmid-Free E. coli T7 Expression System. Bioengineering. 2020; 7(1):8. https://doi.org/10.3390/bioengineering7010008
Chicago/Turabian StyleHausjell, Johanna, Regina Kutscha, Jeannine D. Gesson, Daniela Reinisch, and Oliver Spadiut. 2020. "The Effects of Lactose Induction on a Plasmid-Free E. coli T7 Expression System" Bioengineering 7, no. 1: 8. https://doi.org/10.3390/bioengineering7010008