
Effects of Stroboscopic Vision on Depth Jump Motor Control: A Biomechanical Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedures
2.3. Data Analysis
2.4. Statistical Analysis
3. Results
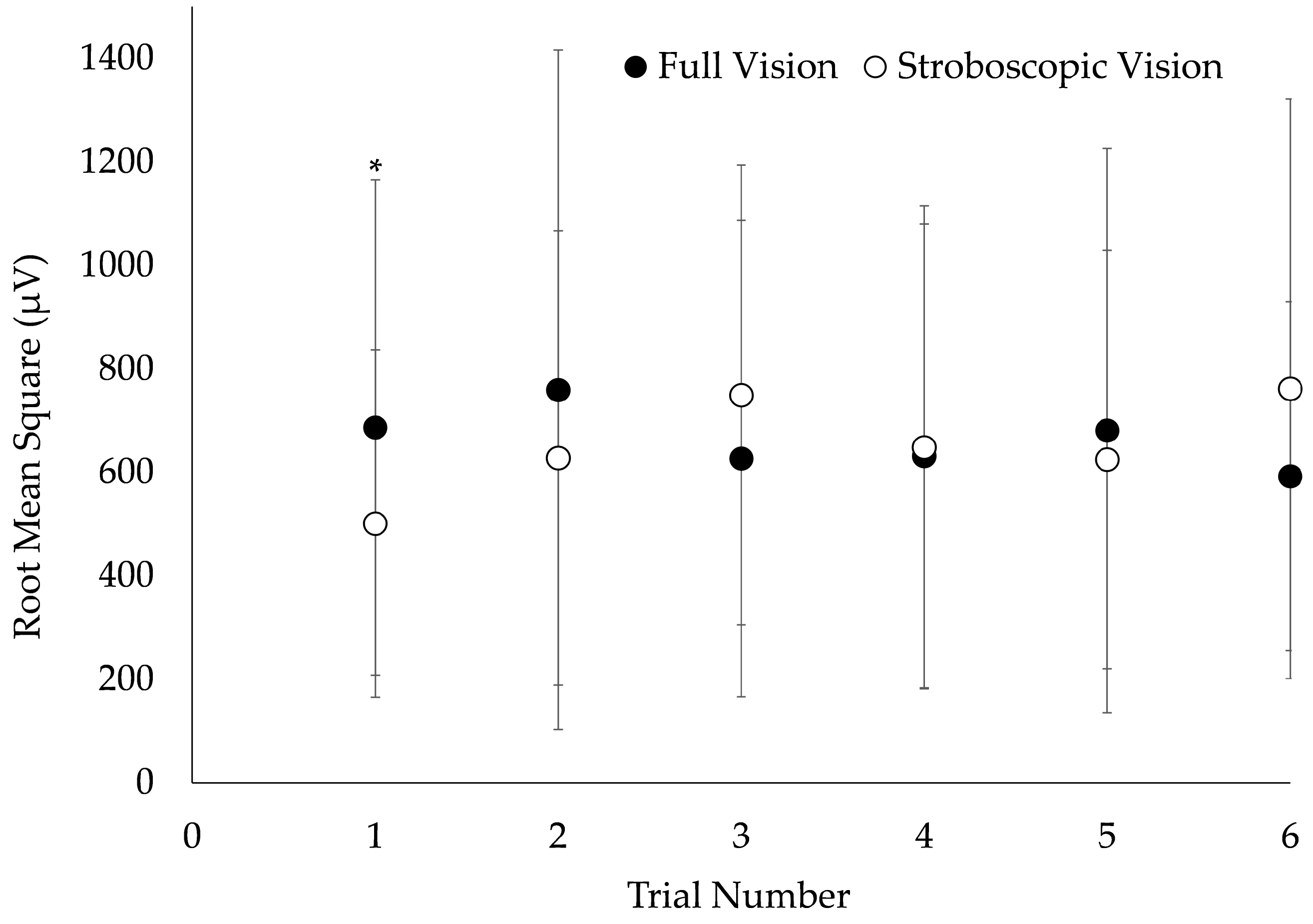
3.1. Inter-Trial Reliability
3.2. RMANOVA
3.2.1. Interactions
3.2.2. Main Effects—GRF
3.2.3. Main Effects—EMG
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Helm, M.; Ritzmann, R.; Gollhofer, A.; Freyler, K. Anticipation modulates neuromechanics of drop jumps in known or unknown ground stiffness. PLoS ONE 2019, 14, e0211276. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.L.; Marusic, U.; Mahoney, J.R. The intersection of cognitive, motor, and sensory processing in aging: Links to functional outcomes. Front. Aging Neurosci. 2022, 14, 1009532. [Google Scholar] [CrossRef]
- Peterka, R.J. Sensory integration for human balance control. Handb. Clin. Neurol. 2018, 159, 27–42. [Google Scholar] [PubMed]
- Kim, K.M.; Kim, J.S.; Oh, J.; Grooms, D.R. Stroboscopic vision as a dynamic sensory reweighting alternative to the sensory organization test. J. Sport Rehabil. 2020, 30, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Anson, E.; Jeka, J. Sensory Reweighting: A Rehabilitative Mechanism? In Handbook of Medical Neuropsychology: Applications of Cognitive Neuroscience; Armstrong, C.L., Morrow, L.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 789–800. ISBN 978-3-030-14895-9. [Google Scholar]
- Kim, K.M.; Kim, J.S.; Grooms, D.R. Stroboscopic vision to induce sensory reweighting during postural control. J. Sport Rehabil. 2017, 26, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Santello, M. Review of motor control mechanisms underlying impact absorption from falls. Gait Posture 2005, 21, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Baudry, S.; Duchateau, J. Age-related influence of vision and proprioception on Ia presynaptic inhibition in soleus muscle during upright stance. J. Physiol. 2012, 590, 5541–5554. [Google Scholar] [CrossRef] [PubMed]
- Haeufle, D.F.; Schmortte, B.; Geyer, H.; Müller, R.; Schmitt, S. The benefit of combining neuronal feedback and feed-forward control for robustness in step down perturbations of simulated human walking depends on the muscle function. Front. Comput. Neurosci. 2018, 12, 80. [Google Scholar] [CrossRef]
- Nashner, L.; Berthoz, A. Visual contribution to rapid motor responses during postural control. Brain Res. 1978, 150, 403–407. [Google Scholar] [CrossRef]
- Liebermann, D.G.; Goodman, D. Pre-landing muscle timing and post-landing effects of falling with continuous vision and in blindfold conditions. J. Electromyogr. Kinesiol. 2007, 17, 212–227. [Google Scholar] [CrossRef]
- Kroll, M.; Preuss, J.; Ness, B.M.; Dolny, M.; Louder, T. Effect of stroboscopic vision on depth jump performance in female NCAA Division I volleyball athletes. Sports Biomech. 2023, 22, 1016–1026. [Google Scholar] [CrossRef] [PubMed]
- Kamibayashi, K.; Muro, M. Modulation of pre-programmed muscle activation and stretch reflex to changes of contact surface and visual input during movement to absorb impact. J. Electromyogr. Kinesiol. 2006, 16, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Shenton, J.T.; Schwoebel, J.; Coslett, H.B. Mental motor imagery and the body schema: Evidence for proprioceptive dominance. Neurosci. Lett. 2004, 370, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Mullineaux, D.R.; Milner, C.E.; Davis, I.S.; Hamill, J. Normalization of ground reaction forces. J. Appl. Biomech. 2006, 22, 230–233. [Google Scholar] [CrossRef]
- Turns, L.J.; Neptune, R.R.; Kautz, S.A. Relationships between muscle activity and anteroposterior ground reaction forces in hemiparetic walking. Arch. Phys. Med. Rehabil. 2007, 88, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Enoka, R.M. Neuromechanics of Human Movement; Human Kinetics: Champaign, IL, USA, 2008. [Google Scholar]
- Santello, M.; McDonagh, M.J.; Challis, J.H. Visual and non-visual control of landing movements in humans. J. Physiol. 2001, 537, 313–327. [Google Scholar] [CrossRef]
- Shin, S.Y.; Crapse, T.B.; Mayo, J.P.; Sommer, M.A. Visuomotor integration. In Encyclopedia of Neuroscience, 1st ed.; Binder, M.D., Hirokawa, N., Windhorst, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 4354–4359. [Google Scholar]
- Garcia, M.C.; Vieira, T.M.M. Surface electromyography: Why, when and how to use it. Rev. Andal. Med. Deporte. 2011, 4, 17–28. [Google Scholar]
- Chaput, M.; Onate, J.A.; Simon, J.E.; Criss, C.R.; Jamison, S.; McNally, M.; Grooms, D.R. Visual cognition associated with knee proprioception, time to stability, and sensory integration neural activity after ACL reconstruction. J. Orthop. Res. 2022, 40, 95–104. [Google Scholar] [CrossRef]
- Knapik, J.J.; Steelman, R.; Grier, T.; Graham, B.; Hoedebecke, K.; Rankin, S.; Klug, K.; Proctor, S.; Jones, B.H. Military parachuting injuries, associated events, and injury risk factors. Aviat. Space Environ. Med. 2011, 82, 797–804. [Google Scholar] [CrossRef]
- Hoffman, J.R. NSCA’s Guide to Program Design; Human Kinetics: Champaign, IL, USA, 2012; p. 51. [Google Scholar]
- Louder, T.; Thompson, B.J.; Banks, N.; Bressel, E. A mixed-methods approach to evaluating the internal validity of the reactive strength index. Sports 2019, 7, 157. [Google Scholar] [CrossRef]
- Louder, T.; Thompson, B.J.; Woster, A.; Bressel, E. Kinetics of depth jumps performed by female and male national collegiate athletics association basketball athletes and young adults. J. Funct. Morphol. Kinesiol. 2023, 8, 108. [Google Scholar] [CrossRef] [PubMed]
- Ambegaonkar, J.P.; Shultz, S.J. Changing filtering parameters affects lower extremity pre-landing muscle activation onset times. Isokinet. Exerc. Sci. 2010, 18, 125–132. [Google Scholar] [CrossRef]
- Mrdakovic, V.; Ilic, D.B.; Jankovic, N.; Rajkovic, Z.; Stefanovic, D. Pre-activity modulation of lower extremity muscles within different types and heights of deep jump. J. Sports Sci. Med. 2008, 7, 269–279. [Google Scholar] [PubMed]
- Han, S.; Son, S.J.; Kim, H.; Lee, H.; Seeley, M.; Hopkins, T. Prelanding movement strategies among chronic ankle instability, coper, and control subjects. Sports Biomech. 2022, 21, 391–407. [Google Scholar] [CrossRef] [PubMed]
- Taube, W.; Schubert, M.; Gruber, M.; Beck, S.; Faist, M.; Gollhofer, A. Direct corticospinal pathways contribute to neuromuscular control of perturbed stance. J. Appl. Physiol. 2006, 101, 420–429. [Google Scholar] [CrossRef]
- Waldvogel, J.; Freyler, K.; Ritzmann, R.; Gollhofer, A. Energy transfer in reactive movements as a function of individual stretch load. Front. Physiol. 2023, 14, 1265443. [Google Scholar] [CrossRef] [PubMed]
- Koo, T.K.; Li, M.Y. A guideline of selecting and reporting intraclass correlation coefficients for reliability research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge Academic: New York, NY, USA, 1988. [Google Scholar]
- Chan, Y.H. Biostatistics 104: Correlational analysis. Singap. Med. J. 2003, 44, 614–619. [Google Scholar]
- Hughes, G.; Watkins, J. Lower limb coordination and stiffness during landing from volleyball block jumps. Res. Sports Med. 2008, 16, 138–154. [Google Scholar] [CrossRef]
- Shih, Y.O.; Teng, H.L.; Powers, C. Lower extremity stiffness predicts ground reaction force loading rate in heel-strike runners. Med. Sci. Sports Exerc. 2019, 51, 1692–1697. [Google Scholar] [CrossRef]
- Duncan, A.D.; McDonagh, M.J. Stretch reflex distinguished from pre-programmed muscle activations following landing impacts in man. J. Physiol. 2000, 526, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Tucker, K.J.; Türker, K.S. Muscle spindle feedback differs between the soleus and gastrocnemius in humans. Somatosens. Mot. Res. 2009, 21, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Ludvig, D.; Cathers, I.; Kearney, R.E. Voluntary modulation of human stretch reflexes. Exp. Brain Res. 2007, 183, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Lamontagne, M.; Kennedy, M.J. The biomechanics of vertical hopping: A review. Res. Sports Med. 2013, 21, 380–394. [Google Scholar] [CrossRef] [PubMed]
- Cordo, P.J. Kinesthetic control of a multijoint movement sequence. J. Neurophysiol. 1990, 63, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Reschechtko, S.; Pruszynski, J.A. Stretch reflexes. Curr. Biol. 2020, 30, R1025–R1030. [Google Scholar] [CrossRef]
- McMahon, J.J.; Lake, J.P.; Stratford, C.; Comfort, P. A proposed method for evaluating drop jump performance with one force platform. Biomechanics 2021, 1, 178–189. [Google Scholar] [CrossRef]
- Polakovičová, M.; Vavák, M.; Ollé, R.; Lehnert, M.; Sigmund, M. Vertical jump development in elite adolescent volleyball players: Effects of sex and age. Acta Gymnica 2018, 48, 115–120. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sex | n | Age (Years) |
|---|---|---|
| Female | 9 | 21.8 (1.7) |
| Male | 11 | 22.9 (1.8) |
| Measure | Full Vision | Stroboscopic Vision |
|---|---|---|
| Peak GRF | 0.637 (0.456–0.808) | 0.478 (0.286–0.697) |
| RFD | 0.587 (0.401–0.775) | 0.643 (0.465–0.812) |
| PFR | 0.619 (0.436–0.797) | 0.696 (0.530–0.844) |
| Measure | Muscle | Full Vision | Stroboscopic Vision |
|---|---|---|---|
| Pre | VM | 0.850 (0.744–0.929) | 0.840 (0.728–0.924) |
| VL | 0.798 (0.667–0.902) | 0.942 (0.895–0.974) | |
| BF | 0.611 (0.428–0.791) | 0.777 (0.636–0.891) | |
| TA | 0.768 (0.625–0.886) | 0.826 (0.708–0.917) | |
| GM | 0.791 (0.656–0.898) | 0.836 (0.722–0.922) | |
| SLR | VM | 0.826 (0.708–0.917) | 0.790 (0.655–0.897) |
| VL | 0.705 (0.541–0.849) | 0.868 (0.772–0.938) | |
| BF | 0.836 (0.723–0.922) | 0.692 (0.524–0.842) | |
| TA | 0.388 (0.202–0.624) | 0.442 (0.252–0.688) | |
| GM | 0.693 (0.461–0.809) | 0.406 (0.220–0.638) | |
| MLR | VM | 0.643 (0.465–0.812) | 0.666 (0.493–0.826) |
| VL | 0.784 (0.647–0.894) | 0.657 (0.482–0.820) | |
| BF | 0.844 (0.736–0.926) | 0.907 (0.835–0.957) | |
| TA | 0.507 (0.318–0.718) | 0.505 (0.314–0.718) | |
| GM | 0.666 (0.493–0.826) | 0.621 (0.440–0.798) | |
| LLR | VM | 0.686 (0.517–0.839) | 0.693 (0.527–0.843) |
| VL | 0.772 (0.630–0.888) | 0.736 (0.582–0.867) | |
| BF | 0.882 (0.794–0.945) | 0.899 (0.822–0.954) | |
| TA | 0.575 (0.388–0.767) | 0.359 (0.177–0.598) | |
| GM | 0.718 (0.559–0.857) | 0.608 (0.425–0.789) |
| Measure | Full Vision | Stroboscopic Vision | Cohen’s d | Pearson r |
|---|---|---|---|---|
| Peak GRF (BW) | 4.66 (1.26) | 4.96 (1.64) * | 0.21 | 0.79 |
| RFD (BW*s-1) | 94.27 (49.74) | 97.34 (43.72) | 0.07 | 0.86 |
| PFR (BW) | 2.28 (1.22) | 2.42 (1.44) | 0.11 | 0.87 |
| Measure | Muscle | Full Vision | Stroboscopic Vision | Cohen’s d | Pearson r |
|---|---|---|---|---|---|
| Pre (μV) | VM | 376.76 (302.92) | 379.30 (340.75) | 0.01 | 0.94 |
| VL | 305.29 (291.25) | 334.17 (369.66) Ұ | 0.09 | 0.96 | |
| BF | 139.32 (198.07) | 147.60 (207.89) | 0.04 | 0.97 | |
| TA | 242.79 (123.46) | 241.34 (145.00) | −0.01 | 0.92 | |
| GM | 225.11 (114.82) | 225.94 (114.77) | 0.00 | 0.98 | |
| SLR (μV) | VM | 846.16 (630.22) | 865.76 (650.52) | 0.03 | 0.95 |
| VL | 613.22 (417.14) | 602.75 (485.01) | −0.02 | 0.94 | |
| BF | 336.75 (487.07) | 393.72 (533.15) | 0.11 | 0.88 | |
| TA | 435.93 (250.43) | 438.72 (233.69) | 0.01 | 0.65 | |
| GM | 172.12 (153.19) | 157.65 (144.89) | −0.10 | 0.77 | |
| MLR (μV) | VM | 663.08 (489.32) | 654.20 (445.49) | −0.02 | 0.91 |
| VL | 522.50 (364.44) | 553.93 (467.56) | 0.08 | 0.91 | |
| BF | 333.83 (604.82) | 387.26 (668.96) Ұ | 0.08 | 0.98 | |
| TA | 455.44 (306.97) | 391.18 (271.92) * | −0.22 | 0.85 | |
| GM | 184.53 (200.94) | 179.99 (208.28) | −0.02 | 0.95 | |
| LLR (μV) | VM | 673.24 (489.39) | 610.51 (430.71) Ұ | −0.14 | 0.91 |
| VL | 518.23 (402.95) | 457.05 (359.28) * | −0.16 | 0.95 | |
| BF | 271.64 (520.39) | 323.03 (669.96) Ұ | 0.08 | 0.98 | |
| TA | 359.53 (279.72) | 341.96 (299.31) | −0.06 | 0.85 | |
| GM | 189.39 (208.99) | 212.81 (265.58) | 0.10 | 0.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harrison, K.D.; Dakin, C.J.; Beethe, A.Z.; Louder, T. Effects of Stroboscopic Vision on Depth Jump Motor Control: A Biomechanical Analysis. Bioengineering 2024, 11, 290. https://doi.org/10.3390/bioengineering11030290
Harrison KD, Dakin CJ, Beethe AZ, Louder T. Effects of Stroboscopic Vision on Depth Jump Motor Control: A Biomechanical Analysis. Bioengineering. 2024; 11(3):290. https://doi.org/10.3390/bioengineering11030290
Chicago/Turabian StyleHarrison, Kenneth D., Christopher J. Dakin, Anne Z. Beethe, and Talin Louder. 2024. "Effects of Stroboscopic Vision on Depth Jump Motor Control: A Biomechanical Analysis" Bioengineering 11, no. 3: 290. https://doi.org/10.3390/bioengineering11030290