
Deep Learning for Generalized EEG Seizure Detection after Hypoxia–Ischemia—Preclinical Validation
 , and
, and
Abstract
:1. Introduction
2. Methods
- (a)
- Can our previous micro-scale EEG pattern classifiers be re-designed for accurate seizure identification in data from fetal sheep models with different gestational ages and/or under the influence of treatment with therapeutic hypothermia?
- (b)
- Can the seizure detection algorithms trained/validated on datasets from certain group sets identify seizures in the EEG sets of other individual groups?
- Study #1: A leave-one-out cross-validation (LOOCV) approach where data from the term sham–normothermia group were included in three different training/test schemes.
- Study #2: A leave-one-out cross-validation (LOOCV) approach where data from the sham–normothermia group were excluded in three different training/test schemes. This was used to study the possible impacts of removing data from the sham–normothermia group.
- Study #3: A k-fold cross-validation (k = 5) approach where data from all groups were randomly combined and included in five different training/test schemes.
- (c)
- Can specific training strategies help to improve the generalization and robustness of pattern classifiers to perform equally well across all groups and identify seizures regardless of what hemisphere the EEG has been recorded from?
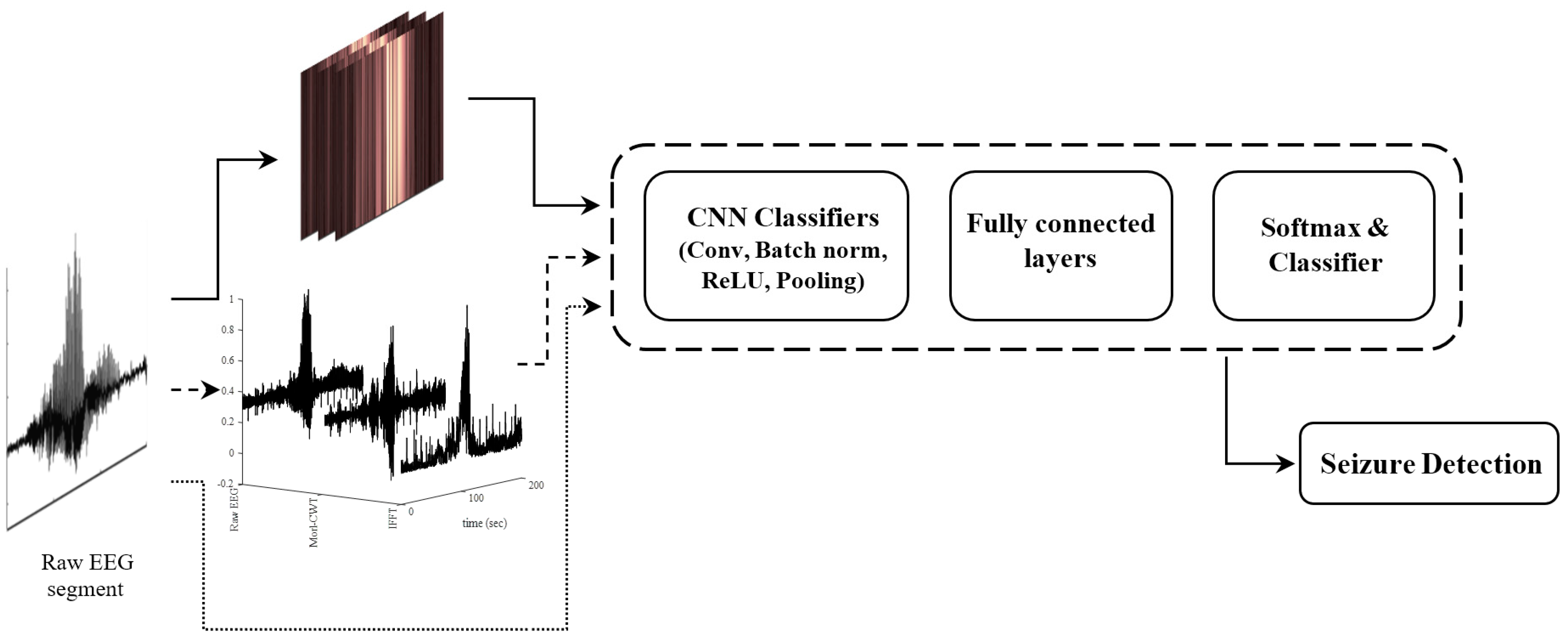
2.1. WS-CNN Seizure Detector
2.2. WF-CNN Seizure Detector
- -
- The CWT coefficients of each EEG segment using morl at an arbitrary scale of 80 (equal to pseudo-frequency of 2.56 Hz). This scale number was chosen to target the embedded spectrums near the mean frequency of the delta-band (0.5–4 Hz).
- -
- -
- The original raw EEG segment.
2.3. 1D-CNN Seizure Detector
2.4. Performance Metrics
2.5. Computing Infrastructure
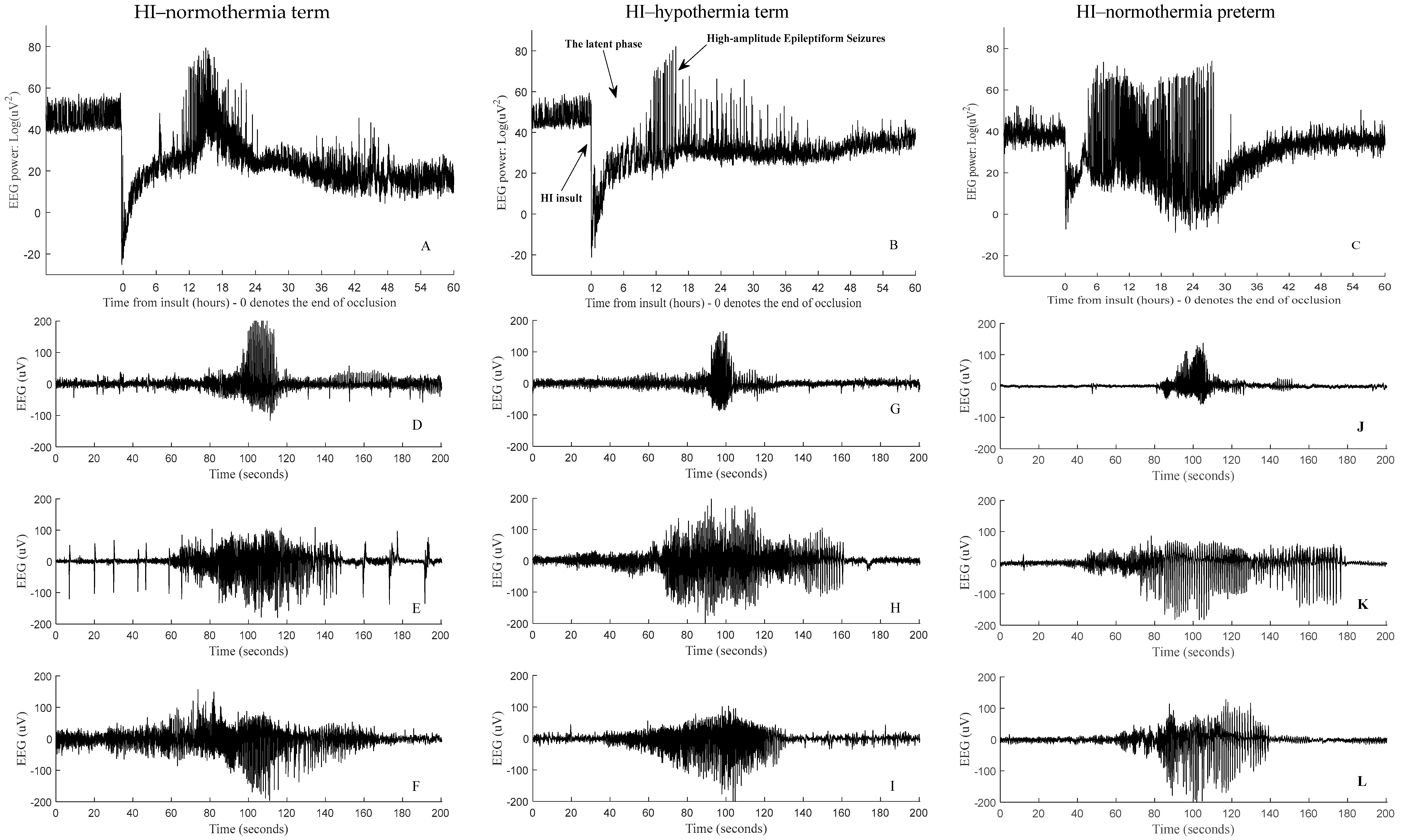
2.6. Experiments, Data Acquisition, and Preparation
Ethics
2.7. Surgical and Experimental Procedures
2.7.1. Fetal Surgery
2.7.2. Experimental Protocols
2.8. Data Acquisition
Preprocessing
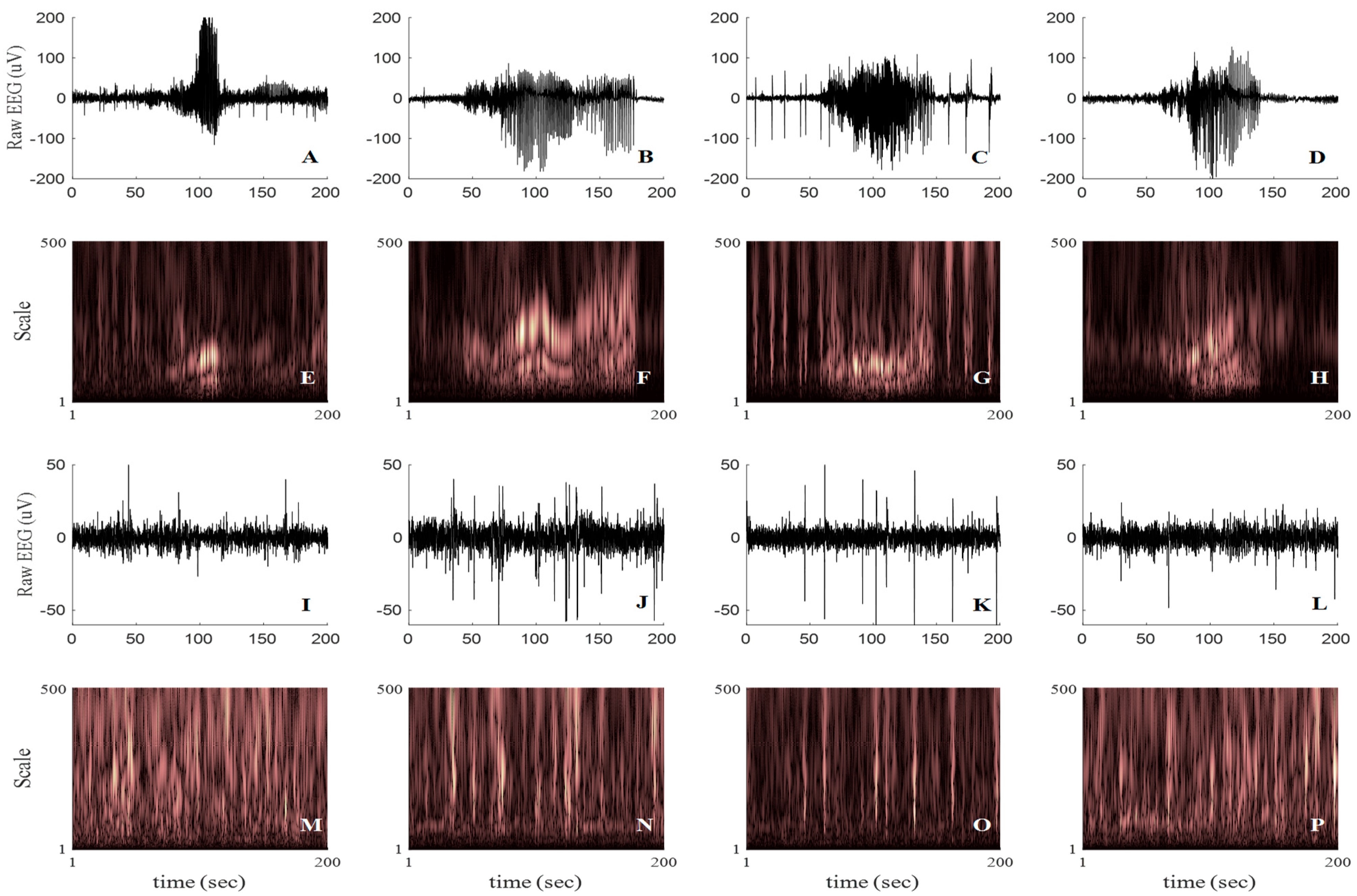
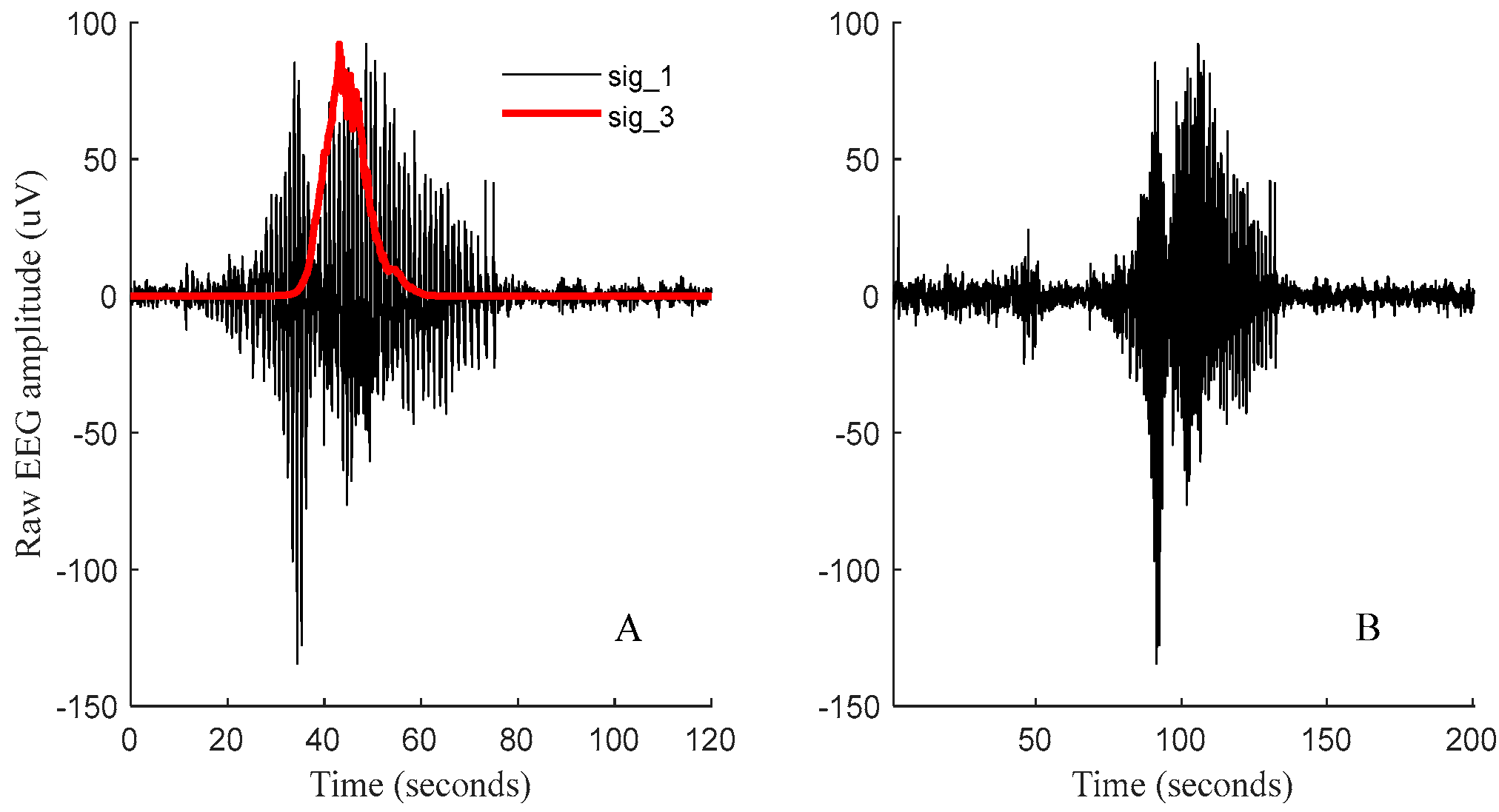
3. Results
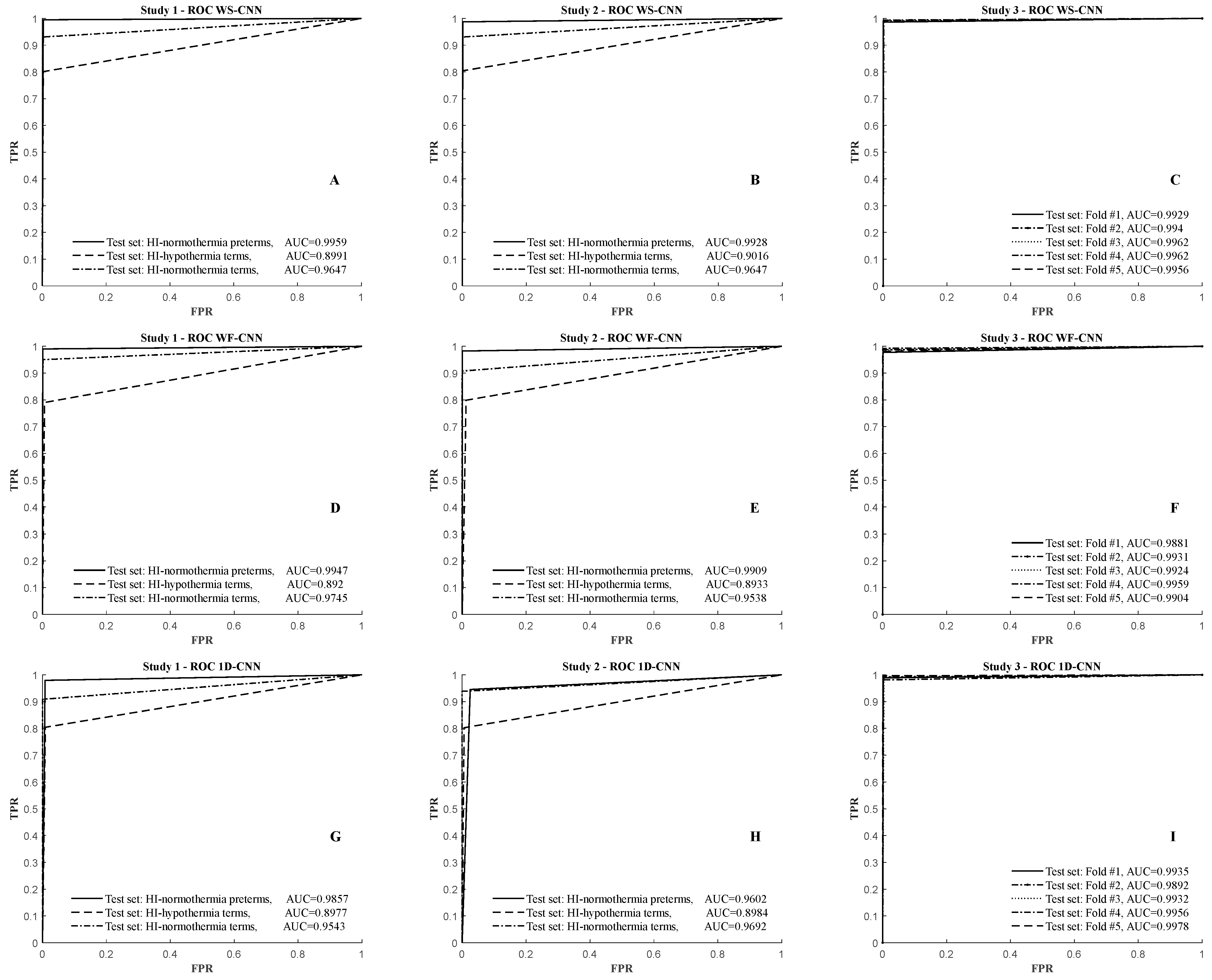
3.1. Results of the WS-CNN Seizure Detector
3.2. Results of the WF-CNN Seizure Detector
3.3. Results of the 1D-CNN Seizure Detector
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Layers | Type | No. of Neurons (Output Layer) | Kernel Size | Stride | Padding | No. of Filters |
|---|---|---|---|---|---|---|
| 0–1 | Conv. | 51,302 × 3 | [3 3] | 1 | 1 | 32 |
| 1–2 | Max_pool | 25,651 × 2 | [2 1] | 2 | 0 | |
| 2–3 | Conv. | 25,651 × 2 | [3 3] | 1 | 1 | 64 |
| 3–4 | Max_pool | 12,825 × 1 | [3 2] | 2 | 0 | |
| 4–5 | Conv. | 12,825 × 1 | [3 3] | 1 | 1 | 96 |
| 5–6 | Max_pool | 6412 × 1 | [3 1] | 2 | 0 | |
| 6–7 | Conv. | 6412 × 1 | [3 3] | 1 | 1 | 128 |
| 7–8 | Max_pool | 3206 × 1 | [2 1] | 2 | 0 | |
| 8–9 | Conv. | 3206 × 1 | [3 3] | 1 | 1 | 256 |
| 9–10 | Max_pool | 1603 × 1 | [2 1] | 2 | 0 | |
| 10–11 | Conv. | 1603 × 1 | [3 3] | 1 | 1 | 512 |
| 11–12 | Max_pool | 801 × 1 | [3 1] | 2 | 0 | |
| 12–14 | Fully_connected | 801 | ||||
| Fully_connected | 20 | |||||
| Fully_connected | 2 | |||||
| Output | Softmax and Classification |
| Layers | Type | No. of Neurons (Output Layer) | Kernel Size | Stride | Padding | No. of Filters |
|---|---|---|---|---|---|---|
| 0–1 | Conv. | 51,302 × 1 | [1024 1] | 1 | 1 | 16 |
| 1–2 | Max_pool | 50,279 × 1 | [3 1] | 2 | 0 | |
| 2–3 | Conv. | 25139 × 1 | [512 1] | 1 | 1 | 32 |
| 3–4 | Max_pool | 24,628 × 1 | [4 1] | 4 | 0 | |
| 4–5 | Conv. | 6157 × 1 | [256 1] | 1 | 1 | 48 |
| 5–6 | Max_pool | 5902 × 1 | [2 1] | 4 | 0 | |
| 6–7 | Conv. | 1476 × 1 | [128 1] | 1 | 1 | 96 |
| 7–8 | Max_pool | 1349 × 1 | [5 1] | 4 | 0 | |
| 8–9 | Conv. | 337 × 1 | [64 1] | 1 | 1 | 128 |
| 9–10 | Max_pool | 274 × 1 | [2 1] | 4 | 0 | |
| 10–11 | Conv. | 69 × 1 | [32 1] | 1 | 1 | 256 |
| 11–12 | Max_pool | 38 × 1 | [2 1] | 4 | 0 | |
| 12–14 | Fully_connected | 2560 | ||||
| Fully_connected | 24 | |||||
| Fully_connected | 2 | |||||
| Output | Softmax and Classification |
References
- Gunn, A.J.; Thoresen, M. Neonatal encephalopathy and hypoxic–ischemic encephalopathy. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 162, pp. 217–237. [Google Scholar]
- Sharpe, C.; Reiner, G.E.; Davis, S.L.; Nespeca, M.; Gold, J.J.; Rasmussen, M.; Kuperman, R.; Harbert, M.J.; Michelson, D.; Joe, P.; et al. Levetiracetam versus phenobarbital for neonatal seizures: A randomized controlled trial. Pediatrics 2020, 145, e20193182. [Google Scholar] [CrossRef] [PubMed]
- DeLaGarza-Pineda, O.; Mailo, J.A.; Boylan, G.; Chau, V.; Glass, H.C.; Mathur, A.M.; Shellhaas, R.A.; Soul, J.S.; Wusthoff, C.J.; Chang, T. Management of seizures in neonates with neonatal encephalopathy treated with hypothermia. Semin. Fetal Neonatal Med. 2021, 26, 101279. [Google Scholar] [CrossRef] [PubMed]
- Bano, S.; Chaudhary, V.; Garga, U.C. Neonatal hypoxic-ischemic encephalopathy: A radiological review. J. Pediatr. Neurosci. 2017, 12, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, C.; Davis, S.L.; Reiner, G.E.; I Lee, L.; Gold, J.J.; Nespeca, M.; Wang, S.G.; Joe, P.; Kuperman, R.; Gardner, M.; et al. Assessing the feasibility of providing a real-time response to seizures detected with continuous long-term neonatal electroencephalography monitoring. J. Clin. Neurophysiol. 2019, 36, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Lynch, N.E.; Stevenson, N.J.; Livingstone, V.; Murphy, B.P.; Rennie, J.M.; Boylan, G.B. The temporal evolution of electrographic seizure burden in neonatal hypoxic ischemic encephalopathy. Epilepsia 2012, 53, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Wietstock, S.O.; Bonifacio, S.L.; Sullivan, J.E.; Nash, K.B.; Glass, H.C. Continuous video electroencephalographic (eeg) monitoring for electrographic seizure diagnosis in neonates: A single-center study. J. Child Neurol. 2016, 31, 328–332. [Google Scholar] [CrossRef]
- Malfilâtre, G.; Mony, L.; Hasaerts, D.; Vignolo-Diard, P.; Lamblin, M.-D.; Bourel-Ponchel, E. Technical recommendations and interpretation guidelines for electroencephalography for premature and full-term newborns. Neurophysiol. Clin. 2021, 51, 35–60. [Google Scholar] [CrossRef]
- Pisani, F.; Pavlidis, E. The role of electroencephalogram in neonatal seizure detection. Expert Rev. Neurother. 2017, 18, 95–100. [Google Scholar] [CrossRef]
- Pavel, A.M.; Rennie, J.M.; de Vries, L.S.; Blennow, M.; Foran, A.; Shah, D.K.; Pressler, R.M.; Kapellou, O.; Dempsey, E.M.; Mathieson, S.R.; et al. Neonatal seizure management: Is the timing of treatment critical? J. Pediatr. 2021, 243, 61–68.e2. [Google Scholar] [CrossRef] [PubMed]
- Bennet, L.; Galinsky, R.; Draghi, V.; Lear, C.A.; Davidson, J.O.; Unsworth, C.P.; Gunn, A.J. Time and sex dependent effects of magnesium sulfate on post-asphyxial seizures in preterm fetal sheep. J. Physiol. 2018, 596, 6079–6092. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Juul, H.M.; Jensen, F.E. Models of hypoxia and ischemia-induced seizures. J. Neurosci. Methods 2016, 260, 252–260. [Google Scholar] [CrossRef]
- McNally, M.A.; Chavez-Valdez, R.; Felling, R.J.; Flock, D.L.; Northington, F.J.; Stafstrom, C.E. Seizure susceptibility correlates with brain injury in male mice treated with hypothermia after neonatal hypoxia-ischemia. Dev. Neurosci. 2019, 40, 576–585. [Google Scholar] [CrossRef]
- Aridas, J.D.; Yawno, T.; Sutherland, A.E.; Nitsos, I.; Wong, F.Y.; Hunt, R.W.; Ditchfield, M.; Fahey, M.C.; Malhotra, A.; Wallace, E.M.; et al. Melatonin augments the neuroprotective effects of hypothermia in lambs following perinatal asphyxia. J. Pineal Res. 2021, 71, e12744. [Google Scholar] [CrossRef]
- Pang, R.; Avdic-Belltheus, A.; Meehan, C.; Martinello, K.; Mutshiya, T.; Yang, Q.; Sokolska, M.; Torrealdea, F.; Hristova, M.; Bainbridge, A.; et al. Melatonin and/or erythropoietin combined with hypothermia in a piglet model of perinatal asphyxia. Brain Commun. 2020, 3, fcaa211. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, H.; Gunn, A.J.; Unsworth, C.P.; Bennet, L. Advanced Deep Learning Spectroscopy of Scalogram Infused CNN Classifiers for Robust Identification of Post-Hypoxic Epileptiform EEG Spikes. Adv. Intell. Syst. 2020, 3, 2000198. [Google Scholar] [CrossRef]
- Abbasi, H.; Drury, P.P.; Lear, C.A.; Gunn, A.J.; Davidson, J.O.; Bennet, L.; Unsworth, C.P. EEG sharp waves are a biomarker of striatal neuronal survival after hypoxia-ischemia in preterm fetal sheep. Sci. Rep. 2018, 8, 16312–16317. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, H.; Gunn, A.J.; Unsworth, C.P.; Bennet, L. Deep Convolutional Neural Networks for the Accurate Identification of High-Amplitude Stereotypic Epileptiform Seizures in the Post-Hypoxic-Ischemic EEG of Preterm Fetal Sheep. In Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Montreal, QC, Canada, 20–24 July 2020; pp. 1–4. [Google Scholar]
- Drury, P.P.; Davidson, J.O.; Heuij, L.G.v.D.; Wassink, G.; Gunn, E.R.; Booth, L.C.; Bennet, L.; Gunn, A.J. Status epilepticus after prolonged umbilical cord occlusion is associated with greater neural injury fetal sheep at term-equivalent. PLoS ONE 2014, 9, e96530. [Google Scholar] [CrossRef] [PubMed]
- Scher, M.S.; Aso, K.; Beggarly, M.E.; Hamid, M.Y.; Steppe, D.A.; Painter, M.J. Electrographic seizures in preterm and full-term neonates: Clinical correlates, associated brain lesions, and risk for neurologic sequelae. Pediatrics 1993, 91, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Clancy, R.R.; Legido, A. The exact ictal and interictal duration of electroencephalographic neonatal seizures. Epilepsia 1987, 28, 537–541. [Google Scholar] [CrossRef]
- Scher, M.S. Seizures in the newborn infant: Diagnosis, treatment, and outcome. Clin. Perinatol. 1997, 24, 735–772. [Google Scholar] [CrossRef]
- Rennie, J.M.; de Vries, L.S.; Blennow, M.; Foran, A.; Shah, D.K.; Livingstone, V.; van Huffelen, A.C.; Mathieson, S.R.; Pavlidis, E.; Weeke, L.C.; et al. Characterisation of neonatal seizures and their treatment using continuous EEG monitoring: A multicentre experience. Arch. Dis. Child. Fetal Neonatal Ed. 2019, 104, F493–F501. [Google Scholar] [CrossRef] [PubMed]
- Kaminiów, K.; Kozak, S.; Paprocka, J. Neonatal Seizures Revisited. Children 2021, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, H.; Dhillon, S.K.; Davidson, J.; Gunn, A.J.; Bennet, L. 2D Wavelet-Scalogram Deep-Learning for Seizures Pattern Identification in the Post-Hypoxic-Ischemic EEG of Preterm Fetal Sheep. In Proceedings of the 45th Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC’23), Sydney, Australia, 24–27 July 2023. [Google Scholar]
- O’shea, A.; Ahmed, R.; Lightbody, G.; Pavlidis, E.; Lloyd, R.; Pisani, F.; Marnane, W.; Mathieson, S.; Boylan, G.; Temko, A. Deep Learning for EEG Seizure Detection in Preterm Infants. Int. J. Neural Syst. 2021, 31, 2150008. [Google Scholar] [CrossRef] [PubMed]
- O’shea, A.; Lightbody, G.; Boylan, G.; Temko, A. Neonatal seizure detection from raw multi-channel EEG using a fully convolutional architecture. Neural Netw. 2020, 123, 12–25. [Google Scholar] [CrossRef]
- Acharya, U.R.; Oh, S.L.; Hagiwara, Y.; Tan, J.H.; Adeli, H. Deep convolutional neural network for the automated detection and diagnosis of seizure using EEG signals. Comput. Biol. Med. 2018, 100, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Gabeff, V.; Teijeiro, T.; Zapater, M.; Cammoun, L.; Rheims, S.; Ryvlin, P.; Atienza, D. Interpreting deep learning models for epileptic seizure detection on EEG signals. Artif. Intell. Med. 2021, 117, 102084. [Google Scholar] [CrossRef]
- Raurale, S.A.; Boylan, G.B.; Mathieson, S.R.; Marnane, W.P.; Lightbody, G.; O’Toole, J.M. Grading hypoxic-ischemic encephalopathy in neonatal EEG with convolutional neural networks and quadratic time-frequency distributions. J. Neural Eng. 2021, 18, 046007. [Google Scholar] [CrossRef]
- Kota, S.; Jasti, K.; Liu, Y.; Liu, H.; Zhang, R.; Chalak, L. EEG spectral power: A proposed physiological biomarker to classify the hypoxic-ischemic encephalopathy severity in real-time. Pediatr. Neurol. 2021, 122, 7–14. [Google Scholar] [CrossRef]
- Moghadam, S.M.; Pinchefsky, E.; Tse, I.; Marchi, V.; Kohonen, J.; Kauppila, M.; Airaksinen, M.; Tapani, K.; Nevalainen, P.; Hahn, C.; et al. Building an open source classifier for the neonatal eeg background: A systematic feature-based approach from expert scoring to clinical visualization. Front. Hum. Neurosci. 2021, 15, 675154. [Google Scholar] [CrossRef]
- Lacan, L.; Betrouni, N.; Lamblin, M.-D.; Chaton, L.; Delval, A.; Bourriez, J.-L.; Storme, L.; Derambure, P.; Tich, S.N. Quantitative approach to early neonatal EEG visual analysis in hypoxic-ischemic encephalopathy severity: Bridging the gap between eyes and machine. Neurophysiol. Clin. 2021, 51, 121–131. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Swetapadma, A.; Pattnaik, P.K. A channel independent generalized seizure detection method for pediatric epileptic seizures. Comput. Methods Programs Biomed. 2021, 209, 106335. [Google Scholar] [CrossRef] [PubMed]
- Gramacki, A.; Gramacki, J. A deep learning framework for epileptic seizure detection based on neonatal EEG signals. Sci. Rep. 2022, 12, 13010. [Google Scholar] [CrossRef] [PubMed]
- Yıldız, İ.; Garner, R.; Lai, M.; Duncan, D. Unsupervised seizure identification on EEG. Comput. Methods Programs Biomed. 2022, 215, 106604. [Google Scholar] [CrossRef]
- Raeisi, K.; Khazaei, M.; Croce, P.; Tamburro, G.; Comani, S.; Zappasodi, F. A graph convolutional neural network for the automated detection of seizures in the neonatal EEG. Comput. Methods Programs Biomed. 2022, 222, 106950. [Google Scholar] [CrossRef] [PubMed]
- Golmohammadi, M.; Shah, V.; Obeid, I.; Picone, J. Deep learning approaches for automated seizure detection from scalp electroencephalograms. In Signal Processing in Medicine and Biology: Emerging Trends in Research and Applications; Springer: Cham, Switzerland, 2020; pp. 235–276. [Google Scholar]
- Pavel, A.M.; O’Toole, J.M.; Proietti, J.; Livingstone, V.; Mitra, S.; Marnane, W.P.; Finder, M.; Dempsey, E.M.; Murray, D.M.; Boylan, G.B.; et al. Machine learning for the early prediction of infants with electrographic seizures in neonatal hypoxic-ischemic encephalopathy. Epilepsia 2023, 64, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Janáčková, S.; Boyd, S.; Yozawitz, E.; Tsuchida, T.; Lamblin, M.D.; Gueden, S.; Pressler, R. Electroencephalographic characteristics of epileptic seizures in preterm neonates. Clin. Neurophysiol. 2016, 127, 2721–2727. [Google Scholar] [CrossRef] [PubMed]
- Patrizi, S.; Holmes, G.L.; Orzalesi, M.; Allemand, F. Neonatal seizures: Characteristics of EEG ictal activity in preterm and full-term infants. Brain Dev. 2003, 25, 427–437. [Google Scholar] [CrossRef]
- Scher, M.S.; Hamid, M.Y.; Steppe, D.A.; Beggarly, M.E.; Painter, M.J. Ictal and interictal electrographic seizure durations in preterm and term neonates. Epilepsia 1993, 34, 284–288. [Google Scholar] [CrossRef]
- Lloyd, R.O.; O’Toole, J.M.; Pavlidis, E.; Filan, P.M.; Boylan, G.B. Electrographic Seizures during the Early Postnatal Period in Preterm Infants. J. Pediatr. 2017, 187, 18–25.e2. [Google Scholar] [CrossRef]
- Wusthoff, C.J.; Dlugos, D.J.; Gutierrez-Colina, A.; Wang, A.; Cook, N.; Donnelly, M.; Clancy, R.; Abend, N.S. Electrographic seizures during therapeutic hypothermia for neonatal hypoxic-ischemic encephalopathy. J. Child Neurol. 2011, 26, 724–728. [Google Scholar] [CrossRef]
- Boylan, G.B.; Kharoshankaya, L.; Wusthoff, C.J. Seizures and hypothermia: Importance of electroencephalographic monitoring and considerations for treatment. Semin. Fetal Neonatal Med. 2015, 20, 103–108. [Google Scholar] [CrossRef]
- Davidson, J.O.; Rout, A.L.; Wassink, G.; Yuill, C.A.; Zhang, F.G.; Green, C.R.; Bennet, L.; Gunn, A.J. Non-Additive Effects of Delayed Connexin Hemichannel Blockade and Hypothermia after Cerebral Ischemia in Near-Term Fetal Sheep. J. Cereb. Blood Flow Metab. 2015, 35, 2052–2061. [Google Scholar] [CrossRef]
- McIntosh, G.H.; Baghurst, K.I.; Potter, B.J.; Hetzel, B.S. Foetal brain development in the sheep. Neuropathol. Appl. Neurobiol. 1979, 5, 103–114. [Google Scholar] [CrossRef]
- Bragin, A.; Engel, J., Jr.; Staba, R.J. High-frequency oscillations in epileptic brain. Curr. Opin. Neurol. 2010, 23, 151–156. [Google Scholar] [CrossRef]
- Hong, S.; Zhou, Y.; Shang, J.; Xiao, C.; Sun, J. Opportunities and challenges of deep learning methods for electrocardiogram data: A systematic review. Comput. Biol. Med. 2020, 122, 103801. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, H.; Unsworth, C.P. Applications of advanced signal processing and machine learning in the neonatal hypoxic-ischemic electroencephalogram. Neural Regen. Res. 2020, 15, 222–231. [Google Scholar]
- Kota, S.; Massaro, A.N.; Chang, T.; Al-Shargabi, T.; Cristante, C.; Vezina, G.; du Plessis, A.; Govindan, R.B. Prognostic Value of Continuous Electroencephalogram Delta Power in Neonates with Hypoxic-Ischemic Encephalopathy. J. Child Neurol. 2020, 35, 517–525. [Google Scholar] [CrossRef] [PubMed]
- MathWorks. Matlab. Available online: https://www.mathworks.com/help/matlab/ref/rng.html (accessed on 28 January 2024).
- New Zealand eScience Infrastructure (NeSI). Available online: https://www.nesi.org.nz/ (accessed on 28 January 2024).
- NVIDIA Corporation. NVIDIA Tesla A100 GPUs. 2020. Available online: https://www.nvidia.com/content/dam/en-zz/Solutions/Data-Center/a100/pdf/nvidia-a100-datasheet-us-nvidia-1758950-r4-web.pdf (accessed on 28 January 2024).
- Patterson, D.; Sweasey, D.; Hebert, C.N. Changes occurring in the chemical composition of the central nervous system during foetal and post-natal development of the sheep. J. Neurochem. 1971, 18, 2027–2040. [Google Scholar] [CrossRef]
- van den Heuij, L.G.; Fraser, M.; Miller, S.L.; Jenkin, G.; Wallace, E.M.; Davidson, J.O.; Lear, C.A.; Lim, R.; Wassink, G.; Gunn, A.J.; et al. Delayed intranasal infusion of human amnion epithelial cells improves white matter maturation after asphyxia in preterm fetal sheep. J. Cereb. Blood Flow Metab. 2019, 39, 223–239. [Google Scholar] [CrossRef]
- Dubois, J.; Dehaene-Lambertz, G.; Kulikova, S.; Poupon, C.; Hüppi, P.S.; Hertz-Pannier, L. The early development of brain white matter: A review of imaging studies in fetuses, newborns, and infants. Neuroscience 2014, 276, 48–71. [Google Scholar] [CrossRef] [PubMed]
- Sannia, A.; Natalizia, A.R.; Parodi, A.; Malova, M.; Fumagalli, M.; Rossi, A.; Ramenghi, L.A. Different gestational ages and changing vulnerability of the premature brain. J. Matern. Neonatal Med. 2015, 28 (Suppl. S1), 2268–2272. [Google Scholar] [CrossRef]
- André, M.; Lamblin, M.D.; d’Allest, A.M.; Curzi-Dascalova, L.; Moussalli-Salefranque, F.; Tich, S.N.T.; Vecchierini-Blineau, M.F.; Wallois, F.; Walls-Esquivel, E.; Plouin, P. Electroencephalography in premature and full-term infants: Developmental features and glossary. Neurophysiol. Clin. 2010, 40, 59–124. [Google Scholar] [CrossRef] [PubMed]
- Bourel-Ponchel, E.; Gueden, S.; Hasaerts, D.; Héberlé, C.; Malfilâtre, G.; Mony, L.; Vignolo-Diard, P.; Lamblin, M.D. Normal EEG during the neonatal period: Maturational aspects from premature to full-term newborns. Neurophysiol. Clin. 2021, 51, 61–88. [Google Scholar] [CrossRef] [PubMed]
- Wallois, F.; Routier, L.; Heberlé, C.; Mahmoudzadeh, M.; Bourel-Ponchel, E.; Moghimi, S. Back to basics: The neuronal substrates and mechanisms that underlie the electroencephalogram in premature neonates. Neurophysiol. Clin. 2021, 51, 5–33. [Google Scholar] [CrossRef]
- Jennekens, W.; Niemarkt, H.J.; Engels, M.; Pasman, J.W.; van Pul, C.; Andriessen, P. Topography of maturational changes in EEG burst spectral power of the preterm infant with a normal follow-up at 2years of age. Clin. Neurophysiol. 2012, 123, 2130–2138. [Google Scholar] [CrossRef]
- Pillay, K.; Dereymaeker, A.; Jansen, K.; Naulaers, G.; De Vos, M. Applying a data-driven approach to quantify EEG maturational deviations in preterms with normal and abnormal neurodevelopmental outcomes. Sci. Rep. 2020, 10, 7288. [Google Scholar] [CrossRef]
- Dereymaeker, A.; Pillay, K.; Vervisch, J.; De Vos, M.; Van Huffel, S.; Jansen, K.; Naulaers, G. Review of sleep-EEG in preterm and term neonates. Early Hum. Dev. 2017, 113, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Scher, M.S.; Sun, M.; Steppe, D.A.; Guthrie, R.D.; Sclabassi, R.J. Comparisons of EEG spectral and correlation measures between healthy term and preterm infants. Pediatr. Neurol. 1994, 10, 104–108. [Google Scholar] [CrossRef]
- Rols, G.; Tallon-Baudry, C.; Girard, P.; Bertrand, O.; Bullier, J. Cortical mapping of gamma oscillations in areas V1 and V4 of the macaque monkey. Vis. Neurosci. 2001, 18, 527–540. [Google Scholar] [CrossRef]
- Ramantani, G.; Maillard, L.; Koessler, L. Correlation of invasive EEG and scalp EEG. Seizure 2016, 41, 196–200. [Google Scholar] [CrossRef]
- Müller-Putz, G.R. Electroencephalography. Handb. Clin. Neurol. 2020, 168, 249–262. [Google Scholar] [PubMed]





| Layers | Type | No. of Neurons (Output Layer) | Kernel Size | Stride | Padding | No. of Filters |
|---|---|---|---|---|---|---|
| 0–1 | Conv. | 333 × 500 | 3 | 1 | 1 | 32 |
| 1–2 | Max_pool | 166 × 250 | [3 2] | 2 | 0 | |
| 2–3 | Conv. | 166 × 250 | 3 | 1 | 1 | 48 |
| 3–4 | Max_pool | 83 × 125 | 2 | 2 | 0 | |
| 4–5 | Conv. | 83 × 125 | 3 | 1 | 1 | 64 |
| 5–6 | Max_pool | 41 × 62 | 3 | 2 | 0 | |
| 6–7 | Conv. | 41 × 62 | 3 | 1 | 1 | 96 |
| 7–8 | Max_pool | 20 × 31 | [3 2] | 2 | 0 | |
| 8–9 | Conv. | 20 × 31 | 3 | 1 | 1 | 128 |
| 9–10 | Max_pool | 10 × 15 | [2 3] | 2 | 0 | |
| 10–11 | Conv. | 10 × 15 | 3 | 1 | 1 | 192 |
| 11–12 | Max_pool | 4 × 6 | [4 5] | 2 | 0 | |
| 12–13 | Conv. | 4 × 6 | 3 | 1 | 1 | 256 |
| 13–14 | Max_pool | 2 × 3 | 2 | 2 | 0 | |
| 14–17 | Fully_connected | 1536 | ||||
| Fully_connected | 24 | |||||
| Fully_connected | 2 | |||||
| Output | Softmax and Classification |
| Trained/Validated on | No. of EEG Patterns in the Training Set (Total/Seizures/Non_Seizures) | Total Length of Training Set (in Hours/Days) | Test on | No. of EEG Patterns in the Testing Set (Total/Seizures/Non_Seizures) | Total Length of Testing Set (in Hours/Days) | Training-to-Testing Ratio | |
|---|---|---|---|---|---|---|---|
| Study #1 3 schemes | G1 + G2 + G3 | 20,491/2311/18,180 | 1024.6/42.7 | G4 | 10,524/1644/8880 | 526.2/21.9 | 1.95 |
| G1 + G4 + G3 | 20,852/2652/18,200 | 1042.6/43.4 | G2 | 10,163/1303/8860 | 508.2/21.2 | 2.05 | |
| G2 + G4 + G3 | 25,582/2947/22,635 | 1279.1/53.3 | G1 | 5433 /1008/4425 | 271.7/11.3 | 4.71 | |
| Study #2 3 schemes | G1 + G2 | 15,596/2311/13,285 | 779.8/32.5 | G4 | 10,524/1644/8880 | 526.2/21.9 | 1.48 |
| G1 + G4 | 15,957/2652/13305 | 797.9/33.2 | G2 | 10,163/1303/8860 | 508.2/21.2 | 1.57 | |
| G2 + G4 | 20,687/2947/17,740 | 1034.4/43.1 | G1 | 5433/1008/4425 | 271.7/11.3 | 3.81 | |
| Study #3 5 folds | G1 + G2 + G3 + G4 (Fold #1) | 24,812/3164/21,648 | 1040.6/43.4 | Fold #1 test-set | 6203/791/5412 | 310.2/12.9 | 4.00 |
| G1 + G2 + G3 + G4 (Fold #2) | 24,812/3164/21,648 | 1040.6/43.4 | Fold #2 test-set | 6203/791/5412 | 310.2/12.9 | 4.00 | |
| G1 + G2 + G3 + G4 (Fold #3) | 24,812/3164/21,648 | 1040.6/43.4 | Fold #3 test-set | 6203/791/5412 | 310.2/12.9 | 4.00 | |
| G1 + G2 + G3 + G4 (Fold #4) | 24,812/3164/21,648 | 1040.6/43.4 | Fold #4 test-set | 6203/791/5412 | 310.2/12.9 | 4.00 | |
| G1 + G2 + G3 + G4 (Fold #5) | 24,812/3164/21,648 | 1040.6/43.4 | Fold #5 test-set | 6203/791/5412 | 310.2/12.9 | 4.00 |
| No. of Animals in the Cohort | No. of Seizures in the Left EEG Channel | No. of Seizures in the Right EEG Channel | No. of Non-Seizures in the Left EEG Channel | No. of Non-Seizures in the Right EEG Channel | |
|---|---|---|---|---|---|
| HI–normothermia terms | 7 | 470 | 538 | 2213 | 2212 |
| HI–hypothermia terms | 14 | 594 | 709 | 4423 | 4437 |
| Sham–normothermia terms | 5 | 0 | 0 | 2438 | 2457 |
| HI–normothermia preterms | 14 | 844 | 800 | 4443 | 4437 |
| Sum | 40 | 1908 | 2047 | 13,517 | 13,543 |
| Total | 3955 | 27,060 | |||
| TP Hits | FP Hits | FN Hits | TN Hits | Sensitivity [%] | Selectivity [%] | Precision [%] | Accuracy [%] | AUC | Average Accuracy [%] | Average AUC | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Study #1 3 schemes | 1636 | 8 | 29 | 8851 | 98.26 | 99.91 | 99.51 | 99.65 | 0.9959 | 98.48 ±1.01 | 0.9532 ±0.0403 |
| 1044 | 259 | 27 | 8833 | 97.48 | 97.15 | 80.12 | 97.19 | 0.8991 | |||
| 938 | 70 | 5 | 4420 | 99.47 | 98.44 | 93.06 | 98.62 | 0.9647 | |||
| Study #2 3 schemes | 1623 | 21 | 15 | 8865 | 99.08 | 99.76 | 98.72 | 99.66 | 0.9928 | 98.56 ±0.92 | 0.9530 ±0.0382 |
| 1048 | 255 | 9 | 8851 | 99.15 | 97.20 | 80.43 | 97.40 | 0.9015 | |||
| 938 | 70 | 5 | 4420 | 99.47 | 98.44 | 93.06 | 98.62 | 0.9647 | |||
| Study #3 5 folds | 780 | 11 | 2 | 5410 | 99.74 | 99.80 | 98.61 | 99.79 | 0.9929 | 99.78 ±0.04 | 0.9950 ±0.0013 |
| 783 | 8 | 10 | 5402 | 98.74 | 99.85 | 98.99 | 99.71 | 0.9940 | |||
| 786 | 5 | 7 | 5405 | 99.12 | 99.91 | 99.37 | 99.81 | 0.9962 | |||
| 786 | 5 | 7 | 5405 | 99.12 | 99.91 | 99.37 | 99.81 | 0.9962 | |||
| 785 | 6 | 7 | 5405 | 99.12 | 99.89 | 99.24 | 99.79 | 0.9956 |
| TP Hits | FP Hits | FN Hits | TN Hits | Sensitivity [%] | Selectivity [%] | Precision [%] | Accuracy [%] | AUC | Average Accuracy [%] | Average AUC | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Study #1 3 schemes | 1628 | 16 | 7 | 8873 | 99.57 | 99.82 | 99.03 | 99.78 | 0.9947 | 98.50 ±1.28 | 0.9507 ±0.0444 |
| 1030 | 273 | 57 | 8803 | 94.76 | 96.99 | 79.05 | 96.75 | 0.892 | |||
| 958 | 50 | 6 | 4419 | 99.38 | 98.88 | 95.04 | 98.97 | 0.9745 | |||
| Study #2 3 schemes | 1615 | 29 | 5 | 8875 | 99.69 | 99.67 | 98.24 | 99.68 | 0.9909 | 98.10 ±1.36 | 0.9460 ±0.0402 |
| 1041 | 262 | 109 | 8751 | 90.52 | 97.09 | 79.89 | 96.35 | 0.8933 | |||
| 915 | 93 | 1 | 4424 | 99.89 | 97.94 | 90.77 | 98.27 | 0.9538 | |||
| Study #3 5 folds | 773 | 18 | 6 | 5406 | 99.23 | 99.67 | 97.72 | 99.61 | 0.9881 | 99.73 ±0.08 | 0.9920 ±0.0026 |
| 781 | 10 | 6 | 5406 | 99.24 | 99.82 | 98.74 | 99.74 | 0.9931 | |||
| 780 | 11 | 7 | 5405 | 99.11 | 99.80 | 98.61 | 99.71 | 0.9924 | |||
| 785 | 6 | 3 | 5409 | 99.62 | 99.89 | 99.24 | 99.85 | 0.9959 | |||
| 776 | 15 | 1 | 5411 | 99.87 | 99.72 | 98.10 | 99.74 | 0.9904 |
| TP hits | FP hits | FN hits | TN hits | Sensitivity [%] | Selectivity [%] | Precision [%] | Accuracy [%] | AUC | Average Accuracy [%] | Average AUC | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Study #1 3 schemes | 1610 | 34 | 70 | 8810 | 95.83 | 99.62 | 97.93 | 99.01 | 0.9857 | 98.00 ±0.96 | 0.9459 ±0.0364 |
| 1048 | 255 | 79 | 8781 | 92.99 | 97.18 | 80.43 | 96.71 | 0.8977 | |||
| 916 | 92 | 1 | 4424 | 99.89 | 97.96 | 90.87 | 98.29 | 0.9543 | |||
| Study #2 3 schemes | 1554 | 90 | 221 | 8659 | 87.55 | 98.97 | 94.53 | 97.04 | 0.9602 | 97.62 ±0.88 | 0.9426 ±0.0315 |
| 1046 | 257 | 53 | 8807 | 95.18 | 97.16 | 80.28 | 96.95 | 0.8984 | |||
| 946 | 62 | 0 | 4425 | 100.00 | 98.62 | 93.85 | 98.86 | 0.9692 | |||
| Study #3 5 folds | 782 | 9 | 9 | 5403 | 98.86 | 99.83 | 98.86 | 99.71 | 0.9935 | 99.70 ±0.14 | 0.9939 ±0.0029 |
| 776 | 15 | 14 | 5398 | 98.23 | 99.72 | 98.10 | 99.53 | 0.9892 | |||
| 783 | 8 | 19 | 5393 | 97.63 | 99.85 | 98.99 | 99.56 | 0.9932 | |||
| 785 | 6 | 7 | 5405 | 99.12 | 99.89 | 99.24 | 99.79 | 0.9956 | |||
| 788 | 3 | 3 | 5409 | 99.62 | 99.94 | 99.62 | 99.90 | 0.9978 |
| Study #1 | Study #2 | Study #3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Scheme 1 | Scheme 2 | Scheme 3 | Scheme 1 | Scheme 2 | Scheme 3 | Fold_1 | Fold_2 | Fold_3 | Fold_4 | Fold_5 | |
| Average overall performance from all classifiers (%) | 99.48 ±0.34 | 96.88 ±0.21 | 98.63 ±0.28 | 98.79 ±1.24 | 96.90 ±0.43 | 98.58 ±0.24 | 99.70 ±0.07 | 99.66 ±0.09 | 99.69 ±0.10 | 99.82 ±0.03 | 99.81 ±0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbasi, H.; Davidson, J.O.; Dhillon, S.K.; Zhou, K.Q.; Wassink, G.; Gunn, A.J.; Bennet, L. Deep Learning for Generalized EEG Seizure Detection after Hypoxia–Ischemia—Preclinical Validation. Bioengineering 2024, 11, 217. https://doi.org/10.3390/bioengineering11030217
Abbasi H, Davidson JO, Dhillon SK, Zhou KQ, Wassink G, Gunn AJ, Bennet L. Deep Learning for Generalized EEG Seizure Detection after Hypoxia–Ischemia—Preclinical Validation. Bioengineering. 2024; 11(3):217. https://doi.org/10.3390/bioengineering11030217
Chicago/Turabian StyleAbbasi, Hamid, Joanne O. Davidson, Simerdeep K. Dhillon, Kelly Q. Zhou, Guido Wassink, Alistair J. Gunn, and Laura Bennet. 2024. "Deep Learning for Generalized EEG Seizure Detection after Hypoxia–Ischemia—Preclinical Validation" Bioengineering 11, no. 3: 217. https://doi.org/10.3390/bioengineering11030217