
The Effects of Habitual Foot Strike Patterns on the Morphology and Mechanical Function of the Medial Gastrocnemius–Achilles Tendon Unit


Abstract
:1. Introduction
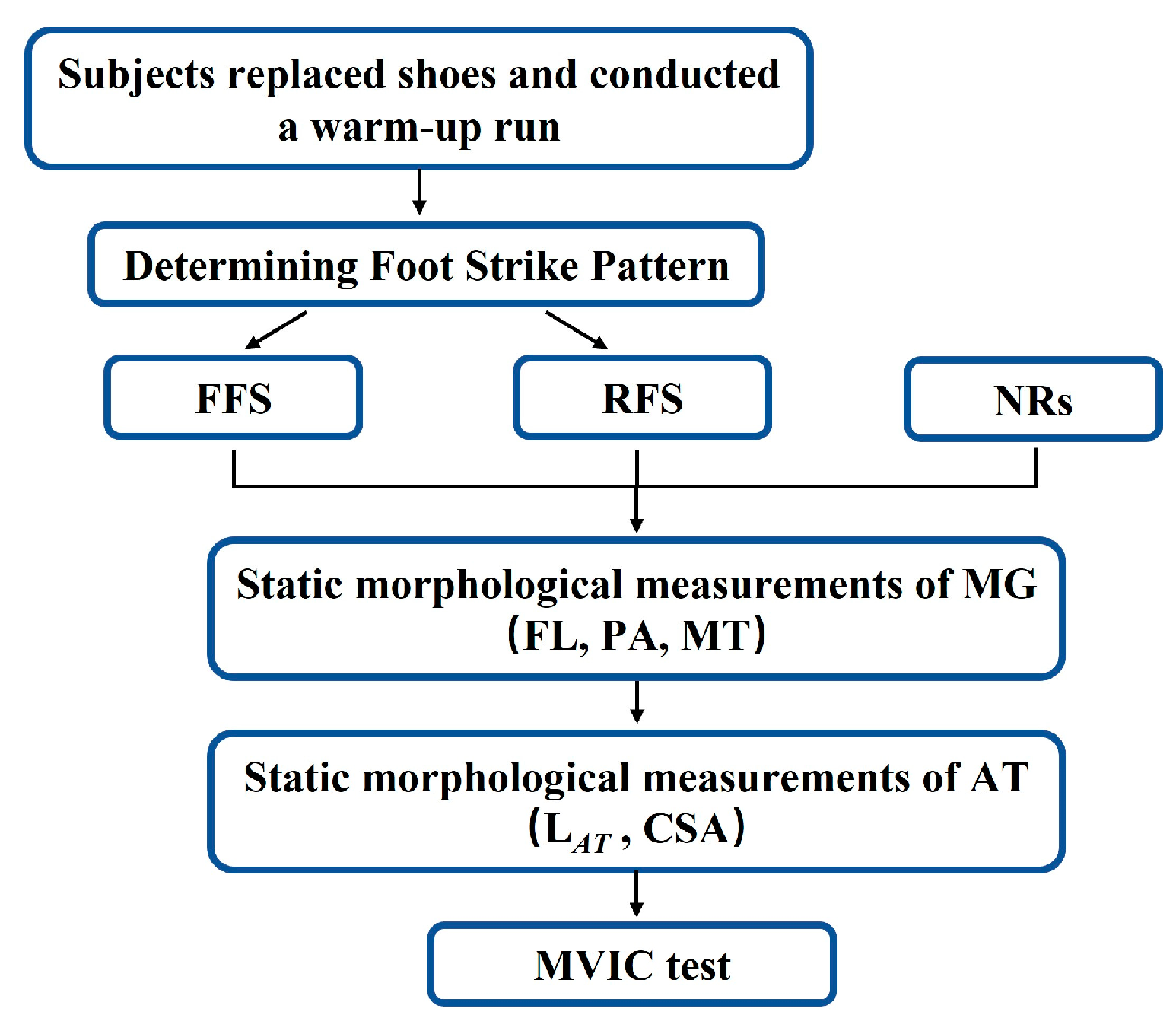
2. Materials and Methods
2.1. Participants
2.2. Instrumentations
2.3. Procedures
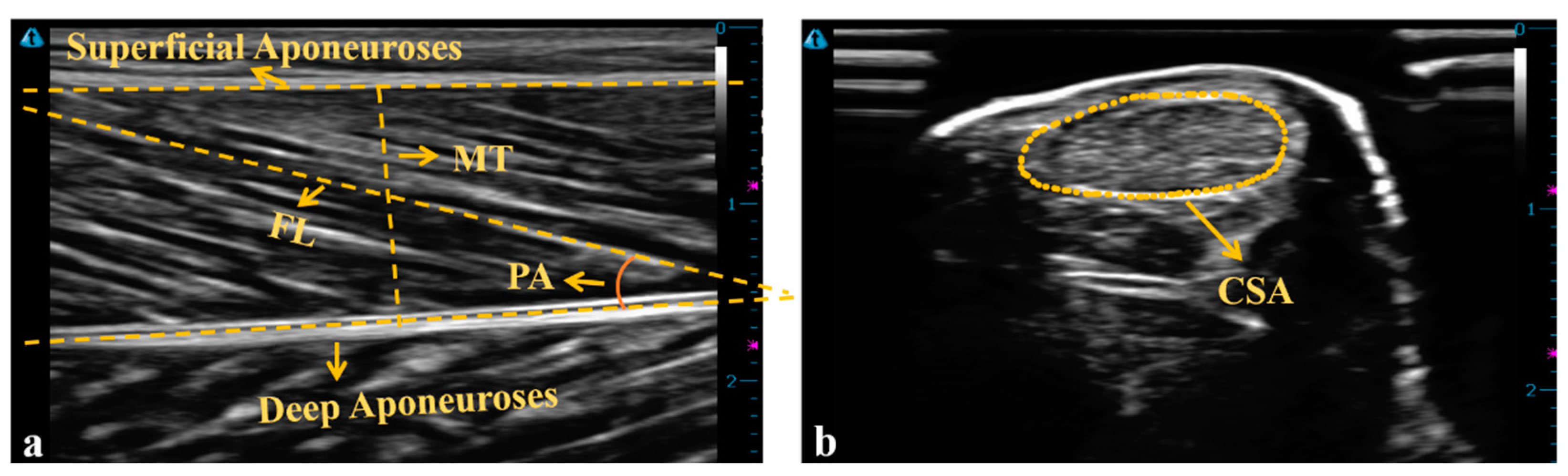
2.4. Data Analysis

2.5. Statistical Analysis
3. Results
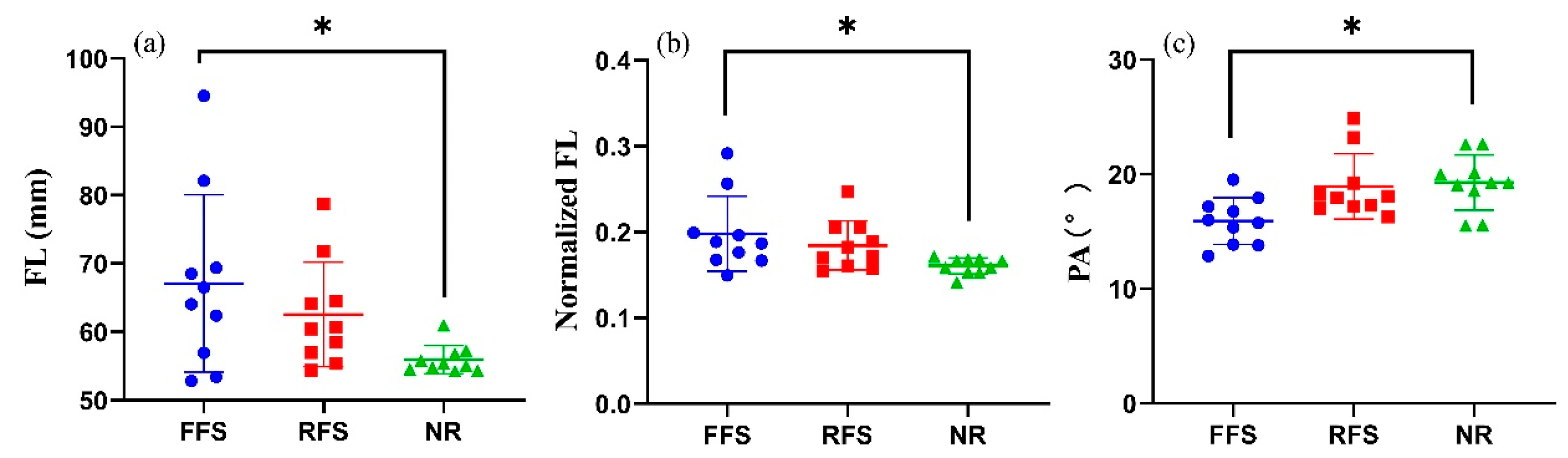
3.1. MG Morphology
3.2. AT Morphology
3.3. Ankle Torque during MVICs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | FFS (n = 10) | RFS (n = 10) | NRs (n = 10) | p Value | η2 |
|---|---|---|---|---|---|
| shank length (cm) | 33.7 ± 2.4 | 34.1 ± 2.1 | 34.9 ± 2.0 | 0.641 | 0.849 |
| FL (mm) | 67.3 ± 12.7 | 62.5 ± 7.6 | 55.9 ± 2.0 * | 0.029 | 0.187 |
| normalized FL | 0.36 ± 0.48 | 0.18 ± 0.03 | 0.16 ± 0.01 * | 0.015 | 0.237 |
| muscle thickness (mm) | 16.8 ± 1.5 | 17.3 ± 2.4 | 18.9 ± 1.9 | 0.065 | 0.183 |
| pennation angle (°) | 16.2 ± 1.9 | 18.9 ± 2.8 | 19.3 ± 2.4 * | 0.008 | 0.280 |
| LAT (cm) | 20.43 ± 2.27 | 20.23 ± 1.73 | 21.57 ± 2.41 | 0.413 | 0.063 |
| normalized LAT | 0.61 ± 0.05 | 0.60 ± 0.07 | 0.62 ± 0.07 | 0.726 | 0.023 |
| CSA (mm2) | 55.82 ± 11.52 | 56.13 ± 8.29 | 51.95 ± 6.95 | 0.387 | 0.068 |
| TMAX (N∙m) | 106.55 ± 31.07 | 93.88 ± 22.50 | 96.62 ± 27.35 | 0.418 | 0.063 |
| normalized TMAX (Nm∙kg−1) | 1.56 ± 0.039 | 1.35 ± 0.30 | 1.41 ± 0.34 | 0.345 | 0.076 |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scheerder, J.; Breedveld, K.; Borgers, J. Running across Europe: The Rise and Size of One of the Largest Sport Markets; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Andersen, J.J. The State of Running 2019. Available online: https://runrepeat.com/state-of-running (accessed on 12 November 2022).
- Hamner, S.R.; Seth, A.; Delp, S.L. Muscle contributions to propulsion and support during running. J. Biomech. 2010, 43, 2709–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, H.; Suga, T.; Takao, K.; Tanaka, T.; Misaki, J.; Miyake, Y.; Nagano, A.; Isaka, T. Relationship between Achilles tendon length and running performance in well-trained male endurance runners. Scand. J. Med. Sci. Sports 2018, 28, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Timmins, R.G.; Bourne, M.N.; Shield, A.J.; Williams, M.D.; Lorenzen, C.; Opar, D.A. Short biceps femoris fascicles and eccentric knee flexor weakness increase the risk of hamstring injury in elite football (soccer): A prospective cohort study. Br. J. Sports Med. 2016, 50, 1524–1535. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, K.; Abe, T.; Brechue, W.F.; Ryushi, T.; Takano, S.; Mizuno, M. Sprint performance is related to muscle fascicle length in male 100-m sprinters. J. Appl. Physiol. 2000, 88, 811–816. [Google Scholar] [CrossRef]
- Arampatzis, A.; De Monte, G.; Karamanidis, K.; Morey-Klapsing, G.; Stafilidis, S.; Bruggemann, G.P. Influence of the muscle-tendon unit’s mechanical and morphological properties on running economy. J. Exp. Biol. 2006, 209, 3345–3357. [Google Scholar] [CrossRef] [Green Version]
- Timmins, R.G.; Shield, A.J.; Williams, M.D.; Lorenzen, C.; Opar, D.A. Architectural adaptations of muscle to training and injury: A narrative review outlining the contributions by fascicle length, pennation angle and muscle thickness. Br. J. Sports Med. 2016, 50, 1467–1472. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, B.; Kobor, I.; Gyimes, Z.; Sebestyen, O.; Tihanyi, J. Lower leg muscle-tendon unit characteristics are related to marathon running performance. Sci. Rep. 2020, 10, 17870. [Google Scholar] [CrossRef]
- Geremia, J.M.; Baroni, B.M.; Lanferdini, F.J.; Bini, R.R.; Sonda, F.C.; Vaz, M.A. Time course of neuromechanical and morphological adaptations to triceps surae isokinetic eccentric training. Phys. Ther. Sports 2018, 34, 84–91. [Google Scholar] [CrossRef]
- Liebl, D.; Willwacher, S.; Hamill, J.; Bruggemann, G.P. Ankle plantarflexion strength in rearfoot and forefoot runners: A novel clusteranalytic approach. Hum. Mov. Sci. 2014, 35, 104–120. [Google Scholar] [CrossRef]
- Almeida, M.O.; Davis, I.S.; Lopes, A.D. Biomechanical Differences of Foot-Strike Patterns During Running: A Systematic Review With Meta-analysis. J. Orthop. Sports Phys. Ther. 2015, 45, 738–755. [Google Scholar] [CrossRef] [Green Version]
- Cibulka, M.; Wenthe, A.; Boyle, Z.; Callier, D.; Schwerdt, A.; Jarman, D.; Strube, M.J. Variation in Medial and Lateral Gastrocnemius Muscle Activity with Foot Position. Int. J. Sports Phys. Ther. 2017, 12, 233–241. [Google Scholar] [PubMed]
- Fukunaga, T.; Roy, R.R.; Shellock, F.G.; Hodgson, J.A.; Day, M.K.; Lee, P.L.; Kwong-Fu, H.; Edgerton, V.R. Physiological cross-sectional area of human leg muscles based on magnetic resonance imaging. J. Orthop. Res. 1992, 10, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Neptune, R.R. Differences in muscle function during walking and running at the same speed. J. Biomech. 2006, 39, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Kumagai, K.; Brechue, W.F. Fascicle length of leg muscles is greater in sprinters than distance runners. Med. Sci. Sports Exerc. 2000, 32, 1125–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchholtz, K.A.; Lambert, M.I.; Bosch, A.; Burgess, T.L. Calf muscle architecture and function in ultra runners and low physical activity individuals: A comparative review. Transl. Sports Med. 2018, 1, 250–256. [Google Scholar] [CrossRef]
- Rosager, S.; Aagaard, P.; Dyhre-Poulsen, P.; Neergaard, K.; Kjaer, M.; Magnusson, S.P. Load-displacement properties of the human triceps surae aponeurosis and tendon in runners and non-runners. Scand. J. Med. Sci. Sports 2002, 12, 90–98. [Google Scholar] [CrossRef]
- Westh, E.; Kongsgaard, M.; Bojsen-Moller, J.; Aagaard, P.; Hansen, M.; Kjaer, M.; Magnusson, S. Effect of habitual exercise on the structural and mechanical properties of human tendon, in vivo, in men and women. Scand. J. Med. Sci. Sports 2008, 18, 23–30. [Google Scholar] [CrossRef]
- Intziegianni, K.; Cassel, M.; Rauf, S.; White, S.; Rector, M.; Kaplick, H.; Wahmkow, G.; Kratzenstein, S.; Mayer, F. Influence of Age and Pathology on Achilles Tendon Properties During a Single-leg Jump. Int. J. Sports Med. 2016, 37, 973–978. [Google Scholar] [CrossRef]
- Greever, C.J. Aerobic and Skeletal Muscle Architectural Adaptations to Concurrent Marathon and Circuit Resistance Training. Master’s Thesis, James Madison University, Harrisonburg, VA, USA, 2012. [Google Scholar]
- Holler, P.; Jaunig, J.; Amort, F.-M.; Tuttner, S.; Hofer-Fischanger, K.; Wallner, D.; Simi, H.; Müller, A.; van Poppel, M.N.M.; Moser, O. Holistic physical exercise training improves physical literacy among physically inactive adults: A pilot intervention study. BMC Public Health 2019, 19, 393. [Google Scholar] [CrossRef]
- Zhang, X.; Deng, L.; Yang, Y.; Xiao, S.; Li, L.; Fu, W. Effects of 12-week transition training with minimalist shoes on Achilles tendon loading in habitual rearfoot strike runners. J. Biomech. 2021, 128, 110807. [Google Scholar] [CrossRef]
- Wang, J.; Luo, Z.; Dai, B.; Fu, W. Effects of 12-week cadence retraining on impact peak, load rates and lower extremity biomechanics in running. PeerJ 2020, 8, e9813. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.; Patritti, B.L.; Stana, L.; Tweedy, S.M. Is increased residual shank length a competitive advantage for elite transtibial amputee long jumpers? Adapt. Phys. Activ. Q. 2011, 28, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geremia, J.M.; Baroni, B.M.; Bini, R.R.; Lanferdini, F.J.; de Lima, A.R.; Herzog, W.; Vaz, M.A. Triceps Surae Muscle Architecture Adaptations to Eccentric Training. Front. Physiol. 2019, 10, 1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Deng, L.; Xiao, S.; Li, L.; Fu, W. Sex Differences in the Morphological and Mechanical Properties of the Achilles Tendon. Int. J. Environ. Res. Public Health 2021, 18, 8974. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Zhang, X.; Xiao, S.; Wang, B.; Fu, W. Gender Difference in Architectural and Mechanical Properties of Medial Gastrocnemius-Achilles Tendon Unit In Vivo. Life 2021, 11, 569. [Google Scholar] [CrossRef] [PubMed]
- Finni, T.; Ikegawa, S.; Komi, P.V. Concentric force enhancement during human movement. Acta Physiol. Scand. 2001, 173, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Histen, K.; Arntsen, J.; L’Hereux, L.; Heeren, J.; Wicki, B.; Saint, S.; Aerni, G.; Denegar, C.R.; Joseph, M.F. Achilles Tendon Properties of Minimalist and Traditionally Shod Runners. J. Sports Rehabil. 2017, 26, 159–164. [Google Scholar] [CrossRef]
- Kudo, S.; Sato, T.; Miyashita, T. Effect of plyometric training on the fascicle length of the gastrocnemius medialis muscle. J. Phys. Ther. Sci. 2020, 32, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Wickiewicz, T.L.; Roy, R.R.; Powell, P.L.; Perrine, J.J.; Edgerton, V.R. Muscle architecture and force-velocity relationships in humans. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1984, 57, 435–443. [Google Scholar] [CrossRef]
- Burkholder, T.J.; Fingado, B.; Baron, S.; Lieber, R.L. Relationship between muscle fiber types and sizes and muscle architectural properties in the mouse hindlimb. J. Morphol. 1994, 221, 177–190. [Google Scholar] [CrossRef]
- Brockett, C.L.; Morgan, D.L.; Proske, U. Human hamstring muscles adapt to eccentric exercise by changing optimum length. Med. Sci. Sports Exerc. 2001, 33, 783–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieber, R.L.; Ward, S.R. Skeletal muscle design to meet functional demands. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 1466–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duclay, J.; Martin, A.; Duclay, A.; Cometti, G.; Pousson, M. Behavior of fascicles and the myotendinous junction of human medial gastrocnemius following eccentric strength training. Muscle Nerve 2009, 39, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Machado, E.; Lanferdini, F.J.; da Silva, E.S.; Geremia, J.M.; Sonda, F.C.; Fletcher, J.R.; Vaz, M.A.; Peyre-Tartaruga, L.A. Triceps Surae Muscle-Tendon Properties as Determinants of the Metabolic Cost in Trained Long-Distance Runners. Front. Physiol. 2021, 12, 767445. [Google Scholar] [CrossRef] [PubMed]
- Young, D.; Degiacomo, C.; Fisher, S.; von Borstel, D. Overcoming Your Achilles Heel: A Review of Achilles Tendon Anatomy, Pathology and Associated MRI Findings. J. Am. Osteopath. Coll. Radiol. 2020, 9. [Google Scholar]
- Hansen, P.; Aagaard, P.; Kjaer, M.; Larsson, B.; Magnusson, S.P. Effect of habitual running on human Achilles tendon load-deformation properties and cross-sectional area. J. Appl. Physiol. 2003, 95, 2375–2380. [Google Scholar] [CrossRef] [Green Version]
- Petrovic, I.; Marinković, M. Influence of morphological characteristics on running performance of endurance athletes. Facta Univ. Ser. Phys. Educ. Sport 2018, 16, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Black, M.I.; Allen, S.J.; Forrester, S.E.; Folland, J.P. The Anthropometry of Economical Running. Med. Sci. Sports Exerc. 2020, 52, 762–770. [Google Scholar] [CrossRef]
- Muraki, S.; Fukumoto, K.; Fukuda, O. Prediction of the muscle strength by the muscle thickness and hardness using ultrasound muscle hardness meter. Springerplus 2013, 2, 457. [Google Scholar] [CrossRef]
- Strasser, E.M.; Draskovits, T.; Praschak, M.; Quittan, M.; Graf, A. Association between ultrasound measurements of muscle thickness, pennation angle, echogenicity and skeletal muscle strength in the elderly. Age 2013, 35, 2377–2388. [Google Scholar] [CrossRef] [Green Version]
- Gonzales, J.M.; Galpin, A.J.; Montgomery, M.M.; Pamukoff, D.N. Comparison of lower limb muscle architecture and geometry in distance runners with rearfoot and forefoot strike pattern. J. Sports Sci. 2019, 37, 2184–2190. [Google Scholar] [CrossRef] [PubMed]
- Potier, T.G.; Alexander, C.M.; Seynnes, O.R. Effects of eccentric strength training on biceps femoris muscle architecture and knee joint range of movement. Eur. J. Appl. Physiol. 2009, 105, 939–944. [Google Scholar] [CrossRef] [PubMed]



| Group | Age (Years) | Height (cm) | Weight (kg) | Weekly Volume (km) | Running Years |
|---|---|---|---|---|---|
| FFS (n = 10) | 27.50 ± 8.68 | 175.75 ± 7.69 | 69.01 ± 6.96 | 39.50 ± 18.92 | 5.40 ± 3.47 |
| RFS (n = 10) | 30.10 ± 5.07 | 173.00 ± 5.16 | 69.59 ± 10.11 | 34.60 ± 13.72 | 4.00 ± 1.70 |
| NR (n = 10) | 25.40 ± 1.78 | 175.00 ± 4.74 | 68.01 ± 6.48 | / | / |
| p value | 0.147 | 0.675 | 0.910 | 0.516 | 0.272 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Wu, K.; Deng, L.; Liu, C.; Fu, W. The Effects of Habitual Foot Strike Patterns on the Morphology and Mechanical Function of the Medial Gastrocnemius–Achilles Tendon Unit. Bioengineering 2023, 10, 264. https://doi.org/10.3390/bioengineering10020264
Li L, Wu K, Deng L, Liu C, Fu W. The Effects of Habitual Foot Strike Patterns on the Morphology and Mechanical Function of the Medial Gastrocnemius–Achilles Tendon Unit. Bioengineering. 2023; 10(2):264. https://doi.org/10.3390/bioengineering10020264
Chicago/Turabian StyleLi, Lu, Kaicheng Wu, Liqin Deng, Cuixian Liu, and Weijie Fu. 2023. "The Effects of Habitual Foot Strike Patterns on the Morphology and Mechanical Function of the Medial Gastrocnemius–Achilles Tendon Unit" Bioengineering 10, no. 2: 264. https://doi.org/10.3390/bioengineering10020264