
Histological Characterization of Class I HLA Molecules in Whole Umbilical Cord Tissue Towards an Inexhaustible Graft Alternative for Reconstructive Surgery
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Immunohistochemistry
2.3. Hematoxylin and Eosin Stain
2.4. Tissue and Cell Lysate Preparation
2.5. Western Immunoblotting
2.6. Immunoprecipitation and Mass Spectrometry
3. Results
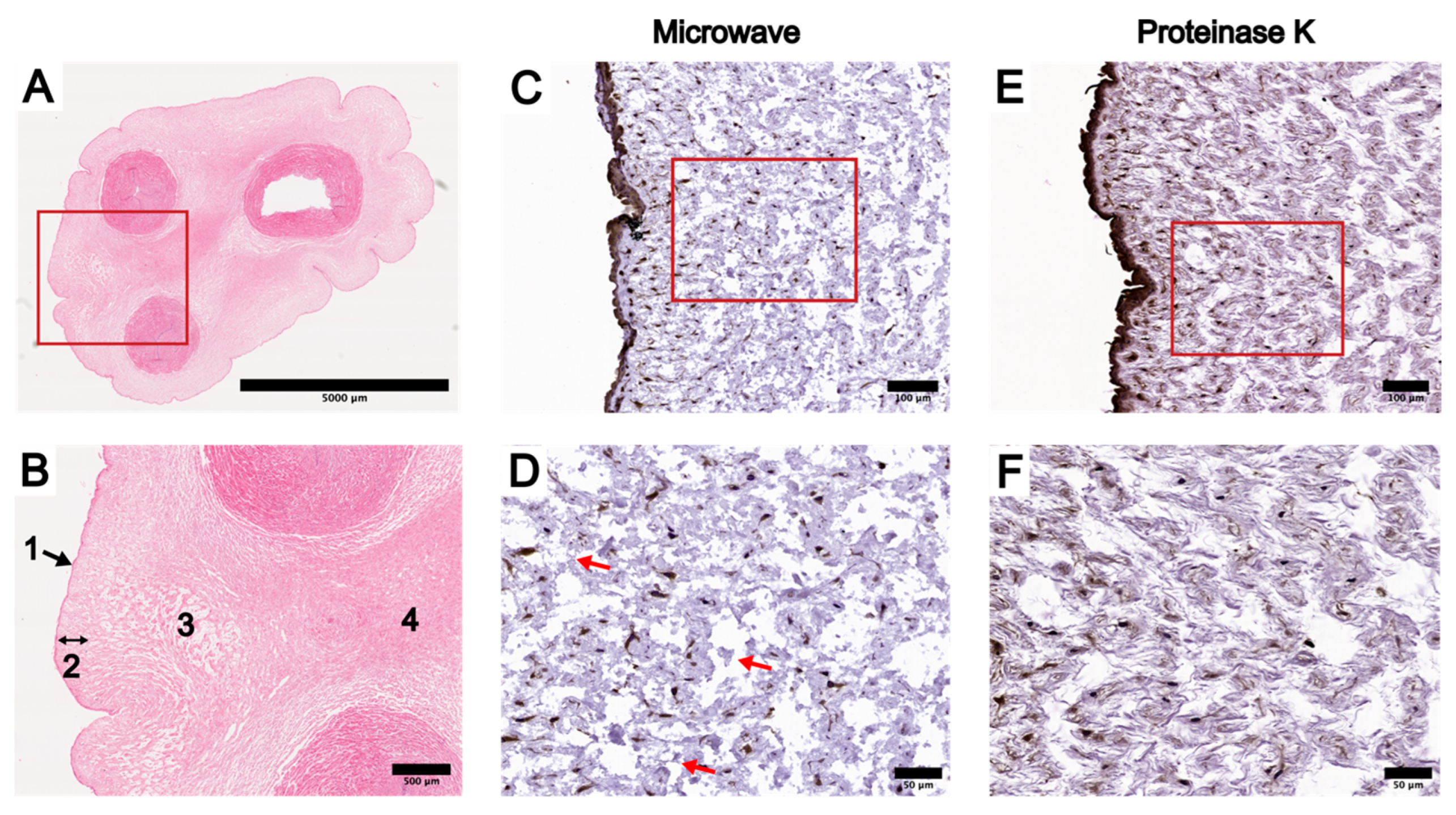
3.1. Optimization of IHC Protocol in UC Tissue Cross-Sections
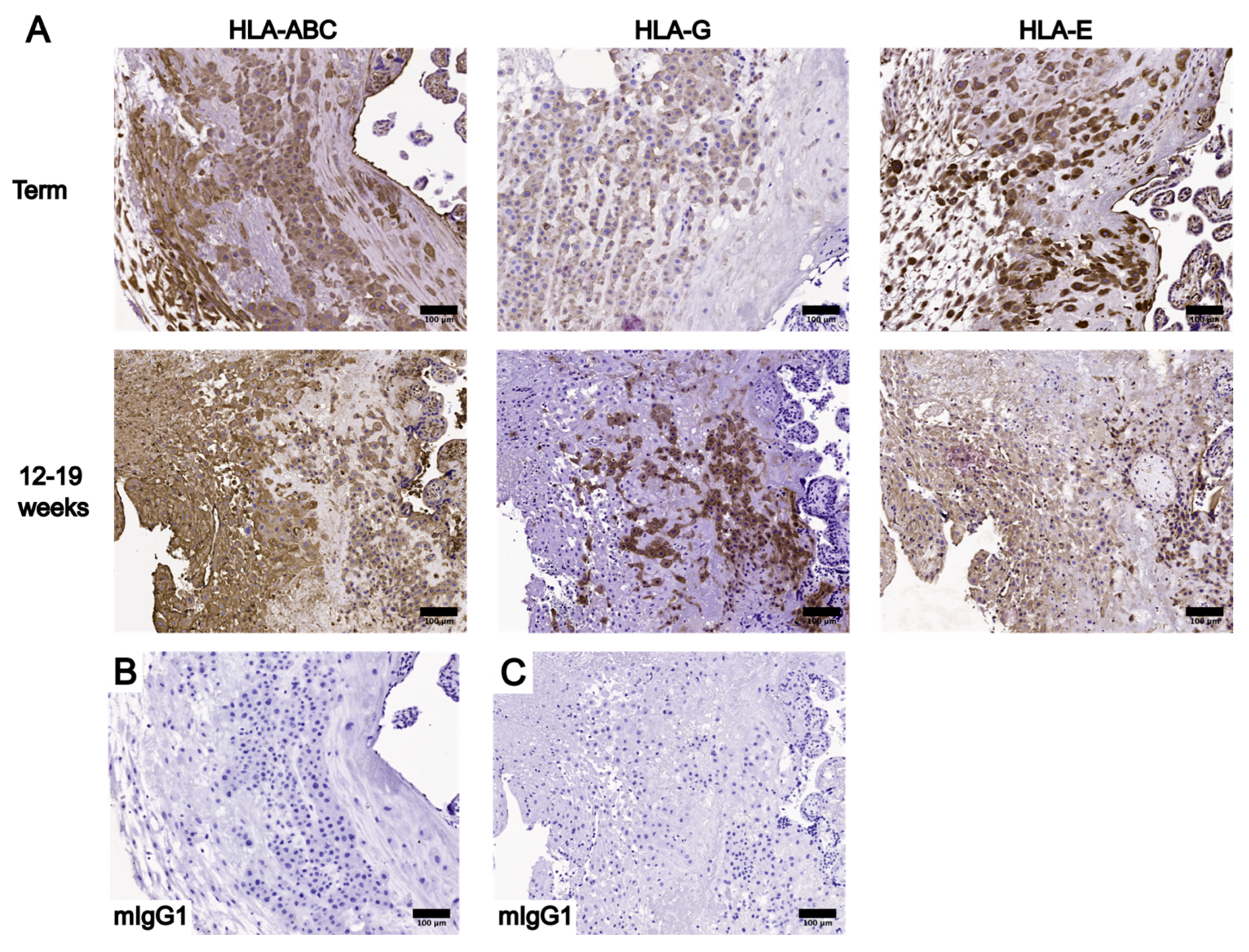
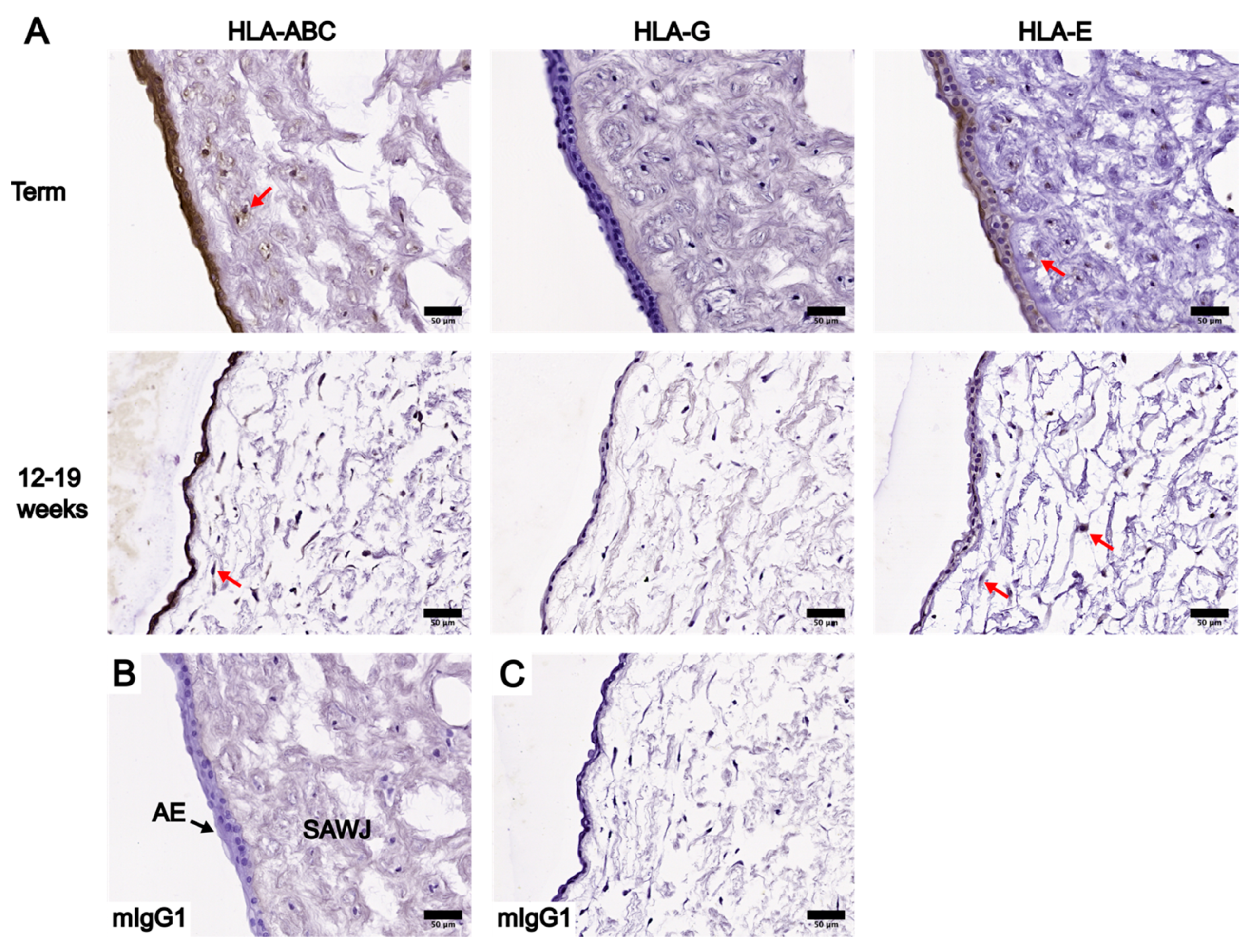
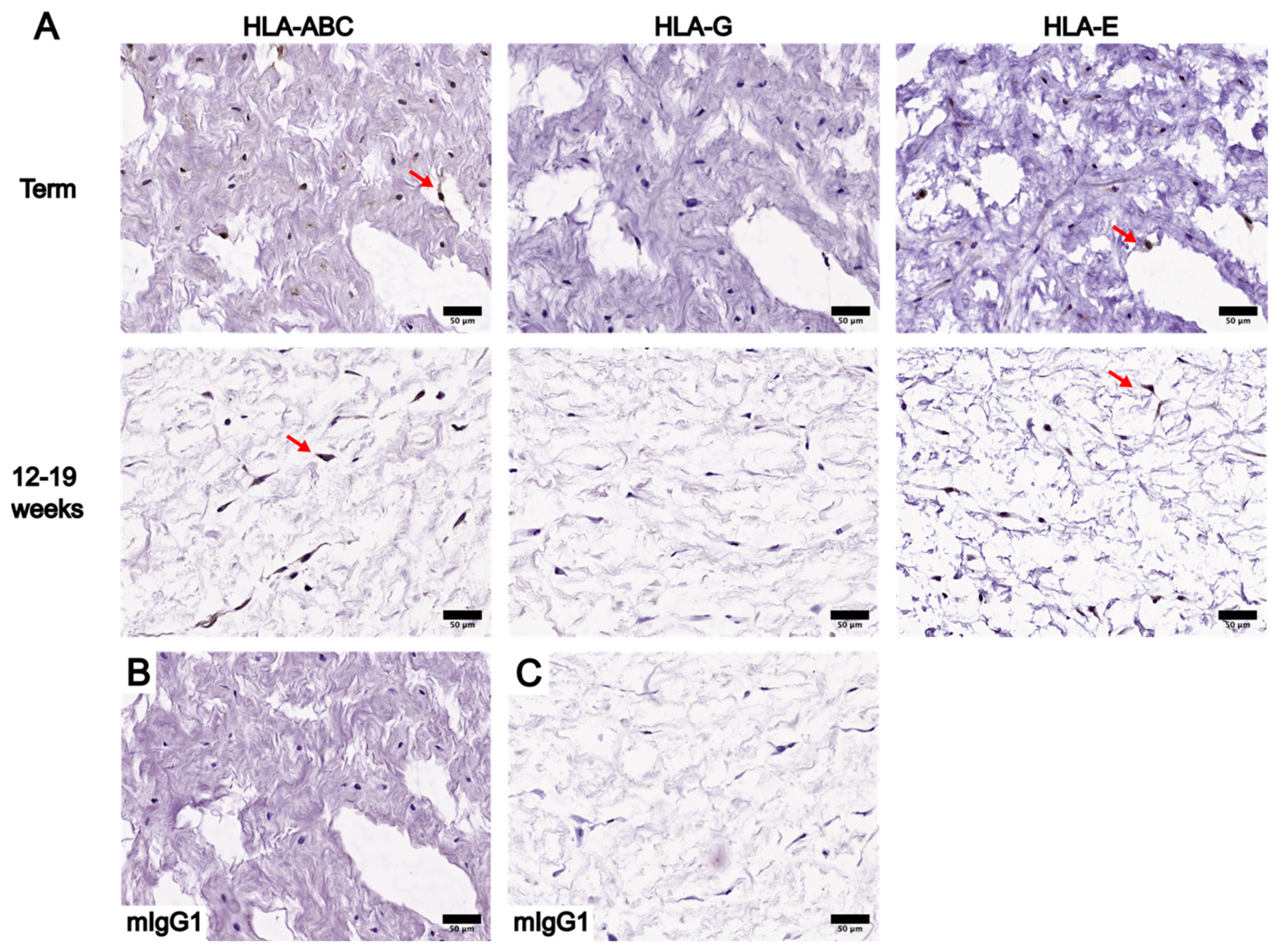
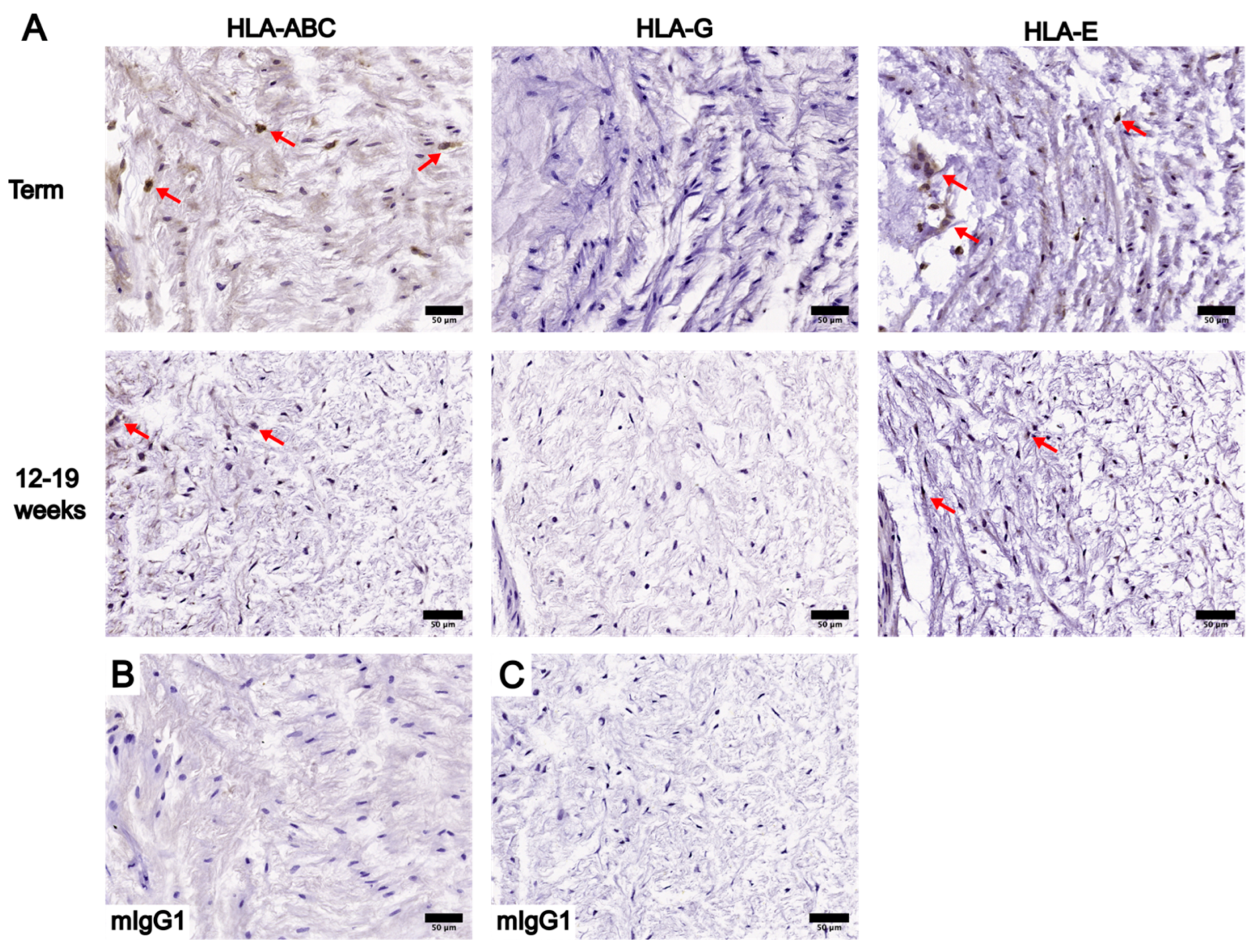
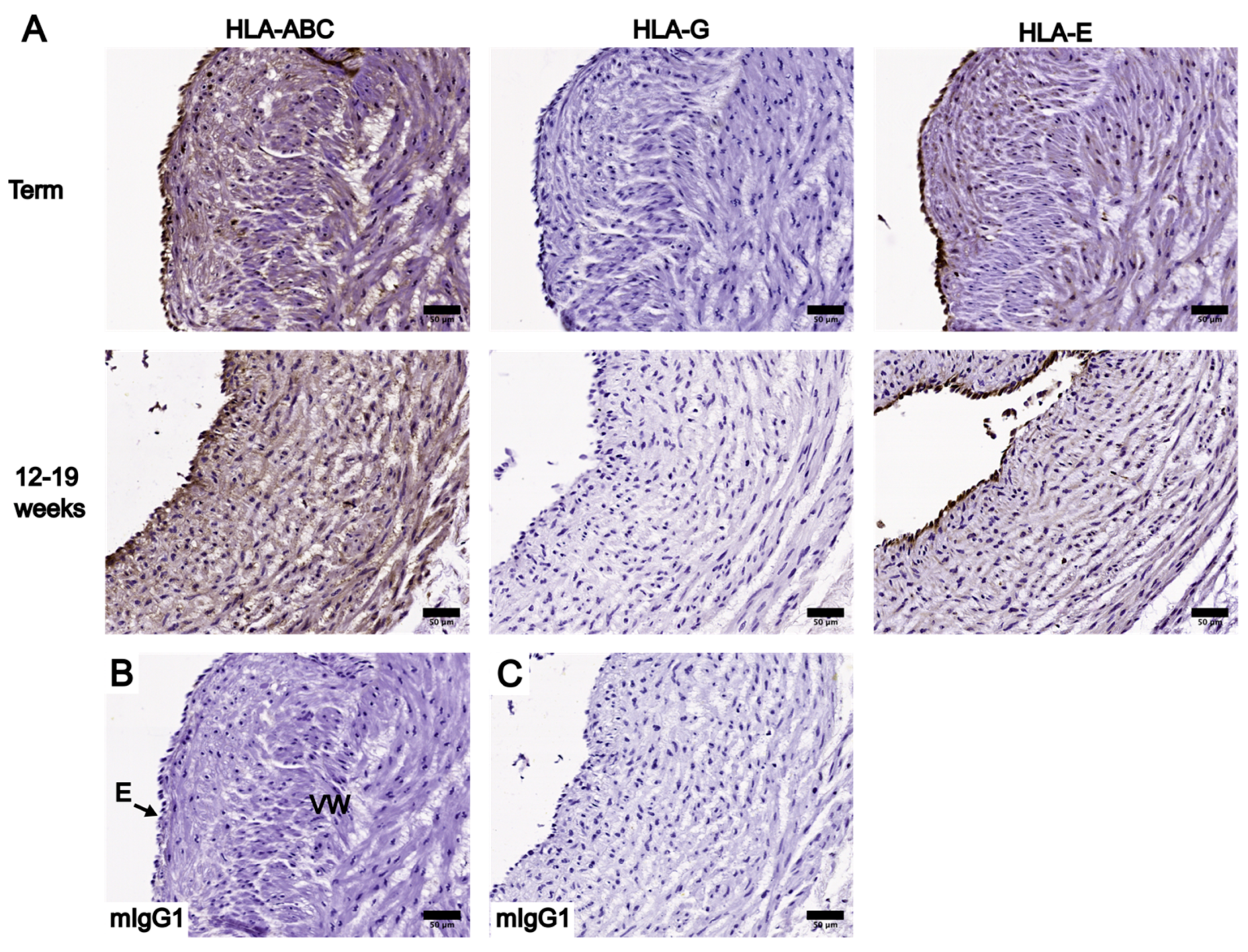
3.2. Localization and Levels of Class I HLA Molecules in UC
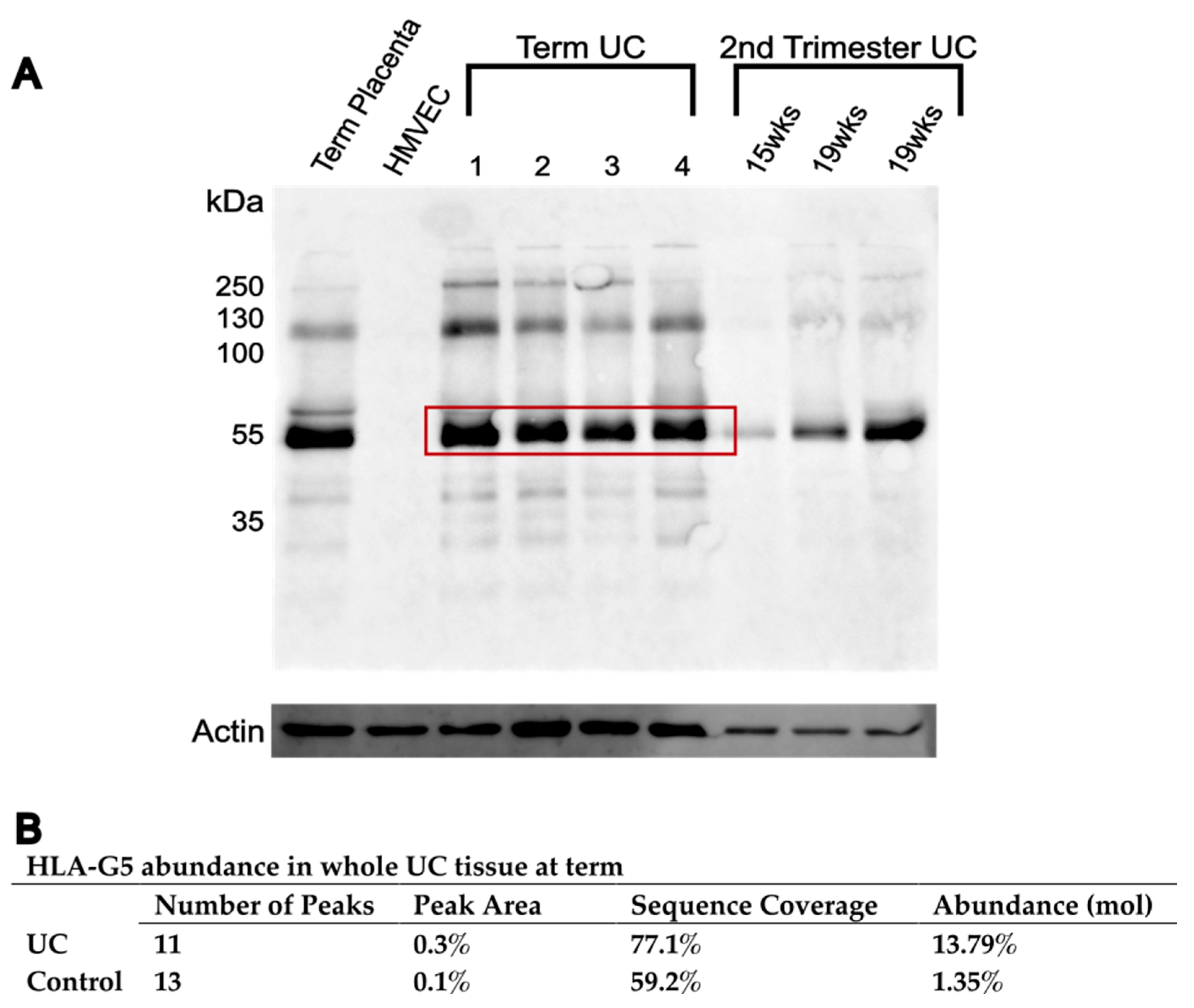
3.3. HLA-G Complex in Whole UC Tissue
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orabi, H.; Bouhout, S.; Morissette, A.; Rousseau, A.; Chabaud, S.; Bolduc, S. Tissue Engineering of Urinary Bladder and Urethra: Advances from Bench to Patients. Sci. World J. 2013, 2013, 154564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, L.S.; Ebbers, M.B. Hypospadias: Anatomy, Etiology, and Technique. J. Pediatr. Surg. 2006, 41, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Mangir, N.; Wilson, K.J.; Osman, N.I.; Chapple, C.R. Current State of Urethral Tissue Engineering. Curr. Opin. Urol. 2019, 29, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, W.; Bush, N.C. Re-Operative Urethroplasty after Failed Hypospadias Repair: How Prior Surgery Impacts Risk for Additional Complications. J. Pediatr. Urol. 2017, 13, 289.e1 –289.e6. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.B.; Luzzani, C.; Andrini, L.B.; Riccillo, F.; Buero, G.; Pelinski, P.; Inda, A.M.; Errecalde, A.L.; Miriuka, S.; Carosella, E.D.; et al. Wound Healing by Allogeneic Transplantation of Specific Subpopulation From Human Umbilical Cord Mesenchymal Stem Cells. Cell Transplant. 2021, 30, 963689721993774. [Google Scholar] [CrossRef]
- Szaraz, P.; Librach, M.; Maghen, L.; Iqbal, F.; Barretto, T.A.; Kenigsberg, S.; Gauthier-Fisher, A.; Librach, C.L. In Vitro Differentiation of First Trimester Human Umbilical Cord Perivascular Cells into Contracting Cardiomyocyte-Like Cells. Stem Cells Int. 2016, 2016, 7513252. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhang, N.; Li, T.; Guo, J.; Wang, Z.; Yang, M.; Gao, L. Human Umbilical Cord Wharton’s Jelly Stem Cells: Immune Property Genes Assay and Effect of Transplantation on the Immune Cells of Heart Failure Patients. Cell. Immunol. 2012, 276, 83–90. [Google Scholar] [CrossRef]
- Liu, S.; Yuan, M.; Hou, K.; Zhang, L.; Zheng, X.; Zhao, B.; Sui, X.; Xu, W.; Lu, S.; Guo, Q. Immune Characterization of Mesenchymal Stem Cells in Human Umbilical Cord Wharton’s Jelly and Derived Cartilage Cells. Cell. Immunol. 2012, 278, 35–44. [Google Scholar] [CrossRef]
- Huang, L.; Wong, Y.P.; Gu, H.; Cai, Y.J.; Ho, Y.; Wang, C.C.; Leung, T.Y.; Burd, A. Stem Cell-like Properties of Human Umbilical Cord Lining Epithelial Cells and the Potential for Epidermal Reconstitution. Cytotherapy 2011, 13, 145–155. [Google Scholar] [CrossRef]
- Zhou, Y.; Gan, S.U.; Lin, G.; Lim, Y.T.; Masilamani, J.; Mustafa, F.B.; Phua, M.L.; Rivino, L.; Phan, T.T.; Lee, K.O.; et al. Characterization of Human Umbilical Cord Lining-Derived Epithelial Cells and Transplantation Potential. Cell Transplant. 2011, 20, 1827–1841. [Google Scholar] [CrossRef]
- Wang, L.; Kang, Y.; Yan, H.; Zhu, X.; Zhu, T.; Jiang, J.; Zhao, J. Tendon Regeneration Induced by Umbilical Cord Graft in a Rabbit Tendon Defect Model. J. Tissue Eng. Regen. Med. 2020, 14, 1009–1018. [Google Scholar] [CrossRef]
- Sundblad, K.W.; Tassis, E.K. A Quality Improvement Pilot Assessment of the Safety and Associated Outcomes of a Viable Cryopreserved Umbilical Tissue Allograft as an Adjunct Surgical Wrap in Peroneus Brevis Tendon Repair. Medicine 2018, 97, e13662. [Google Scholar] [CrossRef]
- Ferdous, K.M.N.U.; Hasan, M.S.; Kabir, K.A.; Zahid, M.K.; Islam, M.K.; Hasanuzzaman, M. Use of Umbilical Cord Flap for Closure of Gastroschisis. J. Shaheed Suhrawardy Med. Coll. 2018, 10, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Marston, W.A.; Lantis, J.C.; Wu, S.C.; Nouvong, A.; Clements, J.R.; Lee, T.D.; McCoy, N.D.; Slade, H.B.; Tseng, S.C. One-Year Safety, Healing and Amputation Rates of Wagner 3–4 Diabetic Foot Ulcers Treated with Cryopreserved Umbilical Cord (TTAX01). Wound Repair Regen. 2020, 28, 526–531. [Google Scholar] [CrossRef]
- Marston, W.A.; Lantis, J.C.; Wu, S.C.; Nouvong, A.; Lee, T.D.; McCoy, N.D.; Slade, H.B.; Tseng, S.C. An Open-Label Trial of Cryopreserved Human Umbilical Cord in the Treatment of Complex Diabetic Foot Ulcers Complicated by Osteomyelitis. Wound Repair Regen. 2019, 27, 680–686. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.E.; Walker, J.T.; Keating, A. Concise Review: Wharton’s Jelly: The Rich, but Enigmatic, Source of Mesenchymal Stromal Cells. Stem. Cells Transl. Med. 2017, 6, 1620–1630. [Google Scholar] [CrossRef]
- Gupta, A.; El-Amin, S.F.; Levy, H.J.; Sze-Tu, R.; Ibim, S.E.; Maffulli, N. Umbilical Cord-Derived Wharton’s Jelly for Regenerative Medicine Applications. J. Orthop. Surg. Res. 2020, 15, 49. [Google Scholar] [CrossRef] [Green Version]
- Leng, Z.; Li, L.; Zhou, X.; Dong, G.; Chen, S.; Shang, G.; Kou, H.; Yang, B.; Liu, H. Novel Insights into the Stemness and Immune Privilege of Mesenchymal Stem Cells from Human Wharton Jelly by Single-Cell RNA Sequencing. Med. Sci. Monit. 2021, 28, e934660. [Google Scholar] [CrossRef]
- Kim, J.H.; Jo, C.H.; Kim, H.R.; Hwang, Y.I. Comparison of Immunological Characteristics of Mesenchymal Stem Cells from the Periodontal Ligament, Umbilical Cord, and Adipose Tissue. Stem Cells Int. 2018, 2018, 8429042. [Google Scholar] [CrossRef]
- Ding, D.C.; Chou, H.L.; Chang, Y.H.; Hung, W.T.; Liu, H.W.; Chu, T.Y. Characterization of HLA-G and Related Immunosuppressive Effects in Human Umbilical Cord Stroma-Derived Stem Cells. Cell Transplant. 2016, 25, 217–228. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Q.; Wang, Z.; Tong, H.; Ma, L.; Zhang, Y.; Shan, F.; Meng, Y.; Yuan, Z. Comparative Analysis of Human Mesenchymal Stem Cells from Fetal-Bone Marrow, Adipose Tissue, and Warton’s Jelly as Sources of Cell Immunomodulatory Therapy. Hum. Vaccines Immunother. 2016, 12, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Mennan, C.; Wright, K.; Bhattacharjee, A.; Balain, B.; Richardson, J.; Roberts, S. Isolation and Characterisation of Mesenchymal Stem Cells from Different Regions of the Human Umbilical Cord. BioMed Res. Int. 2013, 2013, 916136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mennan, C.; Brown, S.; McCarthy, H.; Mavrogonatou, E.; Kletsas, D.; Garcia, J.; Balain, B.; Richardson, J.; Roberts, S. Mesenchymal Stromal Cells Derived from Whole Human Umbilical Cord Exhibit Similar Properties to Those Derived from Wharton’s Jelly and Bone Marrow. FEBS Openbio 2016, 6, 1054–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.J.; Huang, L.; Leung, T.Y.; Burd, A. A Study of the Immune Properties of Human Umbilical Cord Lining Epithelial Cells. Cytotherapy 2014, 16, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Deuse, T.; Stubbendorff, M.; Tang-Quan, K.; Phillips, N.; Kay, M.A.; Eiermann, T.; Phan, T.T.; Volk, H.D.; Reichenspurner, H.; Robbins, R.C.; et al. Immunogenicity and Immunomodulatory Properties of Umbilical Cord Lining Mesenchymal Stem Cells. Cell Transplant. 2011, 20, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Marino, J.; Paster, J.; Benichou, G. Allorecognition by T Lymphocytes and Allograft Rejection. Front. Immunol. 2016, 7, 582. [Google Scholar] [CrossRef] [Green Version]
- Mallis, P.; Boulari, D.; Michalopoulos, E.; Dinou, A.; Spyropoulou-Vlachou, M.; Stavropoulos-Giokas, C. Evaluation of HLA-G Expression in Multipotent Mesenchymal Stromal Cells Derived from Vitrified Wharton’s Jelly Tissue. Bioengineering 2018, 5, 95. [Google Scholar] [CrossRef] [Green Version]
- Stubbendorff, M.; Deuse, T.; Hua, X.; Phan, T.T.; Bieback, K.; Atkinson, K.; Eiermann, T.H.; Velden, J.; Schröder, C.; Reichenspurner, H.; et al. Immunological Properties of Extraembryonic Human Mesenchymal Stromal Cells Derived from Gestational Tissue. Stem Cells Dev. 2013, 22, 2619–2629. [Google Scholar] [CrossRef] [Green Version]
- Alegre, E.; Rizzo, R.; Bortolotti, D.; Fernandez-Landázuri, S.; Fainardi, E.; González, A. Some Basic Aspects of HLA-G Biology. J. Immunol. Res. 2014, 2014, 657625. [Google Scholar] [CrossRef] [Green Version]
- Lila, N.; Carpentier, A.; Amrein, C.; Khalil-Daher, I.; Dausset, J.; Carosella, E.D. Implication of HLA-G Molecule in Heart-Graft Acceptance. Lancet 2000, 355, 2138. [Google Scholar] [CrossRef]
- Iwaszko, M.; Bogunia-Kubik, K. Clinical Significance of the HLA-E and CD94/NKG2 Interaction. Arch Immunol. Ther. Exp. 2011, 59, 353–367. [Google Scholar] [CrossRef]
- King, A.; Allan, D.S.J.; Bowen, M.; Powis, S.J.; Joseph, S.; Verma, S.; Hiby, S.E.; Mcmichael, A.J.; Loke, Y.W.; Braud, V.M. HLA-E Is Expressed on Trophoblast and Interacts with CD94/NKG2 Receptors on Decidual NK Cells. Eur. J. Immunol. 2000, 30, 1623–1631. [Google Scholar] [CrossRef]
- Grant, E.J.; Nguyen, A.T.; Lobos, C.A.; Szeto, C.; Chatzileontiadou, D.S.M.; Gras, S. The Unconventional Role of HLA-E: The Road Less Traveled. Mol. Immunol. 2020, 120, 101–112. [Google Scholar] [CrossRef]
- Battin, C.; Kaufmann, G.; Leitner, J.; Tobias, J.; Wiedermann, U.; Rölle, A.; Meyer, M.; Momburg, F.; Steinberger, P. NKG2A-checkpoint Inhibition and Its Blockade Critically Depends on Peptides Presented by Its Ligand HLA-E. Immunology 2022, 166, 507–521. [Google Scholar] [CrossRef]
- Lee, D.K.; Sengupta, A.; Nevo, O. The Effect of Magnesium Sulfate on Gene Expression in Maternal Microvascular Endothelial Cells. Hypertens. Pregnancy 2018, 37, 30–36. [Google Scholar] [CrossRef]
- Nevo, O.; Soleymanlou, N.; Wu, Y.; Xu, J.; Kingdom, J.; Many, A.; Zamudio, S.; Caniggia, I. Increased Expression of SFlt-1 in In Vivo and In Vitro Models of Human Placental Hypoxia Is Mediated by HIF-1. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R1085–R1093. [Google Scholar] [CrossRef]
- Swets, M.; Wouters, A.; Krijgsman, D.; van Vlierberghe, R.L.P.; Boot, A.; van Eendenburg, J.D.; van Wezel, T.; Gelderblom, H.; van de Velde, C.J.H.; van den Elsen, P.J.; et al. HLA-G Protein Expression in Colorectal Cancer Evaluated by Immunohistochemistry and Western Blot Analysis: Its Expression Characteristics Remain Enigmatic. Clin. Immunol. 2018, 194, 80–86. [Google Scholar] [CrossRef]
- Braud, V.; Jones, E.Y.; McMichael, A. The Human Major Histocompatibility Complex Class Ib Molecule HLA-E Binds Signal Sequence-Derived Peptides with Primary Anchor Residues at Positions 2 and 9. Eur. J. Immunol. 1997, 27, 1164–1169. [Google Scholar] [CrossRef]
- Braud, V.M.; Allan, D.S.J.; Wilson, D.; Mcmichael, A.J. TAP- and Tapasin-Dependent HLA-E Surface Expression Correlates with the Binding of an MHC Class I Leader Peptide. Curr. Biol. 1998, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wieten, L.; Mahaweni, N.M.; Voorter, C.E.M.; Bos, G.M.J.; Tilanus, M.G.J. Clinical and Immunological Significance of HLA-E in Stem Cell Transplantation and Cancer. Tissue Antigens 2014, 84, 523–535. [Google Scholar] [CrossRef]
- Kaiser, B.K.; Barahmand-pour, F.; Paulsene, W.; Medley, S.; Geraghty, D.E.; Strong, R.K. Interactions between NKG2x Immunoreceptors and HLA-E Ligands Display Overlapping Affinities and Thermodynamics. J. Immunol. 2005, 174, 2878–2884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braud, V.M.; Allan, D.S.J.; O’Callaghan, C.A.; Söderström, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E Binds to Natural Killer Cell Receptors CD94/NKG2A, B, and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Tsamadou, C.; Fürst, D.; Vucinic, V.; Bunjes, D.; Neuchel, C.; Mytilineos, D.; Gramatzki, M.; Arnold, R.; Wagner, E.M.; Einsele, H.; et al. Human Leukocyte Antigen-E Mismatch Is Associated with Better Hematopoietic Stem Cell Transplantation Outcome in Acute Leukemia Patients. Haematologica 2017, 102, 1947–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guberina, H.; Rebmann, V.; Wagner, B.; da Silva Nardi, F.; Dziallas, P.; Dolff, S.; Bienholz, A.; Wohlschlaeger, J.; Bankfalvi, A.; Heinemann, F.M.; et al. Association of High HLA-E Expression during Acute Cellular Rejection and Numbers of HLA Class I Leader Peptide Mismatches with Reduced Renal Allograft Survival. Immunobiology 2017, 222, 536–543. [Google Scholar] [CrossRef]
- Choi, J.Y.; Eskandari, S.K.; Cai, S.; Sulkaj, I.; Assaker, J.P.; Allos, H.; Alhaddad, J.; Muhsin, S.A.; Alhussain, E.; Mansouri, A.; et al. Regulatory CD8 T Cells That Recognize Qa-1 Expressed by CD4 T-Helper Cells Inhibit Rejection of Heart Allografts. Proc. Natl. Acad. Sci. USA 2020, 117, 6042–6046. [Google Scholar] [CrossRef]
- Halaseh, S.A.; Halaseh, S.; Ashour, M. Hypospadias: A Comprehensive Review Including Its Embryology, Etiology and Surgical Techniques. Cureus 2022, 14, e27544. [Google Scholar] [CrossRef]
- Cunnane, E.M.; Davis, N.F.; Cunnane, C.V.; Lorentz, K.L.; Ryan, A.J.; Hess, J.; Weinbaum, J.S.; Walsh, M.T.; O’Brien, F.J.; Vorp, D.A. Mechanical, Compositional and Morphological Characterisation of the Human Male Urethra for the Development of a Biomimetic Tissue Engineered Urethral Scaffold. Biomaterials 2021, 269, 120651. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | |
|---|---|
| n | 10 |
| Maternal age (year) | 34.6 ± 2.1 |
| Gestational age (weeks) | 38.8 ± 0.3 |
| Body mass index (kg/m2) | 32 ± 2.9 |
| Gravidity | 3 ± 0.6 |
| Parity | 1 ± 0.3 |
| Blood pressure (mmHg) | |
| Systolic | 109 ± 1.8 |
| Diastolic | 65 ± 1.5 |
| Proteinuria (plus protein) | 0 ± 0 |
| Birth weight (g) | 3594 ± 158.7 |
| Antibody | Clone | Species | Working Concentration (µg/mL) | Source |
|---|---|---|---|---|
| HLA-ABC | EMR8-5 | Mouse | 0.25 | BD Biosciences (cat 565292) |
| HLA-G | MEM-G/1 | Mouse | 2.0 | Thermo Fisher (MA1-19219) |
| 4H84 | Mouse | 2.0 | Exbio (cat 11-499-C100) | |
| HLA-E | MEM-E/02 | Mouse | 5.0 | Abcam (cat ab2216) |
| AR | Fixation Condition | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fixed for 24 h | Fixed for 48 h | ||||||||
| With Agitation | No Agitation | With Agitation | No Agitation | ||||||
| 4C | RT | 4C | RT | 4C | RT | 4C | RT | ||
| Mic | AE | ++# | ++# | ++# | ++# | ++ | ++# | ++ | ++# |
| WJ * | ++ (Present) | ++# (Present) | ++# (Present) | ++# (Present) | ++# (Present) | ++# (Present) | ++# (Present) | ++ (Present) | |
| E | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | |
| VW | + | +# | +# | +# | +# | +# | +# | +# | |
| PC | AE | ++ | ++ | ++ | ++ | ++# | ++ | ++ | ++# |
| WJ* | ++ (Present) | ++# (Present) | ++# (Present) | ++ (Present) | ++ (Present) | ++# (Present) | ++# (Present) | ++ (Present) | |
| E | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | |
| VW | +# | ++# | ++ | ++# | ++# | ++# | +# | +# | |
| Trypsin | AE | +++# | ++# | ++# | +++# | +++# | ++ | +++# | ++# |
| WJ * | ++# (Absent) | ++# (Absent) | ++ (Absent) | ++ (Absent) | ++# (Absent) | ++ (Absent) | ++ (Absent) | ++ (Absent) | |
| E | ++~+++ | + | ++ | ++ | ++ | ++ | +++ | ++ | |
| VW | ++# | + | +# | + | ++# | ++ | ++# | ++ | |
| ProK | AE | +++ | +++# | +++ | +++# | +++# | +++# | +++# | +++# |
| WJ * | ++# (Absent) | ++ (Absent) | ++ (Absent) | ++ (Absent) | ++ (Absent) | ++ (Absent) | ++ (Absent) | ++ (Present) | |
| E | ++ | ++ | ++ | ++ | ++ | +++ | +++# | ++ | |
| VW | ++ | ++ | ++ | + | ++ | ++ | ++# | ++ | |
| No AR | AE | ++ | ++ | ++ | ++ | ++ | ++# | +~++ | ++# |
| WJ * | + (Absent) | + (Absent) | +++ (Absent) | ++ (Absent) | ++ (Absent) | + (Absent) | ++# (Absent) | ++ (Absent) | |
| E | + | + | + | ++ | ++ | ++ | + | ++ | |
| VW | + | + | + | + | ++ | + | + | + | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Y.Y.; Lee, D.K.; Jarvi, S.; Farshadi, M.; Sheng, M.; Mar, S.; Nevo, O.; Leong, H.S. Histological Characterization of Class I HLA Molecules in Whole Umbilical Cord Tissue Towards an Inexhaustible Graft Alternative for Reconstructive Surgery. Bioengineering 2023, 10, 110. https://doi.org/10.3390/bioengineering10010110
Yao YY, Lee DK, Jarvi S, Farshadi M, Sheng M, Mar S, Nevo O, Leong HS. Histological Characterization of Class I HLA Molecules in Whole Umbilical Cord Tissue Towards an Inexhaustible Graft Alternative for Reconstructive Surgery. Bioengineering. 2023; 10(1):110. https://doi.org/10.3390/bioengineering10010110
Chicago/Turabian StyleYao, Yue Ying, Dennis K. Lee, Stephanie Jarvi, Marjan Farshadi, Minzhi Sheng, Sara Mar, Ori Nevo, and Hon S. Leong. 2023. "Histological Characterization of Class I HLA Molecules in Whole Umbilical Cord Tissue Towards an Inexhaustible Graft Alternative for Reconstructive Surgery" Bioengineering 10, no. 1: 110. https://doi.org/10.3390/bioengineering10010110