
Impact of Pb Toxicity on the Freshwater Pearl Mussel, Lamellidens marginalis: Growth Metrics, Hemocyto-Immunology, and Histological Alterations in Gill, Kidney, and Muscle Tissue

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Live Animals and Rearing
2.2. Acute Toxicity Test and Preparing Concentrations for Chronic Exposure
2.3. Sampling and Observation of Hydrological Parameters of Rearing Water
2.4. Tools for Growth Analysis
2.5. Hemocyte Counts, Neutral Red Retention Assay, and Serum Lysosome Activity
2.6. Histology of Gill, Kidney, and Muscle
2.7. Statistical Tools
3. Results
3.1. Acute Toxicity Test
3.2. Water Quality Parameters in Different Trial Units
3.3. Growth Performance, Mortality Rate, and Condition Indices
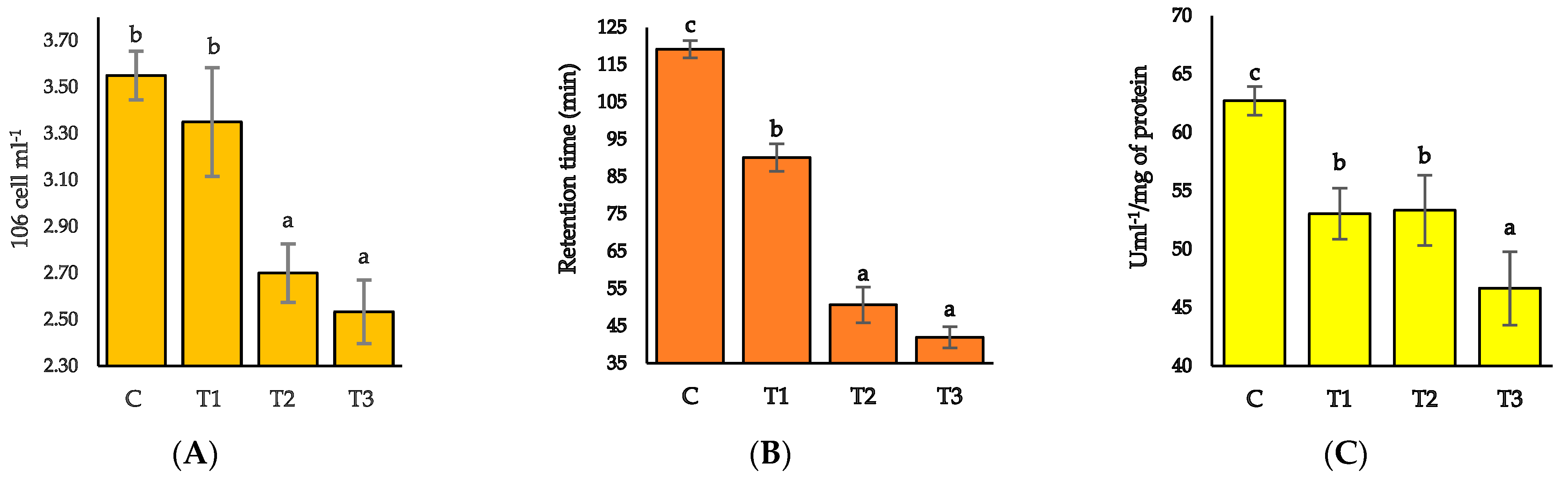
3.4. Hemocytes Count, Lysosomal Membrane Stability (LMS), and Serum Lysosome Activity
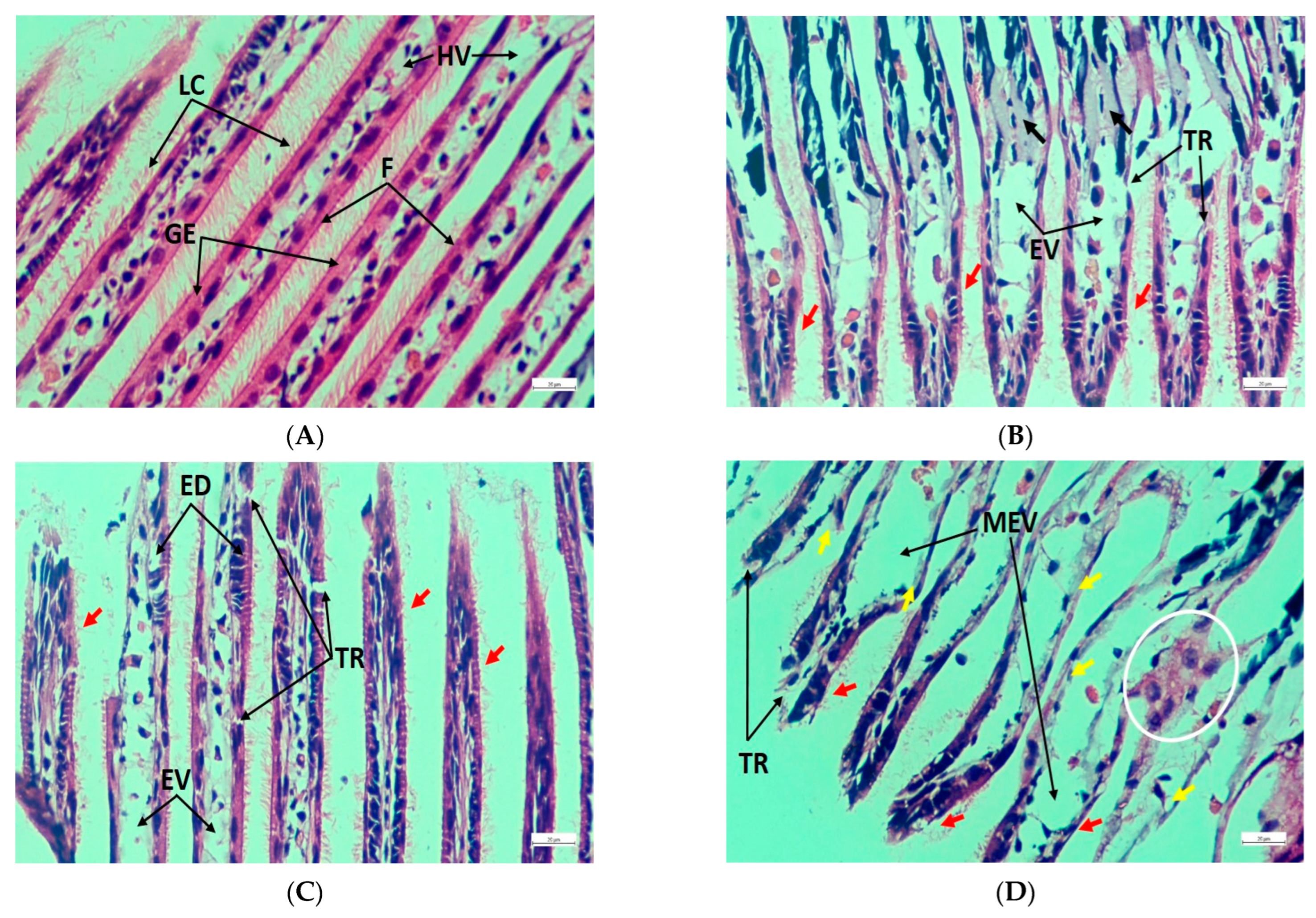
3.5. Histopathology of Gills, Kidneys, and Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, Y.; Rao, J.; Jia, Q. Heavy Metal Pollution and Environmental Risks in the Water of Rongna River Caused by Natural AMD around Tiegelongnan Copper Deposit, Northern Tibet, China. PLoS ONE 2022, 17, e0266700. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, A.H.; Chowdhury, G.; Adikari, D.; Jahan, I.; Andrawina, Y.O.; Hossain, M.A.; Schneider, P.; Iqbal, M.M. Macroplastics Pollution in the Surma River in Bangladesh: A Threat to Fish Diversity and Freshwater Ecosystems. Water 2022, 14, 3263. [Google Scholar] [CrossRef]
- Islam, M.S.; Tanaka, M. Impacts of Pollution on Coastal and Marine Ecosystems Including Coastal and Marine Fisheries and Approach for Management: A Review and Synthesis. Mar. Pollut. Bull. 2004, 48, 624–649. [Google Scholar] [CrossRef] [PubMed]
- Mokarat, M.; Ruangsakunporn, A.; Chaianunporn, T.; Lomthaisong, K.; Robson, M.G.; Keithmaleesatti, S. Heavy Metal Contamination and Bioaccumulation in Khorat Snail-Eating Turtles (Malayemys khoratensis) from an e-Waste Recycling Activity Site in Thailand. Hum. Ecol. Risk Assess. 2022, 28, 133–153. [Google Scholar] [CrossRef]
- Abdel Gawad, S.S. Acute Toxicity of Some Heavy Metals to the Fresh Water Snail, Theodoxus niloticus (Reeve, 1856). Egypt. J. Aquat. Res. 2018, 44, 83–87. [Google Scholar] [CrossRef]
- Mahi, T.F.; Chowdhury, G.; Hossain, M.A.; Baishnab, A.K.; Schneider, P.; Iqbal, M.M. Assessment of Lead (Pb) Toxicity in Juvenile Nile Tilapia, Oreochromis niloticus—Growth, Behaviour, Erythrocytes Abnormalities, and Histological Alterations in Vital Organs. Toxics 2022, 10, 793. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.S.; Routh, J.; Luo, C.; Dario, M.; Miao, Y.; Luo, D.; Wei, L. Metal Accumulations in Aquatic Organisms and Health Risks in an Acid Mine-Affected Site in South China. Environ. Geochem. Health 2021, 43, 4415–4440. [Google Scholar] [CrossRef]
- Inoue, K.; Onitsuka, Y.; Koito, T. Mussel Biology: From the Byssus to Ecology and Physiology, Including Microplastic Ingestion and Deep-Sea Adaptations. Fish. Sci. 2021, 87, 761–771. [Google Scholar] [CrossRef]
- Flora, G.; Gupta, D.; Tiwari, A. Toxicity of Lead: A Review with Recent Updates. Interdiscip. Toxicol. 2012, 5, 47–58. [Google Scholar] [CrossRef]
- Hussain, M.; Hossain, M.A.; Begum, M.; Roy, N.C. Freshwater Mussel (Lamelliedens marginalis) to Reduce the Lead (Pb) Bioaccumulation in Aquaculture of Stinging Catfish, Heteropneustes fossilis. J. Appl. Aquac. 2022, 1–17. [Google Scholar] [CrossRef]
- Usman, K.; Abu-Dieyeh, M.H.; Zouari, N.; Al-Ghouti, M.A. Lead (Pb) Bioaccumulation and Antioxidative Responses in Tetraena qataranse. Sci. Rep. 2020, 10, 17070. [Google Scholar] [CrossRef] [PubMed]
- Zalaghi, R.; Safari-Sinegani, A.-A. The Importance of Different Forms of Pb on Diminishing Biological Activities in a Calcareous Soil. Chem. Ecol. 2014, 30, 446–462. [Google Scholar] [CrossRef]
- Huang, H.; Li, Y.; Zheng, X.; Wang, Z.; Wang, Z.; Cheng, X. Nutritional Value and Bioaccumulation of Heavy Metals in Nine Commercial Fish Species from Dachen Fishing Ground, East China Sea. Sci. Rep. 2022, 12, 6927. [Google Scholar] [CrossRef]
- Chandurvelan, R.; Marsden, I.D.; Glover, C.N.; Gaw, S. Assessment of a Mussel as a Metal Bioindicator of Coastal Contamination: Relationships between Metal Bioaccumulation and Multiple Biomarker Responses. Sci. Total Environ. 2015, 511, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Khalesi, M.K.; Abedi, Z.; Behrouzi, S.; Kohestan Eskandari, S. Haematological, Blood Biochemical and Histopathological Effects of Sublethal Cadmium and Lead Concentrations in Common Carp. Bulg. J. Vet. Med. 2017, 20, 141–150. [Google Scholar] [CrossRef]
- Abdel-Warith, A.W.A.; Younis, E.S.M.I.; Al-Asgah, N.A.; Rady, A.M.; Allam, H.Y. Bioaccumulation of Lead Nitrate in Tissues and Its Effects on Hematological and Biochemical Parameters of Clarias gariepinus. Saudi J. Biol. Sci. 2020, 27, 840–845. [Google Scholar] [CrossRef]
- Shuhaimi-Othman, M.; Nur-Amalina, R.; Nadzifah, Y. Toxicity of Metals to a Freshwater Snail, Melanoides Tuberculata. Sci. World J. 2012, 2012, 1–10. [Google Scholar] [CrossRef]
- Kiran, K.; Midhun, S.J.; Vysakh, A.; James, T.J. Antagonistic Effects of Dietary Moringa oleifera on Hemato-biochemical and Oxidative Stress of Lead Nitrate Intoxicated Nile Tilapia, Oreochromis niloticus. Aquac. Res. 2021, 52, 6164–6178. [Google Scholar] [CrossRef]
- Saleh, I.; Syamsir, S.; Pramaningsih, V.; Hansen, H. The Use of Green Mussel as Bioindicator of Heavy Metal Pollution in Indonesia: A Review. Environ. Health Toxicol. 2021, 36, e2021026. [Google Scholar] [CrossRef]
- Le Saux, A.; David, E.; Betoulle, S.; Bultelle, F.; Rocher, B.; Barjhoux, I.; Cosio, C. New Insights into Cellular Impacts of Metals in Aquatic Animals. Environments 2020, 7, 46. [Google Scholar] [CrossRef]
- Arisekar, U.; Jeya, R.; Shalini, R.; Jeyasekaran, G. Human Health Risk Assessment of Heavy Metals in Aquatic Sediments and Freshwater Fish Caught from Thamirabarani River, the Western Ghats of South Tamil Nadu. Mar. Pollut. Bull. 2020, 159, 111496. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hussain, M.; Sarker, T.R.; Saha, S.; Iqbal, M.M. Reproductive and Morphometric Traits of Freshwater Mussel Lamellidens marginalis and Associated Hydrology in the Ratargul Freshwater Swamp Forest, Bangladesh. Egypt. J. Aquat. Res. 2022. [Google Scholar] [CrossRef]
- Nobles, T.; Zhang, Y. Survival, Growth and Condition of Freshwater Mussels: Effects of Municipal Wastewater Effluent. PLoS ONE 2015, 10, e0128488. [Google Scholar] [CrossRef] [PubMed]
- Yasmeen, S.; Pathan, T.S. Behavioural Response in Freshwater Bivalve Mollusk, Lamellidens marginalis Due to Acute Toxicity of Cadmium Chloride. Int. J. Biol. Innov. 2021, 3, 148–153. [Google Scholar] [CrossRef]
- Costello, D.M.; Brown, L.M.; Lamberti, G.A. Acute Toxic Effects of Ionic Liquids on Zebra Mussel (Dreissena polymorpha) Survival and Feeding. Green Chem. 2009, 11, 548–555. [Google Scholar] [CrossRef]
- Yap, C.K.; Sharifinia, M.; Cheng, W.H.; Al-Shami, S.A.; Wong, K.W.; Al-Mutairi, K.A. A Commentary on the Use of Bivalve Mollusks in Monitoring Metal Pollution Levels. Int. J. Environ. Res. Public Health 2021, 18, 3386. [Google Scholar] [CrossRef]
- EPA, U. Guideline No. 497: Defined Approaches on Skin Sensitisation; OECD Guidelines for the Testing of Chemicals, Section 4; OECD: Paris, France, 2021; ISBN 9789264903005. [Google Scholar]
- Brahma, N.; Gupta, A. Acute Toxicity of Lead in Fresh Water Bivalves Lamellidens jenkinsianus Obesa and Parreysia (Parreysia) Corrugata with Evaluation of Sublethal Effects on Acetylcholinesterase and Catalase Activity, Lipid Peroxidation, and Behavior. Ecotoxicol. Environ. Saf. 2020, 189, 109939. [Google Scholar] [CrossRef]
- Brix, K.V.; Esbaugh, A.J.; Grosell, M. The Toxicity and Physiological Effects of Copper on the Freshwater Pulmonate Snail, Lymnaea stagnalis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 154, 261–267. [Google Scholar] [CrossRef]
- D’Agata, A.; Cappello, T.; Maisano, M.; Parrino, V.; Giannetto, A.; Brundo, M.V.; Ferrante, M.; Mauceri, A. Cellular Biomarkers in the Mussel Mytilus galloprovincialis (Bivalvia: Mytilidae) from Lake Faro (Sicily, Italy). Ital. J. Zool. 2014, 81, 43–54. [Google Scholar] [CrossRef]
- Auguste, M.; Balbi, T.; Ciacci, C.; Canonico, B.; Papa, S.; Borello, A.; Vezzulli, L.; Canesi, L. Shift in Immune Parameters After Repeated Exposure to Nanoplastics in the Marine Bivalve Mytilus. Front. Immunol. 2020, 11, 426. [Google Scholar] [CrossRef]
- Kolyuchkina, G.A.; Ismailov, A.D. Morpho-Functional Characteristics of Bivalve Mollusks under the Experimental Environmental Pollution by Heavy Metals. Oceanology 2011, 51, 804–813. [Google Scholar] [CrossRef]
- Fahmy, S.R.; Sayed, D.A. Toxicological Perturbations of Zinc Oxide Nanoparticles in the Coelatura aegyptiaca Mussel. Toxicol. Ind. Health 2017, 33, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Vasanthi, L.; Revathi, P.; Arulvasu, C.; Munuswamy, N. Biomarkers of Metal Toxicity and Histology of Perna viridis from Ennore Estuary, Chennai, South East Coast of India. Ecotoxicol. Environ. Saf. 2012, 84, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Liberatori, G.; Grassi, G.; Guidi, P.; Bernardeschi, M.; Fiorati, A.; Scarcelli, V.; Genovese, M.; Faleri, C.; Protano, G.; Frenzilli, G.; et al. Effect-based Approach to Assess Nanostructured Cellulose Sponge Removal Efficacy of Zinc Ions from Seawater to Prevent Ecological Risks. Nanomaterials 2020, 10, 1283. [Google Scholar] [CrossRef]
- Paul, S.; Mandal, A.; Bhattacharjee, P.; Chakraborty, S.; Paul, R.; Kumar Mukhopadhyay, B. Evaluation of Water Quality and Toxicity after Exposure of Lead Nitrate in Fresh Water Fish, Major Source of Water Pollution. Egypt. J. Aquat. Res. 2019, 45, 345–351. [Google Scholar] [CrossRef]
- Ramakritinan, C.M.; Chandurvelan, R.; Kumaraguru, A.K. Acute Toxicity of Metals: Cu, Pb, Cd, Hg and Zn on Marine Molluscs, Cerithedia cingulata G., and Modiolus philippinarum H. Indian J. Mar. Sci. 2012, 41, 141–145. [Google Scholar]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: New York, NY, USA, 1971. [Google Scholar]
- Uddin, M.; Yang, H.-S.; Choi, K.-S.; Kim, H.-J.; Hong, J.-S.; Cho, M. Seasonal Changes in Perkinsus Olseni Infection and Gametogenesis in Manila Clam, Ruditapes philippinarum, from Seonjaedo Island in Incheon, off the West Coast of Korea. J. World Aquac. Soc. 2010, 41, 93–101. [Google Scholar] [CrossRef]
- Htun-Han, M. The Reproductive Biology of the Dab Limanda limanda (L.) in the North Sea: Seasonal Changes in the Ovary. J. Fish Biol. 1978, 13, 351–359. [Google Scholar] [CrossRef]
- Juhel, G.; Ramsay, R.M.; Davenport, J.; O’Halloran, J.; Culloty, S.C. Effect of the Microcystin-Producing Cyanobacterium, Microcystis aeruginosa, on Immune Functions of the Zebra Mussel Dreissena polymorpha. J. Shellfish Res. 2015, 34, 433–442. [Google Scholar] [CrossRef]
- Munari, M.; Matozzo, V.; Benetello, G.; Riedl, V.; Pastore, P.; Badocco, D.; Marin, M.G. Exposure to Decreased PH and Caffeine Affects Hemocyte Parameters in the Mussel Mytilus galloprovincialis. J. Mar. Sci. Eng. 2020, 8, 238. [Google Scholar] [CrossRef]
- Chu, F.-L.E.; La Peyre, J.F. Effect of Environmental Factors and Parasitism on Hemolymph Lysozyme and Protein of American Oysters (Crassostrea virginica). J. Invertebr. Pathol. 1989, 54, 224–232. [Google Scholar] [CrossRef]
- Capolupo, M.; Valbonesi, P.; Fabbri, E. A Comparative Assessment of the Chronic Effects of Micro-and Nano-Plastics on the Physiology of the Mediterranean Mussel Mytilus galloprovincialis. Nanomaterials 2021, 11, 649. [Google Scholar] [CrossRef] [PubMed]
- Slaoui, M.; Fiette, L. Histopathology Procedures: From Tissue Sampling to Histopathological Evaluation. Methods Mol. Biol. 2011, 691, 69–82. [Google Scholar] [CrossRef] [PubMed]
- McElwain, A.; Bullard, S.A. Histological Atlas of Freshwater Mussels (Bivalvia, Unionidae): Villosa nebulosa (Ambleminae: Lampsilini), Fusconaia cerina (Ambleminae: Pleurobemini) and Strophitus connasaugaensis (Unioninae: Anodontini). Malacologia 2014, 57, 99–239. [Google Scholar] [CrossRef]
- Carella, F.; Villari, G.; Maio, N.; De Vico, G. Disease and Disorders of Freshwater Unionid Mussels: A Brief Overview of Recent Studies. Front. Physiol. 2016, 7, 489. [Google Scholar] [CrossRef]
- Brahma, N.; Gupta, A. Effects of Copper and Lead on Freshwater Mollusc Corbicula striatella (Deshayes, 1854). J. Energy Res. Environ. Technol. 2019, 6, 205–208. [Google Scholar]
- Hossain, M.A.; Sutradhar, L.; Sarker, T.R.; Saha, S.; Iqbal, M.M. Toxic Effects of Chlorpyrifos on the Growth, Hematology, and Different Organs Histopathology of Nile Tilapia, Oreochromis niloticus. Saudi J. Biol. Sci. 2022, 29, 103316. [Google Scholar] [CrossRef]
- Gilroy, È.A.M.; Gillis, P.L.; King, L.E.; Bendo, N.A.; Salerno, J.; Giacomin, M.; de Solla, S.R. The Effects of Pharmaceuticals on a Unionid Mussel (Lampsilis siliquoidea): An Examination of Acute and Chronic Endpoints of Toxicity across Life Stages. Environ. Toxicol. Chem. 2017, 36, 1572–1583. [Google Scholar] [CrossRef]
- Milam, C.D.; Farris, J.L.; Dwyer, F.J.; Hardesty, D.K. Acute Toxicity of Six Freshwater Mussel Species (Glochidia) to Six Chemicals: Implications for Daphnids and Utterbackia imbecillis as Surrogates for Protection of Freshwater Mussels (Unionidae). Arch. Environ. Contam. Toxicol. 2005, 48, 166–173. [Google Scholar] [CrossRef]
- Wang, N.; Ivey, C.D.; Ingersoll, C.G.; Brumbaugh, W.G.; Alvarez, D.; Hammer, E.J.; Bauer, C.R.; Augspurger, T.; Raimondo, S.; Barnhart, M.C. Acute Sensitivity of a Broad Range of Freshwater Mussels to Chemicals with Different Modes of Toxic Action. Environ. Toxicol. Chem. 2017, 36, 786–796. [Google Scholar] [CrossRef]
- Chapman, D.V.; Bradley, C.; Gettel, G.M.; Hatvani, I.G.; Hein, T.; Kovács, J.; Liska, I.; Oliver, D.M.; Tanos, P.; Trásy, B.; et al. Developments in Water Quality Monitoring and Management in Large River Catchments Using the Danube River as an Example. Environ. Sci. Policy 2016, 64, 141–154. [Google Scholar] [CrossRef]
- Tedengren, M.; Olsson, B.; Bradley, B.; Zhou, L. Heavy Metal Uptake, Physiological Response and Survival of the Blue Mussel (Mytilus edulis) from Marine and Brackish Waters in Relation to the Induction of Heat-Shock Protein 70. Hydrobiologia 1999, 393, 261–269. [Google Scholar] [CrossRef]
- Wang, N.; Ingersoll, C.G.; Greer, I.E.; Hardesty, D.K.; Ivey, C.D.; Kunz, J.L.; Brumbaugh, W.G.; Dwyer, F.J.; Roberts, A.D.; Augspurger, T.; et al. Chronic Toxicity of Copper and Ammonia to Juvenile Freshwater Mussels (Unionidae). Environ. Toxicol. Chem. 2007, 26, 2048–2056. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Lin, D.; Shang, Y.; Hu, Y.; Lu, W.; Huang, X.; Ning, K.; Chen, Y.; Wang, Y. CO2-Induced PH Reduction Increases Physiological Toxicity of Nano-TiO2 in the Mussel Mytilus coruscus. Sci. Rep. 2017, 7, 40015. [Google Scholar] [CrossRef]
- Álvarez-González, C.A.; Martínez-Sánchez, L.; Peña-Marín, E.S.; Guerrero-Zárate, R.; Jesús-Ramírez, F.; Morales-García, V.; Uribe-López, M.; Núñez-Nogueira, G. Effects on the Growth and Digestive Enzyme Activity in Nile Tilapia Fry (Oreochromis niloticus) by Lead Exposure. Water. Air. Soil Pollut. 2020, 231, 1–15. [Google Scholar] [CrossRef]
- Jorge, M.B.; Loro, V.L.; Bianchini, A.; Wood, C.M.; Gillis, P.L. Mortality, Bioaccumulation and Physiological Responses in Juvenile Freshwater Mussels (Lampsilis siliquoidea) Chronically Exposed to Copper. Aquat. Toxicol. 2013, 126, 137–147. [Google Scholar] [CrossRef]
- Paulino, A.T.; Santos, L.B.; Nozaki, J. Protective Action of Zinc against Lead Poisoning in Tilapia Oreochromis niloticus. Toxicol. Environ. Chem. 2007, 89, 363–370. [Google Scholar] [CrossRef]
- Ballina, N.R.; Maresca, F.; Cao, A.; Villalba, A. Bivalve Haemocyte Subpopulations: A Review. Front. Immunol. 2022, 13, 826255. [Google Scholar] [CrossRef]
- Hossain, M.A.; Rani Sarker, T.; Sutradhar, L.; Hussain, M.; Mahbub Iqbal, M. Toxic Effects of Chlorpyrifos on the Growth, Hemocytes Counts, and Vital Organ’s Histopathology of Freshwater Mussel, Lamellidens marginalis. J. King Saud Univ. Sci. 2022, 29, 102482. [Google Scholar] [CrossRef]
- Coles, J.; Farley, S.; Pipe, R. Alteration of the Immune Response of the Common Marine Mussel Mytilus edulis Resulting from Exposure to Cadmium. Dis. Aquat. Organ. 1995, 22, 59–65. [Google Scholar] [CrossRef]
- Evariste, L.; Rioult, D.; Brousseau, P.; Geffard, A.; David, E.; Auffret, M.; Fournier, M.; Betoulle, S. Differential Sensitivity to Cadmium of Immunomarkers Measured in Hemocyte Subpopulations of Zebra Mussel Dreissena polymorpha. Ecotoxicol. Environ. Saf. 2017, 137, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, A.; Fortier, M.; Masson, S.; Auffret, M.; Brousseau, P.; Fournier, M. Effect of Temperature on Immunocompetence of the Blue Mussel (Mytilus edulis). J. Xenobiotics 2016, 6, 5889. [Google Scholar] [CrossRef]
- Alonso, A.; Suárez, P.; Ruiz, Y.; Dobal, V.; San Juan, F. Gonadal Histopathological Disorders in Mytilus Galloprovincialis Male Exposed to Tars Used in Mussel Farms. Front. Mar. Sci. 2019, 6, 577. [Google Scholar] [CrossRef]
- Costa, P.M.; Carreira, S.; Costa, M.H.; Caeiro, S. Development of Histopathological Indices in a Commercial Marine Bivalve (Ruditapes decussatus) to Determine Environmental Quality. Aquat. Toxicol. 2013, 126, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J.; Costa, M.; Leite, C.; Borges, C.; Coppola, F.; Henriques, B.; Monteiro, R.; Russo, T.; Di Cosmo, A.; Soares, A.M.V.M.; et al. Ecotoxicological Effects of Lanthanum in Mytilus galloprovincialis: Biochemical and Histopathological Impacts. Aquat. Toxicol. 2019, 211, 181–192. [Google Scholar] [CrossRef]
- Mona, M.H.; El-Khodary, G.M.; Abdel-Halim, K.Y.; Omran, N.E.; Abd El-Aziz, K.K.; El-Saidy, S.A. Histopathological Alterations Induced by Marine Environmental Pollutants on the Bivalve Cerastoderma glaucum (Bruguière 1789) from Temsah Lake, Suez Canal, Egypt. Environ. Sci. Pollut. Res. 2022, 29, 9971–9989. [Google Scholar] [CrossRef]
- Knowles, G.; Handlinger, J.; Jones, B.; Moltschaniwskyj, N. Hemolymph Chemistry and Histopathological Changes in Pacific Oysters (Crassostrea gigas) in Response to Low Salinity Stress. J. Invertebr. Pathol. 2014, 121, 78–84. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration of Pb(NO₃)₂ (mgL−1) | Stocking Density (No./Treatment) | Replication |
|---|---|---|---|
| C (Control) | 00.00 (00% of LC50) | 20 | 3 |
| T1 | 21.93 (10% of LC50) | 20 | 3 |
| T2 | 43.86 (20% of LC50) | 20 | 3 |
| T3 | 87.72 (40% of LC50) | 20 | 3 |
| Species | Value of 96 h LC50 (mgL−1) | References | |
|---|---|---|---|
| Bivalves | Lamellidense marginalis | 219.32 | Current study |
| L. jenkinsianus | 192.14 | [28] | |
| Corbicula striatella | 231.01 | [48] | |
| Gastropods | Theodoxus niloticus | 18 | [5] |
| Melanoides tuberculata | 6.82 | [17] | |
| Cerithedia cingulate | 15.50 | [37] | |
| Bivalves | Modiolus philippinarum | 2.86 |
| Treatment | Day 0 | Day 15 | Day 40 | |
|---|---|---|---|---|
| Temperature | C | 20.73 ± 0.23 | 19.56 ± 0.37 | 20.66 ± 0.50 |
| T1 | 20.93 ± 0.11 | 20.23 ± 0.41 | 20.66 ± 0.20 | |
| T2 | 21.00 ± 0.17 | 19.70 ± 0.17 | 20.76 ± 0.15 | |
| T3 | 21.03 ± 0.28 | 20.13 ± 0.49 | 20.80 ± 0.10 | |
| pH | C | 7.96 ± 0.05 | 8.20 ± 0.10 | 8.23 ± 0.10 |
| T1 | 7.83 ± 0.05 | 8.19 ± 0.10 | 7.96 ± 0.05 | |
| T2 | 7.70 ± 0.03 | 8.16 ± 0.05 | 7.76 ± 0.05 | |
| T3 | 7.60 ± 0.01 | 8.06 ± 0.05 | 7.53 ± 0.05 | |
| Salinity | C | 0.07 ± 0 | 0.08 ± 0 | 0.10 ± 0 |
| T1 | 0.07 ± 0 | 0.08 ± 0 | 0.11 ± 0 | |
| T2 | 0.07 ± 0 | 0.08 ± 0 | 0.11 ± 0 | |
| T3 | 0.07 ± 0 | 0.08 ± 0 | 0.09 ± 0 | |
| NH3 | C | 0.05 ± 0 | 0.07 ± 0 | 0.07 ± 0 |
| T1 | 0.01 ± 0 | 0.02 ± 0 | 0.02 ± 0 | |
| T2 | 0.02 ± 0 | 0.02 ± 0 | 0.02 ± 0 | |
| T3 | 0.02 ± 0 | 0.01 ± 0 | 0.02 ± 0 | |
| DO | C | 7.26 ± 0.49 | 7.48 ± 0.47 | 7.39 ± 0.63 |
| T1 | 7.58 ± 0.29 | 7.58 ± 0.29 | 7.60 ± 0.48 | |
| T2 | 7.54 ± 0.075 | 7.54 ± 0.07 | 7.43 ± 0.08 | |
| T3 | 7.64 ± 0.104 | 7.64 ± 0.10 | 7.77 ± 0.08 |
| C | T1 | T2 | T3 | |
|---|---|---|---|---|
| Shell Length Initial (cm) | 7.23 ± 0.16 | 7.16 ± 0.12 | 7.26 ± 0.17 | 7.22 ± 0.20 |
| Shell Weight Initial (g) | 41.40 ± 1.8 | 41.28 ± 2.01 | 41.06 ± 2.59 | 41.16 ± 2.52 |
| Shell Depth Initial (cm) | 4.06 ± 0.11 | 4.02 ± 0.09 | 4.09 ± 0.06 | 4.01 ± 0.10 |
| Shell Length Final (cm) | 7.33 ± 0.09 | 7.28 ± 0.10 | 7.31 ± 0.10 | 7.28 ± 0.13 |
| Shell Weight Final(g) | 50.32 ± 2.58 b | 40.69 ± 1.58 a | 41.41 ± 1.58 a | 38.67 ± 2.24 a |
| Shell Depth Final (cm) | 4.32 ± 0.05 | 4.25 ± 0.05 | 4.36 ± 0.04 | 4.36 ± 0.05 |
| Soft Tissue Wet Weight (g) | 9.41 ± 0.64 b | 7.88 ± 0.26 a | 7.85 ± 0.31 a | 7.01 ± 0.42 a |
| Dry Shell Weight (g) | 16.45 ± 0.98 | 15.05 ± 0.85 | 16.23 ± 1.07 | 15.09 ± 1.50 |
| Length Gain (cm) | 0.10 ± 0.21 | 0.12 ± 0.13 | 0.05 ± 0.20 | 0.06 ± 0.25 |
| Weight Gain (g) | 8.92 ± 3.52 b | −0.59 ± 2.65 ab | 0.35 ± 3.29 ab | −2.49 ± 3.95 a |
| Specific Growth Rate % | 0.48 ± 0.18 b | −0.01 ± 0.17 a | 0.07 ± 0.19 a | −0.13 ± 0.25 a |
| Average Daily Length Gain | 0.002 ± 0.05 | 0.003 ± 0.003 | 0.001 ± 0.005 | 0.001 ± 0.006 |
| Average Daily Weight Gain | 0.22 ± 0.08 b | −0.01 ± 0.06 ab | 0.01 ± 0.08 ab | −0.06 ± 0.09 a |
| Organ | Abnormality | Control | T1 | T2 | T3 |
|---|---|---|---|---|---|
| Gill | Tissue Rupture | --- | * | * | ** |
| Extended Vessel | --- | ** | ** | *** | |
| A Massive Expansion of The Vessel | --- | * | --- | ** | |
| Epithelial Deformities | --- | --- | * | * | |
| Filament Congestion | --- | * | --- | * | |
| Cilia Damage | --- | * | ** | *** | |
| Epithelial Damage | --- | --- | * | *** | |
| Filament Attachment And Abnormalities | --- | --- | --- | * | |
| Kidney | Enormous Granules | --- | * | ** | ** |
| Extended Lumen | --- | --- | * | ** | |
| Damage to Connective Tissue | --- | --- | --- | * | |
| Vacuoles Due to Massive Tissue Rupture | --- | --- | * | ** | |
| Enlarged Brown Intracellular Granules | --- | --- | * | *** | |
| Necrosis | --- | ** | ** | ** | |
| Tissue Rupture | --- | --- | ** | ** | |
| Epithelial Lifting | --- | --- | * | --- | |
| Muscle | Cell Rupture | --- | * | *** | ** |
| Massive Vacuoles Due to Cell Rupture | --- | --- | ** | * | |
| Necrosis | --- | * | * | ** | |
| Fibrous Tissue Damage | --- | --- | * | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.A.; Chowdhury, T.; Chowdhury, G.; Schneider, P.; Hussain, M.; Das, B.; Iqbal, M.M. Impact of Pb Toxicity on the Freshwater Pearl Mussel, Lamellidens marginalis: Growth Metrics, Hemocyto-Immunology, and Histological Alterations in Gill, Kidney, and Muscle Tissue. Toxics 2023, 11, 475. https://doi.org/10.3390/toxics11060475
Hossain MA, Chowdhury T, Chowdhury G, Schneider P, Hussain M, Das B, Iqbal MM. Impact of Pb Toxicity on the Freshwater Pearl Mussel, Lamellidens marginalis: Growth Metrics, Hemocyto-Immunology, and Histological Alterations in Gill, Kidney, and Muscle Tissue. Toxics. 2023; 11(6):475. https://doi.org/10.3390/toxics11060475
Chicago/Turabian StyleHossain, Mohammad Amzad, Toma Chowdhury, Gourab Chowdhury, Petra Schneider, Monayem Hussain, Bipresh Das, and Mohammed Mahbub Iqbal. 2023. "Impact of Pb Toxicity on the Freshwater Pearl Mussel, Lamellidens marginalis: Growth Metrics, Hemocyto-Immunology, and Histological Alterations in Gill, Kidney, and Muscle Tissue" Toxics 11, no. 6: 475. https://doi.org/10.3390/toxics11060475