

The Role of G3BP1 Gene Mediates P38 MAPK/JNK Pathway in Testicular Spermatogenic Dysfunction Caused by Cyfluthrin
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Experimental Animal
2.3. Hematoxylin-Eosin Staining and Ultrastructural Observation of Testicular Tissue
2.4. Homogenization of Testicular Tissue
2.5. Determination of MDA Content
2.6. Determination of T-AOC
2.7. The Determination of FSH, T, LH and GnRH by Elisa
2.8. Quantitative Real-Time PCR (qPCR)
2.9. Western Blot
2.10. Immunohistochemical Analysis (IHC)
2.11. Double-Label Immunofluorescence (Double-IF)
2.12. Statistical Methods
3. Results
3.1. The Effect of Cyfluthrin on Body Weight and Testicular Organ Coefficient in Rats
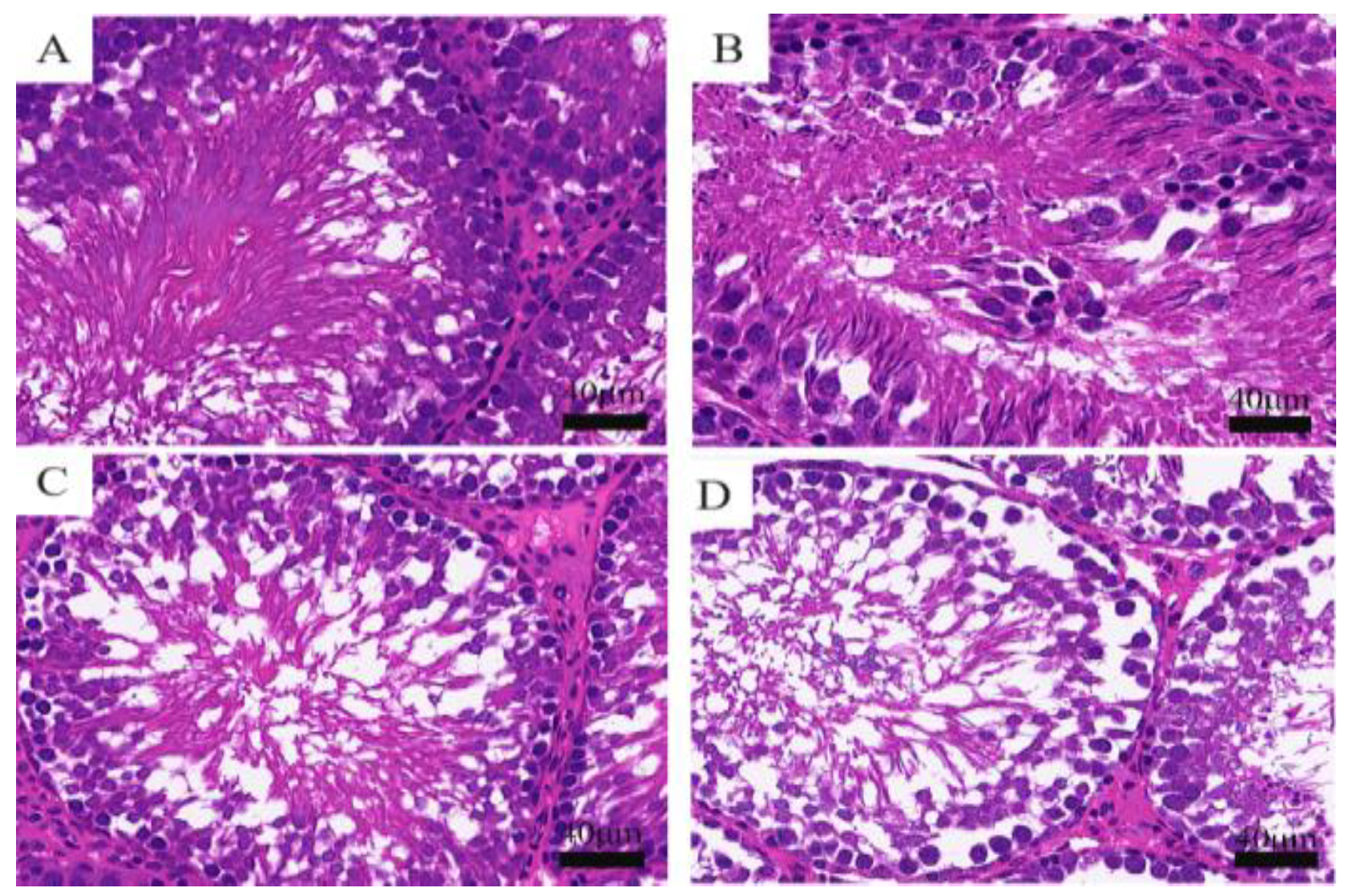
3.2. Cyfluthrin Could Cause Histopathological Changes in Testis and Ultrastructural Damage in Spermatocytes
3.2.1. Histopathological Changes of Rat Testis Observed by HE Staining
3.2.2. Cyfluthrin Could Cause Ultrastructural Damage in Spermatocytes
3.3. Cyfluthrin Could Interfere with the Normal Secretion of the Hypothalamic–Pituitary–Testicular Axis, Causing Hypergonadotropic Dysfunction
3.4. Cyfluthrin Could Cause the Oxidation–Oxidation Steady-State Imbalance and Inhibit G3BP1 Expressionin in Testicular Tissue
3.5. Determination of MAPK Pathway Key Factors Expression in Testicular Tissues by Western Blot and q-PCR
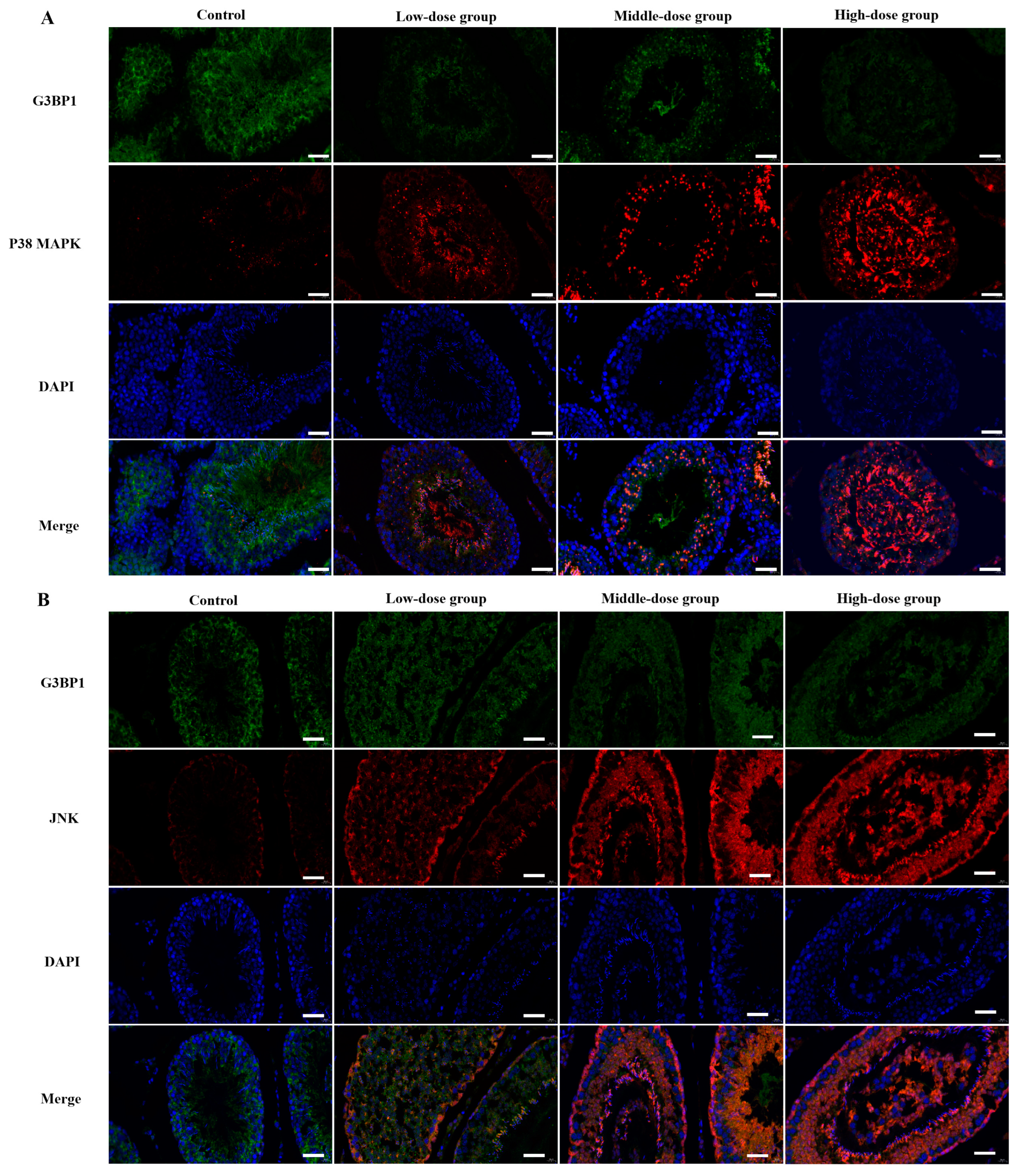
3.6. The Determination of Co-Located Situation of JNK/G3BP1, P38 MAPK/G3BP1 by Doudle-IF
3.7. Western Blot and q-PCR to Detect the Expression of Cytochrome c Oxidase 1 and 4 in Testicular Tissues
3.8. Western Blot and q-PCR to Detect Caspases 8/9/3 in Testicular Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Acr | acrosome |
| Cy | Cyfluthrin |
| EEDs | environmental endocrine disruptors |
| FSH | follicle-stimulating hormone |
| G3BP1 | Gtpase-activated protein SH3 domain-binding protein 1 |
| GnRH | gonadotropin-releasing hormone |
| LH | luteinizing hormone |
| LD | lipid droplets |
| M | mitochondria |
| N | nuclear |
| RER | rough endoplasmic reticulum |
| T | testosterone |
| TEM | transmission electron microscope |
References
- Tijani, J.O.; Fatoba, O.O.; Babajide, O.O.; Petrik, L.F. Pharmaceuticals, endocrine disruptors, personal care products, nanomaterials and perfluorinated pollutants: A review. Environ. Chem. Lett. 2016, 14, 27–49. [Google Scholar] [CrossRef]
- Syed, F.; Soni, I.; John, P.; Bhatnagar, P. Embryotoxic and teratogenic evaluation of cyfluthrin in Swiss albino mice. Toxicol. Ind. Health 2018, 16, 121–126. [Google Scholar] [CrossRef]
- Hedges, L.; Brown, S.; MacLeod, A.K.; Moreau, M.; Yoon, M.; Creek, M.R.; Osimitz, T.G.; Lake, B.G. Metabolism of bifenthrin, β-cyfluthrin, λ-cyhalothrin, cyphenothrin and esfenvalerate by rat and human cytochrome P450 and carboxylesterase enzymes. Xenobiotica 2020, 50, 1434–1442. [Google Scholar] [CrossRef]
- Shi, Y.; Qi, W.; Xu, Q.; Wang, Z.; Cao, X.; Zhou, L.; Ye, L. The role of epigenetics in the reproductive toxicity of environmental endocrine disruptors. Environ. Mol. Mutagen. 2021, 62, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, J.; Imai, K.; Shiraishi, H.; Nozawa, S.; Yoshiike, M.; Mieno, M.N.; Andersson, A.-M.; Iwamoto, T. Pyrethroid insecticide exposure and reproductive hormone levels in healthy Japanese male subjects. Andrology 2014, 2, 416–420. [Google Scholar] [CrossRef]
- Patrick, S.M.; Bornman, M.S.; Joubert, A.M.; Pitts, N.; Naidoo, V.; de Jager, C. Effects of environmental endocrine disruptors, including insecticides used for malaria vector control on reproductive parameters of male rats. Reprod. Toxicol. 2016, 61, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Pauluhn, J. Upper respiratory tract nociceptor stimulation and stress response following acute and repeated Cyfluthrin inhalation in normal and pregnant rats: Physiological rat-specific adaptions can easily be misunderstood as adversities. Toxicol. Lett. 2018, 282, 8–24. [Google Scholar] [CrossRef]
- Ravula, A.R.; Yenugu, S. Transgenerational effects on the fecundity and sperm proteome in rats exposed to a mixture of pyrethroids at doses similar to human consumption. Chemosphere 2022, 290, 133242. [Google Scholar] [CrossRef]
- Ila, H.B.; Topaktas, M.; Rencuzogullari, E.; Kayraldiz, A.; Donbak, L.; Daglioglu, Y.K. Genotoxic potential of cyfluthrin. Mutat. Res. Toxicol. Environ. Mutagen. 2008, 656, 49–54. [Google Scholar] [CrossRef]
- Savelli, C.J.; Mateus, C. A mixed-method exploration into the experience of members of the FAO/WHO International Food Safety Authorities Network (INFOSAN): Study protocol. BMJ Open 2019, 9, e027091. [Google Scholar] [CrossRef]
- Green, M.P.; Harvey, A.J.; Finger, B.J.; Tarulli, G.A. Endocrine disrupting chemicals: Impacts on human fertility and fecundity during the peri-conception period. Environ. Res. 2021, 194, 110694. [Google Scholar] [CrossRef]
- Cuenca, J.B.; Tirado, N.; Barral, J.; Ali, I.; Levi, M.; Stenius, U.; Berglund, M.; Dreij, K. Increased levels of genotoxic damage in a Bolivian agricultural population exposed to mixtures of pesticides. Sci. Total. Environ. 2019, 695, 133942. [Google Scholar] [CrossRef]
- Wu, B. The Current Situation of Pesticide Residues in Vegetable Greenhouses in the Suburbs of Yinchuan City and Analysis of Influencing Factors on Cardiovascular Health of Planters; Ningxia Medical University: Yinchuan, China, 2016. [Google Scholar]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef]
- Martínez, M.-A.; Lopez-Torres, B.; Rodríguez, J.-L.; Martínez, M.; Maximiliano, J.-E.; Martínez-Larrañaga, M.-R.; Anadón, A.; Ares, I. Toxicologic evidence of developmental neurotoxicity of Type II pyrethroids cyfluthrin and alpha-cypermethrin in SH-SY5Y cells. Food Chem. Toxicol. 2020, 137, 111173. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Ares, I.; Ramos, E.; Castellano, V.; Martínez, M.; Martínez-Larrañaga, M.; Anadón, A. Evidence for dose-additive effects of a type II pyrethroid mixture. in vitro assessment. Environ. Res. 2015, 138, 58–66. [Google Scholar] [CrossRef]
- Lu, Y.-C.; Liang, W.-Z.; Kuo, C.-C.; Hao, L.-J.; Chou, C.-T.; Jan, C.-R. Action of the insecticide cyfluthrin on Ca2+ signal transduction and cytotoxicity in human osteosarcoma cells. Hum. Exp. Toxicol. 2020, 39, 1268–1276. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, L.; Zhang, L.; Zhang, W.; Deng, Y.; Liu, R.; Qin, Y.; Zhou, Z.; Diao, J. Thermal effects on tissue distribution, liver biotransformation, metabolism and toxic responses in Mongolia racerunner (Eremias argus) after oral administration of beta-cyfluthrin. Environ. Res. 2020, 185, 109393. [Google Scholar] [CrossRef]
- Li, A.J.; Martinez-Moral, M.-P.; Kannan, K. Temporal variability in urinary pesticide concentrations in repeated-spot and first-morning-void samples and its association with oxidative stress in healthy individuals. Environ. Int. 2019, 130, 104904. [Google Scholar] [CrossRef]
- Ye, X.; Li, F.; Zhang, J.; Ma, H.; Ji, D.; Huang, X.; Curry, T.E., Jr.; Liu, W.; Liu, J. Pyrethroid insecticide cypermethrin accelerates pubertal onset in male mice via disrupting hypothalamic-pituitary-gonadal axis. Environ. Sci. Technol. 2017, 51, 10212–10221. [Google Scholar] [CrossRef]
- Omer, A.; Barrera, M.C.; Moran, J.L.; Lian, X.J.; Di Marco, S.; Beausejour, C.; Gallouzi, I.-E. G3BP1 controls the senescence-associated secretome and its impact on cancer progression. Nat. Commun. 2020, 11, 4979. [Google Scholar] [CrossRef]
- Sharma, A.; Minhas, S.; Dhillo, W.S.; Jayasena, C.N. Male infertility due to testicular disorders. J. Clin. Endocrinol. Metab. 2021, 106, e442–e459. [Google Scholar] [CrossRef] [PubMed]
- Almog, T.; Naor, Z. The role of Mitogen activated protein kinase (MAPK) in sperm functions. Mol. Cell. Endocrinol. 2010, 314, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; He, Z.; Yu, C.; Guan, Q. Role of p38 MAPK Signalling in Testis Development and Male Fertility. Oxidative Med. Cell. Longev. 2022, 2022, 6891897. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ma, L.; Xie, Y.; Zhu, L.; Ni, W.; Wang, R.; Song, Y.; Li, X.; Yang, H. The role of stimulator of interferon genes-mediated AMPK/mTOR/P70S6K autophagy pathway in cyfluthrin-induced testicular injury. Environ. Toxicol. 2023, 38, 727–742. [Google Scholar] [CrossRef] [PubMed]
- Saillenfait, A.-M.; Ndiaye, D.; Sabaté, J.-P. The estrogenic and androgenic potential of pyrethroids in vitro. Review. Toxicol. Vitro 2016, 34, 321–332. [Google Scholar] [CrossRef]
- Maksoud, H.A.; Mahfouz, M.; Soliman, M.; Elharrif, M.G.; Abbass, M.; El-Badry, M. Harmful effects of pyrethroid ester insecticide on the male reproductive system mainly through affecting testicular function and inflammatory markers. BIOCELL 2020, 44, 111–115. [Google Scholar] [CrossRef]
- Gupta, A.S.; Deshpande, P.S. Causes and prevalence of factors causing infertility in a public health facility. J. Hum. Reprod. Sci. 2019, 12, 287–293. [Google Scholar] [CrossRef]
- Plunk, E.C.; Richards, S.M. Endocrine-disrupting air pollutants and their effects on the hypothalamus-pituitary-gonadal axis. Int. J. Mol. Sci. 2020, 21, 9191. [Google Scholar] [CrossRef]
- Salonia, A.; Rastrelli, G.; Hackett, G.; Seminara, S.B.; Huhtaniemi, I.T.; Rey, R.A.; Hellstrom, W.J.G.; Palmert, M.R.; Corona, G.; Dohle, G.R.; et al. Paediatric and adult-onset male hypogonadism. Nat. Rev. Dis. Prim. 2019, 5, 38. [Google Scholar] [CrossRef]
- Hamzeh, M.; Robaire, B. Androgens activate mitogen-activated protein kinase via epidermal growth factor receptor/insulin-like growth factor 1 receptor in the mouse PC-1 cell line. J. Endocrinol. 2019, 209, 55–64. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, T.; Ren, X.; Chen, X.; Wang, S.; Qin, C. Pyrethroids toxicity to male reproductive system and offspring as a function of oxidative stress induction: Rodent studies. Front. Endocrinol. 2021, 12, 656106. [Google Scholar] [CrossRef]
- Herman, A.B.; Afonso, M.; Kelemen, S.E.; Ray, M.; Vrakas, C.N.; Burke, A.C.; Scalia, R.G.; Moore, K.; Autieri, M.V. Regulation of stress granule formation by inflammation, vascular injury, and atherosclerosis. Arter. Thromb. Vasc. Biol. 2019, 39, 2014–2027. [Google Scholar] [CrossRef]
- Albornoz, A.; Carletti, T.; Corazza, G.; Marcello, A. The stress granule component TIA-1 binds tick-borne encephalitis virus RNA and is recruited to perinuclear sites of viral replication to inhibit viral translation. J. Virol. 2014, 88, 6611–6622. [Google Scholar] [CrossRef]
- Lee, A.K.; Klein, J.; Tacer, K.F.; Lord, T.; Oatley, M.J.; Oatley, J.M.; Porter, S.N.; Pruett-Miller, S.M.; Tikhonova, E.B.; Karamyshev, A.L.; et al. Translational repression of G3BP in cancer and germ cells suppresses stress granules and enhances stress tolerance. Mol. Cell 2020, 79, 645–659.e9. [Google Scholar] [CrossRef]
- Somasekharan, S.P.; Zhang, F.; Saxena, N.; Ni Huang, J.; Kuo, I.-C.; Low, C.; Bell, R.; Adomat, H.; Stoynov, N.; Foster, L.; et al. G3BP1-linked mRNA partitioning supports selective protein synthesis in response to oxidative stress. Nucleic Acids Res. 2020, 48, 6855–6873. [Google Scholar] [CrossRef]
- Fontana, F.; Marzagalli, M.; Marelli, M.M.; Raimondi, M.; Moretti, R.M.; Limonta, P. Gonadotropin-releasing hormone receptors in prostate cancer: Molecular aspects and biological functions. Int. J. Mol. Sci. 2020, 21, 9511. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, W.; Zhang, J.; Li, H. Role of JNK and ERK1/2 MAPK signaling pathway in testicular injury of rats induced by di-N-butyl-phthalate (DBP). Biol. Res. 2019, 52, 41. [Google Scholar] [CrossRef]
- Azad, F.; Nejati, V.; Shalizar-Jalali, A.; Najafi, G.; Rahmani, F. Antioxidant and anti-apoptotic effects of royal jelly against nicotine-induced testicular injury in mice. Environ. Toxicol. 2019, 34, 708–718. [Google Scholar] [CrossRef]
- Barati, E.; Nikzad, H.; Karimian, M. Oxidative stress and male infertility: Current knowledge of pathophysiology and role of antioxidant therapy in disease management. Cell. Mol. Life Sci. 2020, 77, 93–113. [Google Scholar] [CrossRef]
- Cho, E.; Than, T.T.; Kim, S.-H.; Park, E.-R.; Kim, M.-Y.; Lee, K.H.; Shin, H.J. G3BP1 depletion increases radiosensitisation by inducing oxidative stress in response to DNA damage. Anticancer Res. 2019, 39, 6087–6095. [Google Scholar] [CrossRef]
- Kuribayashi, K.; Mayes, P.A.; El-Deiry, W.S. What are Caspases 3 and 7 doing upstream of the mitochondria? Cancer Biol. Ther. 2016, 5, 763–765. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Before Exposure | Exposure for 14 Days | Exposure for 28 Days |
|---|---|---|---|
| Control group | 337.60 ± 16.34 | 418 ± 18.47 | 473.7 ± 21.22 |
| Low-dose group | 338.20 ± 8.07 | 420.6 ± 18.29 | 473.2 ± 21.96 |
| Middle-dose group | 335.20 ± 14.67 | 408.1 ± 21.17 | 432.8 ± 78.28 |
| High-dose group | 345.30 ± 10.77 | 419.3 ± 16.51 | 454.2 ± 18.44 |
| F | 1.755 | 0.9222 | 2.03 |
| p value | 0.1731 | 0.4399 | 0.127 |
| Groups | Testicular Organ Coefficient | F | p-Value |
|---|---|---|---|
| Control group | 0.74 ± 0.06 | 0.3841 | 0.6847 |
| Low-dose group | 0.84 ± 0.10 | ||
| Middle-dose group | 0.81 ± 0.08 | ||
| High-dose group | 0.73 ± 0.25 |
| Groups | MDA (nmol/mg Prot) | T-AOC (mmol/mg) |
|---|---|---|
| Control group | 59.77 ± 11.41 | 46.68 ± 8.44 |
| Low-dose group | 74.61 ± 24.21 * | 18.47 ± 5.59 * |
| Middle-dose group | 104.50 ± 10.61 * | 6.41 ± 2.14 * |
| High-dose group | 182.40 ± 12.21 ** | 1.55 ± 1.51 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-Y.; Sun, J.; Ma, L.-Y.; Xie, Y.-X.; Zhang, N.; Zhao, J.; Yang, H.-F. The Role of G3BP1 Gene Mediates P38 MAPK/JNK Pathway in Testicular Spermatogenic Dysfunction Caused by Cyfluthrin. Toxics 2023, 11, 451. https://doi.org/10.3390/toxics11050451
Li X-Y, Sun J, Ma L-Y, Xie Y-X, Zhang N, Zhao J, Yang H-F. The Role of G3BP1 Gene Mediates P38 MAPK/JNK Pathway in Testicular Spermatogenic Dysfunction Caused by Cyfluthrin. Toxics. 2023; 11(5):451. https://doi.org/10.3390/toxics11050451
Chicago/Turabian StyleLi, Xiao-Yu, Jian Sun, Li-Ya Ma, Yong-Xin Xie, Na Zhang, Ji Zhao, and Hui-Fang Yang. 2023. "The Role of G3BP1 Gene Mediates P38 MAPK/JNK Pathway in Testicular Spermatogenic Dysfunction Caused by Cyfluthrin" Toxics 11, no. 5: 451. https://doi.org/10.3390/toxics11050451