

Natural Polyphenols—Resveratrol, Quercetin, Magnolol, and β-Catechin—Block Certain Aspects of Heroin Addiction and Modulate Striatal IL-6 and TNF-α
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Groups
2.3. Drugs
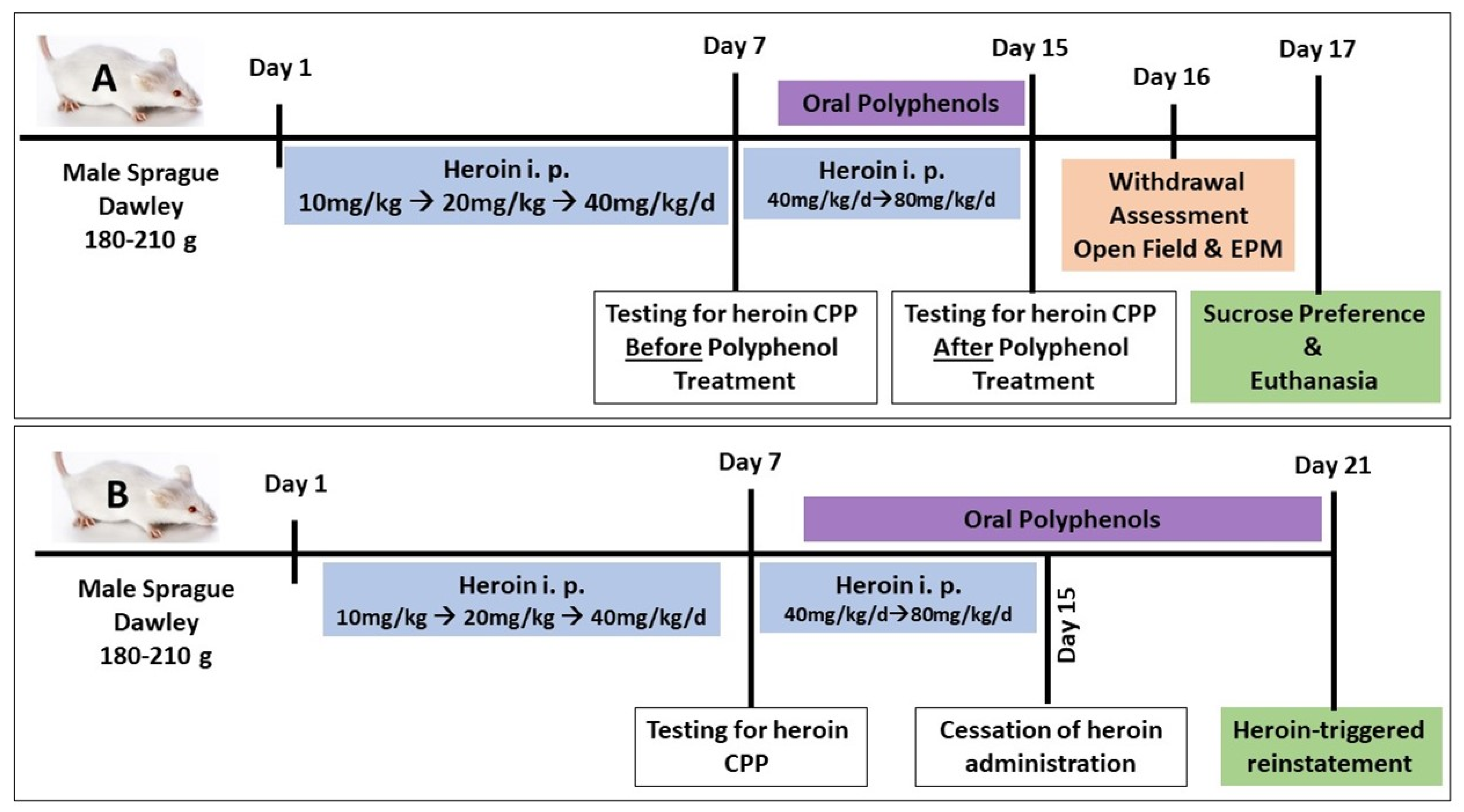
2.4. Behavioral Study
- (A)
- Conditioned place preference (first cohort)
- Habituation: Before the start of the procedure, the rats were habituated to the place preference laboratory room for one hour. During the preconditioning phase (1 day), the animals were allowed to freely explore the whole apparatus for 15 min. The time spent in each chamber, while the door is open, was recorded (unconditioned preference), and then the animals were returned to their home cages.
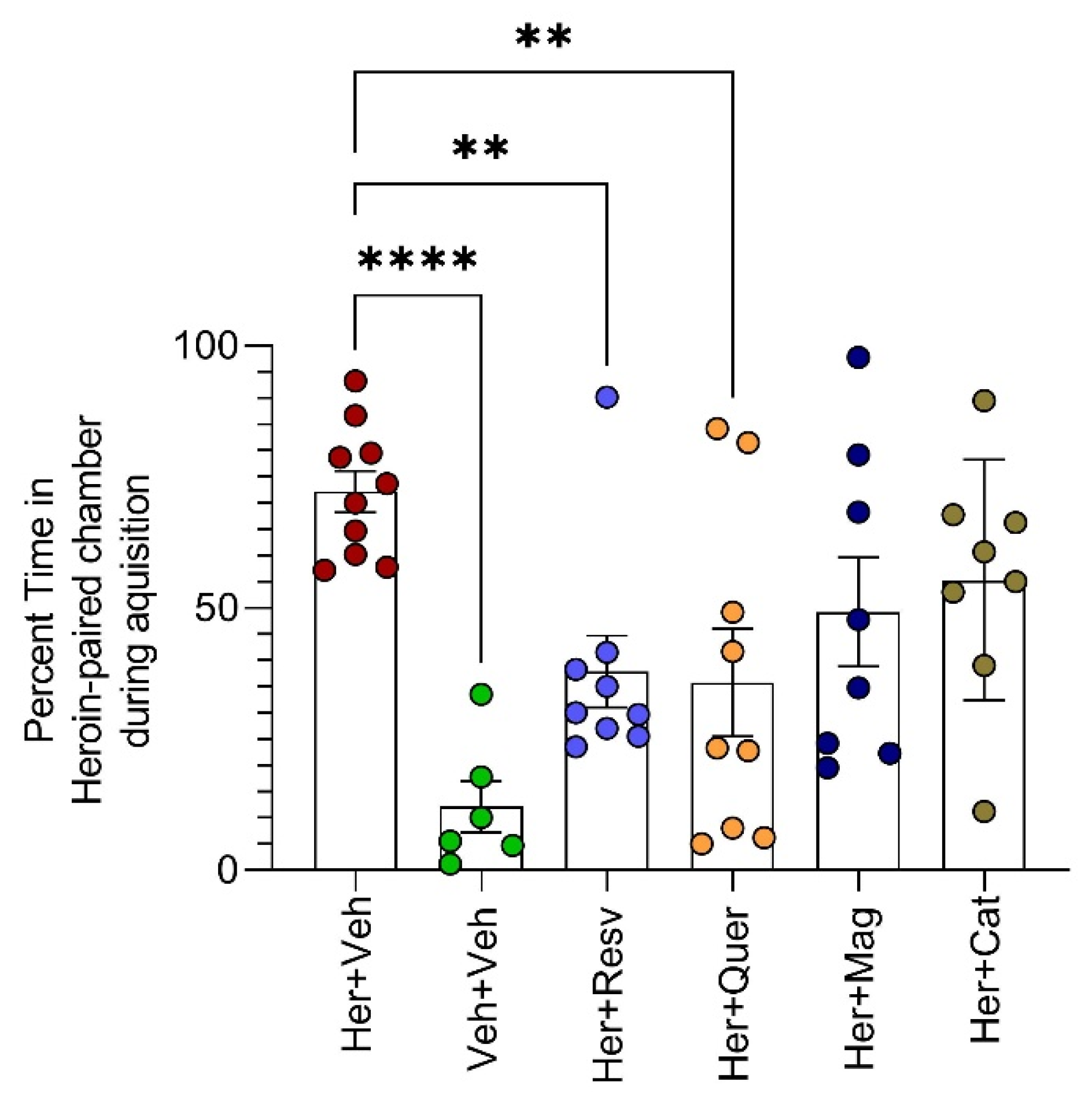
- Establishing CPP: The following day, control rats received saline (0.5 mL/kg, i.p.), while heroin-primed rats received heroin in the least preferred chamber in escalating doses, starting from 10 mg/kg, i.p. up to 80 mg/kg daily for 14 consecutive days (10 mg/kg/d for 4 days, then 20 mg/kg/d for 4 days, then 40 mg/kg/d for 4 days, then 80 mg/kg/d for 2 days). The rats received treatments for heroin CPP starting on day 8. Animals were administered polyphenols (quercetin 50 mg/kg/d [36], or β-catechin 100 mg/kg/d [37], or resveratrol 30 mg/kg/d [38], p.o., or magnolol 50 mg/kg/d, p.o. [39]) or distilled water (1 mL) through oral gavage for 7 consecutive days starting at day 8 of heroin administration.
- Testing for heroin CPP and testing the efficacy of polyphenols in blocking heroin CPP: On day 15, the animals were tested for heroin preference during 10 min of free access to both chambers. The percentage of time spent in the drug-paired chamber was recorded manually by a blinded observer in real time.
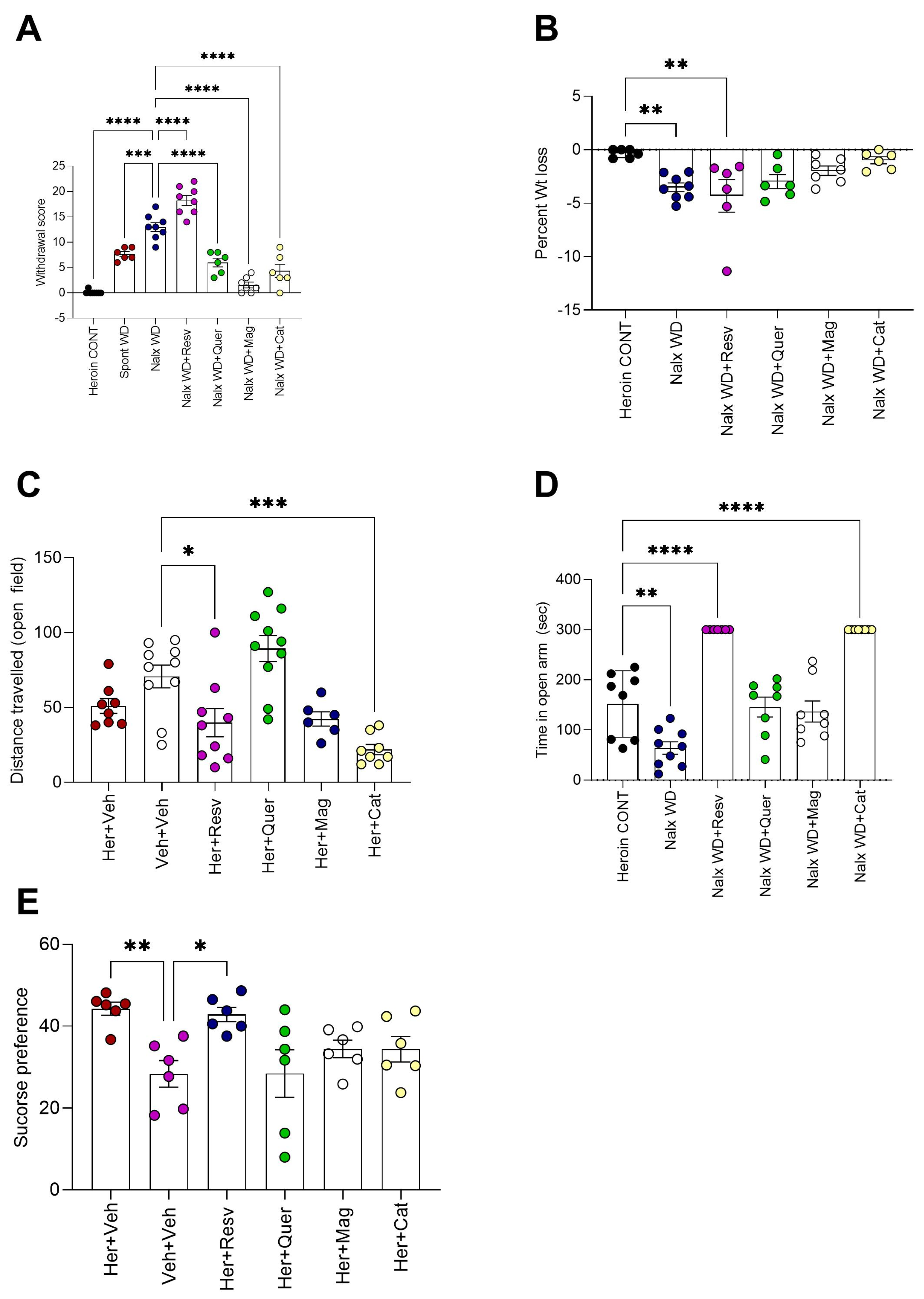
- Testing for the efficacy of polyphenols in attenuating naloxone-precipitated heroin withdrawal: On day 16 after testing for heroin CPP, rats were challenged with naloxone (1 mg/kg, i.p.) after 24 h of the last heroin dose (between 8:00 a.m. and 12:00 p.m.) to precipitate withdrawal, and they were observed in a transparent cylinder arena for a 30 min test period to detect withdrawal symptoms; scores were recorded manually in real time by a blinded observer [40]. Specific withdrawal signs, including jumping, wet dog shakes, head shakes, teeth chattering, tremors, and rearing movements, were counted during every 5 min observation. Irritation, piloerection, salivation, diarrhea, and grooming were observed and scored on a four-point scale: 0 = absent; 1 = mild; 2 = moderate; 3 = severe. The scores for each time period were combined [1].
- Testing for the effect of naloxone-precipitated withdrawal on spontaneous locomotor activity using an open field: On day 16, the rats were screened in an open field at the end of the experiment for 5 min (Fernandes et al., 2012). Each rat was placed in the center of the field (100 × 100 cm white box), and the number of squares crossed, as well as the vertical rears, were monitored by a blind observer in real time.
- Testing for the effect of naloxone-precipitated withdrawal on anxiety using elevated plus maze (EPM): Following the open field experiment, the rats were tested for anxiety using EPM. The maze was raised 40 cm off the floor with two equal crossed arms (10 cm wide and 100 cm long), and one arm was closed by 30 cm high walls. Animals were placed at the intersection facing one open arm and allowed to freely move; the time spent in the open or closed arm, in addition to the number of entries into any of the arms, was recorded in real time by a blinded observer. Maze sessions of 5 min each were held after 90 min of naloxone-induced withdrawal [41].
- Testing for the effect of naloxone-precipitated withdrawal on sucrose preference: On day 17 (next day of naloxone-precipitated withdrawal), the rats were deprived of food for 12 h, starting at 8:00 p.m. and continuing until 8:00 a.m. the next day, and were placed in individual cages and provided two regular 200 mL bottles: one containing 3% sucrose solution and the other containing tap water. On the next day (8:00 a.m.), the volumes of sucrose-containing water and plain water were recorded after 24 h. Sucrose intake was calculated: sucrose preference = sucrose intake/total intake (sucrose + water intake) × 100 (28).
- Euthanasia and brain tissue collection: The rats were euthanized by decapitation under light anesthesia after the end of the sucrose preference test on day 17. Brain tissues were dissected and stored at −80 °C for molecular assay (Figure 1A).
- (B)
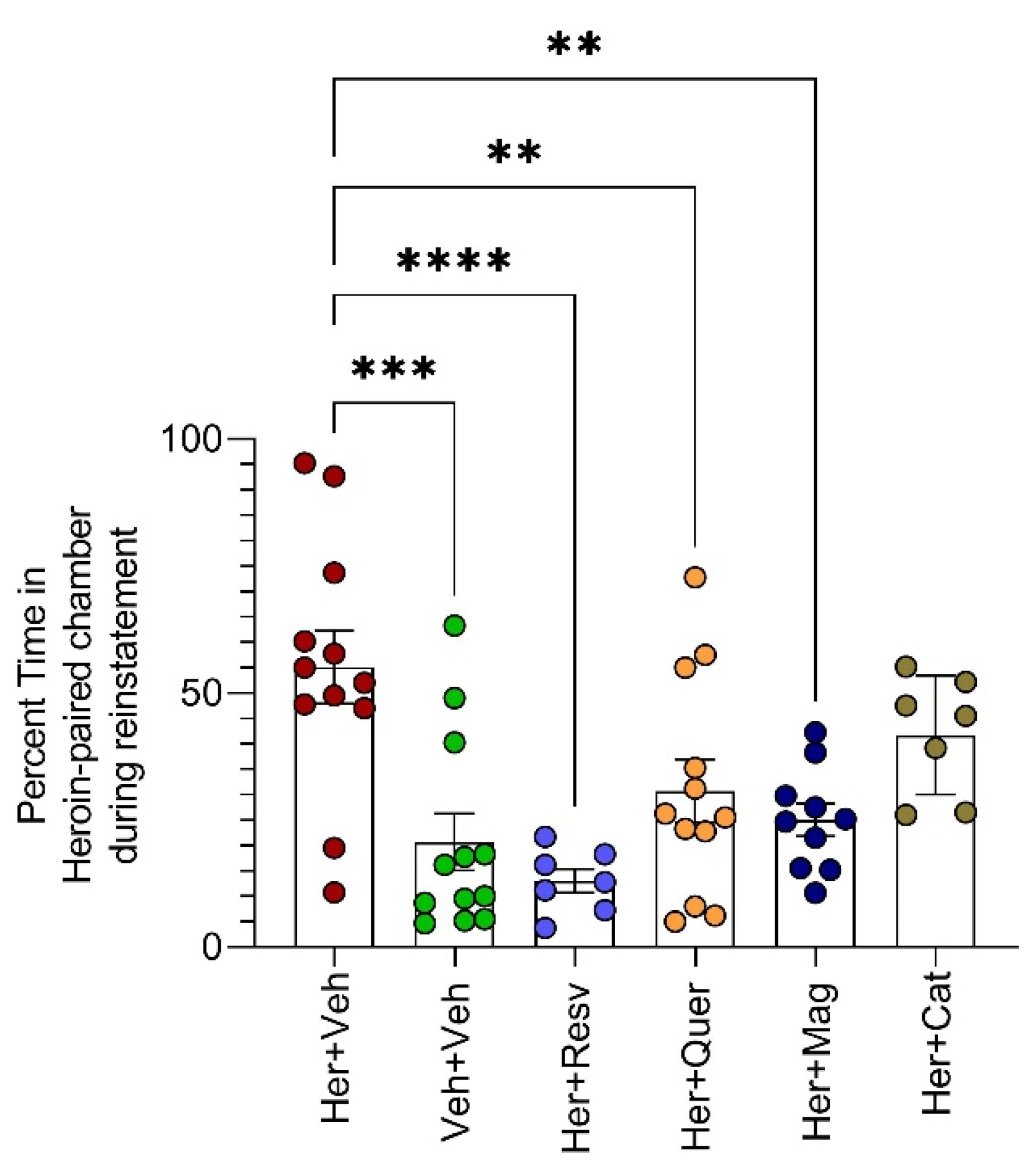
- Heroin-triggered-reinstatement (second cohort)
2.5. Striatal TNF-α and IL-6 Assay
2.6. Statistical Analysis
3. Results
3.1. Behavioral Effects
3.2. Resveratrol Worsens Naloxone-Induced Heroin Withdrawal
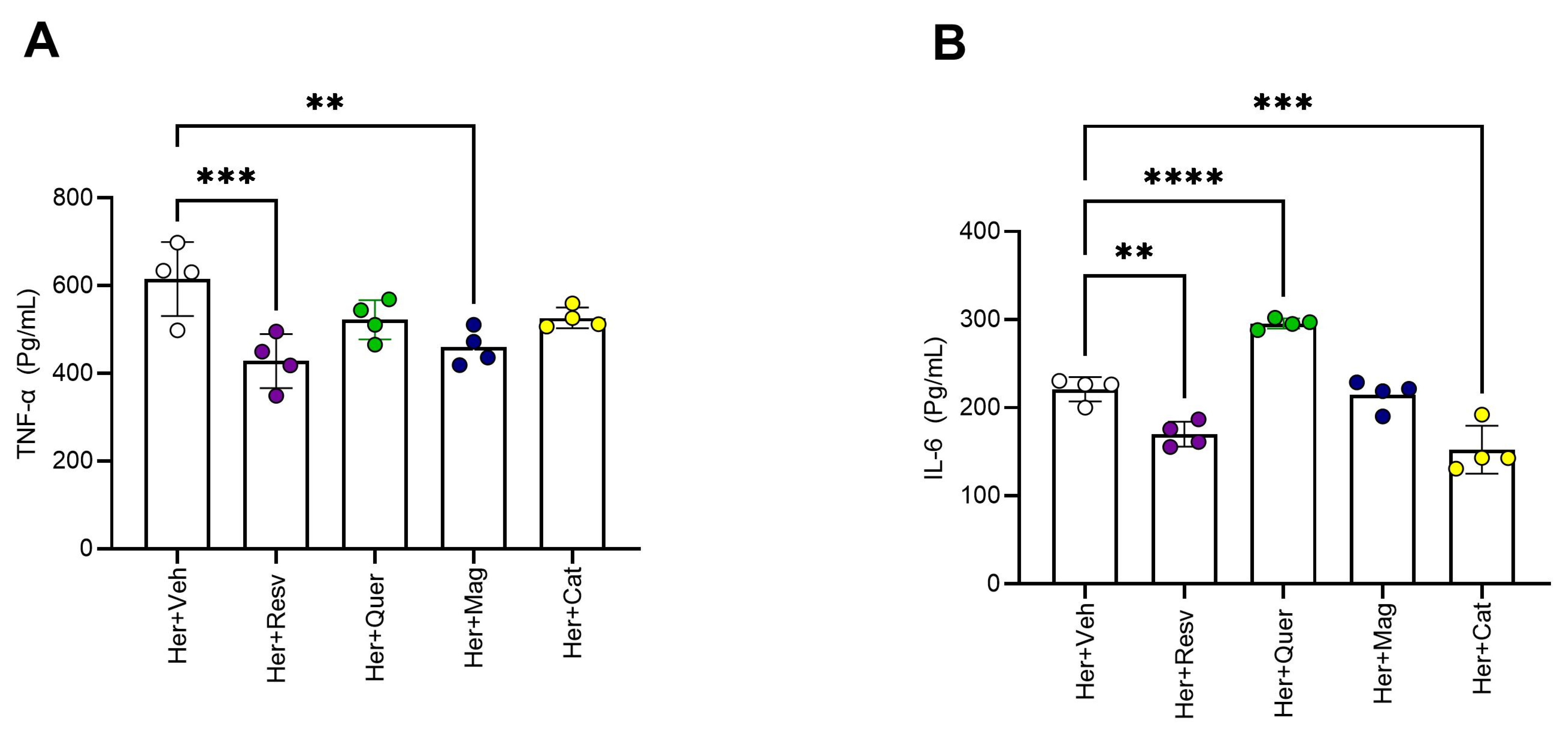
3.3. Molecular Effects
4. Discussion
4.1. Quercetin Abolished Heroin Dependence Acquisition and Inhibited Reinstatement Attributed to Anti-Inflammatory Effects
4.2. Resveratrol Blocked Heroin Acquisition and Drug-Induced Reinstatement Effectively, but Accentuated Withdrawal Manifestations
4.3. β-Catechin Blocked Reinstatement, but Not Acquisition of Heroin Dependence and Reduced the Withdrawal Manifestations
4.4. Magnolol Blocked Heroin-Induced Reinstatement, but Did Not Affect Acquisition and Withdrawal
4.5. Could Polyphenols Act through Dopaminergic Mechanisms?
4.6. Could the Current Results Be Related to an Anti-Inflammatory Mechanism?
4.7. The Role of Immunomodulatory Mechanisms
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Degenhardt, L.; Whiteford, H.A.; Ferrari, A.J.; Baxter, A.J.; Charlson, F.J.; Hall, W.D.; Freedman, G.; Burstein, R.; Johns, N.; Engell, R.E.; et al. Global burden of disease attributable to illicit drug use and dependence: Findings from the Global Burden of Disease Study 2010. Lancet 2013, 382, 1564–1574. [Google Scholar] [CrossRef]
- Lipari, R.N.; Hughes, A. Trends in Heroin Use in the United States: 2002 to 2013. In The CBHSQ Report; Substance Abuse and Mental Health Services Administration: Rockville, MD, USA, 2013; pp. 1–11. [Google Scholar]
- Martins, S.S.; Sarvet, A.; Santaella-Tenorio, J.; Saha, T.; Grant, B.F.; Hasin, D.S. Changes in US Lifetime Heroin Use and Heroin Use Disorder: Prevalence From the 2001–2002 to 2012–2013 National Epidemiologic Survey on Alcohol and Related Conditions. JAMA Psychiatry 2017, 74, 445–455. [Google Scholar] [CrossRef]
- Jones, C.M. Heroin use and heroin use risk behaviors among nonmedical users of prescription opioid pain relievers—United States, 2002–2004 and 2008–2010. Drug Alcohol Depend. 2013, 132, 95–100. [Google Scholar] [CrossRef]
- Azab, S.M.S.; Tawfik, H.; Hayes, B.D. Intoxication related to substances use in patients presenting to Ain Shams University Poisoning Treatment Center, Cairo, Egypt (2015–2019). Drug Alcohol Rev. 2022, 41, 1109–1118. [Google Scholar] [CrossRef]
- Connery, H.S. Medication-assisted treatment of opioid use disorder: Review of the evidence and future directions. Harv. Rev. Psychiatry 2015, 23, 63–75. [Google Scholar] [CrossRef]
- Kosten, T.R.; George, T.P. The neurobiology of opioid dependence: Implications for treatment. Sci. Pract. Perspect 2002, 1, 13–20. [Google Scholar] [CrossRef]
- Koehl, J.L.; Zimmerman, D.E.; Bridgeman, P.J. Medications for management of opioid use disorder. Am. J. Health Syst. Pharm. 2019, 76, 1097–1103. [Google Scholar] [CrossRef]
- Eastwood, B.; Strang, J.; Marsden, J. Effectiveness of treatment for opioid use disorder: A national, five-year, prospective, observational study in England. Drug Alcohol Depend. 2017, 176, 139–147. [Google Scholar] [CrossRef]
- Woods, J.S.; Joseph, H. From Narcotic to Normalizer: The Misperception of Methadone Treatment and the Persistence of Prejudice and Bias. Subst. Use Misuse 2018, 53, 323–329. [Google Scholar] [CrossRef]
- Polydorou, S.; Ross, S.; Coleman, P.; Duncan, L.; Roxas, N.; Thomas, A.; Mendoza, S.; Hansen, H. Integrating Buprenorphine Into an Opioid Treatment Program: Tailoring Care for Patients with Opioid Use Disorders. Psychiatr. Serv. 2017, 68, 295–298. [Google Scholar] [CrossRef]
- Johnson, B.; Richert, T. Diversion of methadone and buprenorphine by patients in opioid substitution treatment in Sweden: Prevalence estimates and risk factors. Int. J. Drug Policy 2015, 26, 183–190. [Google Scholar] [CrossRef]
- Hofford, R.S.; Russo, S.J.; Kiraly, D.D. Neuroimmune mechanisms of psychostimulant and opioid use disorders. Eur. J. Neurosci. 2019, 50, 2562–2573. [Google Scholar] [CrossRef]
- Roy, S.; Barke, R.A.; Loh, H.H. MU-opioid receptor-knockout mice: Role of mu-opioid receptor in morphine mediated immune functions. Brain Res. Mol. Brain Res. 1998, 61, 190–194. [Google Scholar] [CrossRef]
- Merighi, S.; Gessi, S.; Varani, K.; Fazzi, D.; Stefanelli, A.; Borea, P.A. Morphine mediates a proinflammatory phenotype via mu-opioid receptor-PKCvarepsilon-Akt-ERK1/2 signaling pathway in activated microglial cells. Biochem. Pharmacol. 2013, 86, 487–496. [Google Scholar] [CrossRef]
- Morcuende, A.; Navarrete, F.; Nieto, E.; Manzanares, J.; Femenia, T. Inflammatory Biomarkers in Addictive Disorders. Biomolecules 2021, 11, 1824. [Google Scholar] [CrossRef]
- Chan, Y.Y.; Yang, S.N.; Lin, J.C.; Chang, J.L.; Lin, J.G.; Lo, W.Y. Inflammatory response in heroin addicts undergoing methadone maintenance treatment. Psychiatry Res. 2015, 226, 230–234. [Google Scholar] [CrossRef]
- Bastami, S.; Norling, C.; Trinks, C.; Holmlund, B.; Walz, T.M.; Ahlner, J.; Uppugunduri, S. Inhibitory effect of opiates on LPS mediated release of TNF and IL-8. Acta Oncol. 2013, 52, 1022–1033. [Google Scholar] [CrossRef]
- Belkowski, S.M.; Alicea, C.; Eisenstein, T.K.; Adler, M.W.; Rogers, T.J. Inhibition of interleukin-1 and tumor necrosis factor-alpha synthesis following treatment of macrophages with the kappa opioid agonist U50, 488H. J. Pharmacol. Exp. Ther. 1995, 273, 1491–1496. [Google Scholar]
- Wang, W.; Lin, Q.; Lin, R.; Zhang, J.; Ren, F.; Zhang, J.; Ji, M.; Li, Y. PPARalpha agonist fenofibrate attenuates TNF-alpha-induced CD40 expression in 3T3-L1 adipocytes via the SIRT1-dependent signaling pathway. Exp. Cell Res. 2013, 319, 1523–1533. [Google Scholar] [CrossRef]
- Grabacka, M.; Pierzchalska, M.; Plonka, P.M.; Pierzchalski, P. The Role of PPAR Alpha in the Modulation of Innate Immunity. Int. J. Mol. Sci. 2021, 22, 10545. [Google Scholar] [CrossRef] [PubMed]
- Gehm, B.D.; McAndrews, J.M.; Chien, P.Y.; Jameson, J.L. Resveratrol, a polyphenolic compound found in grapes and wine, is an agonist for the estrogen receptor. Proc. Natl. Acad. Sci. USA 1997, 94, 14138–14143. [Google Scholar] [CrossRef] [PubMed]
- Erlund, I.; Freese, R.; Marniemi, J.; Hakala, P.; Alfthan, G. Bioavailability of quercetin from berries and the diet. Nutr. Cancer 2006, 54, 13–17. [Google Scholar] [CrossRef]
- Johnson, R.; Bryant, S.; Huntley, A.L. Green tea and green tea catechin extracts: An overview of the clinical evidence. Maturitas 2012, 73, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Sarrica, A.; Kirika, N.; Romeo, M.; Salmona, M.; Diomede, L. Safety and Toxicology of Magnolol and Honokiol. Planta Med. 2018, 84, 1151–1164. [Google Scholar] [CrossRef]
- Bhullar, K.S.; Rupasinghe, H.P. Polyphenols: Multipotent therapeutic agents in neurodegenerative diseases. Oxid. Med. Cell Longev. 2013, 2013, 891748. [Google Scholar] [CrossRef] [PubMed]
- Lee, K. Transactivation of peroxisome proliferator-activated receptor alpha by green tea extracts. J. Vet. Sci. 2004, 5, 325–330. [Google Scholar] [CrossRef]
- Marinovic, M.P.; Sousa-Filho, C.P.B.; Batista, F.A.H.; Avelino, T.M.; Cogliati, B.; Figueira, A.C.M.; Otton, R.; Rodrigues, A.C. Green tea extract increases adiponectin and PPAR alpha levels to improve hepatic steatosis. J. Nutr. Biochem. 2022, 103, 108957. [Google Scholar] [CrossRef]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: A concise review. J. Food Biochem. 2022, 46, e14264. [Google Scholar] [CrossRef] [PubMed]
- Yunusoglu, O. Resveratrol impairs acquisition, reinstatement and precipitates extinction of alcohol-induced place preference in mice. Neurol. Res. 2021, 43, 985–994. [Google Scholar] [CrossRef]
- Singh, A.; Naidu, P.S.; Kulkarni, S.K. Quercetin, a bioflavonoid, reverses development of tolerance and dependence to morphine. Drug Dev. Res. 2002, 57, 167–172. [Google Scholar] [CrossRef]
- Yunusoglu, O. Evaluation of the effects of quercetin on the rewarding property of ethanol in mice. Neurosci. Lett. 2022, 768, 136383. [Google Scholar] [CrossRef]
- Capasso, A.; Piacente, S.; Pizza, C.; Sorrentino, L. Flavonoids reduce morphine withdrawal in-vitro. J. Pharm. Pharmacol. 1998, 50, 561–564. [Google Scholar] [CrossRef]
- Abulseoud, O.A.; Miller, J.D.; Wu, J.; Choi, D.S.; Holschneider, D.P. Ceftriaxone upregulates the glutamate transporter in medial prefrontal cortex and blocks reinstatement of methamphetamine seeking in a condition place preference paradigm. Brain Res. 2012, 1456, 14–21. [Google Scholar] [CrossRef]
- Aguilar, M.A.; Rodriguez-Arias, M.; Minarro, J. Neurobiological mechanisms of the reinstatement of drug-conditioned place preference. Brain Res. Rev. 2009, 59, 253–277. [Google Scholar] [CrossRef]
- Joshi, D.; Naidu, P.S.; Singh, A.; Kulkarni, S.K. Protective effect of quercetin on alcohol abstinence-induced anxiety and convulsions. J. Med. Food 2005, 8, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. Protective effect of catechin on humoral and cell mediated immunity in rat model. Int. Immunopharmacol. 2018, 54, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Pei, Q.; Song, X.; Zhou, X.; Yin, Z.; Jia, R.; Zou, Y.; Li, L.; Yue, G.; Liang, X.; et al. The enhancement of immune function and activation of NF-kappaB by resveratrol-treatment in immunosuppressive mice. Int. Immunopharmacol. 2016, 33, 42–47. [Google Scholar] [CrossRef]
- Bai, Y.; Song, L.; Dai, G.; Xu, M.; Zhu, L.; Zhang, W.; Jing, W.; Ju, W. Antidepressant effects of magnolol in a mouse model of depression induced by chronic corticosterone injection. Steroids 2018, 135, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Popik, P.; Skolnick, P. The NMDA antagonist memantine blocks the expression and maintenance of morphine dependence. Pharmacol. Biochem. Behav. 1996, 53, 791–797. [Google Scholar] [CrossRef]
- Pum, M.E.; Huston, J.P.; Muller, C.P. The role of cortical serotonin in anxiety and locomotor activity in Wistar rats. Behav. Neurosci. 2009, 123, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Rahmadi, M.; Suasana, D.; Lailis, S.R.; Ratri, D.M.N.; Ardianto, C. The effects of quercetin on nicotine-induced reward effects in mice. J. Basic Clin. Physiol. Pharmacol. 2021, 32, 327–333. [Google Scholar] [CrossRef]
- Chen, F.; Sun, J.; Chen, C.; Zhang, Y.; Zou, L.; Zhang, Z.; Chen, M.; Wu, H.; Tian, W.; Liu, Y.; et al. Quercetin Mitigates Methamphetamine-Induced Anxiety-Like Behavior through Ameliorating Mitochondrial Dysfunction and Neuroinflammation. Front. Mol. Neurosci. 2022, 15, 829886. [Google Scholar] [CrossRef] [PubMed]
- Naidu, P.S.; Singh, A.; Joshi, D.; Kulkarni, S.K. Possible mechanisms of action in quercetin reversal of morphine tolerance and dependence. Addict. Biol. 2003, 8, 327–336. [Google Scholar] [CrossRef]
- Zeng, Q.; Xiong, Q.; Zhou, M.; Tian, X.; Yue, K.; Li, Y.; Ru, Q. Resveratrol attenuates methamphetamine-induced memory impairment via inhibition of oxidative stress and apoptosis in mice. J. Food Biochem. 2021, 45, e13622. [Google Scholar] [CrossRef] [PubMed]
- Calleri, E.; Pochetti, G.; Dossou, K.S.S.; Laghezza, A.; Montanari, R.; Capelli, D.; Prada, E.; Loiodice, F.; Massolini, G.; Bernier, M.; et al. Resveratrol and its metabolites bind to PPARs. Chembiochem 2014, 15, 1154–1160. [Google Scholar] [CrossRef]
- Shuto, T.; Kuroiwa, M.; Koga, Y.; Kawahara, Y.; Sotogaku, N.; Toyomasu, K.; Nishi, A. Acute effects of resveratrol to enhance cocaine-induced dopamine neurotransmission in the striatum. Neurosci. Lett. 2013, 542, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Panlilio, L.V.; Justinova, Z.; Goldberg, S.R. Inhibition of FAAH and activation of PPAR: New approaches to the treatment of cognitive dysfunction and drug addiction. Pharmacol. Ther. 2013, 138, 84–102. [Google Scholar] [CrossRef]
- Quiroga, C.; Barberena, J.J.; Alcaraz-Silva, J.; Machado, S.; Imperatori, C.; Yadollahpour, A.; Budde, H.; Yamamoto, T.; Telles-Correia, D.; Murillo-Rodríguez, E. The role of peroxisome proliferator-activated receptor in addiction: A novel drug target. Curr. Top. Med. Chem. 2021, 21, 964–975. [Google Scholar] [CrossRef]
- Singh, S.; Jamwal, S.; Kumar, P. Neuroprotective potential of Quercetin in combination with piperine against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurotoxicity. Neural Regen. Res. 2017, 12, 1137–1144. [Google Scholar]
- Jain, J.; Hasan, W.; Biswas, P.; Yadav, R.S.; Jat, D. Neuroprotective effect of quercetin against rotenone-induced neuroinflammation and alterations in mice behavior. J. Biochem. Mol. Toxicol. 2022, 36, e23165. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, R.; Chen, C.; Du, X.; Ruan, L.; Sun, J.; Li, J.; Zhang, L.; O’Donnell, J.M.; Pan, J. Antidepressant-like effect of trans-resveratrol in chronic stress model: Behavioral and neurochemical evidences. J. Psychiatr. Res. 2013, 47, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Dong, S.; Yi, L.; Geng, D.; Liu, Q. Magnolol abrogates chronic mild stress-induced depressive-like behaviors by inhibiting neuroinflammation and oxidative stress in the prefrontal cortex of mice. Int. Immunopharmacol. 2018, 59, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Kou, D.Q.; Jiang, Y.L.; Qin, J.H.; Huang, Y.H. Magnolol attenuates the inflammation and apoptosis through the activation of SIRT1 in experimental stroke rats. Pharmacol. Rep. 2017, 69, 642–647. [Google Scholar] [CrossRef]
- Tse, A.K.; Wan, C.K.; Zhu, G.Y.; Shen, X.L.; Cheung, H.Y.; Yang, M.; Fong, W.F. Magnolol suppresses NF-kappaB activation and NF-kappaB regulated gene expression through inhibition of IkappaB kinase activation. Mol. Immunol. 2007, 44, 2647–2658. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Shou, L.; Guo, X.; Wei, M.; Zheng, H.; Tao, T. Magnolol attenuates the locomotor impairment, cognitive deficit, and neuroinflammation in Alzheimer’s disease mice with brain insulin resistance via up-regulating miR-200c. Bioengineered 2022, 13, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Fu, W.; Chen, H.; You, S.; Liu, X.; Yang, Y.; Wei, Y.; Huang, J.; Rui, W. Magnolol attenuates the inflammation and enhances phagocytosis through the activation of MAPK, NF-kappaB signal pathways in vitro and in vivo. Mol. Immunol. 2019, 105, 96–106. [Google Scholar] [CrossRef]
- Xian, Y.F.; Qu, C.; Liu, Y.; Ip, S.P.; Yuan, Q.J.; Yang, W.; Lin, Z.X. Magnolol Ameliorates Behavioral Impairments and Neuropathology in a Transgenic Mouse Model of Alzheimer’s Disease. Oxid. Med. Cell Longev. 2020, 2020, 5920476. [Google Scholar] [CrossRef]
- Huang, S.Y.; Tai, S.H.; Chang, C.C.; Tu, Y.F.; Chang, C.H.; Lee, E.J. Magnolol protects against ischemic-reperfusion brain damage following oxygen-glucose deprivation and transient focal cerebral ischemia. Int. J. Mol. Med. 2018, 41, 2252–2262. [Google Scholar] [CrossRef]
- Lee, W.T.; Lin, M.H.; Lee, E.J.; Hung, Y.C.; Tai, S.H.; Chen, H.Y.; Chen, T.Y.; Wu, T.S. Magnolol reduces glutamate-induced neuronal excitotoxicity and protects against permanent focal cerebral ischemia up to 4 hours. PLoS ONE 2012, 7, e39952. [Google Scholar] [CrossRef]
- Zhou, F.; Jiang, Z.; Yang, B.; Hu, Z. Magnolol exhibits anti-inflammatory and neuroprotective effects in a rat model of intracerebral haemorrhage. Brain Behav. Immun. 2019, 77, 161–167. [Google Scholar] [CrossRef]
- Li, L.F.; Yang, J.; Ma, S.P.; Qu, R. Magnolol treatment reversed the glial pathology in an unpredictable chronic mild stress-induced rat model of depression. Eur. J. Pharmacol. 2013, 711, 42–49. [Google Scholar] [CrossRef]
- Matsui, N.; Akae, H.; Hirashima, N.; Kido, Y.; Tanabe, S.; Koseki, M.; Fukuyama, Y.; Akagi, M. Magnolol Enhances Hippocampal Neurogenesis and Exerts Antidepressant-Like Effects in Olfactory Bulbectomized Mice. Phytother. Res. 2016, 30, 1856–1861. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Hu, Y.; Chen, Z.; Dai, Y.; Hu, Y.; Qi, M. Magnolol attenuates depressive-like behaviors by polarizing microglia towards the M2 phenotype through the regulation of Nrf2/HO-1/NLRP3 signaling pathway. Phytomedicine 2021, 91, 153692. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Liu, R.; Chen, C.; Ji, F.; Li, T. Opioid System Modulates the Immune Function: A Review. Transl. Perioper. Pain Med. 2016, 1, 5–13. [Google Scholar] [PubMed]
- Eisenstein, T.K.; Rahim, R.T.; Feng, P.; Thingalaya, N.K.; Meissler, J.J. Effects of opioid tolerance and withdrawal on the immune system. J. Neuroimmune Pharmacol. 2006, 1, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Govitrapong, P.; Suttitum, T.; Kotchabhakdi, N.; Uneklabh, T. Alterations of immune functions in heroin addicts and heroin withdrawal subjects. J. Pharmacol. Exp. Ther. 1998, 286, 883–889. [Google Scholar]
- Weber, R.J.; Gomez-Flores, R.; Smith, J.E.; Martin, T.J. Immune, neuroendocrine, and somatic alterations in animal models of human heroin abuse. J. Neuroimmunol. 2004, 147, 134–137. [Google Scholar] [CrossRef]
- Piepenbrink, M.S.; Samuel, M.; Zheng, B.; Carter, B.; Fucile, C.; Bunce, C.; Kiebala, M.; Khan, A.A.; Thakar, J.; Maggirwar, S.B.; et al. Humoral Dysregulation Associated with Increased Systemic Inflammation among Injection Heroin Users. PLoS ONE 2016, 11, e0158641. [Google Scholar] [CrossRef]
- Sheng, W.S.; Hu, S.; Gekker, G.; Zhu, S.; Peterson, P.K.; Chao, C.C. Immunomodulatory role of opioids in the central nervous system. Arch. Immunol. Ther. Exp. 1997, 45, 359–366. [Google Scholar]
- Coller, J.K.; Hutchinson, M.R. Implications of central immune signaling caused by drugs of abuse: Mechanisms, mediators and new therapeutic approaches for prediction and treatment of drug dependence. Pharmacol. Ther. 2012, 134, 219–245. [Google Scholar] [CrossRef]
- Alesci, A.; Nicosia, N.; Fumia, A.; Giorgianni, F.; Santini, A.; Cicero, N. Resveratrol and Immune Cells: A Link to Improve Human Health. Molecules 2022, 27, 424. [Google Scholar] [CrossRef]
- Shakibaei, M.; Harikumar, K.B.; Aggarwal, B.B. Resveratrol addiction: To die or not to die. Mol. Nutr. Food Res. 2009, 53, 115–128. [Google Scholar] [CrossRef]
- Gualdoni, G.A.; Kovarik, J.J.; Hofer, J.; Dose, F.; Pignitter, M.; Doberer, D.; Steinberger, P.; Somoza, V.; Wolzt, M.; Zlabinger, G.J. Resveratrol enhances TNF-alpha production in human monocytes upon bacterial stimulation. Biochim. Biophys. Acta 2014, 1840, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.H.; Zhou, W.L.; Wu, Q.L.; Li, X.Y.; Zhao, W.M.; Zou, J.P. Low dose of resveratrol enhanced immune response of mice. Acta Pharmacol. Sin. 2002, 23, 893–897. [Google Scholar] [PubMed]
- Falchetti, R.; Fuggetta, M.P.; Lanzilli, G.; Tricarico, M.; Ravagnan, G. Effects of resveratrol on human immune cell function. Life Sci. 2001, 70, 81–96. [Google Scholar] [CrossRef]
- Malaguarnera, L. Influence of Resveratrol on the Immune Response. Nutrients 2019, 11, 946. [Google Scholar] [CrossRef]
- Xu, L.; Botchway, B.O.A.; Zhang, S.; Zhou, J.; Liu, X. Inhibition of NF-kappaB Signaling Pathway by Resveratrol Improves Spinal Cord Injury. Front. Neurosci. 2018, 12, 690. [Google Scholar] [CrossRef]
- Gao, X.; Xu, Y.X.; Janakiraman, N.; Chapman, R.A.; Gautam, S.C. Immunomodulatory activity of resveratrol: Suppression of lymphocyte proliferation, development of cell-mediated cytotoxicity, and cytokine production. Biochem. Pharmacol. 2001, 62, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Gangwar, V.; Garg, A.; Lomore, K.; Korla, K.; Bhat, S.S.; Rao, R.P.; Rafiq, M.; Kumawath, R.; Uddagiri, B.V.; Kareenhalli, V.V. Immunomodulatory Effects of a Concoction of Natural Bioactive Compounds-Mechanistic Insights. Biomedicines 2021, 9, 1522. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, J.; Yang, F.; Li, S.; Ma, W.; Chang, X.; Yang, L. Neuroprotective Effects of Quercetin on Ischemic Stroke: A Literature Review. Front. Pharmacol. 2022, 13, 854249. [Google Scholar] [CrossRef]
- Azeem, M.; Hanif, M.; Mahmood, K.; Ameer, N.; Chughtai, F.R.S.; Abid, U. An insight into anticancer, antioxidant, antimicrobial, antidiabetic and anti-inflammatory effects of quercetin: A review. Polym. Bull. 2022, 80, 241–262. [Google Scholar] [CrossRef] [PubMed]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Park, W. Anti-Inflammatory Effect of Quercetin on RAW 264.7 Mouse Macrophages Induced with Polyinosinic-Polycytidylic Acid. Molecules 2016, 21, 450. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Zhang, J.; Xie, J.; Yang, L.; Xing, Y.; Li, Z. The effects of quercetin on immunity, antioxidant indices, and disease resistance in zebrafish (Danio rerio). Fish Physiol Biochem. 2020, 46, 759–770. [Google Scholar] [CrossRef]
- Yang, J.X.; Maria, T.C.; Zhou, B.; Xiao, F.L.; Wang, M.; Mao, Y.J.; Li, Y. Quercetin improves immune function in Arbor Acre broilers through activation of NF-kappaB signaling pathway. Poult. Sci. 2020, 99, 2305. [Google Scholar] [CrossRef]
- Li, J.; Du, L.; He, J.N.; Chu, K.O.; Guo, C.L.; Wong, M.O.M.; Pang, C.P.; Chu, W.K. Anti-inflammatory Effects of GTE in Eye Diseases. Front. Nutr. 2021, 8, 753955. [Google Scholar] [CrossRef]
- Kim, J.M.; Heo, H.J. The roles of catechins in regulation of systemic inflammation. Food Sci. Biotechnol. 2022, 31, 957–970. [Google Scholar] [CrossRef]
- Baranwal, A.; Aggarwal, P.; Rai, A.; Kumar, N. Pharmacological Actions and Underlying Mechanisms of Catechin: A Review. Mini Rev. Med. Chem. 2022, 22, 821–833. [Google Scholar] [CrossRef]
- Cheng, A.W.; Tan, X.; Sun, J.Y.; Gu, C.M.; Liu, C.; Guo, X. Catechin attenuates TNF-alpha induced inflammatory response via AMPK-SIRT1 pathway in 3T3-L1 adipocytes. PLoS ONE 2019, 14, e0217090. [Google Scholar] [CrossRef]
- Mehta, R.; Bhandari, R.; Kuhad, A. Effects of catechin on a rodent model of autism spectrum disorder: Implications for the role of nitric oxide in neuroinflammatory pathway. Psychopharmacology 2021, 238, 3249–3271. [Google Scholar] [CrossRef]
- Kumari, M.V.; Yoneda, T.; Hiramatsu, M. Effect of “beta CATECHIN” on the life span of senescence accelerated mice (SAM-P8 strain). Biochem. Mol. Biol. Int. 1997, 41, 1005–1011. [Google Scholar] [PubMed]
- Wang, J.; Guo, X.; Chen, C.; Sun, S.; Liu, G.; Liu, M.; Hao, M.; Che, H. Gender differences in food allergy depend on the PPAR gamma/NF-kappaB in the intestines of mice. Life Sci. 2021, 278, 119606. [Google Scholar] [CrossRef] [PubMed]
- Santambrogio, P.; Cozzi, A.; Di Meo, I.; Cavestro, C.; Vergara, C.; Rodriguez-Pascau, L.; Martinell, M.; Pizcueta, P.; Tiranti, V.; Levi, S. PPAR Gamma Agonist Leriglitazone Recovers Alterations Due to Pank2-Deficiency in hiPS-Derived Astrocytes. Pharmaceutics 2023, 15, 202. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Zhao, J.; Song, X. Protective effects of fraxin on cerebral ischemia-reperfusion injury by mediating neuroinflammation and oxidative stress through PPAR-γ/NF-ΚB pathway. Brain Res. Bull. 2022, 187, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Soto, A.; Chavez-Santoscoy, R.A.; Porras, O.; Hidalgo-Ledesma, M.; Serrano-Medina, A.; Ramírez-Rodríguez, A.A.; Castillo-Martinez, N.A. Epicatechin and quercetin exhibit in vitro antioxidant effect, improve biochemical parameters related to metabolic syndrome, and decrease cellular genotoxicity in humans. Food Res. Int. 2021, 142, 110101. [Google Scholar] [CrossRef]
- Chhabra, G.; Singh, C.K.; Ndiaye, M.A.; Fedorowicz, S.; Molot, A.; Ahmad, N. Prostate cancer chemoprevention by natural agents: Clinical evidence and potential implications. Cancer Lett. 2018, 422, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Moussa, C.; Hebron, M.; Huang, X.; Ahn, J.; Rissman, R.A.; Aisen, P.S.; Turner, R.S. Resveratrol regulates neuro-inflammation and induces adaptive immunity in Alzheimer’s disease. J. Neuroinflamm. 2017, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Delpino, F.M.; Figueiredo, L.M. Resveratrol supplementation and type 2 diabetes: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2022, 62, 4465–4480. [Google Scholar] [CrossRef]
- Henning, S.M.; Wang, P.; Lee, R.P.; Trang, A.; Husari, G.; Yang, J.; Aronson, W.J. Prospective randomized trial evaluating blood and prostate tissue concentrations of green tea polyphenols and quercetin in men with prostate cancer. Food Funct. 2020, 11, 4114–4122. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
ElShebiney, S.; Elgohary, R.; El-Shamarka, M.; Mowaad, N.; Abulseoud, O.A. Natural Polyphenols—Resveratrol, Quercetin, Magnolol, and β-Catechin—Block Certain Aspects of Heroin Addiction and Modulate Striatal IL-6 and TNF-α. Toxics 2023, 11, 379. https://doi.org/10.3390/toxics11040379
ElShebiney S, Elgohary R, El-Shamarka M, Mowaad N, Abulseoud OA. Natural Polyphenols—Resveratrol, Quercetin, Magnolol, and β-Catechin—Block Certain Aspects of Heroin Addiction and Modulate Striatal IL-6 and TNF-α. Toxics. 2023; 11(4):379. https://doi.org/10.3390/toxics11040379
Chicago/Turabian StyleElShebiney, Shaimaa, Rania Elgohary, Marwa El-Shamarka, Noha Mowaad, and Osama A. Abulseoud. 2023. "Natural Polyphenols—Resveratrol, Quercetin, Magnolol, and β-Catechin—Block Certain Aspects of Heroin Addiction and Modulate Striatal IL-6 and TNF-α" Toxics 11, no. 4: 379. https://doi.org/10.3390/toxics11040379