
Advances in the Utilization of Zebrafish for Assessing and Understanding the Mechanisms of Nano-/Microparticles Toxicity in Water


 and
and
Abstract
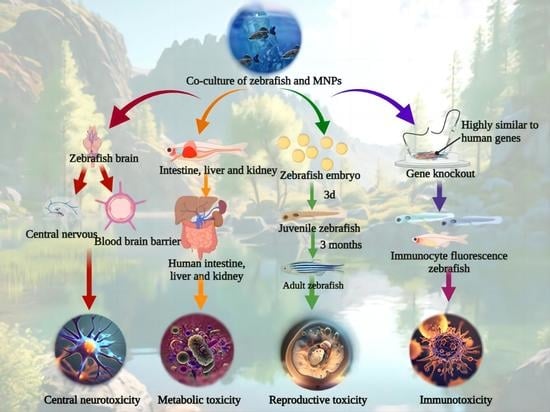
:
1. Introduction
2. Reliability of the Zebrafish for Toxicity Evaluation
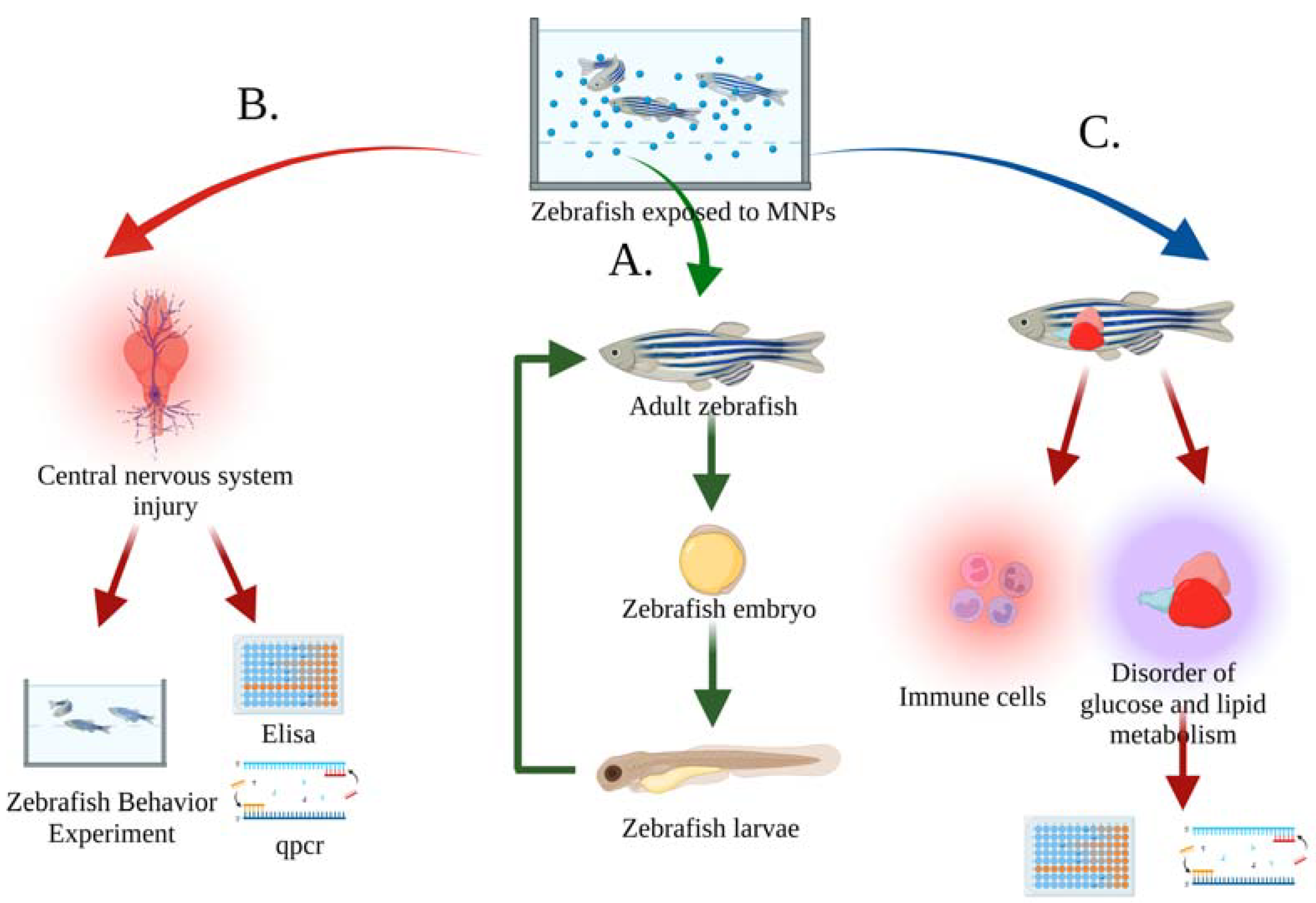
3. Toxicity of MNPs in Water to Zebrafish
3.1. Growth and Reproduction of Zebrafish
3.2. The Behavior and Nervous System of Zebrafish
3.3. Metabolism and Immune System of Zebrafish
4. Exploring the Toxicological Mechanisms of MNPs in Water with Zebrafish
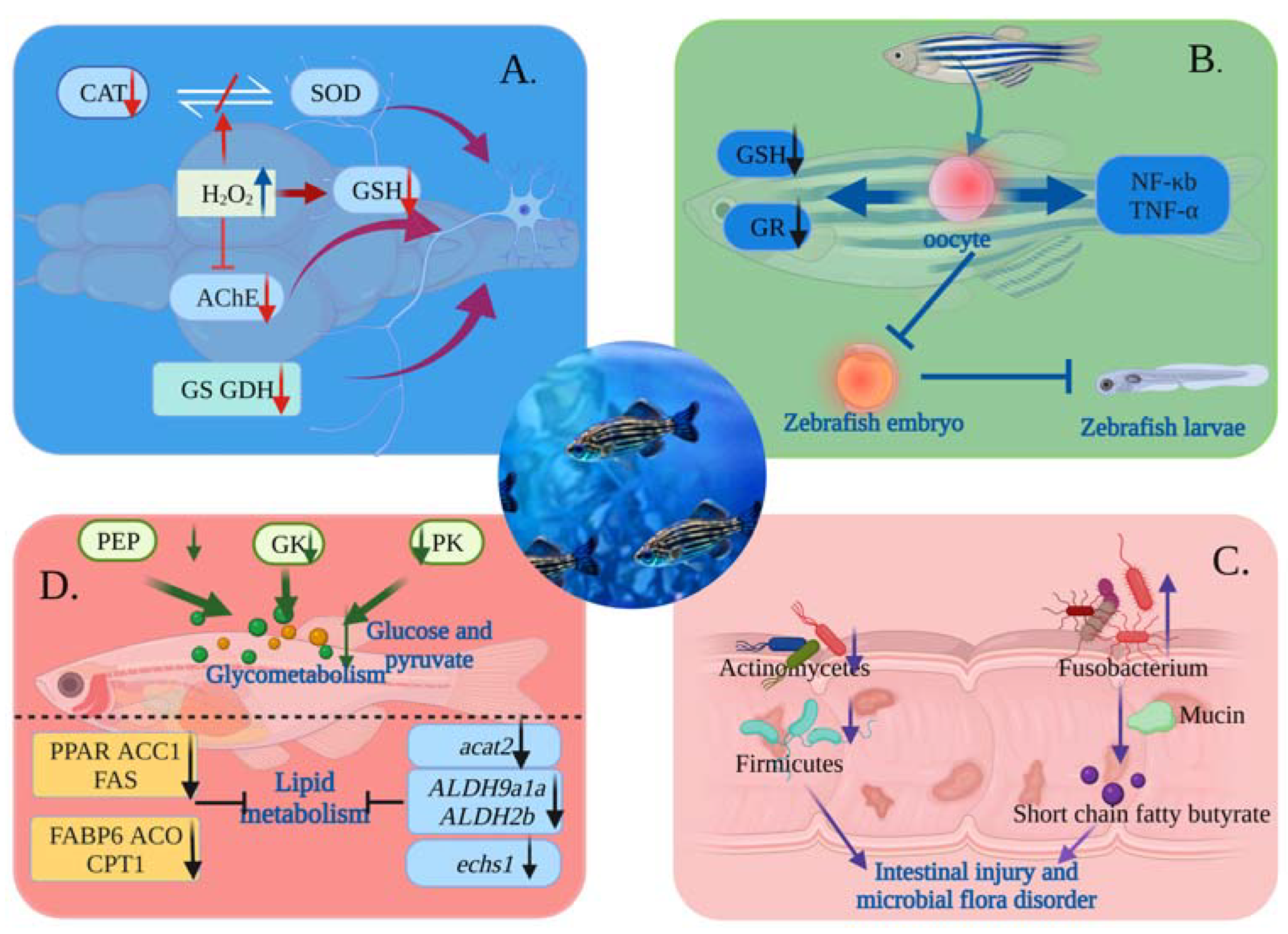
4.1. Toxicological Mechanism of MNPs on Nervous System
4.2. Toxicological Mechanism of MNPs Affecting Reproduction
4.3. Effect of MNPs on Gut Histopathology and Microbiota in Zebrafish
4.4. Toxicological Mechanisms of MNPs Causing Metabolic Disorders
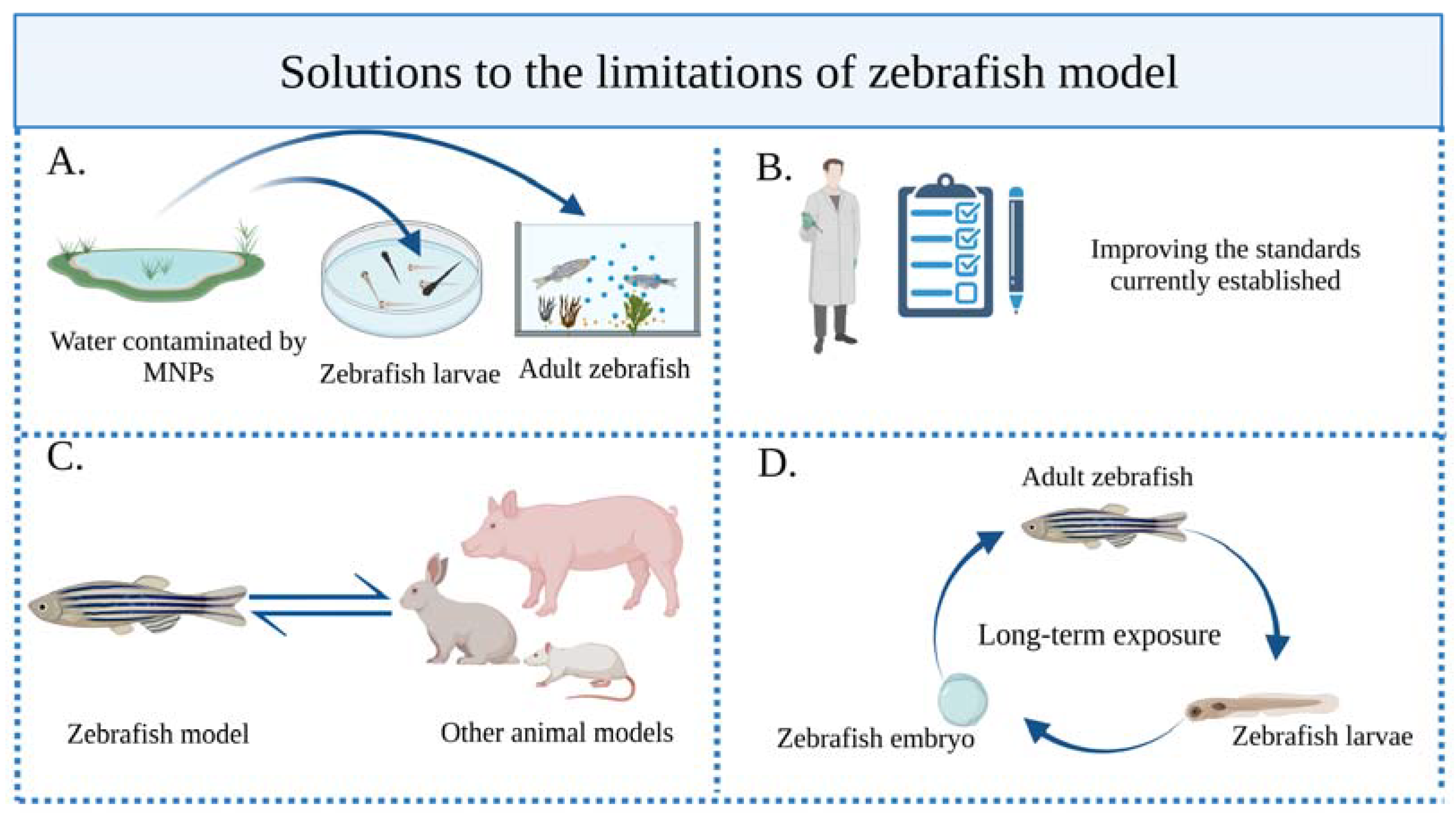
5. Limitations and Possible Solutions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MNPs | Nano-/microplastics |
| MPs | Microplastics |
| NPs | Nanoplastics |
| PE | Polyethylene |
| PS | Polystyrene |
| PP | Polypropylene |
| PVC | Polyvinyl chloride |
| PET | Polyethylene terephthalate |
| PD | Parkinson’s syndrome |
| AD | Alzheimer’s disease |
| ROS | Reactive oxygen species |
| AChE | Activity of acetylcholinesterase |
| IL-1α | Interleukin-1α |
| NF-κb | Nuclear factor-κb |
| CAT | Catalase |
| GSH | Glutathione |
| SOD | Superoxide dismutase |
| ACh | Acetylcholine neurotransmitter |
| GS | Glutamine synthetase |
| GDH | Glutamate dehydrogenase |
| GR | Glutathione reductase |
| TNF-α | Tumor necrosis factor-α |
| PEP | Phosphoenolpyruvate carboxykinase |
| GK | Glucokinase |
| PK | Pyruvate kinase |
| PPAR | Peroxisome proliferator-activated receptor |
| ACC1 | Acetyl-coa carboxylase 1 |
| FAS | Fatty acid synthase |
| ICD | Isocitrate dehydrogenase |
| FABP6 | Fatty acid binding protein 6 |
| ACO | Acyl-coa oxidase |
| CPT1 | Carnitine palmitoyltransferase 1 |
| Echs1 | Enoyl-coa hydratase 1 |
| TG | Triglyceride |
References
- Prata, J.C.; da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental exposure to microplastics: An overview on possible human health effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef]
- Banerjee, A.; Shelver, W.L. Micro- and nanoplastic induced cellular toxicity in mammals: A review. Sci. Total Environ. 2021, 755, 142518. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Kauffman, A.E.; Li, L.; McFee, W.; Cai, B.; Weinstein, J.; Lead, J.R.; Chatterjee, S.; Scott, G.I.; Xiao, S. Health impacts of environmental contamination of micro- and nanoplastics: A review. Environ. Health Prev. Med. 2020, 25, 29. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; He, Y.; Yan, Y.; Junaid, M.; Wang, J. Characteristics, Toxic Effects, and Analytical Methods of Microplastics in the Atmosphere. Nanomaterials 2021, 11, 2747. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, B.; Dissanayake, P.D.; Bolan, N.S.; Dar, J.Y.; Kumar, M.; Haque, M.N.; Mukhopadhyay, R.; Ramanayaka, S.; Biswas, J.K.; Tsang, D.C.W.; et al. Challenges and opportunities in sustainable management of microplastics and nanoplastics in the environment. Environ. Res. 2022, 207, 112179. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Chen, H.; Ma, J.; Fang, Y.; Qu, L.; Yang, Q.; Peng, B.; Zhang, X.; Jin, L.; Sun, D. Research progress on extraction technology and biomedical function of natural sugar substitutes. Front. Nutr. 2022, 9, 952147. [Google Scholar] [CrossRef]
- Qu, L.; Wang, L.; Ji, H.; Fang, Y.; Lei, P.; Zhang, X.; Jin, L.; Sun, D.; Dong, H. Toxic Mechanism and Biological Detoxification of Fumonisins. Toxins 2022, 14, 182. [Google Scholar] [CrossRef]
- Zhang, B.; Chao, J.; Chen, L.; Liu, L.; Yang, X.; Wang, Q. Research progress of nanoplastics in freshwater. Sci. Total Environ. 2021, 757, 143791. [Google Scholar] [CrossRef]
- Horton, A.A. Plastic pollution: When do we know enough? J. Hazard. Mater. 2022, 422, 126885. [Google Scholar] [CrossRef]
- Lionetto, F.; Corcione, C.E.; Rizzo, A.; Maffezzoli, A. Production and Characterization of Polyethylene Terephthalate Nanoparticles. Polymers 2021, 13, 3745. [Google Scholar] [CrossRef]
- Menon, V.; Sharma, S.; Gupta, S.; Ghosal, A.; Nadda, A.K.; Jose, R.; Sharma, P.; Kumar, S.; Singh, P.; Raizada, P. Prevalence and implications of microplastics in potable water system: An update. Chemosphere 2023, 317, 137848. [Google Scholar] [CrossRef]
- Liu, Z.; Xia, X.; Lv, X.; Song, E.; Song, Y. Iron-bearing nanoparticles trigger human umbilical vein endothelial cells ferroptotic responses by promoting intracellular iron level. Environ. Pollut. 2021, 287, 117345. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Lv, X.; Yang, B.; Qin, Q.; Song, E.; Song, Y. Tetrachlorobenzoquinone exposure triggers ferroptosis contributing to its neurotoxicity. Chemosphere 2021, 264, 128413. [Google Scholar] [CrossRef]
- Zhang, Q.; He, Y.; Cheng, R.; Li, Q.; Qian, Z.; Lin, X. Recent advances in toxicological research and potential health impact of microplastics and nanoplastics in vivo. Environ. Sci. Pollut. Res. Int. 2022, 29, 40415–40448. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.A.; Gedik, K.; Gaga, E.O. Atmospheric micro (nano) plastics: Future growing concerns for human health. Air Qual. Atmosphere Health 2023, 16, 233–262. [Google Scholar] [CrossRef]
- Huang, J.; Dong, G.; Liang, M.; Wu, X.; Xian, M.; An, Y.; Zhan, J.; Xu, L.; Xu, J.; Sun, W.; et al. Toxicity of micro(nano)plastics with different size and surface charge on human nasal epithelial cells and rats via intranasal exposure. Chemosphere 2022, 307, 136093. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Martins, M.A.; Soares, A.M.V.; Cuesta, A.; Oliveira, M. Polystyrene nanoplastics alter the cytotoxicity of human pharmaceuticals on marine fish cell lines. Environ. Toxicol. Pharmacol. 2019, 69, 57–65. [Google Scholar] [CrossRef]
- Banerjee, A.; Shelver, W.L. Micro- and Nanoplastic-Mediated Pathophysiological Changes in Rodents, Rabbits, and Chickens: A Review. J. Food Prot. 2021, 84, 1480–1495. [Google Scholar] [CrossRef]
- Lim, D.; Jeong, J.; Song, K.S.; Sung, J.H.; Oh, S.M.; Choi, J. Inhalation toxicity of polystyrene micro(nano)plastics using modified OECD TG 412. Chemosphere 2021, 262, 128330. [Google Scholar] [CrossRef]
- Ma, C.; Chen, Q.; Li, J.; Li, B.; Liang, W.; Su, L.; Shi, H. Distribution and translocation of micro- and nanoplastics in fish. Crit. Rev. Toxicol. 2021, 51, 740–753. [Google Scholar] [CrossRef]
- Yong, C.Q.Y.; Valiyaveettil, S.; Tang, B.L. Toxicity of Microplastics and Nanoplastics in Mammalian Systems. Int. J. Environ. Res. Public. Health 2020, 17, 1509. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kong, A.; Shelton, D.; Dong, H.; Li, J.; Zhao, F.; Bai, C.; Huang, K.; Mo, W.; Chen, S.; et al. Early life stage transient aristolochic acid exposure induces behavioral hyperactivity but not nephrotoxicity in larval zebrafish. Aquat. Toxicol. 2021, 238, 105916. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Ma, J.; Fang, Y.; Lei, P.; Wang, L.; Qu, L.; Wu, W.; Jin, L.; Sun, D. Advances in Zebrafish for Diabetes Mellitus with Wound Model. Bioengineering 2023, 10, 330. [Google Scholar] [CrossRef]
- Bhagat, J.; Zang, L.; Nishimura, N.; Shimada, Y. Zebrafish: An emerging model to study microplastic and nanoplastic toxicity. Sci. Total Environ. 2020, 728, 138707. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Freeman, J.L. Making Waves: New Developments in Toxicology with the Zebrafish. Toxicol. Sci. Off. J. Soc. Toxicol. 2018, 163, 5–12. [Google Scholar] [CrossRef]
- Bauer, B.; Mally, A.; Liedtke, D. Zebrafish Embryos and Larvae as Alternative Animal Models for Toxicity Testing. Int. J. Mol. Sci. 2021, 22, 13417. [Google Scholar] [CrossRef] [PubMed]
- Clément, Y.; Torbey, P.; Gilardi-Hebenstreit, P.; Crollius, H.R. Enhancer-gene maps in the human and zebrafish genomes using evolutionary linkage conservation. Nucleic Acids Res. 2020, 48, 2357–2371. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Batel, A.; Borchert, F.; Reinwald, H.; Erdinger, L.; Braunbeck, T. Microplastic accumulation patterns and transfer of benzo[a]pyrene to adult zebrafish (Danio rerio) gills and zebrafish embryos. Environ. Pollut. 2018, 235, 918–930. [Google Scholar] [CrossRef]
- Trevisan, R.; Voy, C.; Chen, S.; Di Giulio, R.T. Nanoplastics Decrease the Toxicity of a Complex PAH Mixture but Impair Mitochondrial Energy Production in Developing Zebrafish. Environ. Sci. Technol. 2019, 53, 8405–8415. [Google Scholar] [CrossRef]
- Li, Y.; Chen, T.; Miao, X.; Yi, X.; Wang, X.; Zhao, H.; Lee, S.M.-Y.; Zheng, Y. Zebrafish: A promising in vivo model for assessing the delivery of natural products, fluorescence dyes and drugs across the blood-brain barrier. Pharmacol. Res. 2017, 125, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef]
- Otsuna, H.; Hutcheson, D.A.; Duncan, R.N.; McPherson, A.D.; Scoresby, A.N.; Gaynes, B.F.; Tong, Z.; Fujimoto, E.; Kwan, K.M.; Chien, C.-B.; et al. High-resolution analysis of central nervous system expression patterns in zebrafish Gal4 enhancer-trap lines. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2015, 244, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.J.; Zhang, C.; Ojiaku, P.; Bell, V.; Devkota, S.; Mukhopadhyay, S. Effects of ethanol exposure on nervous system development in zebrafish. Int. Rev. Cell Mol. Biol. 2012, 299, 255–315. [Google Scholar] [CrossRef] [PubMed]
- Goessling, W.; Sadler, K.C. Zebrafish: An important tool for liver disease research. Gastroenterology 2015, 149, 1361–1377. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jin, Q.; Gao, C.; Chen, Y.; Sun, Z.; Guo, G.; Peng, J. Unraveling Differential Transcriptomes and Cell Types in Zebrafish Larvae Intestine and Liver. Cells 2022, 11, 3290. [Google Scholar] [CrossRef]
- Brugman, S. The zebrafish as a model to study intestinal inflammation. Dev. Comp. Immunol. 2016, 64, 82–92. [Google Scholar] [CrossRef]
- Sulukan, E.; Şenol, O.; Baran, A.; Kankaynar, M.; Yıldırım, S.; Kızıltan, T.; Bolat, İ.; Ceyhun, S.B. Nano-sized polystyrene plastic particles affect many cancer-related biological processes even in the next generations; zebrafish modeling. Sci. Total. Environ. 2022, 838, 156391. [Google Scholar] [CrossRef]
- Zhang, R.; Silic, M.R.; Schaber, A.; Wasel, O.; Freeman, J.L.; Sepúlveda, M.S. Exposure route affects the distribution and toxicity of polystyrene nanoplastics in zebrafish. Sci. Total. Environ. 2020, 724, 138065. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Maity, S.; Banerjee, S.; Dutta, S.; Adhikari, M.; Guchhait, R.; Biswas, C.; De, S.; Pramanick, K. Toxicological impacts of nanopolystyrene on zebrafish oocyte with insight into the mechanism of action: An expression-based analysis. Sci. Total. Environ. 2022, 830, 154796. [Google Scholar] [CrossRef]
- Pitt, J.A.; Trevisan, R.; Massarsky, A.; Kozal, J.S.; Levin, E.D.; Di Giulio, R.T. Maternal transfer of nanoplastics to offspring in zebrafish (Danio rerio): A case study with nanopolystyrene. Sci. Total. Environ. 2018, 643, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Malafaia, G.; de Souza, A.M.; Pereira, A.C.; Gonçalves, S.; da Costa Araújo, A.P.; Ribeiro, R.X.; Rocha, T.L. Developmental toxicity in zebrafish exposed to polyethylene microplastics under static and semi-static aquatic systems. Sci. Total. Environ. 2020, 700, 134867. [Google Scholar] [CrossRef]
- Teng, M.; Zhao, X.; Wu, F.; Wang, C.; Wang, C.; White, J.C.; Zhao, W.; Zhou, L.; Yan, S.; Tian, S. Charge-specific adverse effects of polystyrene nanoplastics on zebrafish (Danio rerio) development and behavior. Environ. Int. 2022, 163, 107154. [Google Scholar] [CrossRef] [PubMed]
- Aliakbarzadeh, F.; Rafiee, M.; Khodagholi, F.; Khorramizadeh, M.R.; Manouchehri, H.; Eslami, A.; Sayehmiri, F.; Mohseni-Bandpei, A. Adverse effects of polystyrene nanoplastic and its binary mixtures with nonylphenol on zebrafish nervous system: From oxidative stress to impaired neurotransmitter system. Environ. Pollut. 2023, 317, 120587. [Google Scholar] [CrossRef] [PubMed]
- Mak, C.W.; Ching-Fong Yeung, K.; Chan, K.M. Acute toxic effects of polyethylene microplastic on adult zebrafish. Ecotoxicol. Environ. Saf. 2019, 182, 109442. [Google Scholar] [CrossRef]
- Zhao, Y.; Qin, Z.; Huang, Z.; Bao, Z.; Luo, T.; Jin, Y. Effects of polyethylene microplastics on the microbiome and metabolism in larval zebrafish. Environ. Pollut. 2021, 282, 117039. [Google Scholar] [CrossRef]
- Cheng, H.; Duan, Z.; Wu, Y.; Wang, Y.; Zhang, H.; Shi, Y.; Zhang, H.; Wei, Y.; Sun, H. Immunotoxicity responses to polystyrene nanoplastics and their related mechanisms in the liver of zebrafish (Danio rerio) larvae. Environ. Int. 2022, 161, 107128. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bao, Z.; Wan, Z.; Fu, Z.; Jin, Y. Polystyrene microplastic exposure disturbs hepatic glycolipid metabolism at the physiological, biochemical, and transcriptomic levels in adult zebrafish. Sci. Total Environ. 2020, 710, 136279. [Google Scholar] [CrossRef]
- Sökmen, T.Ö.; Sulukan, E.; Türkoğlu, M.; Baran, A.; Özkaraca, M.; Ceyhun, S.B. Polystyrene nanoplastics (20 nm) are able to bioaccumulate and cause oxidative DNA damages in the brain tissue of zebrafish embryo (Danio rerio). Neurotoxicology 2020, 77, 51–59. [Google Scholar] [CrossRef]
- Bashirova, N.; Poppitz, D.; Klüver, N.; Scholz, S.; Matysik, J.; Alia, A. A mechanistic understanding of the effects of polyethylene terephthalate nanoplastics in the zebrafish (Danio rerio) embryo. Sci. Rep. 2023, 13, 1891. [Google Scholar] [CrossRef]
- Sarasamma, S.; Audira, G.; Siregar, P.; Malhotra, N.; Lai, Y.-H.; Liang, S.-T.; Chen, J.-R.; Chen, K.H.-C.; Hsiao, C.-D. Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure. Int. J. Mol. Sci. 2020, 21, 1410. [Google Scholar] [CrossRef]
- Tarasco, M.; Gavaia, P.J.; Bensimon-Brito, A.; Cordelières, F.P.; Santos, T.; Martins, G.; de Castro, D.T.; Silva, N.; Cabrita, E.; Bebianno, M.J.; et al. Effects of pristine or contaminated polyethylene microplastics on zebrafish development. Chemosphere 2022, 303, 135198. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.P.d.C.; da Luz, T.M.; Rocha, T.L.; Ahmed, M.A.I.; e Silva, D.D.D.M.; Rahman, M.M.; Malafaia, G. Toxicity evaluation of the combination of emerging pollutants with polyethylene microplastics in zebrafish: Perspective study of genotoxicity, mutagenicity, and redox unbalance. J. Hazard. Mater. 2022, 432, 128691. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.A.; Kozal, J.S.; Jayasundara, N.; Massarsky, A.; Trevisan, R.; Geitner, N.; Wiesner, M.; Levin, E.D.; Di Giulio, R.T. Uptake, tissue distribution, and toxicity of polystyrene nanoparticles in developing zebrafish (Danio rerio). Aquat. Toxicol. 2018, 194, 185–194. [Google Scholar] [CrossRef]
- Chen, Q.; Yin, D.; Jia, Y.; Schiwy, S.; Legradi, J.; Yang, S.; Hollert, H. Enhanced uptake of BPA in the presence of nanoplastics can lead to neurotoxic effects in adult zebrafish. Sci. Total. Environ. 2017, 609, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Xiao, Y.; Liu, Y.; Lei, Y.; Cai, Y.; Liang, Q.; Nie, S.; Jia, Y.; Chen, S.; Huang, C.; et al. Leachate from plastic food packaging induced reproductive and neurobehavioral toxicity in zebrafish. Ecotoxicol. Environ. Saf. 2022, 231, 113189. [Google Scholar] [CrossRef]
- Chen, Q.; Lackmann, C.; Wang, W.; Seiler, T.-B.; Hollert, H.; Shi, H. Microplastics Lead to Hyperactive Swimming Behaviour in Adult Zebrafish. Aquat. Toxicol. 2020, 224, 105521. [Google Scholar] [CrossRef]
- Yu, H.; Chen, Q.; Qiu, W.; Ma, C.; Gao, Z.; Chu, W.; Shi, H. Concurrent water- and foodborne exposure to microplastics leads to differential microplastic ingestion and neurotoxic effects in zebrafish. Water Res. 2022, 219, 118582. [Google Scholar] [CrossRef]
- Santos, D.; Luzio, A.; Matos, C.; Bellas, J.; Monteiro, S.M.; Félix, L. Microplastics alone or co-exposed with copper induce neurotoxicity and behavioral alterations on zebrafish larvae after a subchronic exposure. Aquat. Toxicol. 2021, 235, 105814. [Google Scholar] [CrossRef]
- Wright, S.L.; Rowe, D.; Thompson, R.C.; Galloway, T.S. Microplastic ingestion decreases energy reserves in marine worms. Curr. Biol. 2013, 23, R1031–R1033. [Google Scholar] [CrossRef]
- Rabezanahary, A.N.A.; Piette, M.; Missawi, O.; Garigliany, M.-M.; Kestemont, P.; Cornet, V. Microplastics alter development, behavior, and innate immunity responses following bacterial infection during zebrafish embryo-larval development. Chemosphere 2023, 311, 136969. [Google Scholar] [CrossRef]
- Limonta, G.; Mancia, A.; Benkhalqui, A.; Bertolucci, C.; Abelli, L.; Fossi, M.C.; Panti, C. Microplastics induce transcriptional changes, immune response and behavioral alterations in adult zebrafish. Sci. Rep. 2019, 9, 15775. [Google Scholar] [CrossRef] [PubMed]
- Bobori, D.C.; Feidantsis, K.; Dimitriadi, A.; Datsi, N.; Ripis, P.; Kalogiannis, S.; Sampsonidis, I.; Kastrinaki, G.; Ainali, N.M.; Lambropoulou, D.A.; et al. Dose-Dependent Cytotoxicity of Polypropylene Microplastics (PP-MPs) in Two Freshwater Fishes. Int. J. Mol. Sci. 2022, 23, 13878. [Google Scholar] [CrossRef] [PubMed]
- Bobori, D.C.; Dimitriadi, A.; Feidantsis, K.; Samiotaki, A.; Fafouti, D.; Sampsonidis, I.; Kalogiannis, S.; Kastrinaki, G.; Lambropoulou, D.A.; Kyzas, G.Z.; et al. Differentiation in the expression of toxic effects of polyethylene-microplastics on two freshwater fish species: Size matters. Sci. Total. Environ. 2022, 830, 154603. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Y.; Wang, Q.; Lv, M.; Zhao, X.; Ji, Y.; Han, X.; Wang, X.; Chen, L. The combined toxic effects of polyvinyl chloride microplastics and di(2-ethylhexyl) phthalate on the juvenile zebrafish (Danio rerio). J. Hazard. Mater. 2022, 440, 129711. [Google Scholar] [CrossRef]
- Zhou, R.; Zhou, D.; Yang, S.; Shi, Z.; Pan, H.; Jin, Q.; Ding, Z. Neurotoxicity of polystyrene nanoplastics with different particle sizes at environment-related concentrations on early zebrafish embryos. Sci. Total Environ. 2023, 872, 162096. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef]
- Kim, E.H.; Jeong, J.A.; Choi, E.K.; Jeong, T.-Y. Antioxidant enzyme activity in Daphnia magna under microscopic observation and shed carapace length as an alternative growth endpoint. Sci. Total. Environ. 2021, 794, 148771. [Google Scholar] [CrossRef]
- Qu, R.; Wang, X.; Wang, Z.; Wei, Z.; Wang, L. Metal accumulation and antioxidant defenses in the freshwater fish Carassius auratus in response to single and combined exposure to cadmium and hydroxylated multi-walled carbon nanotubes. J. Hazard. Mater. 2014, 275, 89–98. [Google Scholar] [CrossRef]
- Santos, D.; Félix, L.; Luzio, A.; Parra, S.; Cabecinha, E.; Bellas, J.; Monteiro, S.M. Toxicological effects induced on early life stages of zebrafish (Danio rerio) after an acute exposure to microplastics alone or co-exposed with copper. Chemosphere 2020, 261, 127748. [Google Scholar] [CrossRef]
- Singh, N.; Bhagat, J.; Tiwari, E.; Khandelwal, N.; Darbha, G.K.; Shyama, S.K. Metal oxide nanoparticles and polycyclic aromatic hydrocarbons alter nanoplastic’s stability and toxicity to zebrafish. J. Hazard. Mater. 2021, 407, 124382. [Google Scholar] [CrossRef] [PubMed]
- Schmidel, A.J.; Assmann, K.L.; Werlang, C.C.; Bertoncello, K.T.; Francescon, F.; Rambo, C.L.; Beltrame, G.M.; Calegari, D.; Batista, C.B.; Blaser, R.E.; et al. Subchronic atrazine exposure changes defensive behaviour profile and disrupts brain acetylcholinesterase activity of zebrafish. Neurotoxicol. Teratol. 2014, 44, 62–69. [Google Scholar] [CrossRef]
- Bashirzade, A.A.; Zabegalov, K.N.; Volgin, A.D.; Belova, A.S.; Demin, K.A.; de Abreu, M.S.; Babchenko, V.Y.; Bashirzade, K.A.; Yenkoyan, K.B.; Tikhonova, M.A.; et al. Modeling neurodegenerative disorders in zebrafish. Neurosci. Biobehav. Rev. 2022, 138, 104679. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cao, H. Zebrafish and Medaka: Important Animal Models for Human Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 10766. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Pal, R.; Siddiqi, N.J.; Sharma, B. Acetylcholinesterase from Human Erythrocytes as a Surrogate Biomarker of Lead Induced Neurotoxicity. Enzym. Res. 2015, 2015, e370705. [Google Scholar] [CrossRef] [PubMed]
- Garcimartín, A.; López-Oliva, M.E.; González, M.P.; Sánchez-Muniz, F.J.; Benedí, J. Hydrogen peroxide modifies both activity and isoforms of acetylcholinesterase in human neuroblastoma SH-SY5Y cells. Redox Biol. 2017, 12, 719–726. [Google Scholar] [CrossRef]
- Dadsetan, S.; Kukolj, E.; Bak, L.K.; SØrensen, M.; Ott, P.; Vilstrup, H.; Schousboe, A.; Keiding, S.; Waagepetersen, H.S. Brain Alanine Formation as an Ammonia-Scavenging Pathway during Hyperammonemia: Effects of Glutamine Synthetase Inhibition in Rats and Astrocyte—Neuron Co-Cultures. J. Cereb. Blood Flow Metab. 2013, 33, 1235–1241. [Google Scholar] [CrossRef]
- Gao, J.; Lin, L.; Wei, A.; Sepúlveda, M.S. Protein Corona Analysis of Silver Nanoparticles Exposed to Fish Plasma. Environ. Sci. Technol. Lett. 2017, 4, 174–179. [Google Scholar] [CrossRef]
- Rossi, G.; Barnoud, J.; Monticelli, L. Polystyrene Nanoparticles Perturb Lipid Membranes. J. Phys. Chem. Lett. 2014, 5, 241–246. [Google Scholar] [CrossRef]
- Sendra, M.; Pereiro, P.; Yeste, M.P.; Mercado, L.; Figueras, A.; Novoa, B. Size matters: Zebrafish (Danio rerio) as a model to study toxicity of nanoplastics from cells to the whole organism. Environ. Pollut. 2021, 268, 115769. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.; Wang, Q.; Sun, Y. Microplastics as a Vector for HOC Bioaccumulation in Earthworm Eisenia fetida in Soil: Importance of Chemical Diffusion and Particle Size. Environ. Sci. Technol. 2020, 54, 12154–12163. [Google Scholar] [CrossRef] [PubMed]
- Grigorakis, S.; Mason, S.A.; Drouillard, K.G. Determination of the gut retention of plastic microbeads and microfibers in goldfish (Carassius auratus). Chemosphere 2017, 169, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-M.; Byeon, E.; Jeong, H.; Kim, M.-S.; Chen, Q.; Lee, J.-S. Different effects of nano- and microplastics on oxidative status and gut microbiota in the marine medaka Oryzias melastigma. J. Hazard. Mater. 2021, 405, 124207. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Huang, A.; Cao, S.; Sun, F.; Wang, L.; Guo, H.; Ji, R. Effects of nanoplastics and microplastics on toxicity, bioaccumulation, and environmental fate of phenanthrene in fresh water. Environ. Pollut. 2016, 219, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, L.; Huang, Q.; Dong, S.; Wang, X.; Yan, C. Combined effects of micro-/nano-plastics and oxytetracycline on the intestinal histopathology and microbiome in zebrafish (Danio rerio). Sci. Total Environ. 2022, 843, 156917. [Google Scholar] [CrossRef] [PubMed]
- Engevik, M.A.; Danhof, H.A.; Ruan, W.; Engevik, A.C.; Chang-Graham, A.L.; Engevik, K.A.; Shi, Z.; Zhao, Y.; Brand, C.K.; Krystofiak, E.S.; et al. Fusobacterium nucleatum Secretes Outer Membrane Vesicles and Promotes Intestinal Inflammation. mBio 2021, 12, e02706–e02720. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef]
- Alvarez, A.; Saez, J.M.; Davila Costa, J.S.; Colin, V.L.; Fuentes, M.S.; Cuozzo, S.A.; Benimeli, C.S.; Polti, M.A.; Amoroso, M.J. Actinobacteria: Current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere 2017, 166, 41–62. [Google Scholar] [CrossRef]
- Collinder, E.; Björnhag, G.; Cardona, M.; Norin, E.; Rehbinder, C.; Midtvedt, T. Gastrointestinal Host–Microbial Interactions in Mammals and Fish: Comparative Studies in Man, Mice, Rats, Pigs, Horses, Cows, Elks, Reindeers, Salmon and Cod. Microb. Ecol. Health Dis. 2003, 15, 66–78. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Marx, T. Chapter 35—Immunoprotective Effects of Probiotics in the Elderly. In Foods and Dietary Supplements in the Prevention and Treatment of Disease in Older Adults; Watson, R.R., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 363–372. ISBN 978-0-12-418680-4. [Google Scholar]
- Barletta, M.; Lima, A.R.A.; Costa, M.F. Distribution, sources and consequences of nutrients, persistent organic pollutants, metals and microplastics in South American estuaries. Sci. Total Environ. 2019, 651, 1199–1218. [Google Scholar] [CrossRef] [PubMed]
- Mizraji, R.; Ahrendt, C.; Perez-Venegas, D.; Vargas, J.; Pulgar, J.; Aldana, M.; Patricio Ojeda, F.; Duarte, C.; Galbán-Malagón, C. Is the feeding type related with the content of microplastics in intertidal fish gut? Mar. Pollut. Bull. 2017, 116, 498–500. [Google Scholar] [CrossRef] [PubMed]
- Santangeli, S.; Notarstefano, V.; Maradonna, F.; Giorgini, E.; Gioacchini, G.; Forner-Piquer, I.; Habibi, H.R.; Carnevali, O. Effects of diethylene glycol dibenzoate and Bisphenol A on the lipid metabolism of Danio rerio. Sci. Total Environ. 2018, 636, 641–655. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, Y.; Deng, M.; Wang, X.; Tu, W.; Fu, Z.; Jin, Y. Bioaccumulation in the gut and liver causes gut barrier dysfunction and hepatic metabolism disorder in mice after exposure to low doses of OBS. Environ. Int. 2019, 129, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lei, P.; Chen, H.; Wang, L.; Fang, Y.; Yan, X.; Yang, Q.; Peng, B.; Jin, L.; Sun, D. Advances in lncRNAs from stem cell-derived exosome for the treatment of cardiovascular diseases. Front. Pharmacol. 2022, 13, 986683. [Google Scholar] [CrossRef]
- Zuo, Q.; Zhang, G.; He, L.; Ma, S.; Ma, H.; Zhai, J.; Wang, Z.; Zhang, T.; Wang, Y.; Guo, Y. Canagliflozin Attenuates Hepatic Steatosis and Atherosclerosis Progression in Western Diet-Fed ApoE-Knockout Mice. Drug Des. Devel. Ther. 2022, 16, 4161–4177. [Google Scholar] [CrossRef]
- Oaxaca-Castillo, D.; Andreoletti, P.; Vluggens, A.; Yu, S.; van Veldhoven, P.P.; Reddy, J.K.; Cherkaoui-Malki, M. Biochemical characterization of two functional human liver acyl-CoA oxidase isoforms 1a and 1b encoded by a single gene. Biochem. Biophys. Res. Commun. 2007, 360, 314–319. [Google Scholar] [CrossRef]
- Parini, P.; Jiang, Z.-Y.; Einarsson, C.; Eggertsen, G.; Zhang, S.-D.; Rudel, L.L.; Han, T.-Q.; Eriksson, M. ACAT2 and human hepatic cholesterol metabolism: Identification of important gender-related differences in normolipidemic, non-obese Chinese patients. Atherosclerosis 2009, 207, 266–271. [Google Scholar] [CrossRef]
- Dalby, P.A.; Aucamp, J.P.; George, R.; Martinez-Torres, R.J. Structural stability of an enzyme biocatalyst. Biochem. Soc. Trans. 2007, 35, 1606–1609. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Zhang, H.; Dai, J. Proteomic analysis of male zebrafish livers chronically exposed to perfluorononanoic acid. Environ. Int. 2012, 42, 20–30. [Google Scholar] [CrossRef]
- Zhang, J.; Ibrahim, M.M.; Sun, M.; Tang, J. Enoyl-coenzyme A hydratase in cancer. Clin. Chim. Acta Int. J. Clin. Chem. 2015, 448, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Sobanska, M.; Scholz, S.; Nyman, A.-M.; Cesnaitis, R.; Gutierrez Alonso, S.; Klüver, N.; Kühne, R.; Tyle, H.; de Knecht, J.; Dang, Z.; et al. Applicability of the fish embryo acute toxicity (FET) test (OECD 236) in the regulatory context of Registration, Evaluation, Authorisation, and Restriction of Chemicals (REACH). Environ. Toxicol. Chem. 2018, 37, 657–670. [Google Scholar] [CrossRef] [PubMed]






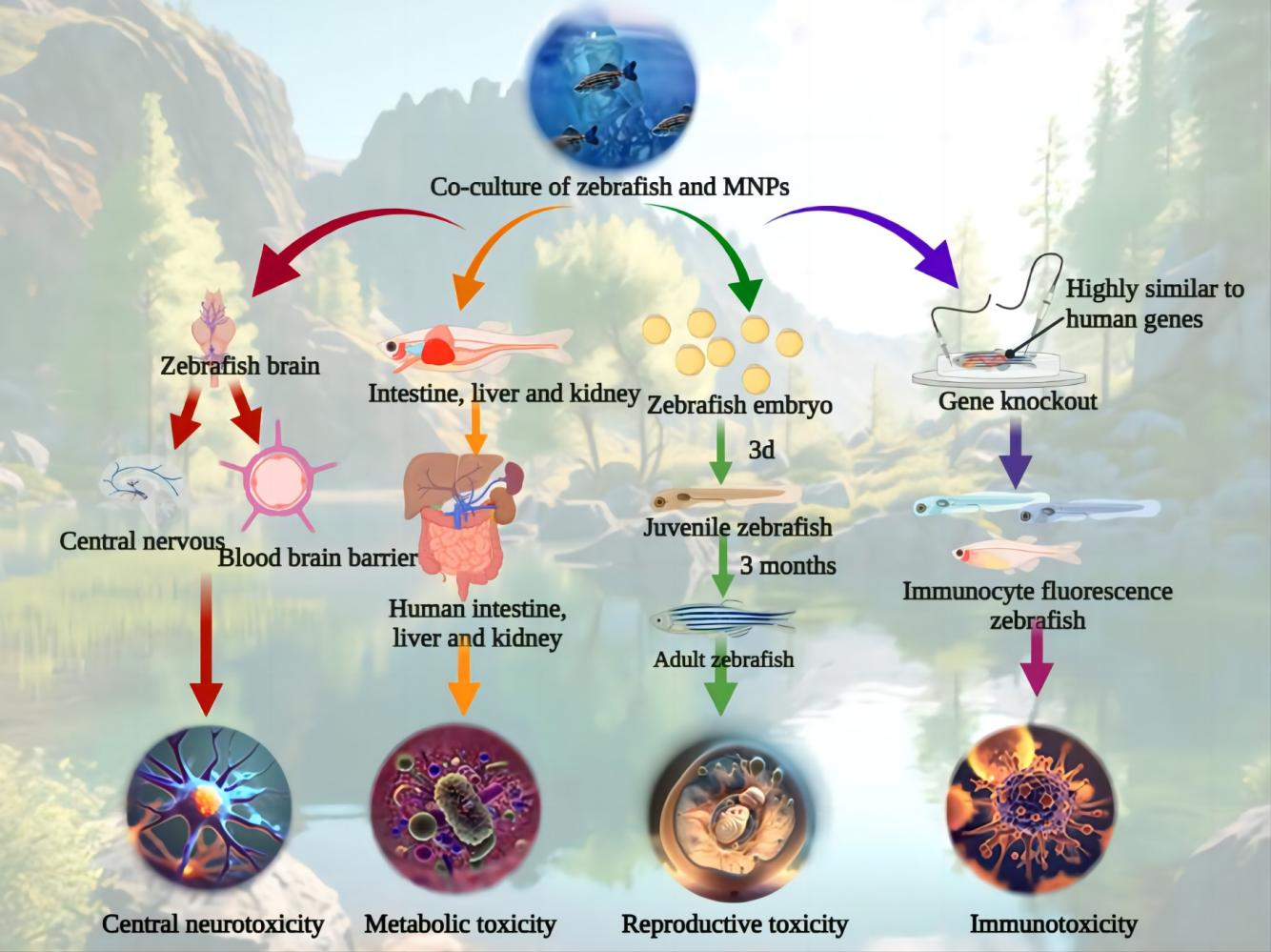{kind=link}
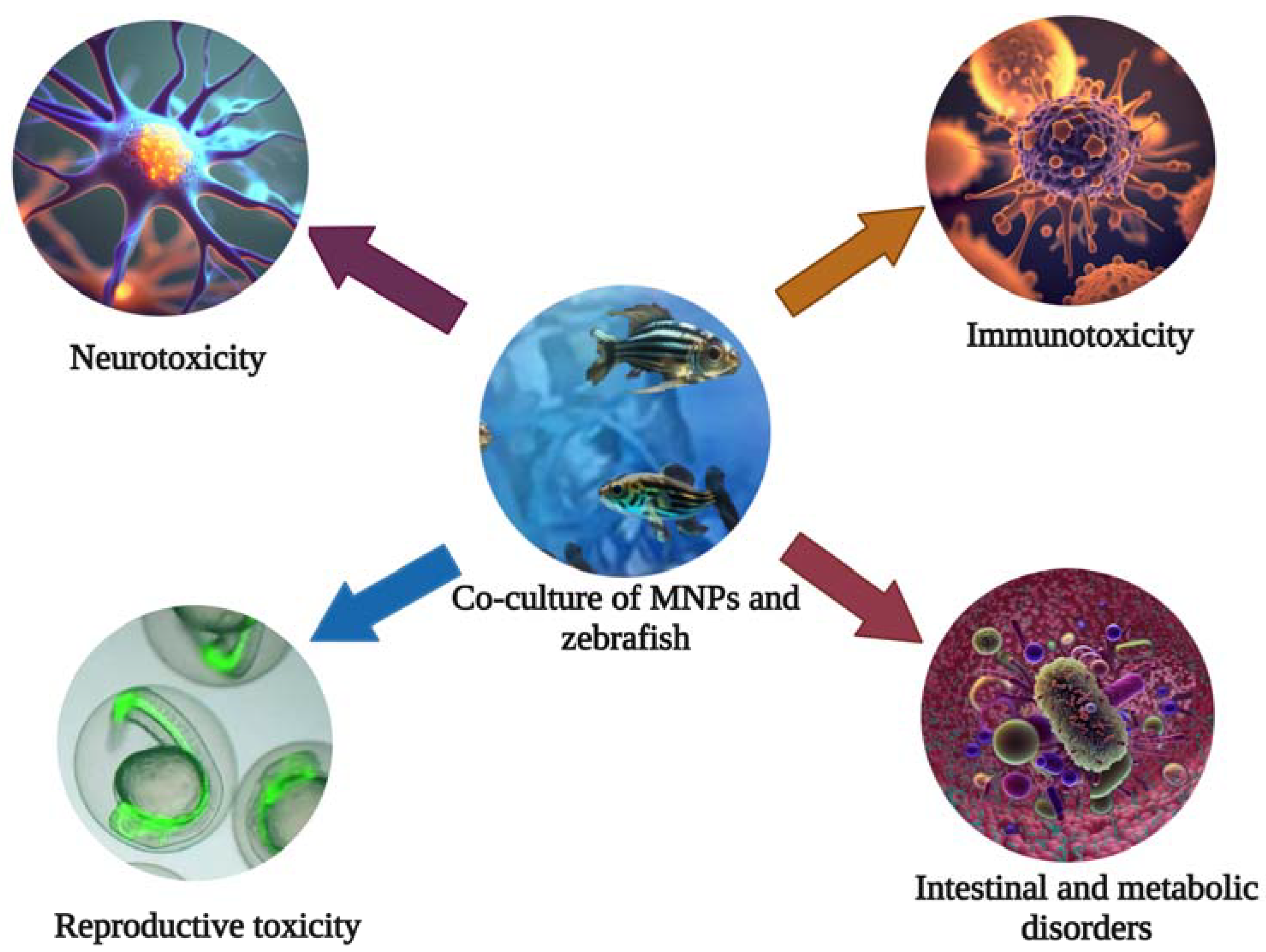{kind=link}
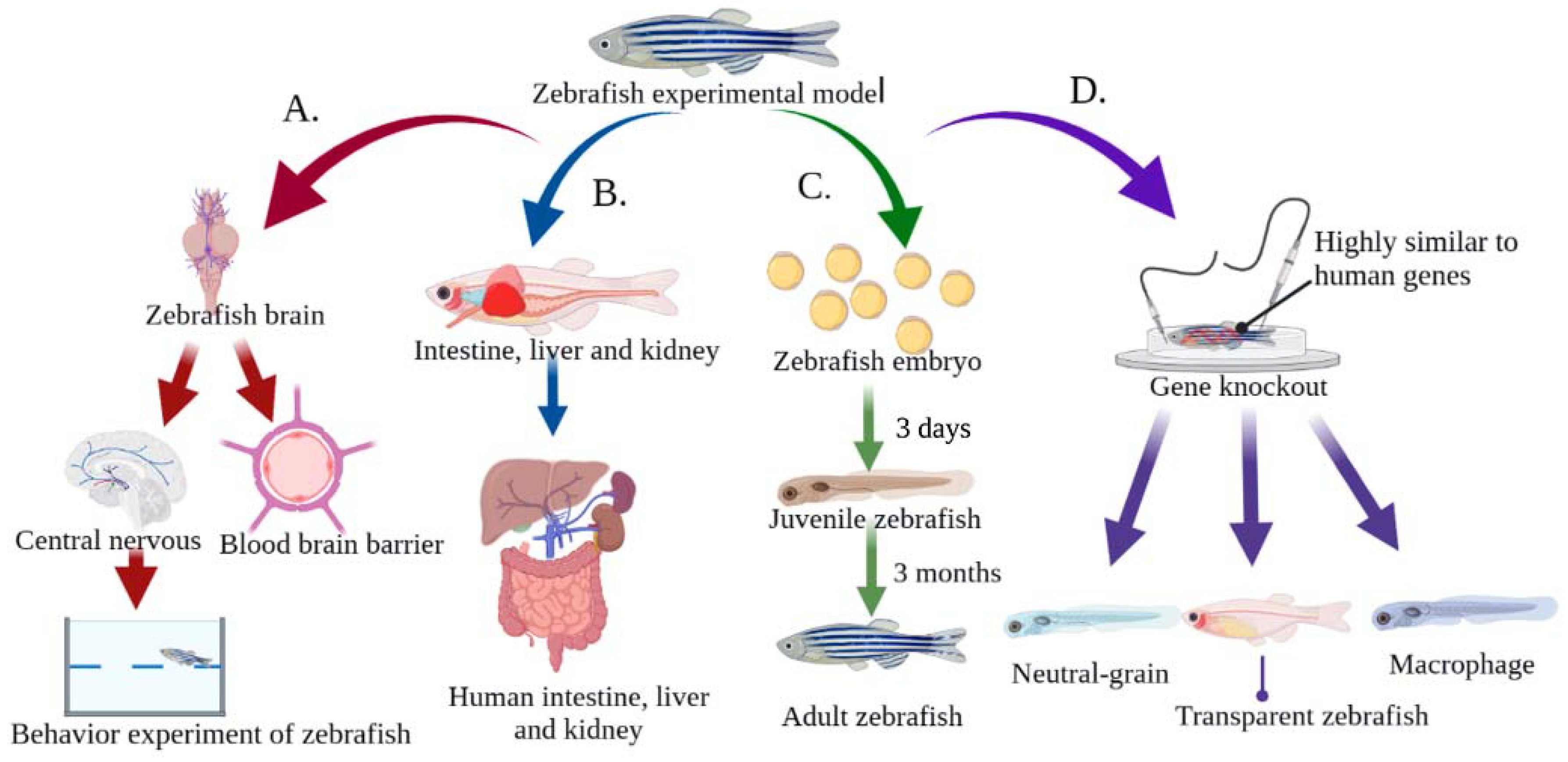{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxicity of MNPs | Types of MNPs | Experimental Model | Days of Exposure | Harm Caused | References |
|---|---|---|---|---|---|
| Growth and reproductive toxicity | PS-NPs | Adult zebrafish and their offspring | Adult zebrafish: 7 days, larvae: 96 h | Significantly reduced glutathione reductase activity in adult zebrafish brain, muscle, and testis. In addition, GSH reductase activity and thiol levels were decreased and the oxidative system was disrupted in embryos and larvae of female and co-parent exposed offspring. | [41] |
| PET-NPs | Zebrafish embryos and larvae | 96 h | Embryo hatching rate and survival rate decreased dramatically, accompanied by impaired mitochondrial-membrane integrity and significant alterations in oxidative-stress-related pathways. | [50] | |
| PE-NPs | Adult zebrafish | 360 days | Zebrafish fecundity, egg morphology, and yolk area were impaired and skeletal deformities and impaired development of the caudal fins and scales. Malformations were also observed in the offspring. | [52] | |
| PVC-MPs | Zebrafish embryos and larvae | 9 days | It delayed embryo hatching, cause zebrafish death and induce oxidative stress, and inhibited heart development. | [66] | |
| PE-MPs | Adult zebrafish | 15 days | It caused DNA damage and nuclear abnormality in red blood cells, as well as a decrease in SOD and CAT activities, and a large amount of ROS was produced, which caused oxidative stress. | [53] | |
| PS-NPs | Zebrafish oocytes | 6 h | It caused oxidative stress, immunotoxicity, and apoptosis in oocytes. | [40] | |
| Neurotoxicity | PS-MPs | Adult zebrafish | 7 days | Zebrafish exhibited anxious behavior, increased swimming distance, and longer periods of manic and active states. | [58] |
| PS-NPs | Adult zebrafish | 7 days | PS-NPs accumulated in zebrafish brains and the fish exhibited circadian rhythm activity, aggressive behavior, anxiety behavior, and predator-avoidance behavior. In addition, the expression of neurotoxicity-related neurotransmitters such as dopamine, 5-hydroxytryptamine, and γ-aminobutyric acid was decreased. | [51] | |
| PE-MPs | Adult zebrafish | 4 days | The zebrafish showed erratic movements and epileptic behavior with the tail bent downward or upward | [45] | |
| Mixture of MPs | Zebrafish larvae | 14 days | It reduced the average swimming speed and total distance of zebrafish larvae. It affected avoidance behavior and aversive stimulus response, and significantly inhibited the AChE activity of zebrafish larvae. | [60] | |
| PS-NPs | Zebrafish larvae | 120 h | Comparing PS-NH2 (positively charged) to PS-COOH (negatively charged), PS-NH2 caused greater developmental toxicity and apoptosis in the brain and greater neurobehavioral impairment as well as a reduction in glycine, cysteine, glutathione, and glutamate levels. | [43] | |
| PS-NPs | Zebrafish embryos and larvae | 120 h | PS-NPs entered the embryo and brain, harmed embryonic development, induced neuronal loss, specifically interfered with the GABAergic, cholinergic, and serotonergic systems, and affected neuronal signaling, resulting in behavioral abnormalities. | [67] | |
| Metabolic and immune system toxicity | PS-MPs | Zebrafish larvae | 8 days | The expression of genes related to oxidative stress and immune system response was upregulated, and cell apoptosis occurred. | [62] |
| PE-MPs | Zebrafish embryos and larvae | 7 days | Glucose and lipid metabolism was disturbed, and the metabolism of triglycerides, total cholesterol, non-esterified fatty acids, total bile acids, glucose, and pyruvate was disturbed. | [46] | |
| PS-MPs | Adult male zebrafish | 21 days | Glucose, lipid, and amino-acid metabolism in the liver was disordered, and the amounts of glucose, pyruvate, and α-ketoglutarate in the liver were decreased. | [48] | |
| PE-MPs | Adult zebrafish | 21 days | It caused DNA damage and increased ubiquitination levels, while causing oxidative stress, leading to lipid peroxidation and apoptosis. | [65] | |
| PP-MPs | Adult zebrafish | 21 days | It caused lipid peroxidation, DNA damage, protein ubiquitination, cell apoptosis, and autophagy, and inhibited the functions of zebrafish gill and liver cells. | [64] | |
| PS-MPs | Adult zebrafish | 7 days | It induced oxidative stress and disrupted fat and energy metabolism, resulting in liver inflammation and lipid accumulation in zebrafish. | [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, P.; Zhang, W.; Ma, J.; Xia, Y.; Yu, H.; Du, J.; Fang, Y.; Wang, L.; Zhang, K.; Jin, L.; et al. Advances in the Utilization of Zebrafish for Assessing and Understanding the Mechanisms of Nano-/Microparticles Toxicity in Water. Toxics 2023, 11, 380. https://doi.org/10.3390/toxics11040380
Lei P, Zhang W, Ma J, Xia Y, Yu H, Du J, Fang Y, Wang L, Zhang K, Jin L, et al. Advances in the Utilization of Zebrafish for Assessing and Understanding the Mechanisms of Nano-/Microparticles Toxicity in Water. Toxics. 2023; 11(4):380. https://doi.org/10.3390/toxics11040380
Chicago/Turabian StyleLei, Pengyu, Wenxia Zhang, Jiahui Ma, Yuping Xia, Haiyang Yu, Jiao Du, Yimeng Fang, Lei Wang, Kun Zhang, Libo Jin, and et al. 2023. "Advances in the Utilization of Zebrafish for Assessing and Understanding the Mechanisms of Nano-/Microparticles Toxicity in Water" Toxics 11, no. 4: 380. https://doi.org/10.3390/toxics11040380