
The Difference between Rhizosphere and Endophytic Bacteria on the Safe Cultivation of Lettuce in Cr-Contaminated Farmland


Abstract
:
1. Introduction
2. Materials and Methods
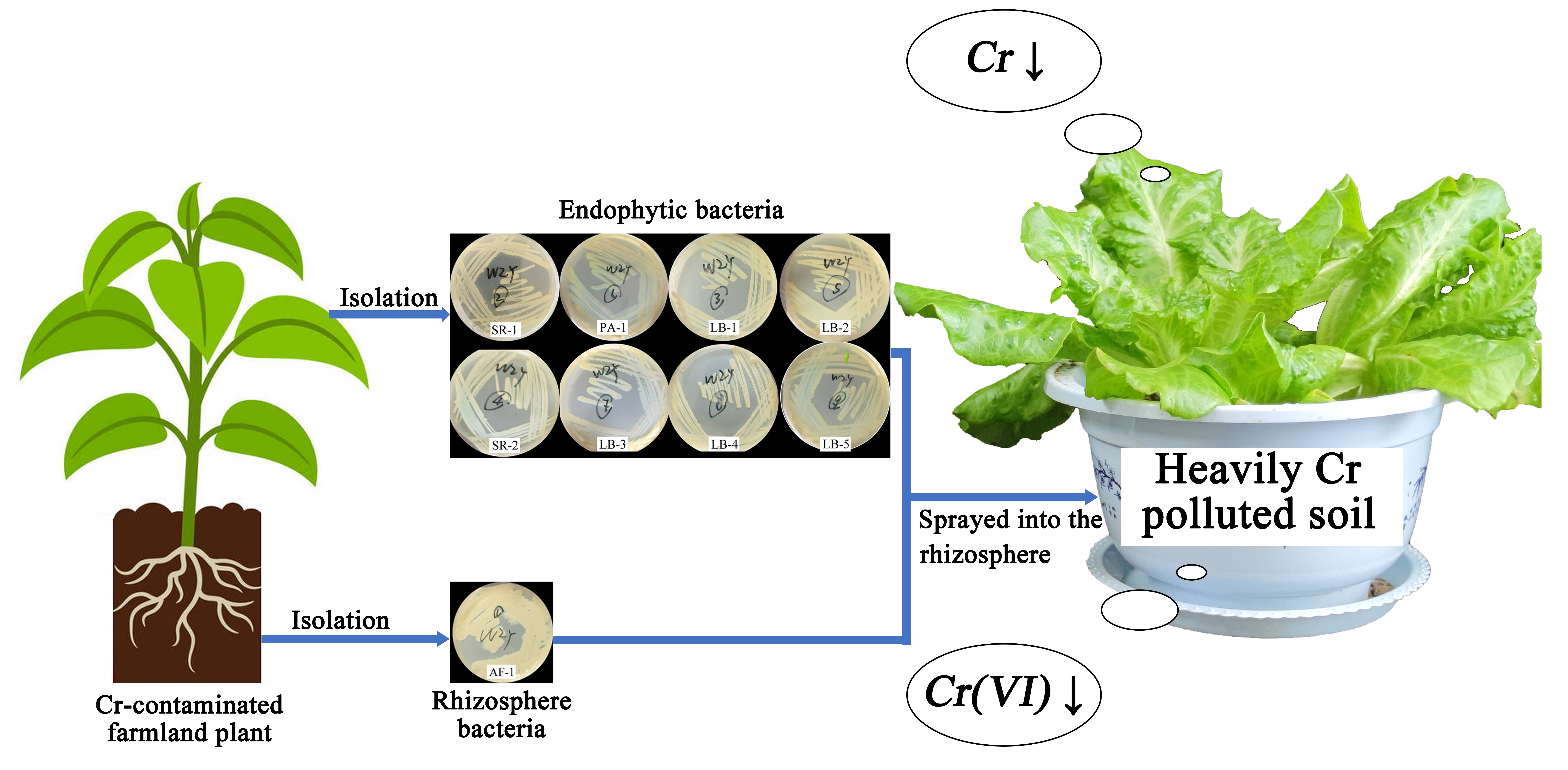
2.1. The Collection of Cr-Contaminated Soil and Plant Breeding
2.2. Isolation and Identification of Bacterial Strains
2.3. Experimental Design
2.4. Plant Sample Analysis
2.5. Rhizosphere Soil Sample Analysis
2.6. Quality Control
2.7. Translocation Factor (TF) and Bioaccumulation Factor (BAF)
2.8. Statistical Analysis
3. Results
3.1. Isolation and Identification of Cr-Tolerant Bacteria
3.2. Influence of Bacteria on Rhizosphere Soil Physiochemical Properties
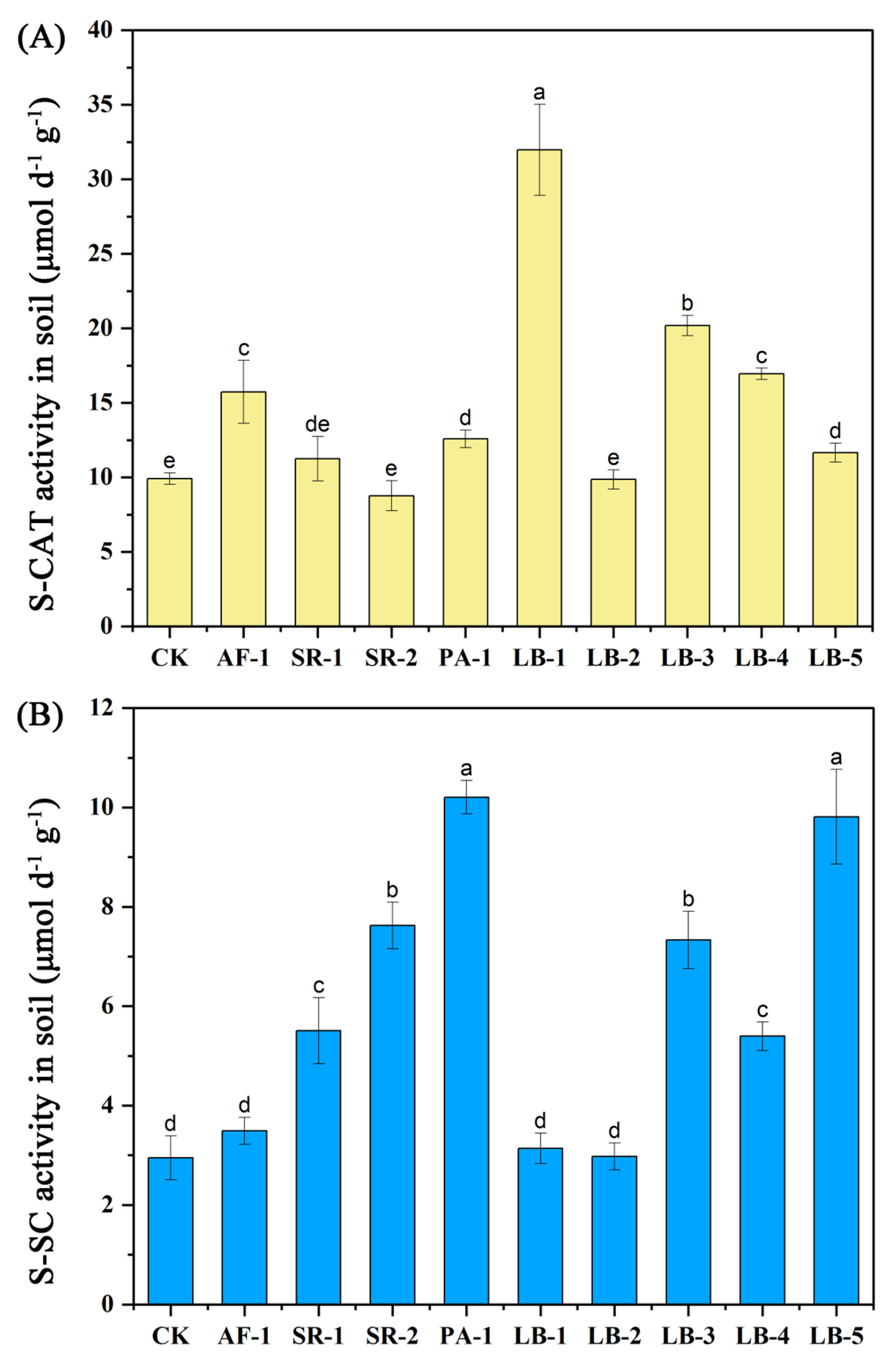
3.3. Soil Enzyme Activities
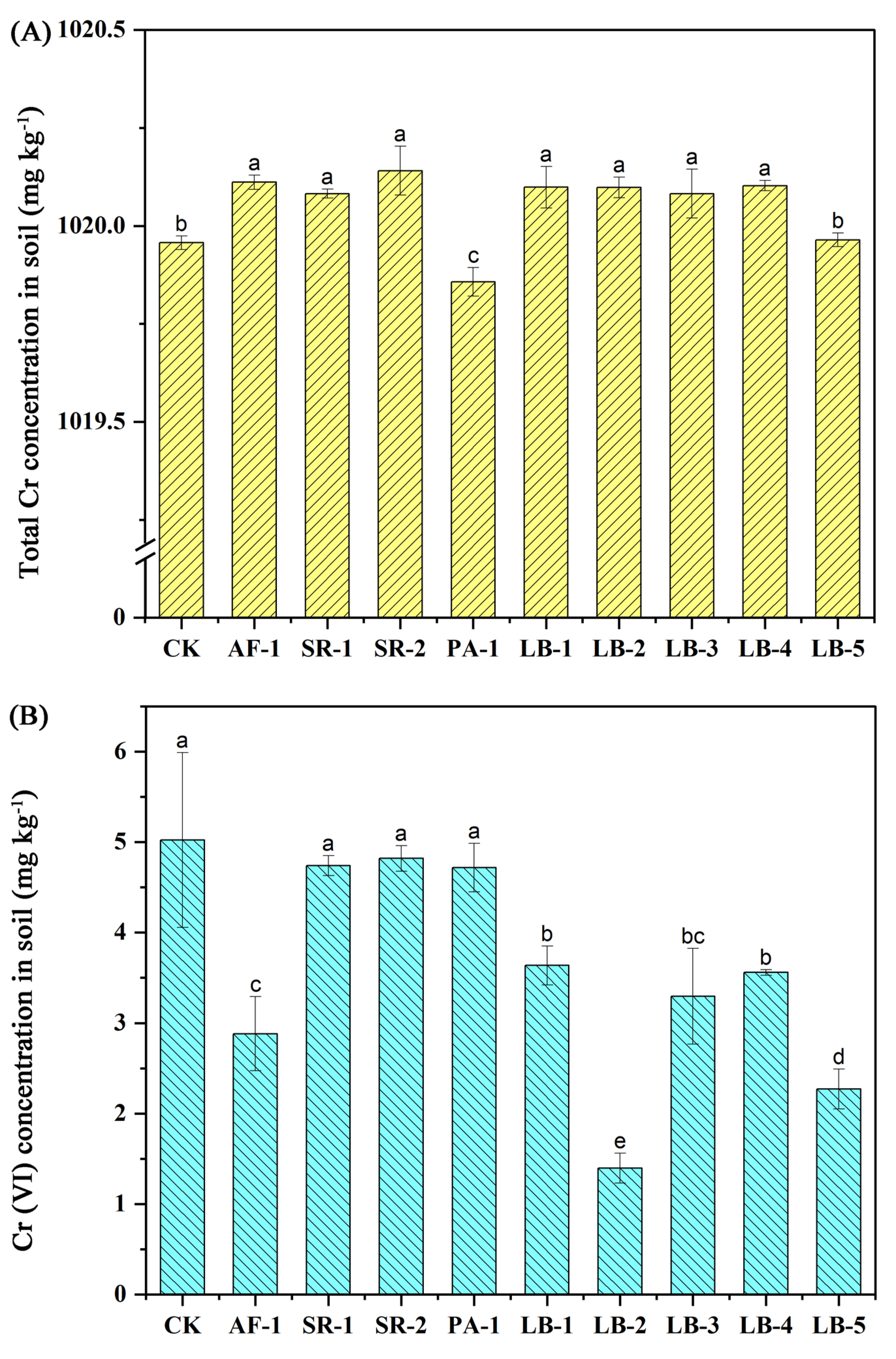
3.4. Soil Total Cr and Cr (VI) Concentration
3.5. Plant Growth and Photosynthetic Pigments
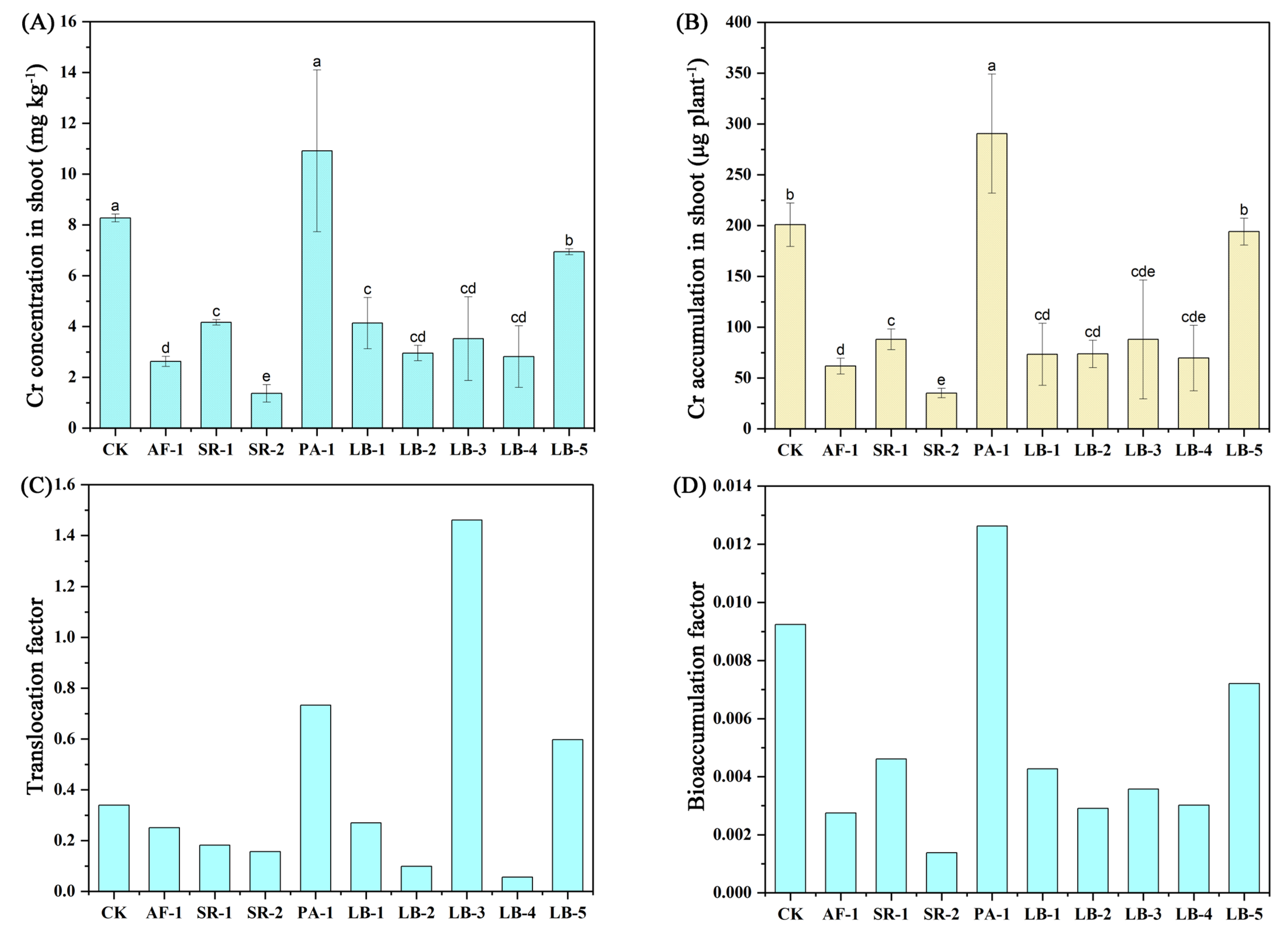
3.6. Cr Uptake and Accumulation in Plant
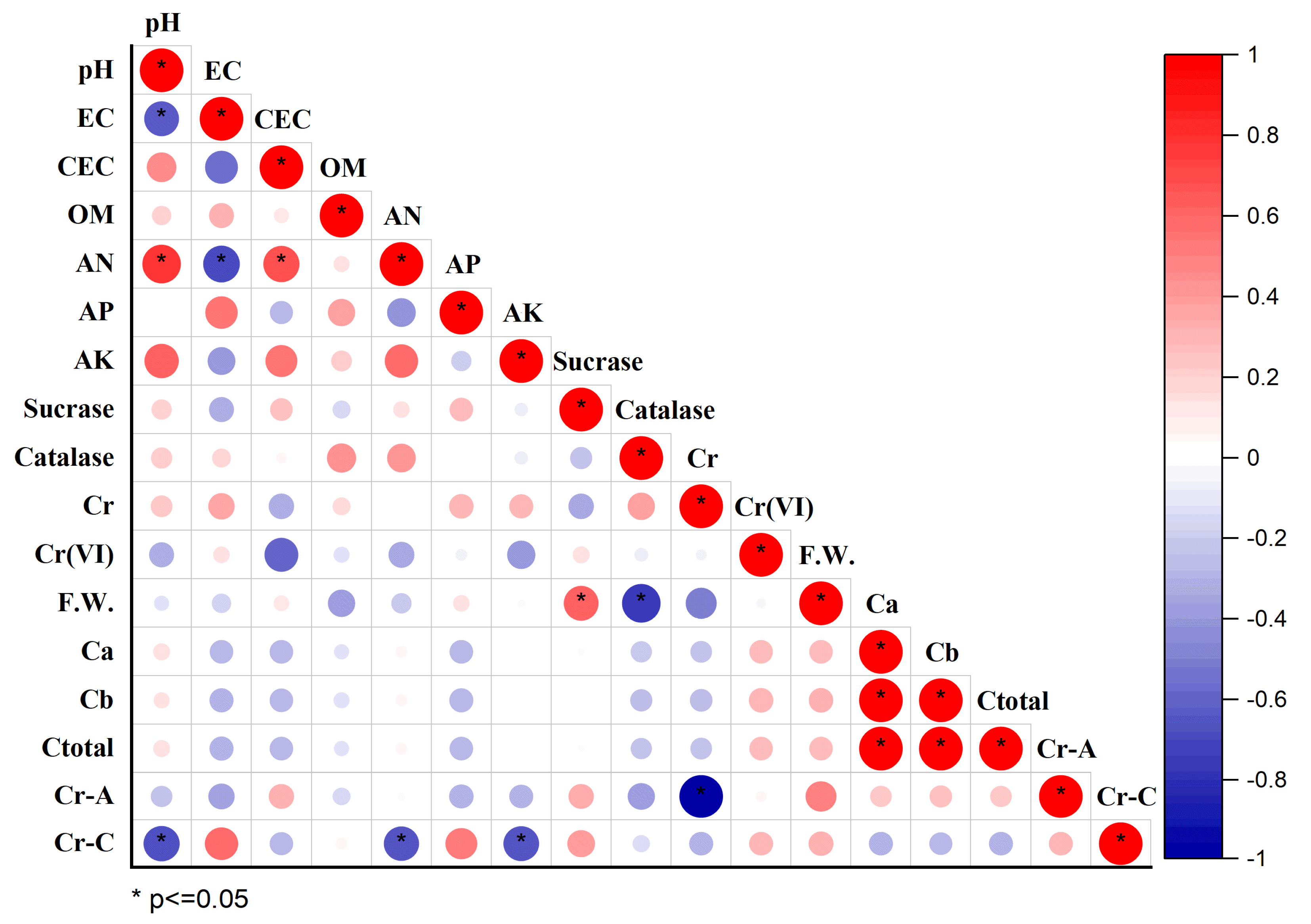
3.7. Correlation Analysis between Soil Environmental Variables and Plant Indexes
4. Discussion
4.1. Influence of Cr-Tolerant Bacteria on Soil Physicochemical Properties
4.2. Influence of Cr-Tolerant Bacteria on Soil Enzyme Activities
4.3. Influence of Cr-Tolerant Bacteria on Lettuce Growth and Photosynthetic Pigments
4.4. Influence of Cr-Tolerant Bacteria on Soil Total Cr Concentration and Soil Cr (VI) Concentration
4.5. Influence of Cr-Tolerant Bacteria on Cr Concentration in Lettuce
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kováčik, J.; Babula, P.; Hedbavny, J.; Kryštofová, O.; Provaznik, I. Physiology and methodology of Cr toxicity using alga Scenedesmus quadricauda as model object. Chemosphere 2015, 120, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Katz, S.A.; Salem, H. The Biological and Environmental Chemistry of Cr; VCH: New York, NY, USA, 1994; p. 214. [Google Scholar]
- Zhang, Q.; Amor, K.; Galer, S.J.G.; Thompson, I.; Porcelli, D. Using stable isotope fractionation factors to identify Cr (VI) reduction pathways: Metal-mineral-microbe interactions. Water Res. 2019, 151, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Arisah, F.M.; Amir, A.F.; Ramli, N.; Ariffin, H.; Maeda, T.; Hassan, M.A.; Yusoff, M.Z.M. Bacterial resistance against heavy metals in pseudomonas aeruginosa rw9 involving hexavalent Cr removal. Sustainability 2021, 13, 9797. [Google Scholar] [CrossRef]
- Yang, Q.Q.; Li, Z.Y.; Lu, X.N.; Duan, Q.N.; Huang, L.; Bi, J. A review of soil heavy metal pollution from industrial and agricultural regions in China: Pollution and risk assessment. Sci. Total Environ. 2018, 642, 690–700. [Google Scholar] [CrossRef]
- Li, L.; Shang, X.; Sun, X.; Xiao, X.; Xue, J.; Gao, Y.; Gao, H. Bioremediation potential of hexavalent Cr by a novel bacterium Stenotrophomonas acidaminiphila 4-1. Environ. Technol. Innov. 2021, 22, 101409. [Google Scholar] [CrossRef]
- Anderson, R.A. Cr as an essential nutrient for humans. Regul. Toxicol. Pharmacol. 1997, 26, S35–S41. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Bharagava, R.N. Toxic and genotoxic effects of hexavalent Cr in environment and its bioremediation strategies. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2016, 34, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, M.; Tian, T.; Lin, S.; Xu, P.; Zhou, L.; Dai, C.; Hao, Q.; Wu, Y.; Zhai, Z.; et al. The effect of hexavalent Cr on the incidence and mortality of human cancers: A meta-analysis based on published epidemiological cohort studies. Front. Oncol. 2019, 9, 103389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA. Vegetables 2018 Summary; USDA: Syracuse, NY, USA, 2019.
- Vijayaraj, A.S.; Mohandass, C.; Joshi, D.; Rajput, N. Effective bioremediation and toxicity assessment of tannery wastewaters treated with indigenous bacteria. 3 Biotech 2018, 8, 428. [Google Scholar] [CrossRef]
- Hu, J.; Meng, D.; Liu, X.; Liang, Y.; Yin, H.; Liu, H. Response of soil fungal community to long-term Cr contamination. Trans. Nonferrous Metals Soc. China 2018, 28, 1838–1846. [Google Scholar] [CrossRef]
- Abhilash, P.C.; Tripathi, V.; Edrisi, S.A.; Dubey, R.K.; Bakshi, M.; Dubey, P.K.; Singh, H.B.; Ebbs, S.D. Sustainability of crop production from polluted lands. Energy Ecol. Environ. 2016, 1, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Sumiahadi, A.; Acar, R. A review of phytoremediation technology: Heavy metals uptake by plants. IOP Conf. Ser. Earth Environ. Sci. 2018, 142, 012023. [Google Scholar] [CrossRef]
- Fan, Y.; Li, Z.; Zhou, T.; Zhou, S.; Wu, L.; Luo, Y.; Christie, P. Phytoextraction potential of soils highly polluted with cadmium using the cadmium/zinc hyperaccumulator Sedum plumbizincicola. Int. J. Phytoremediation 2019, 21, 733–741. [Google Scholar] [CrossRef]
- Banerjee, S.; Misra, A.; Chaudhury, S.; Dam, B. A Bacillus strain TCL isolated from Jharia coalmine with remarkable stress responses, Cr reduction capability and bioremediation potential. J. Hazard. Mater. 2019, 367, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Xiao, C.; Zhou, N.; Chi, R. Speciation, toxicity, microbial remediation and phytoremediation of soil chromium contamination. Environ. Chem. Lett. 2021, 19, 1413–1431. [Google Scholar] [CrossRef]
- Zheng, Q.; Na, S.; Li, X.; Li, N.; Hai, R.; Wang, X. Acute effects of hexavalent Cr on the performance and microbial community of activated sludge in aerobiotic reactors. Environ. Technol. 2018, 40, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, A.; Winkler, W.C. Magnesium-sensing riboswitches in bacteria. RNA Biol. 2010, 7, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Bansal, N.; Coetzee, J.J.; Chirwa, E.M.N. In situ bioremediation of hexavalent Cr in presence of iron by dried sludge bacteria exposed to high Cr concentration. Ecotoxicol. Environ. Saf. 2019, 172, 281–289. [Google Scholar] [CrossRef]
- Elahi, A.; Rehman, A. Comparative behavior of two-gram positive Cr6+ resistant bacterial strains Bacillus aerius S1 and Brevibacterium iodinum S2 under hexavalent chromium Cr stress. Biotechnol. Rep. 2019, 21, e00307. [Google Scholar] [CrossRef]
- Murthy, M.K.; Khandayataray, P.; Samal, D. Chromium toxicity and its remediation by using endophytic bacteria and nanomaterials: A review. J. Environ. Manag. 2022, 318, 115620. [Google Scholar] [CrossRef]
- Zhou, S.M.; Zhang, M.; Zhang, K.K.; Yang, X.W.; He, D.X.; Yin, J.; Wang, C.Y. Effects of reduced nitrogen and suitable soil moisture on wheat (Triticum aestivum L.) rhizosphere soil microbiological, biochemical properties and yield in the Huanghuai Plain. J. Integrat. Agric. 2020, 19, 234–250. [Google Scholar] [CrossRef]
- Guo, J.K.; Lv, X.; Jia, H.L.; Hua, L.; Ren, X.H.; Muhammad, H.; Wei, T.; Ding, Y.Z. Effects of EDTA and plant growth-promoting rhizobacteria on plant growth and heavy metal uptake of hyperaccumulator Sedum alfredii Hance. J. Environ. Sci. 2020, 88, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Xiao, A.W.; Li, Z.; Li, W.C.; Ye, Z.H. The effect of plant growth-promoting rhizobacteria (PGPR) on arsenic accumulation and the growth of rice plants (Oryza sativa L.). Chemosphere 2020, 242, 125136. [Google Scholar]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, S.; Xia, Y.; Cui, J.; Wang, J.; Wang, M. Correlation analysis between differential metabolites and bacterial endophytes of Ephedra sinica in different years. Ind. Crop Prod. 2022, 175, 114250. [Google Scholar] [CrossRef]
- Deng, Z.; Cao, L. Fungal endophytes and their interactions with plants in phytoremediation: A review. Chemosphere 2017, 168, 1100–1106. [Google Scholar] [CrossRef]
- Sharma, P.; Tripathi, S.; Chandra, R. Phytoremediation potential of heavy metal accumulator plants for waste management in the pulp and paper industry. Heliyon 2020, 6, 04559. [Google Scholar] [CrossRef]
- Eljounaidi, K.; Lee, S.K.; Bae, H. Bacterial endophytes as potential biocontrol agents of vascular wilt diseases—Review and future prospects. Biol. Control 2016, 103, 62–68. [Google Scholar] [CrossRef]
- Leroy, C.; Jauneau, A.; Martinez, Y.; Cabin-Flaman, A.; Gibouin, D.; Orivel, J.; Séjalon-Delmas, N. Exploring fungus–plant N transfer in a tripartite ant–plant–fungus mutualism. Ann. Bot. 2017, 120, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.S.; Dunfield, K.E. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lin, H.; Dong, Y.; He, Y.; Liu, C. Isolation of vanadium-resistance endophytic bacterium PRE01 from Pterisvittata in stone coal smelting district and characterization for potential use in phytoremediation. J. Hazard. Mater. 2018, 341, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kaul, S.; Sharma, T.; Dhar, M.K. Omics” tools for better understanding the plant–endophyte interactions. Front. Plant Sci. 2016, 7, 955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.J.; Li, B.; Dong, Y.B.; Lin, H. Endophyte colonization enhanced cadmium phytoremediation by improving endosphere and rhizosphere microecology characteristics. J. Hazard. Mater. 2022, 434, 128829. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, Q.; Huang, L.; Xu, S.; Fu, Y.; Hou, D.; Feng, Y.; Yang, X. Cadmium phytoextraction through Brassica juncea L. under different consortia of plant growth-promoting bacteria from different ecological niches. Ecotoxicol. Environ. Saf. 2022, 237, 113541. [Google Scholar] [CrossRef]
- Xu, S.A.; Yu, C.; Wang, Q.; Liao, J.Y.; Liu, C.J.; Huang, L.K.; Liu, Q.Z.; Wen, Z.Y.; Feng, Y. Chromium contamination and health risk assessment of soil and agricultural products in a rural area in southern China. Toxics 2023, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Wang, Y.N.; Zhang, W.J.; Zhang, D.M. Isolating and high-density culturing of Nitrosomonas sp.THD-1. J. Biol. 2012, 29, 81–84. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts, polyphenol oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xue, J.; Wei, X.; Su, H.; Xu, R. Optimization of Cr6+ removal by Bacillus subtilis strain SZMC 6179J from Cr-containing soil. Indian J. Microbiol. 2020, 60, 430–435. [Google Scholar] [CrossRef]
- Hu, X.; Huang, X.; Zhao, H.; Liu, F.; Wang, L.; Zhao, X.; Gao, P.; Li, X.; Ji, P. Possibility of using modified fly ash and organic fertilizers for remediation of heavy-metal-contaminated soils. J. Clean. Prod. 2021, 284, 124713. [Google Scholar] [CrossRef]
- Hamid, Y.; Tang, L.; Hussain, B.; Usman, M.; Rehman Hashmi, M.L.U.; Bilal Khan, M.; Yang, X.; He, Z. Immobilization and sorption of Cd and Pb in contaminated stagnic anthrosols as amended with biochar and manure combined with inorganic additives. J. Environ. Manag. 2020, 257, 109999. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Khan, A.Z.; Ding, X.; Khan, S.; Ayaz, T.; Fidel, R.; Khan, M.A. Biochar efficacy for reducing heavy metals uptake by Cilantro (Coriandrum sativum) and spinach (Spinaccia oleracea) to minimize human health risk. Chemosphere 2020, 244, 125543. [Google Scholar] [CrossRef] [PubMed]
- Reijonen, I.; Hartikainen, H. Oxidation mechanisms and chemical bioavailability of Cr in agricultural soil—pH as the master variable. Appl. Geochem. 2016, 74, 84–93. [Google Scholar] [CrossRef]
- Yaashikaa, P.R.; Kumar, P.S.; Babu, V.P.M.; Durga, R.K.; Manivasagan, V.; Saranya, K.; Saravanan, A. Modelling on the removal of Cr (VI) ions from aquatic system using mixed biosorbent (Pseudomonas stutzeri and acid treated Banyan tree bark). J. Mol. Liq. 2019, 276, 362–370. [Google Scholar] [CrossRef]
- Shahid, M.; Dumat, C.; Aslam, M.; Pinelli, E. Assessment of lead speciation by organic ligands using speciation models. Chem. Speciat. Bioavailab. 2012, 24, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Shekofteh, H.; Ramazani, F.; Shirani, H. Optimal feature selection for predicting soil CEC: Comparing the hybrid of ant colony organization algorithm and adaptive network-based fuzzy system with multiple linear regression. Geoderma 2017, 298, 27–34. [Google Scholar] [CrossRef]
- Arias, M.; Pérez-Novo, C.; Osorio, F.; López, E.; Soto, B. Adsorption and desorption of copper and zinc in the surface layer of acid soils. J. Colloid Interface Sci. 2005, 288, 21–29. [Google Scholar] [CrossRef]
- Thorgersen, M.P.; Lancaster, W.A.; Ge, X.X.; Zane, G.M.; Wetmore, K.M.; Vaccaro, B.J.; Poole, F.L.; Younkin, A.D.; Deutschbauer, A.M.; Arkin, A.P.; et al. Mechanisms of Chromium and Uranium Toxicity in Pseudomonas stutzeri RCH2 Grown under Anaerobic Nitrate-Reducing Conditions. Front. Microbiol. 2017, 8, 1529. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.L.; Xue, W.J.; Zeng, G.M.; Wan, J.; Chen, G.M.; Huang, C.; Zhang, C.; Cheng, M.; Xu, P. Immobilization of Cd in river sediments by sodium alginate modified nanoscale zero-valent iron: Impact on enzyme activities and microbial community diversity. Water Res. 2016, 106, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.L.; Liu, L.S.; Zeng, G.M.; Xu, P.; Huang, C.; Deng, L.J.; Wang, R.Z.; Wan, J. The effects of rice straw biochar on indigenous microbial community and enzymes activity in heavy metal-contaminated sediment. Chemosphere 2017, 174, 545–553. [Google Scholar] [CrossRef]
- Han, B.; Song, L.; Li, H.; Song, H. Naked oats biochar-supported nanoscale zero-valent iron composite: Effects on Cd immobilization and enzyme activities in Ulansuhai River sediments of China. J. Soils Sediments 2019, 19, 2650–2662. [Google Scholar] [CrossRef]
- Yao, X.H.; Min, H.; Lu, Z.H.; Yuan, H. Influence of acetamiprid on soil enzymatic activities and respiration. Eur. J. Soil Biol. 2006, 42, 120–126. [Google Scholar] [CrossRef]
- Gianfreda, L.; Bollag, J.M. Influence of natural and anthropogenic factors on enzyme activity in soil. In Soil Biochemistry; Stotzky, G., Bollag, J.M., Eds.; Marcel Dekker: New York, NY, USA, 1996; pp. 123–193. [Google Scholar]
- Luo, A.L.; Yu, X.Y. Effects of fluazinam on soil sucrase activitiy and respiration. Chinese CN. J. Eco-Agric. 2011, 19, 902–906. [Google Scholar]
- Ozturk, S.; Kava, T.; Aslim, B.; Tan, S. Removal and reduction of Cr by Pseudomonas spp. and their correlation to rhamnolipid production. J. Hazard. Mater. 2012, 231–232, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.R.; Prajapati, S.; Das, J.; Dangar, T.K.; Das, N.; Thatoi, H. Reduction of selenite to red elemental selenium by moderately halotolerant Bacillus megaterium strains isolated from Bhitarkanika mangrove soil and characterization of reduced product. Chemosphere 2011, 84, 1231–1237. [Google Scholar] [CrossRef]
- Arshad, M.; Khan, A.H.A.; Hussain, I.; Anees, M.; Iqbal, M.; Soja, G.; Linde, C.; Yousaf, S. The reduction of Cr (VI) phytotoxicity and phytoavailability to wheat (Triticum aestivum L.) using biochar and bacteria. Appl. Soil Ecol. 2018, 114, 90–98. [Google Scholar] [CrossRef]
- Wu, Y.; Deng, P.; Jia, Y.; Bai, Q.; Xiao, H. Identification of Alcaligenes faecalis and its characteristics of tolarating for hexavalent chromium. Chin. J. Environ. Eng. 2015, 9, 3545–3550. [Google Scholar]
- Liu, Q.; Chen, Z.; Huang, L.; Mujtaba Munir, M.A.; Wu, Y.; Wang, Q.; Ma, L.; Xu, S.; Wen, Z.; Feng, Y. The effects of a combined amendment on growth, cadmium adsorption by five fruit vegetables, and soil fertility in contaminated greenhouse under rotation system. Chemosphere 2021, 285, 131499. [Google Scholar] [CrossRef]
- Christou, A.; Georgiadou, E.C.; Zissimos, A.M.; Christoforou, I.C.; Christofi, C.; Neocleous, D.; Dalias, P.; Fotopoulos, V. Uptake of hexavalent Cr by Lactuca sativa and Triticum aestivum plants and mediated effects on their performance, linked with associated public health risks. Chemosphere 2021, 267, 128912. [Google Scholar] [CrossRef]
- Agathokleous, E.; Feng, Z.; Peñuelas, J. Chlorophyll hormesis: Are chlorophylls major components of stress biology in higher plants? Sci. Total Environ. 2020, 726, 138637. [Google Scholar] [CrossRef]
- Pushkar, B.; Sevak, P.; Parab, S.; Nilkanth, N. Cr pollution and its bioremediation mechanisms in bacteria: A review. J. Environ. Manag. 2012, 287, 112279. [Google Scholar] [CrossRef] [PubMed]
- Chai, L.Y.; Huang, S.H.; Yang, Z.H.; Peng, B.; Huang, Y.; Chen, Y.H. Cr (VI) remediation by indigenous bacteria in soils contaminated by Cr-containing slag. J. Hazard. Mater. 2009, 167, 516–522. [Google Scholar] [CrossRef]
- Minari, G.D.; Saran, L.M.; Constancio, M.Y.L.; da Silva, R.C.; Rosalen, D.L.; de Melo, W.J.; Alves, L.M.C. Bioremediation potential of new cadmium, Cr, and nickel-resistant bacteria isolated from tropical agricultural soil. Ecotoxicol. Environ. Saf. 2020, 204, 111038. [Google Scholar] [CrossRef]
- Sarankumar, R.K.; Arulprakash, A.; Devanesan, S.; Selvi, A.; AlSalhi, M.S.; Rajasekar, A.; Ahamed, A. Bioreduction of hexavalent Cr by Cr resistant alkalophilic bacteria isolated from tannery effluent. J. King Saud. Univ. Sci. 2020, 32, 1969–1977. [Google Scholar] [CrossRef]
- Shameer, S.; Prasad, T. Plant growth promoting rhizobacteria for sustainable agricultural practices with special reference to biotic and abiotic stresses. Plant Growth Regul. 2018, 84, 603–615. [Google Scholar] [CrossRef]
- De Oliveira, L.M.; Gress, J.; De, J.; Rathinasabapathi, B.; Marchi, G.; Chen, Y.; Ma, L.Q. Sulfate and chromate increased each other’s uptake and translocation in As-hyperaccumulator Pteris vittata. Chemosphere 2016, 147, 36–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ao, M.; Chen, X.; Deng, T.; Sun, S.; Tang, Y.; Morel, J.L.; Wang, S. Cr biogeochemical behaviour in soil-plant systems and remediation strategies: A critical review. J. Hazard. Mater. 2022, 424, 1272. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Wang, X.Y.; Yao, L.G.; Chen, Z.J. Lettuce-derived rhizosphere polyamine-producing bacteria and their potential to reduce Cd and Pb accumulation in lettuce (Lactuca sativa L.). Environ. Exp. Bot. 2020, 178, 104161. [Google Scholar] [CrossRef]
- Song, Y. Study on Remediation of Cr-Contaminated Soil by Microbial Agents and Reduction Mechanism. Master’s Thesis, Inner Mongolia University of Science and Technology, Baotou, China, 2020. [Google Scholar]
- Tirry, N.; Kouchou, A.; El Omari, B.; Ferioun, M.; El Ghachtouli, N. Improved Cr tolerance of Medicago sativa by plant growth-promoting rhizobacteria (PGPR). J. Genet. Eng. Biotechnol. 2021, 19, 149. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, N.; Vishwakarma, K.; Singh, J.; Mishra, M.; Kumar, V.; Rani, R.; Sharma, S. Tolerance and reduction of Cr (VI) by Bacillus sp. MNU16 isolated from contaminated coal mining soil. Front. Plant Sci. 2017, 8, 778. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Bacteria | Species | Scientific Name | Source Area | Host Plant | Separation Site | MIC * (mg kg−1) | Cr (VI) * Removal Rate |
|---|---|---|---|---|---|---|---|
| AF-1 | Alcaligenes | Alcaligenes faecalis | Xinchang | maize | Rhizosphere soil | 800 | 99.8% |
| SR-1 | Serratia | Serratia sp. | Xinchang | rice | leaf | 400 | 99.6% |
| SR-2 | Serratia | Serratia nematodiphila | Yuyao | rice | root | 400 | 71.1% |
| PA-1 | Pseudomonas | Pseudomonas aeruginosa | Yuyao | maize | leaf | 400 | 99.5% |
| LB-1 | Lysinebacillus | Lysinibacillus sp. Strain SePC-36 | Xinchang | rice | root | 800 | 77.0% |
| LB-2 | Lysinebacillus | Lysinibacillus mangiferihumi strain WK63 | Yuyao | rice | leaf | 800 | 99.8% |
| LB-3 | Lysinebacillus | Lysinibacillus sphaericus strain HBUM07034 | Yuyao | rice | root | 800 | 99.8% |
| LB-4 | Lysinebacillus | Lysinibacillus magniferhumi strain M-GX18 | Yuyao | rice | leaf | 800 | 99.9% |
| LB-5 | Lysinebacillus | Lysinibacillus sp. Strain M-3 | Yuyao | rice | root | 800 | 99.8% |
| Item | pH | Electric Conductivity (ds cm−1) | Cation Exchange Capacity (cmol kg−1) | Organic Matter (g kg−1) | Available Nitrogen (mg kg−1) | Available Phosphorus (mg kg−1) | Available Potassium (mg kg−1) |
|---|---|---|---|---|---|---|---|
| CK | 4.78 ± 0.05 d | 1.08 ± 0.04 a | 8.98 ± 0.08 f | 29.06 ± 0.47 b | 114.02 ± 2.49 d | 164.98 ± 3.75 b | 151.24 ± 1.60 c |
| AF-1 | 5.00 ± 0.03 bc | 1.05 ± 0.07 ab | 9.13 ± 0.06 e | 29.05 ± 0.28 b | 160.43 ± 1.56 b | 165.74 ± 5.38 b | 165.77 ± 4.17 b |
| SR-1 | 5.03 ± 0.05 b | 1.05 ± 0.04 ab | 9.51 ± 0.03 d | 30.07 ± 0.44 b | 156.84 ± 8.16 b | 167.80 ± 6.50 b | 175.38 ± 3.83 ab |
| SR-2 | 5.12 ± 0.03 a | 1.03 ± 0.01 b | 9.55 ± 0.08 d | 29.43 ± 1.32 b | 169.82 ± 18.50 ab | 164.54 ± 6.92 b | 170.74 ± 4.61 ab |
| PA-1 | 4.96 ± 0.02 c | 1.08 ± 0.02 a | 8.97 ± 0.20 ef | 30.54 ± 0.68 b | 129.21 ± 4.12 c | 183.09 ± 2.90 a | 158.09 ± 1.54 b |
| LB-1 | 5.05 ± 0.03 b | 1.07 ± 0.05 ab | 9.61 ± 0.11 cd | 31.84 ± 0.28 a | 173.65 ± 18.61 ab | 170.10 ± 4.88 b | 163.09 ± 3.22 b |
| LB-2 | 5.05 ± 0.05 ab | 1.07 ± 0.05 ab | 9.87 ± 0.32 b–d | 30.74 ± 1.93 ab | 145.88 ± 14.90 bc | 175.34 ± 12.62 ab | 179.10 ± 3.45 ab |
| LB-3 | 5.07 ± 0.03 ab | 1.05 ± 0.04 ab | 9.77 ± 0.08 c | 28.60 ± 1.10 b | 166.07 ± 4.55 ab | 169.92 ± 6.46 b | 165.04 ± 5.19 b |
| LB-4 | 5.01 ± 0.02 b | 1.06 ± 0.08 ab | 10.02 ± 0.16 b | 30.29 ± 1.07 b | 178.39 ± 8.24 a | 162.74 ±2.23 b | 183.29 ± 5.95 a |
| LB-5 | 5.01 ± 0.0.3 b | 1.04 ± 0.05 ab | 10.63 ± 0.10 a | 29.73 ± 2.04 ab | 174.71 ± 7.25 a | 166.46 ±5.71 b | 166.54 ± 4.15 b |
| Treatments | Fresh Weight (g/plant) | Chlorophyll a (mg kg−1) | Chlorophyll b (mg kg−1) | Total Chlorophyll (mg kg−1) |
|---|---|---|---|---|
| CK | 24.27 ± 2.51 a | 428.73 ± 46.92 c | 139.27 ± 12.21 cd | 568.00 ± 59.13 c |
| AF-1 | 23.55 ± 3.08 ab | 577.98 ± 96.77 b | 186.33 ± 14.57 b | 764.31 ± 111.34 b |
| SR-1 | 21.10 ± 1.96 ab | 261.94 ± 38.85 e | 91.71 ± 13.84 de | 353.66 ± 52.69 d |
| SR-2 | 26.78 ± 4.55 a | 798.81 ± 76.50 a | 253.44 ± 15.64 a | 1052.25 ± 92.14 a |
| PA-1 | 27.55 ± 3.64 a | 414.04 ± 52.32 cd | 138.06 ± 18.96 cd | 552.11 ± 71.28 c |
| LB-1 | 16.90 ± 4.23 b | 389.06 ± 42.16 cd | 123.05 ± 21.20 d | 512.11 ± 63.36 c |
| LB-2 | 24.76 ± 2.06 a | 345.69 ± 24.55 d | 108.49 ± 13.99 d | 454.18 ± 38.54 cd |
| LB-3 | 22.72 ± 4.96 ab | 215.46 ± 61.47 e | 70.35 ± 16.48 e | 285.81 ± 77.95 d |
| LB-4 | 24.19 ± 6.70 ab | 446.63 ± 48.96 c | 146.42 ± 11.10 c | 593.05 ± 60.06 bc |
| LB-5 | 27.92 ± 1.43 a | 316.34 ± 52.00 d | 109.07 ± 14.66 d | 425.41 ± 66.66 cd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Z.; Liu, Q.; Yu, C.; Huang, L.; Liu, Y.; Xu, S.; Li, Z.; Liu, C.; Feng, Y. The Difference between Rhizosphere and Endophytic Bacteria on the Safe Cultivation of Lettuce in Cr-Contaminated Farmland. Toxics 2023, 11, 371. https://doi.org/10.3390/toxics11040371
Wen Z, Liu Q, Yu C, Huang L, Liu Y, Xu S, Li Z, Liu C, Feng Y. The Difference between Rhizosphere and Endophytic Bacteria on the Safe Cultivation of Lettuce in Cr-Contaminated Farmland. Toxics. 2023; 11(4):371. https://doi.org/10.3390/toxics11040371
Chicago/Turabian StyleWen, Zheyu, Qizhen Liu, Chao Yu, Lukuan Huang, Yaru Liu, Shun’an Xu, Zhesi Li, Chanjuan Liu, and Ying Feng. 2023. "The Difference between Rhizosphere and Endophytic Bacteria on the Safe Cultivation of Lettuce in Cr-Contaminated Farmland" Toxics 11, no. 4: 371. https://doi.org/10.3390/toxics11040371