
Acute Toxicity of the Dinoflagellate Amphidinium carterae on Early Life Stages of Zebrafish (Danio rerio)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture of A. carterae
2.2. Preparation of A. carterae Toxin
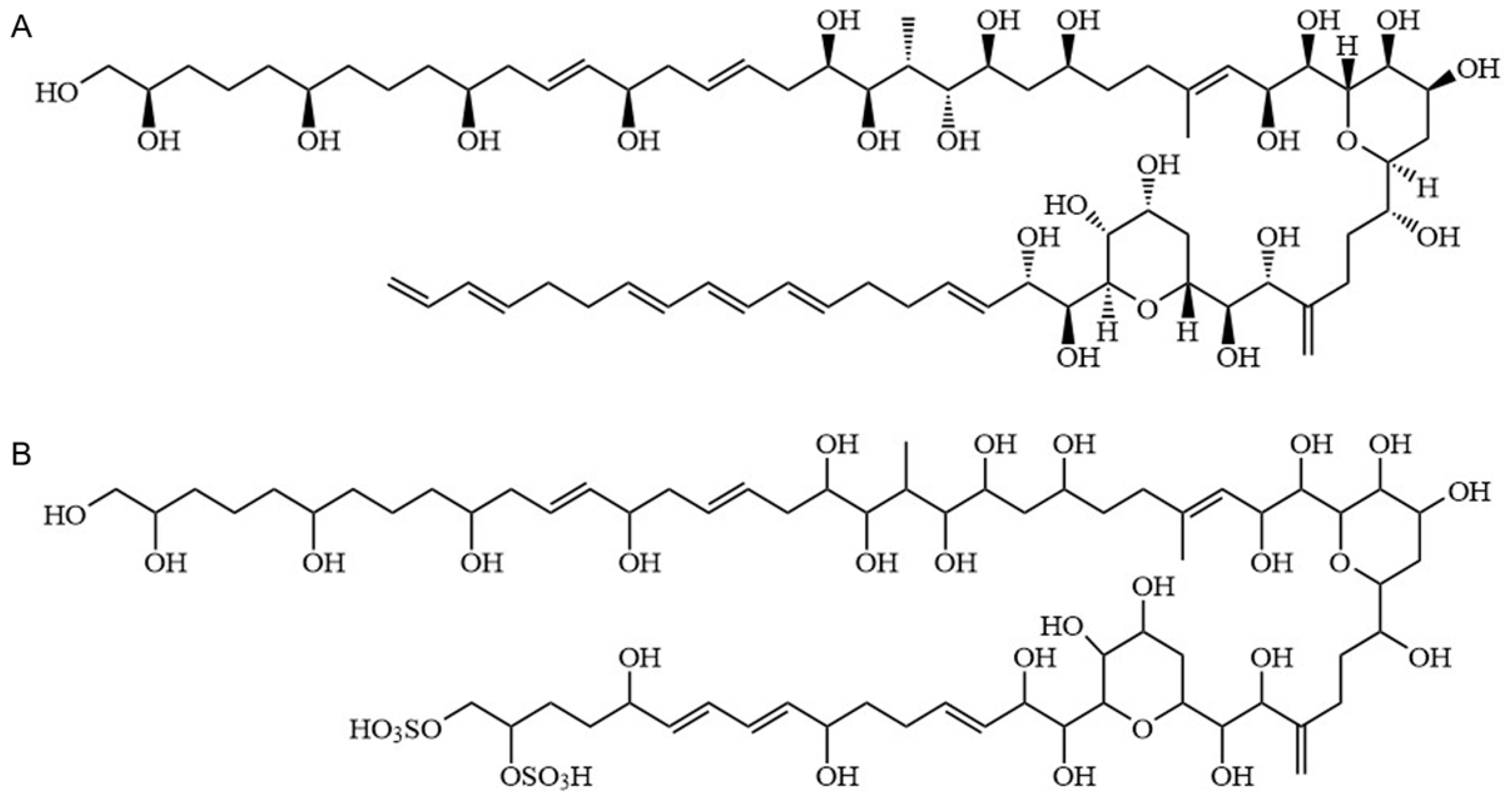
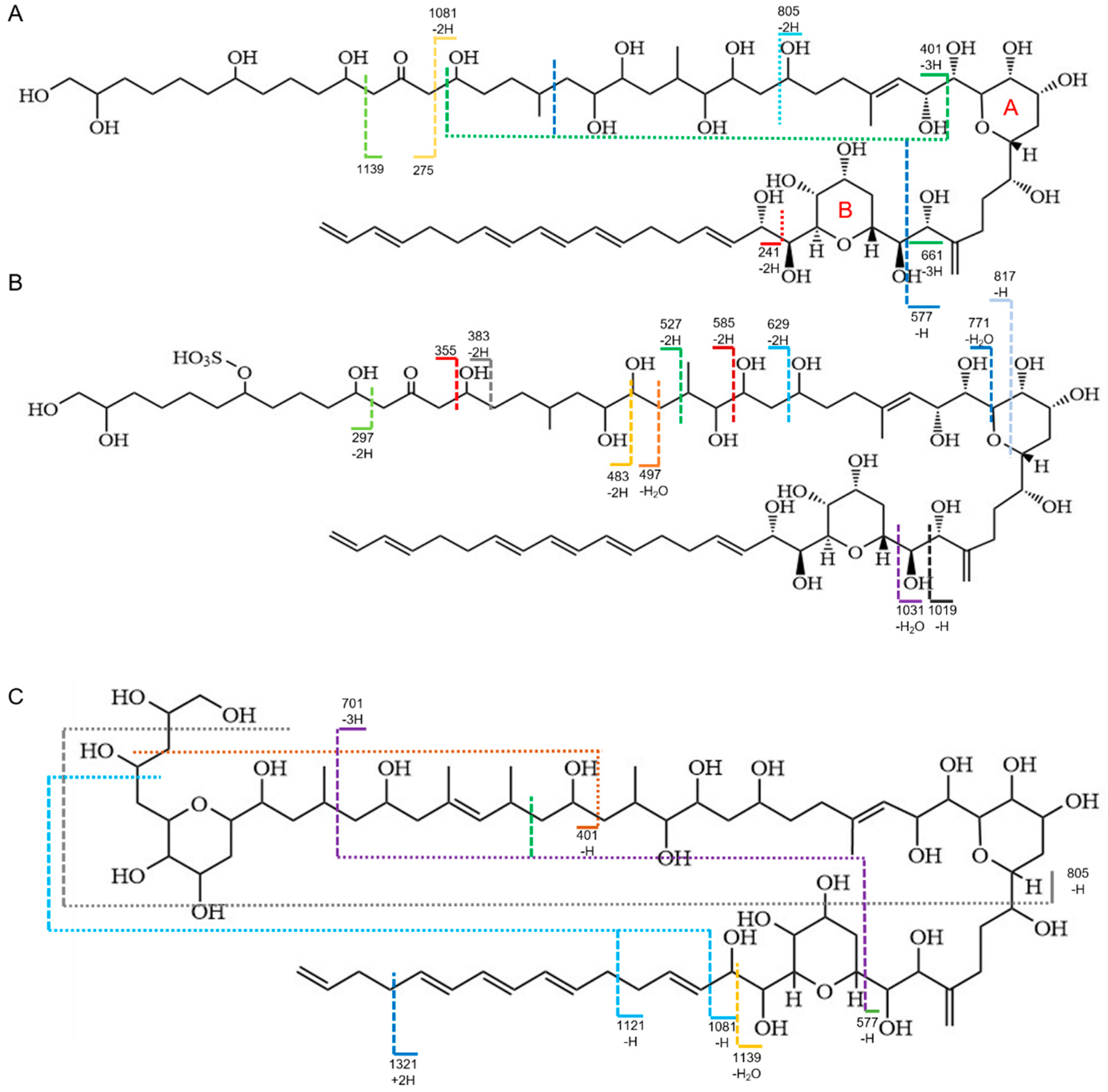
2.3. Identification of AMs
2.3.1. Conditions for Chromatographic Separation
2.3.2. Conditions for High-Resolution Mass Determination
2.4. Measurement of Hemolytic Activity
2.5. Zebrafish Maintenance and Embryo Collection
2.6. Zebrafish Embryo Acute Toxicity Tests
2.7. Statistical Analysis
3. Results and Discussion
3.1. Distribution Pattern of AMs between Extracellular and Intracellular
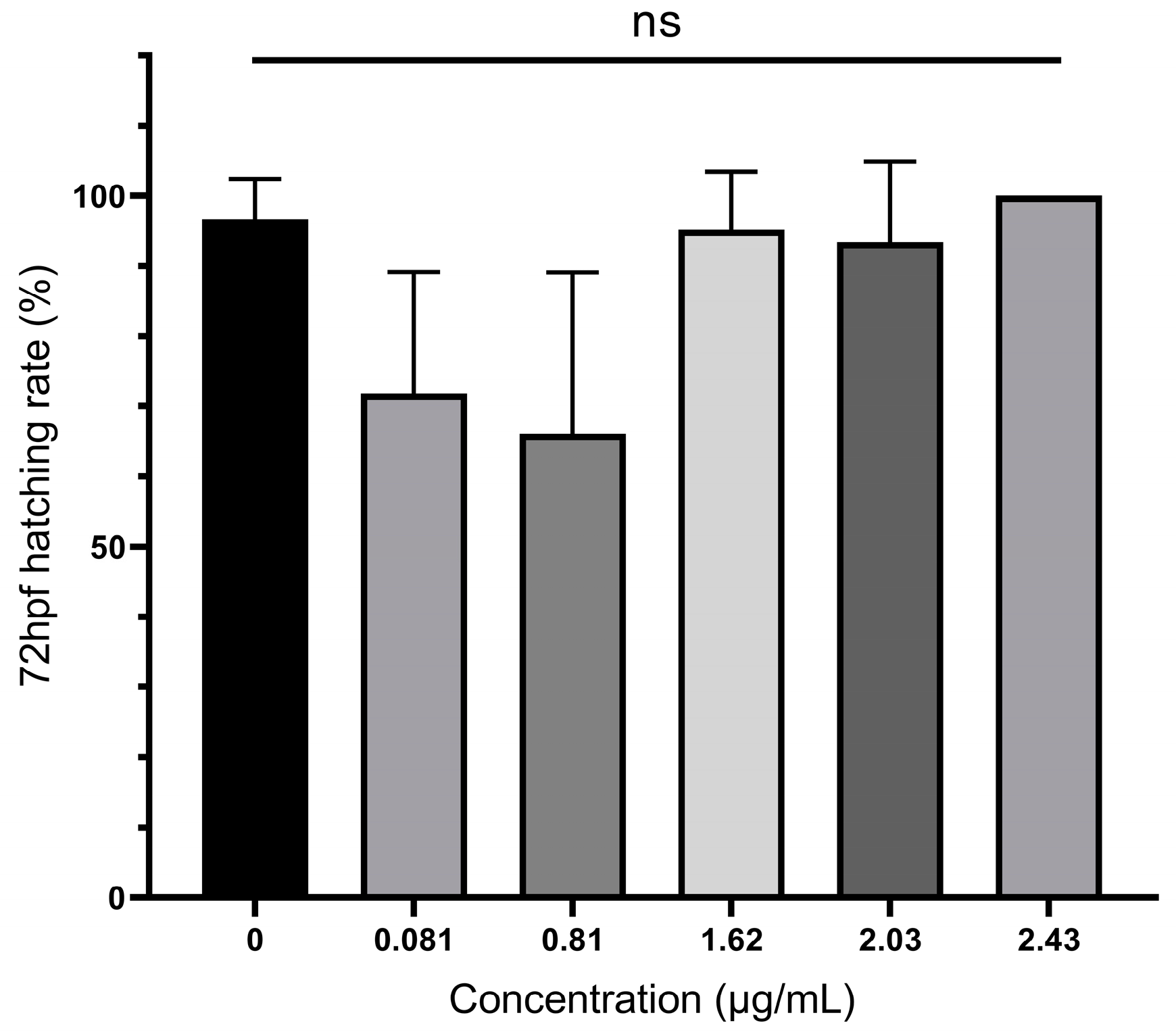
3.2. Mortality and Hatching Rates of Zebrafish Embryos Treated with A. carterae Extracellular AMs
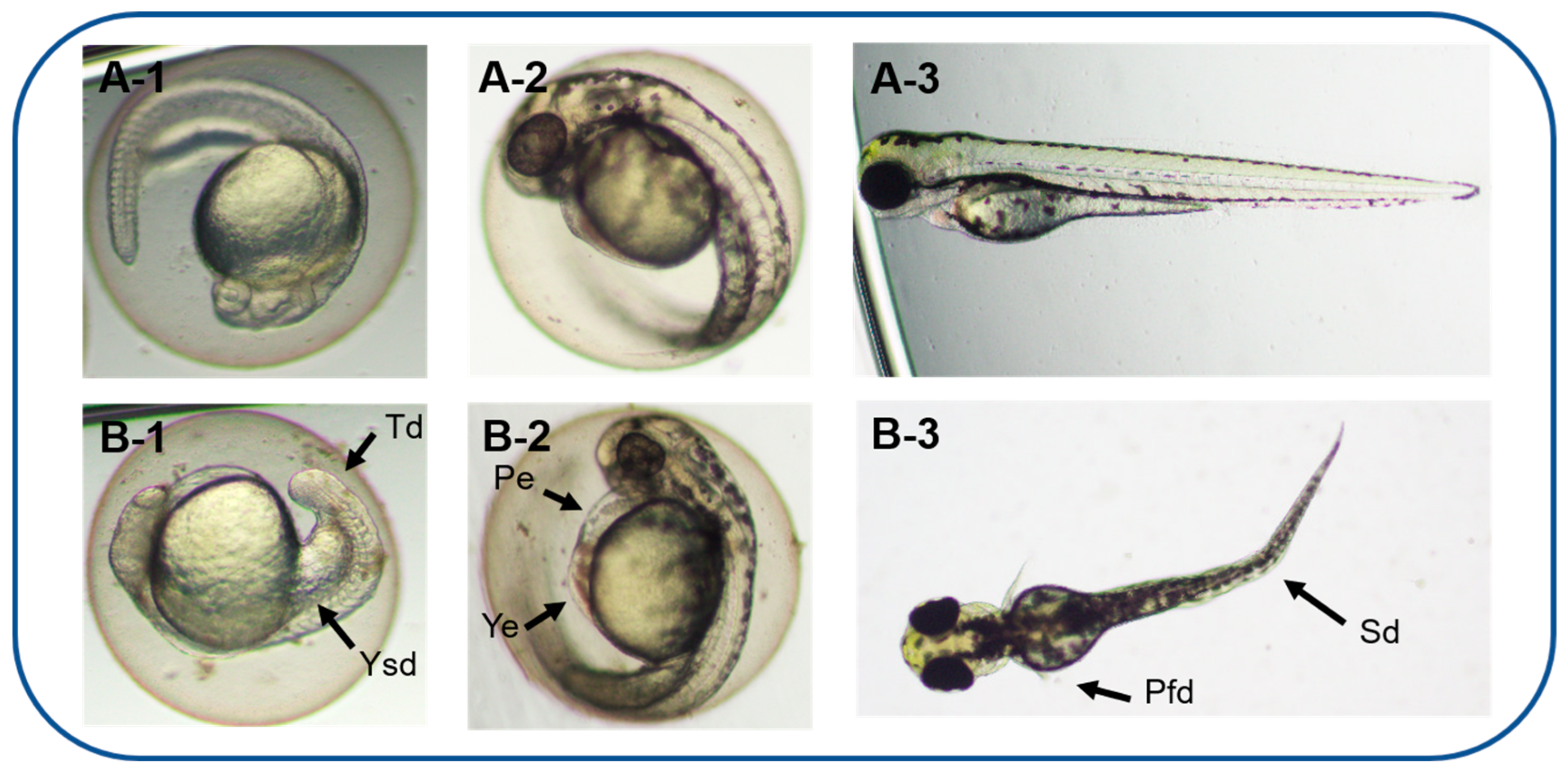
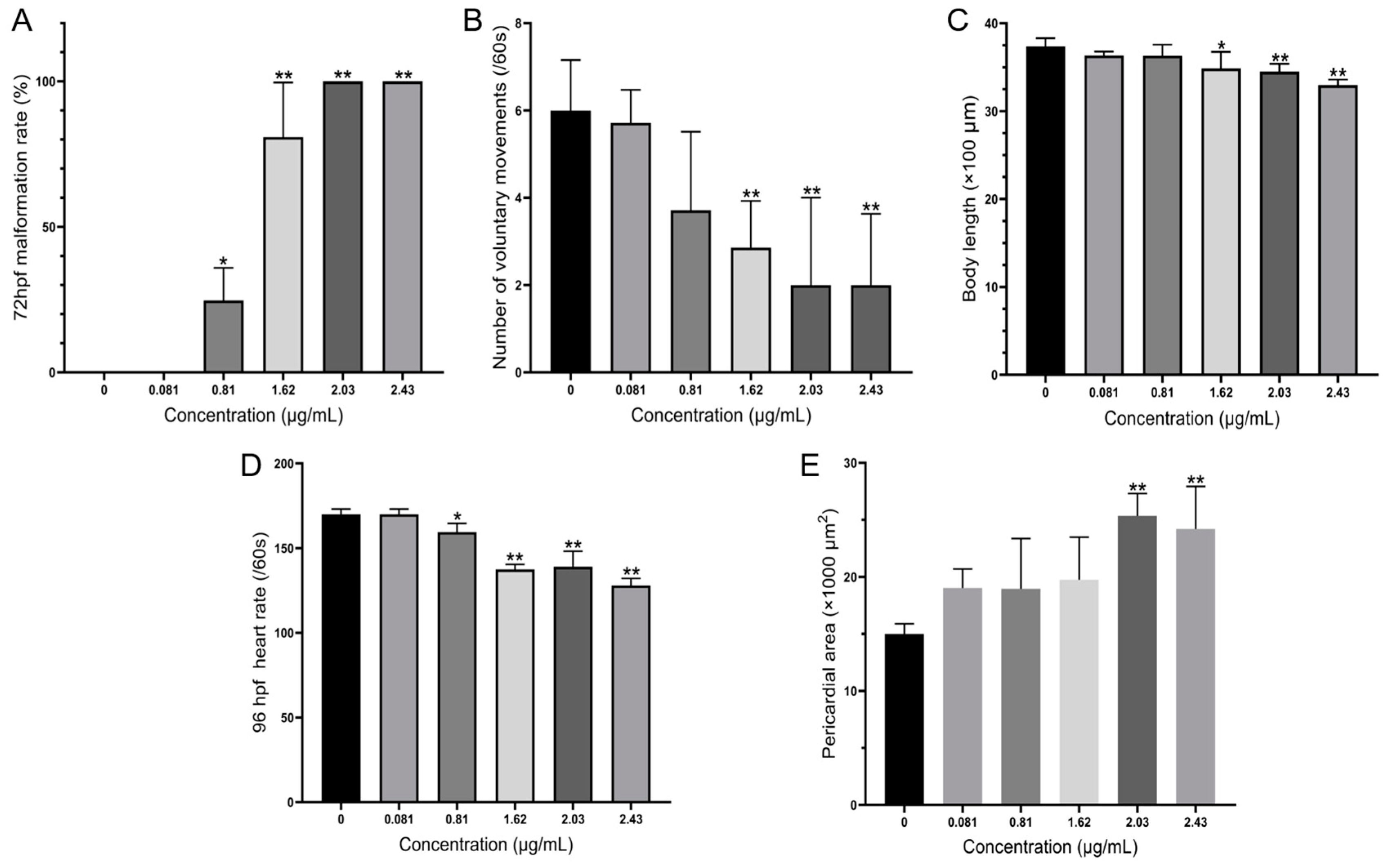
3.3. Teratogenicity of Zebrafish Embryos Treated by A. carterae Extracellular AMs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maria Duran-Riveroll, L.; Cembella, A.D.; Okolodkov, Y.B. A review on the biodiversity and biogeography of toxigenic benthic marine dinoflagellates of the coasts of Latin America. Front. Mar. Sci. 2019, 6, 148. [Google Scholar] [CrossRef] [Green Version]
- Gallardo-Rodriguez, J.; Sanchez-Miron, A.; Garcia-Camacho, F.; Lopez-Rosales, L.; Chisti, Y.; Molina-Grima, E. Bioactives from microalgal dinoflagellates. Biotechnol. Adv. 2012, 30, 1673–1684. [Google Scholar] [CrossRef] [PubMed]
- Berdalet, E.; Tester, P.A.; Chinain, M.; Fraga, S.; Lemee, R.; Litaker, W.; Penna, A.; Usup, G.; Vila, M.; Zingone, A. Harmful algal blooms in benthic systems: Recent progress and future research. Oceanography 2017, 30, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Lim, A.S.; Jeong, H.J. Benthic dinoflagellates in Korean waters. Algae 2021, 36, 91–109. [Google Scholar] [CrossRef]
- Lee, J.J.; Shpigel, M.; Freeman, S.; Zmora, O.; McLeod, S.; Bowen, S.; Pearson, M.; Szostek, A. Physiological ecology and possible control strategy of a toxic marine dinoflagellate, Amphidinium sp., from the benthos of a mariculture pond. Aquaculture 2003, 217, 351–371. [Google Scholar] [CrossRef]
- Zingone, A.; Berdalet, E.; Bienfang, P.; Enevoldsen, H.; Evans, J.; Kudela, R.; Tester, P. Harmful algae in benthic systems: A GEOHAB core research program. Cryptogam. Algol. 2012, 33, 225–230. [Google Scholar] [CrossRef]
- Usami, M.; Satake, M.; Ishida, S.; Inoue, A.; Kan, Y.; Yasumoto, T. Palytoxin analogs from the dinoflagellate Ostreopsis siamnsis. J. Am. Chem. Soc. 1995, 117, 5389–5390. [Google Scholar] [CrossRef]
- Cruz-Rivera, E.; Villareal, T.A. Macroalgal palatability and the flux of ciguatera toxins through marine food webs. Harmful Algae 2006, 5, 497–525. [Google Scholar] [CrossRef]
- Lee, T.C.H.; Fong, F.L.Y.; Ho, K.C.; Lee, F.W.F. The mechanism of diarrhetic shellfish poisoning toxin production in Prorocentrum spp.: Physiological and molecular perspectives. Toxins 2016, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.Q.; Han, X.T.; Yang, B.J.; Yu, Z.M. Analysis on allelochemicals in the cell-free Filtrates of Amphidinium carterae. Acta Ecol. Sin. 2012, 32, 1745–1754. [Google Scholar]
- Gain, G.; Peltekis, A.; Fontana, A.; Bailleul, B. Measurements of photosynthesis in mixtures reveal allelopathy between a dinoflagellate (Amphidinium carterae) and a diatom (Thalassiosira pseudonana). Biochim. Biophys. Acta-Bioenerg. 2022, 1863, 148823. [Google Scholar] [CrossRef]
- Holmes, M.J.; Venables, B.; Lewis, R.J. Critical review and conceptual and quantitative models for the transfer and depuration of ciguatoxins in fishes. Toxins 2021, 13, 515. [Google Scholar] [CrossRef] [PubMed]
- Sosa, S.; Pelin, M.; Ponti, C.; Carlin, M.; Tubaro, A. Acute toxicity by oral co-exposure to palytoxin and okadaic acid in mice. Mar. Drugs 2022, 20, 735. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.; Patterson, D.J. The benthic dinoflagellate genus Amphidinium in south-eastern Australian waters, including three new species. Eur. J. Phycol. 2002, 37, 279–298. [Google Scholar] [CrossRef]
- Jiang, L.C.; Li, Q.; Lu, S.H. Morphology and Phylogenetics study on the species of Amphidinium (Gymnodinials, Dinophyceae) from Weizhou Island, Guangxi. J. Trop. Oceanogr. 2020, 39, 106–115. [Google Scholar]
- Murray, S.A.; Garby, T.; Hoppenrath, M.; Neilan, B.A. Genetic diversity, morphological uniformity and polyketide production in dinoflagellates (Amphidinium, Dinoflagellata). PLoS ONE 2012, 7, e38253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristina Abreu, A.; Molina-Miras, A.; Aguilera-Saez, L.M.; Lopez-Rosales, L.; del Carmen Ceron-Garcia, M.; Sanchez-Miron, A.; Olmo-Garcia, L.; Carrasco-Pancorbo, A.; Garcia-Camacho, F.; Molina-Grima, E.; et al. Production of Amphidinols and Other Bioproducts of Interest by the Marine Microalga Amphidinium carterae Unraveled by Nuclear Magnetic Resonance Metabolomics Approach Coupled to Multivariate Data Analysis. J. Agric. Food Chem. 2019, 67, 9667–9682. [Google Scholar] [CrossRef]
- Ge, W.; Wang, J.Y.; Chai, C. Hemolytic activity of toxin in Amphidinium carterae Hulburt. Oceanol. Limnol. Sin. 2009, 40, 732–737. [Google Scholar]
- Morsy, N.; Houdai, T.; Konoki, K.; Matsumori, N.; Oishi, T.; Murata, M. Effects of lipid constituents on membrane-permeabilizing activity of amphidinols. Bioorganic Med. Chem. 2008, 16, 3084–3090. [Google Scholar] [CrossRef]
- Echigoya, R.; Rhodes, L.; Oshima, Y.; Satake, M. The structures of five new antifungal and hemolytic amphidinol analogs from Amphidinium carterae collected in New Zealand. Harmful Algae 2005, 4, 383–389. [Google Scholar] [CrossRef]
- Martínez, K.A.; Lauritano, C.; Druka, D.; Romano, G.; Grohmann, T.; Jaspars, M.; Martín, J.; Díaz, C.; Cautain, B.; de la Cruz, M.; et al. Amphidinol 22, a new cytotoxic and antifungal amphidinol from the dinoflagellate Amphidinium carterae. Mar. Drugs 2019, 17, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Van Wagoner, R.M.; Misner, I.; Tomas, C.; Wright, J.L.C. Structure and Biosynthesis of Amphidinol 17, a Hemolytic Compound from Amphidinium carterae. J. Nat. Prod. 2010, 73, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Cossy, J.; Tsuchiya, T.; Ferrie, L.; Reymond, S.; Kreuzer, T.; Colobert, F.; Jourdain, P.; Marko, I.E. Efficient syntheses of the polyene fragments present in amphidinols. Synlett 2007, 2007, 2286–2288. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T.; Fujita, T.; Naoki, H. Amphidinol, a polyhydroxy-polyene antifungal agent with an unprecedented structure, from a marine dinoflagellate, Amphidinium klebsii. J. Am. Chem. Soc. 1991, 113, 9859–9861. [Google Scholar] [CrossRef]
- Nuzzo, G.; Cutignano, A.; Sardo, A.; Fontana, A. Antifungal Amphidinol 18 and Its 7-Sulfate Derivative from the Marine Dinoflagellate Amphidinium carterae. J. Nat. Prod. 2014, 77, 1524–1527. [Google Scholar] [CrossRef] [PubMed]
- Morales-Amador, A.; Molina-Miras, A.; López-Rosales, L.; Sánchez-Mirón, A.; García-Camacho, F.; Souto, M.L.; Fernández, J.J. Isolation and structural elucidation of new amphidinol analogues from Amphidinium carterae cultivated in a pilot-scale photobioreactor. Mar. Drugs 2021, 19, 432. [Google Scholar] [CrossRef]
- Morsy, N.; Houdai, T.; Matsuoka, S.; Matsumori, N.; Adachi, S.; Oishi, T.; Fernandez, J.J. Structures of new amphidinols with truncated polyhydroxyl chain and their membrane-permeabilizing activities. Bioorganic Med. Chem. 2006, 14, 6548–6554. [Google Scholar] [CrossRef]
- Houdai, T.; Matsuoka, S.; Matsumori, N.; Murata, M. Membrane-permeabilizing activities of amphidinol 3, polyene-polyhydroxy antifungal from a marine dinoflagellate. Biochim. Biophys. Acta-Biomembr. 2004, 1667, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Wellkamp, M.; Garcia-Camacho, F.; Duran-Riveroll, L.M.; Tebben, J.; Tillmann, U.; Krock, B. LC-MS/MS Method Development for the Discovery and Identification of Amphidinols Produced by Amphidinium. Mar. Drugs 2020, 18, 497. [Google Scholar] [CrossRef] [PubMed]
- Souto, M.L.; Hertweck, C.; Fernandez, J.J.; Morales-Amador, A.; Garcia-Altares, M. Rapid Screening of Polyol Polyketides from Marine Dinoflagellates. Anal. Chem. 2022, 94, 14205–14213. [Google Scholar]
- Lefebvre, K.A.; Trainer, V.L.; Scholz, N.L. Morphological abnormalities and sensorimotor deficits in larval fish exposed to dissolved saxitoxin. Aquat. Toxicol. 2004, 66, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, K.A.; Elder, N.E.; Hershberger, P.K.; Trainer, V.L.; Stehr, C.M.; Scholz, N.L. Dissolved saxitoxin causes transient inhibition of sensorimotor function in larval Pacific herring (Clupea harengus pallasi). Mar. Biol. 2005, 147, 1393–1402. [Google Scholar] [CrossRef]
- Wang, X.J.; Feng, X.Q.; Zhuang, Y.; Lu, J.H.; Wang, Y.; Goncalves, R.J.; Li, X.; Lou, Y.L.; Guan, W.C. Effects of ocean acidification and solar ultraviolet radiation on physiology and toxicity of dinoflagellate Karenia mikimotoi. Harmful Algae 2019, 81, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.P.; Gantar, M.; Gibbs, P.D.L.; Schmale, M.C. The zebrafish (Danio rerio) embryo as a model system for identification and characterization of developmental toxins from marine and freshwater microalgae. Comp. Biochem. Physiol. C-Toxicol. Pharmacol. 2007, 145, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Copmans, D.; Rateb, M.; Tabudravu, J.N.; Perez-Bonilla, M.; Dirkx, N.; Vallorani, R.; Diaz, C.; Perez, D.P.J.; Smith, A.J.; Ebel, R. Zebrafish-based discovery of antiseizure compounds from the red sea: Pseurotin A(2) and azaspirofuran A. ACS Chem. Neurosci. 2018, 9, 1652–1662. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Rodriguez, M.; Lopez-Rosales, L.; Diletta, G.; del Carmen Ceron-Garcia, M.; Navarro-Lopez, E.; Jose Gallardo-Rodriguez, J.J.; Tristan, A.I.; Abreu, A.C.; Garcia-Camacho, F. The isolation of specialty compounds from Amphidinium carterae biomass by two-step solid-phase and liquid-liquid extraction. Toxins 2022, 14, 593. [Google Scholar] [CrossRef]
- Pan, L.; Chen, J.H.; Shen, H.H.; He, X.P.; Li, G.J.; Song, X.C.; Zhou, D.S.; Sun, C.J. Profiling of extracellular toxins associated with diarrhetic shellfish poison in Prorocentrum lima culture medium by high-performance liquid chromatography coupled with mass spectrometry. Toxins 2017, 9, 308. [Google Scholar] [CrossRef]
- Bao, J.Y.; Gao, X.L.; Jones, A.D. Unusual negative charge-directed fragmentation: Collision-induced dissociation of cyclopentenone oxylipins in negative ion mode. Rapid Commun. Mass Spectrom. 2014, 28, 457–464. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book; A Guide for the Laboratory Use of Zebrafish (Danio rerio); University of Oregon Press: Eugene, OR, USA, 1993. [Google Scholar]
- Mullins, M.C.; Hammerschmidt, M.; Haffter, P.; Nussleinvolhard, C. Large-scale mutagenesis in the zebrafish: In search of genes controlling development in a vertebrate. Curr. Biol. 1994, 4, 189–202. [Google Scholar] [CrossRef]
- (OECD) OfECaDP. OECD Guidelines for the Testing of Chemicals: Fish Embryo Acute Toxicity (FET) Test; OECD ilibrary: Paris, France, 2013. [Google Scholar]
- Huang, M.; Jiao, J.; Wang, J.; Xia, Z.; Zhang, Y. Exposure to acrylamide induces cardiac developmental toxicity in zebrafish during cardiogenesis. Environ. Pollut. 2018, 234, 656–666. [Google Scholar] [CrossRef]
- Ahmad, F.; Liu, X.; Zhou, Y.; Yao, H. An in vivo evaluation of acute toxicity of cobalt ferrite (CoFe2O4) nanoparticles in larval-embryo Zebrafish (Danio rerio). Aquat. Toxicol. 2015, 166, 21–28. [Google Scholar] [CrossRef]
- Wang, H.; Niu, X.Q.; Feng, X.Q.; Goncalves, R.J.; Guan, W.C. Effects of ocean acidification and phosphate limitation on physiology and toxicity of the dinoflagellate Karenia mikimotoi. Harmful Algae 2019, 87, 101621. [Google Scholar] [CrossRef] [PubMed]
- Llanos-Rivera, A.; Alvarez-Munoz, K.; Astuya-Villalon, A.; Lopez-Rosales, L.; Garcia-Camacho, F.; Sanchez-Miron, A.; Krock, B.; Jose Gallardo-Rodriguez, J. Sublethal effect of the toxic dinoflagellate Karlodinium veneficum on early life stages of zebrafish (Danio rerio). Environ. Sci. Pollut. Res. 2022, 30, 27113–27124. [Google Scholar] [CrossRef]
- Zeng, C.; Sun, H.; Xie, P.; Wang, J.H.; Zhang, G.R.; Chen, N.; Yan, W.; Li, G.Y. The role of apoptosis in MCLR-induced developmental toxicity in zebrafish embryos. Aquat. Toxicol. 2014, 149, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Parsons, A.; Lange, A.; Hutchinson, T.H.; Miyagawa, S.; Iguchi, T.; Kudoh, T.; Tyler, C.R. Molecular mechanisms and tissue targets of brominated flame retardants, BDE-47 and TBBPA, in embryo-larval life stages of zebrafish (Danio rerio). Aquat. Toxicol. 2019, 209, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.A.; Kohli, G.S.; Farrell, H.; Spiers, Z.B.; Place, A.R.; Dorantes-Aranda, J.J.; Ruszczyk, J. A fish kill associated with a bloom of Amphidinium carterae in a coastal lagoon in Sydney, Australia. Harmful Algae 2015, 49, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.J.; Kuo, C.M.; Lin, Y.C.; Chen, Y.M.; Lu, C.K. Carteraol E, a potent polyhydroxyl ichthyotoxin from the dinoflagellate Amphidinium carterae. Tetrahedron Lett. 2009, 50, 2512–2515. [Google Scholar] [CrossRef]
- Jiang, Z.B.; Liu, J.J.; Chen, J.F.; Chen, Q.Z.; Yan, X.J.; Xuan, J.L.; Zeng, J.N. Responses of summer phytoplankton community to drastic environmental changes in the Changjiang (Yangtze River) estuary during the past 50 years. Water Res. 2014, 54, 1–11. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | m/z | Formula | Calc. | mDa | ppm | |
|---|---|---|---|---|---|---|
| AM18 | 1 | 1357.8248 | C71H122O24 | 1357.8248 | 0.0 | 0.0 |
| 2 | 1139.6733 | C60H100O20 | 1139.6730 | 0.3 | 0.3 | |
| 3 | 1081.6312 | C57H94O19 | 1081.6311 | 0.1 | 0.1 | |
| 4 | 805.4382 | C43H66O14 | 805.4374 | 0.8 | 1.0 | |
| 5 | 661.3799 | C33H58O13 | 661.3799 | 0.2 | 0.3 | |
| 6 | 577.3227 | C28H50O12 | 577.3224 | 0.3 | 0.5 | |
| 7 | 561.3064 | C31H46O9 | 561.3064 | 0.0 | 0.0 | |
| 8 | 401.2540 | C21H38O7 | 401.2539 | 0.1 | 0.2 | |
| 9 | 275.1856 | C14H28O5 | 275.1858 | −0.2 | −0.7 | |
| 10 | 241.1533 | C17H22O | 241.1440 | −5.9 | −24.5 | |
| AM19 | 1 | 1437.7759 | C71H122O27S | 1437.7816 | −5.7 | −4.0 |
| 2 | 1031.5466 | C48H88O21S | 1031.5461 | 0.5 | 0.5 | |
| 3 | 1019.5461 | C47H88O21S | 1019.5461 | 0.3 | 0.3 | |
| 4 | 771.4200 | C36H68O15S | 771.4201 | −0.1 | −0.1 | |
| 5 | 629.3210 | C28H54O13S | 629.3207 | 0.3 | 0.5 | |
| 6 | 585.2947 | C26H50O12S | 585.2945 | 0.2 | 0.3 | |
| 7 | 527.2529 | C23H44O11S | 527.2526 | 0.3 | 0.6 | |
| 8 | 497.2423 | C22H42O10S | 497.2420 | 0.3 | 0.6 | |
| 9 | 483.2266 | C21H40O10S | 483.2264 | 0.2 | 0.4 | |
| 10 | 383.1382 | C15H28O9S | 383.1376 | 0.6 | 1.6 | |
| 11 | 355.1435 | C14H28O8S | 355.1427 | 0.8 | 2.3 | |
| 12 | 297.1017 | C11H22O7S | 297.1008 | 0.9 | 3.0 | |
| AM2 | 1 | 1373.8121 | C71H122O25 | 1373.8197 | −7.6 | −5.5 |
| 2 | 1321.8035 | C67H118O25 | 1321.7884 | 16.0 | 12.1 | |
| 2 | 1139.6661 | C56H100O23 | 1139.6577 | 8.4 | 7.4 | |
| 3 | 1121.6622 | C56H98O22 | 1121.6471 | 15.1 | 13.5 | |
| 4 | 1081.6318 | C53H94O22 | 1081.6158 | 16.0 | 14.8 | |
| 5 | 701.4111 | C36H62O13 | 701.4112 | −0.1 | −0.1 | |
| 6 | 577.3232 | C28H50O12 | 577.3224 | 0.8 | 1.4 | |
| 7 | 401.2544 | C21H38O7 | 401.2539 | 0.5 | 1.2 | |
| AD1 | 1 | 1487.7990 | C73H125O27SNa | 1487.7948 | 4.2 | 2.8 |
| 2 | 701.4068 | C36H62O13 | 701.4112 | −4.4 | −6.3 | |
| 3 | 661.3774 | C33H58O13 | 661.3799 | −2.5 | −3.8 | |
| 4 | 577.3196 | C28H50O12 | 577.3224 | −7.8 | −13.5 | |
| 5 | 355.1416 | C14H28O8S | 355.1427 | −1.1 | −3.1 | |
| 6 | 297.1006 | C11H22O7S | 297.1008 | −0.2 | −0.7 | |
| AD2 | 1 | 1183.6519 | C54H100O25 | 1183.6475 | 4.4 | 3.7 |
| 2 | 661.3774 | C33H58O13 | 661.3799 | −2.5 | −3.8 | |
| AD3 | 1 | 1453.7662 | C71H122O28S | 1453.7765 | −10.3 | −7.1 |
| 2 | 701.4092 | C36H62O13 | 701.4112 | −2.0 | −2.9 | |
| 3 | 661.3794 | C33H58O13 | 661.3799 | −0.5 | −0.8 | |
| 4 | 577.3221 | C28H50O12 | 577.3224 | −0.3 | −0.5 | |
| 5 | 355.1422 | C14H28O8S | 355.1427 | −0.5 | −1.4 | |
| 6 | 275.1858 | C14H28O5 | 275.1858 | −7.9 | −28.7 | |
| 7 | 297.1014 | C11H22O7S | 297.1008 | 0.6 | 2.0 | |
| AM | AM18 | AM19 | AM2 | AD1 | AD2 | AD3 |
|---|---|---|---|---|---|---|
| Extracellular | 0.0061 | 99.7250 | 0.0248 | 0.2342 | 0.0078 | 0.0021 |
| Intracellular | 0.0009 | 99.9899 | 0.0048 | 0.0008 | 0.0023 | 0.0013 |
| Time of Exposure (hpf) | Concentration (µg/mL) | LC50 (95% Confidence) | X2 (p) | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 0.081 | 0.81 | 1.62 | 2.03 | 2.43 | |||
| 12 | 0, 0, 0 | 0, 0, 1 | 2, 0, 4 | 5, 4, 2 * | 5, 3, 4 * | 6, 9, 7 ** | 2.07 (1.30–5.58) | 49.97 (p < 0.001) |
| 24 | 0, 0, 0 | 0, 0, 1 | 2, 4, 5 * | 5, 4, 2 * | 5, 3, 4 * | 6, 9, 7 ** | 1.85 (1.27–3.15) | 43.384 (p < 0.001) |
| 48 | 0, 0, 0 | 0, 0, 1 | 2, 4, 5 * | 6, 5, 3 ** | 5, 4, 5 ** | 6, 9, 7 ** | 1.539 (1.08–2.35) | 48.57 (p < 0.001) |
| 72 | 0, 0, 0 | 0, 0, 1 | 3, 4, 5 ** | 6, 5, 3 ** | 5, 4, 5 ** | 6, 9, 7 ** | 1.49 (1.04–2.28) | 47.397 (p < 0.001) |
| 96 | 0, 0, 0 | 0, 0, 1 | 3, 4, 5 * | 6, 5, 3 ** | 5, 4, 5 ** | 6, 9, 7 ** | 1.49 (1.04–2.28) | 47.397 (p < 0.001) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Yan, Z.; Chen, J.; Wang, D.; Li, K. Acute Toxicity of the Dinoflagellate Amphidinium carterae on Early Life Stages of Zebrafish (Danio rerio). Toxics 2023, 11, 370. https://doi.org/10.3390/toxics11040370
Yang X, Yan Z, Chen J, Wang D, Li K. Acute Toxicity of the Dinoflagellate Amphidinium carterae on Early Life Stages of Zebrafish (Danio rerio). Toxics. 2023; 11(4):370. https://doi.org/10.3390/toxics11040370
Chicago/Turabian StyleYang, Xiao, Zhi Yan, Jingjing Chen, Derui Wang, and Ke Li. 2023. "Acute Toxicity of the Dinoflagellate Amphidinium carterae on Early Life Stages of Zebrafish (Danio rerio)" Toxics 11, no. 4: 370. https://doi.org/10.3390/toxics11040370