
PFAS Biotransformation Pathways: A Species Comparison Study
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Metabolic Map Collection
2.2. Data Quality Assurance (QA)
- Tracking of the data source in files and folders with systematic file naming;
- Checklists/How To guidance for initial coders;
- QA accuracy checklists for reviewers of the initial coded xml and the final MetaPath pathway.
2.3. Analysis of Metabolic Maps
2.4. Automated PFAS Pipeline Profiler
3. Results
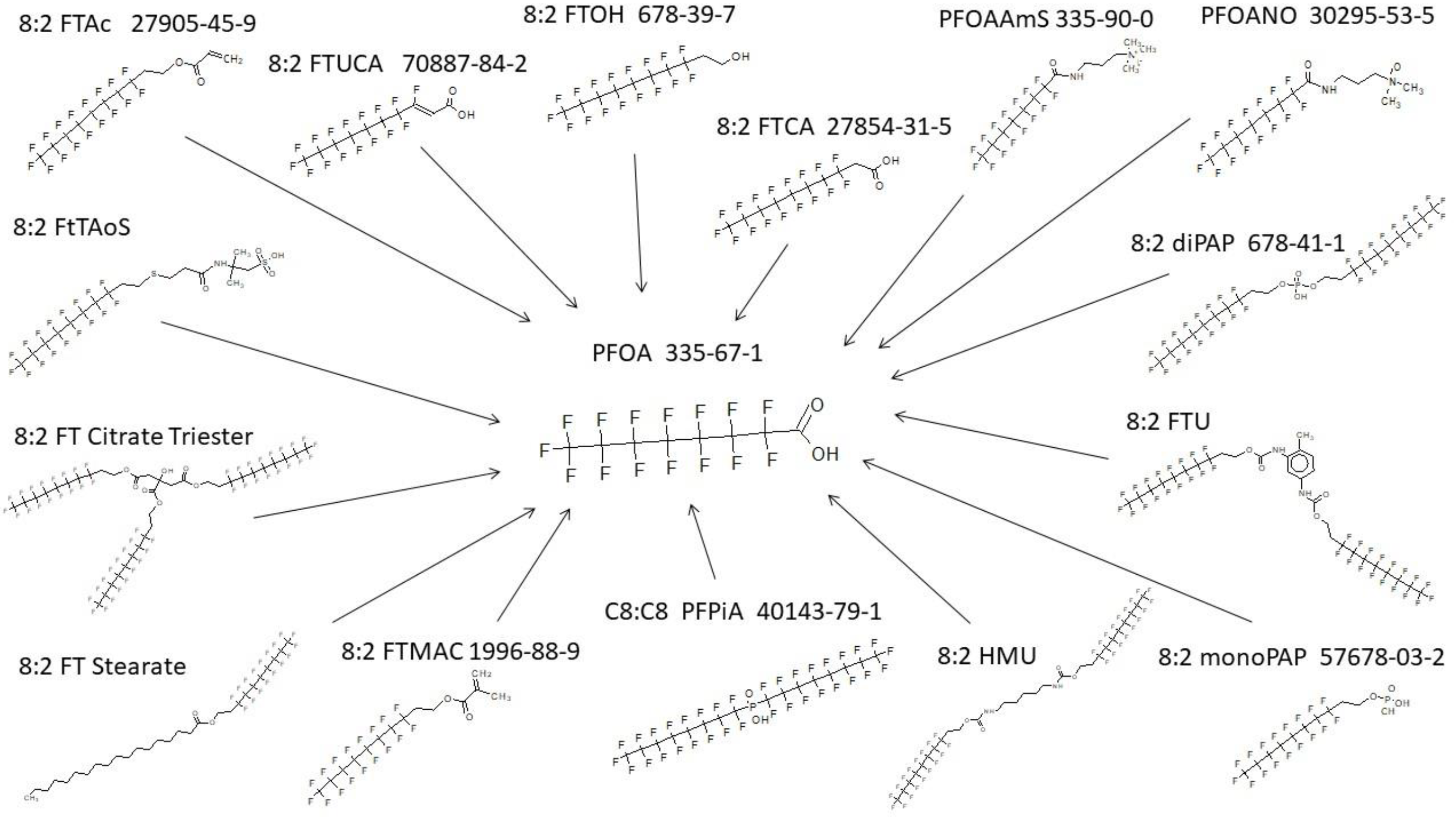
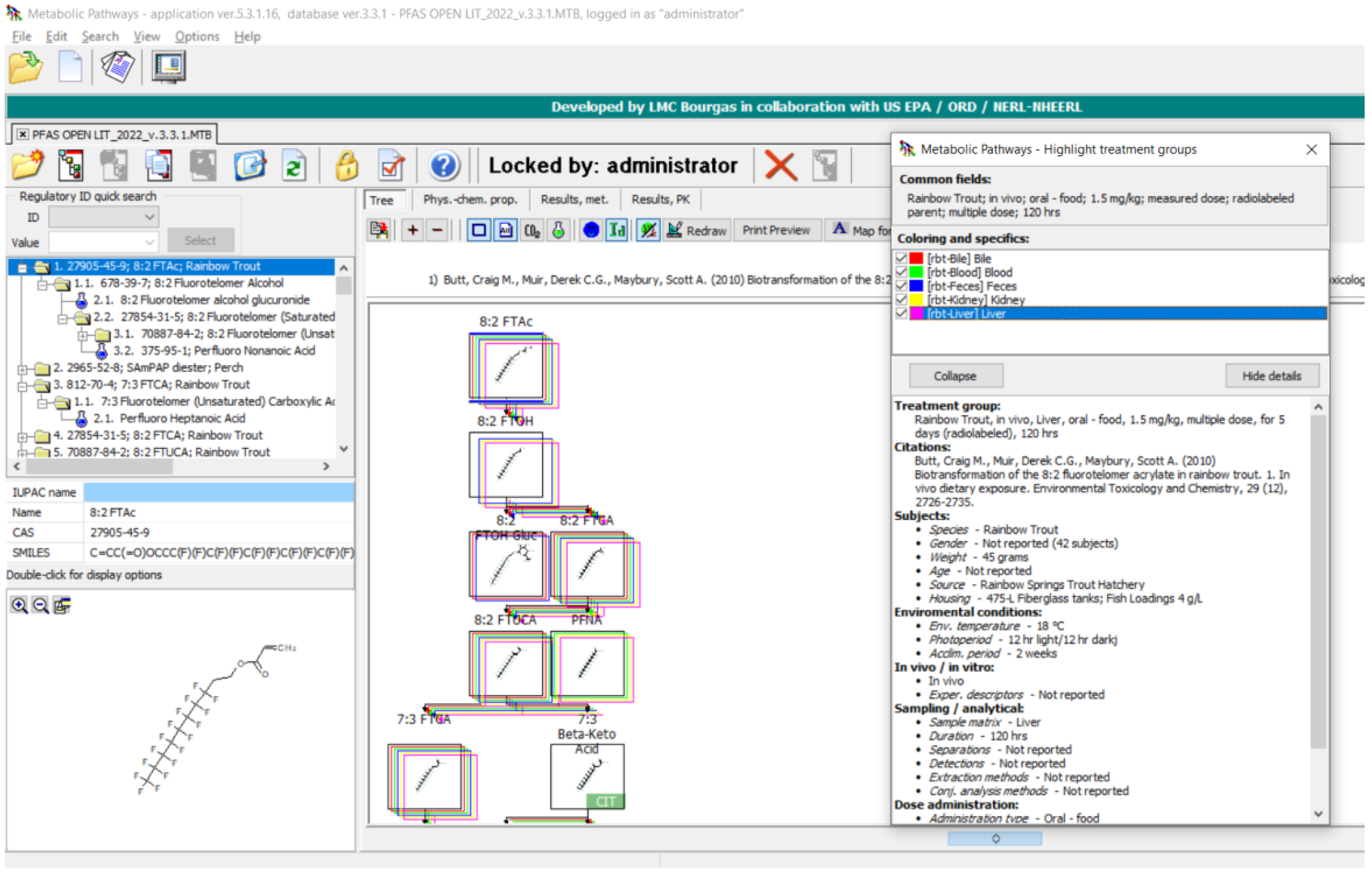
3.1. PFAS Biotransformation Database
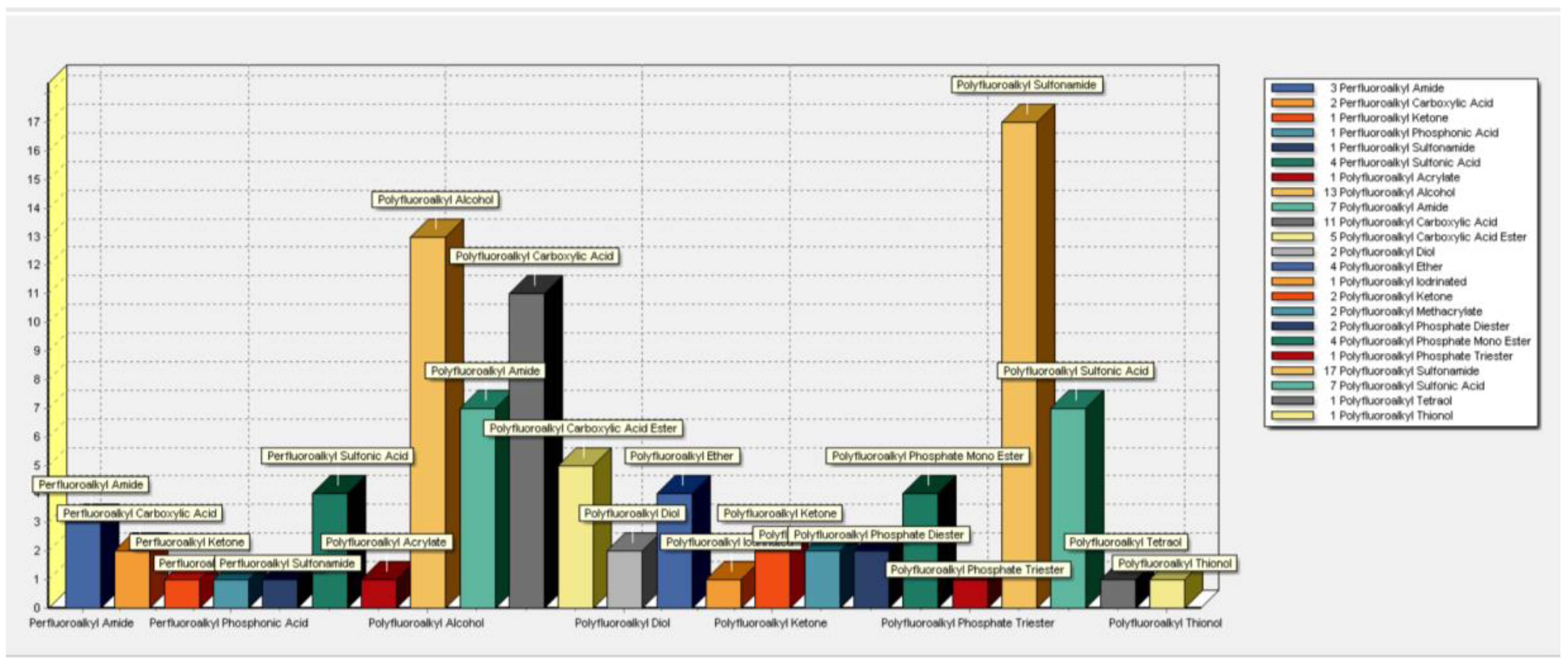
3.2. Terminal Product(s) Observed in PFAS Pathways
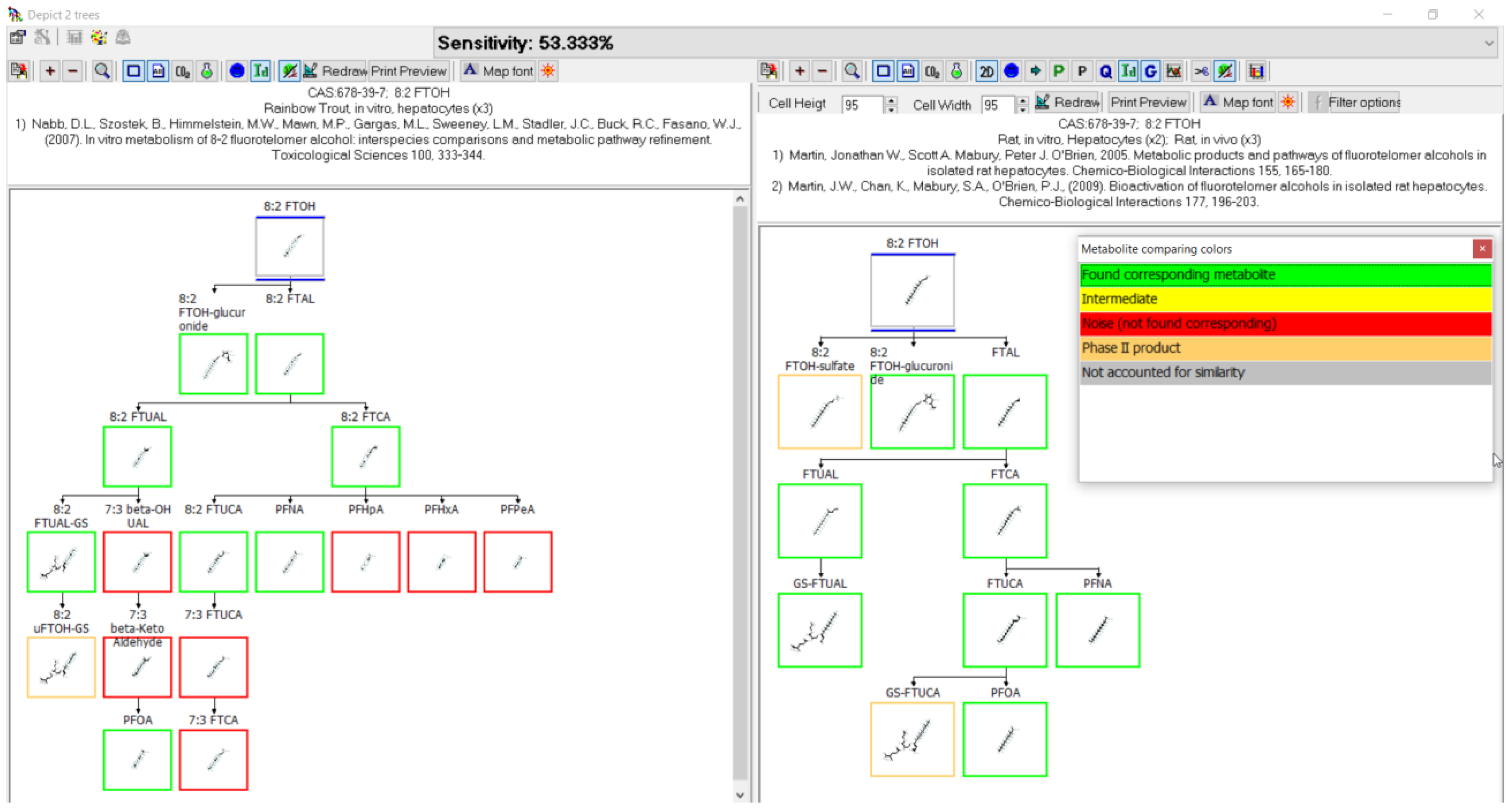
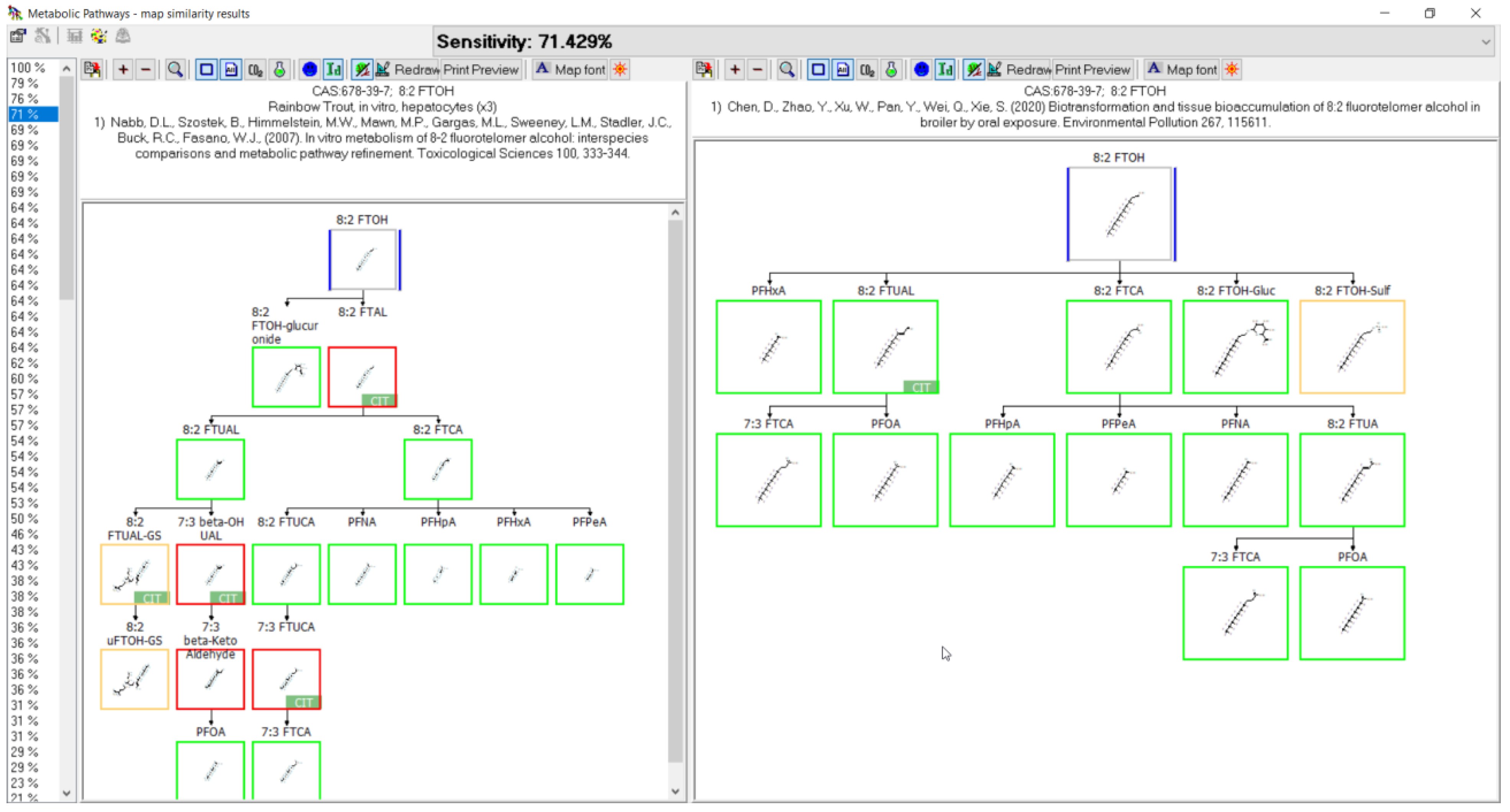
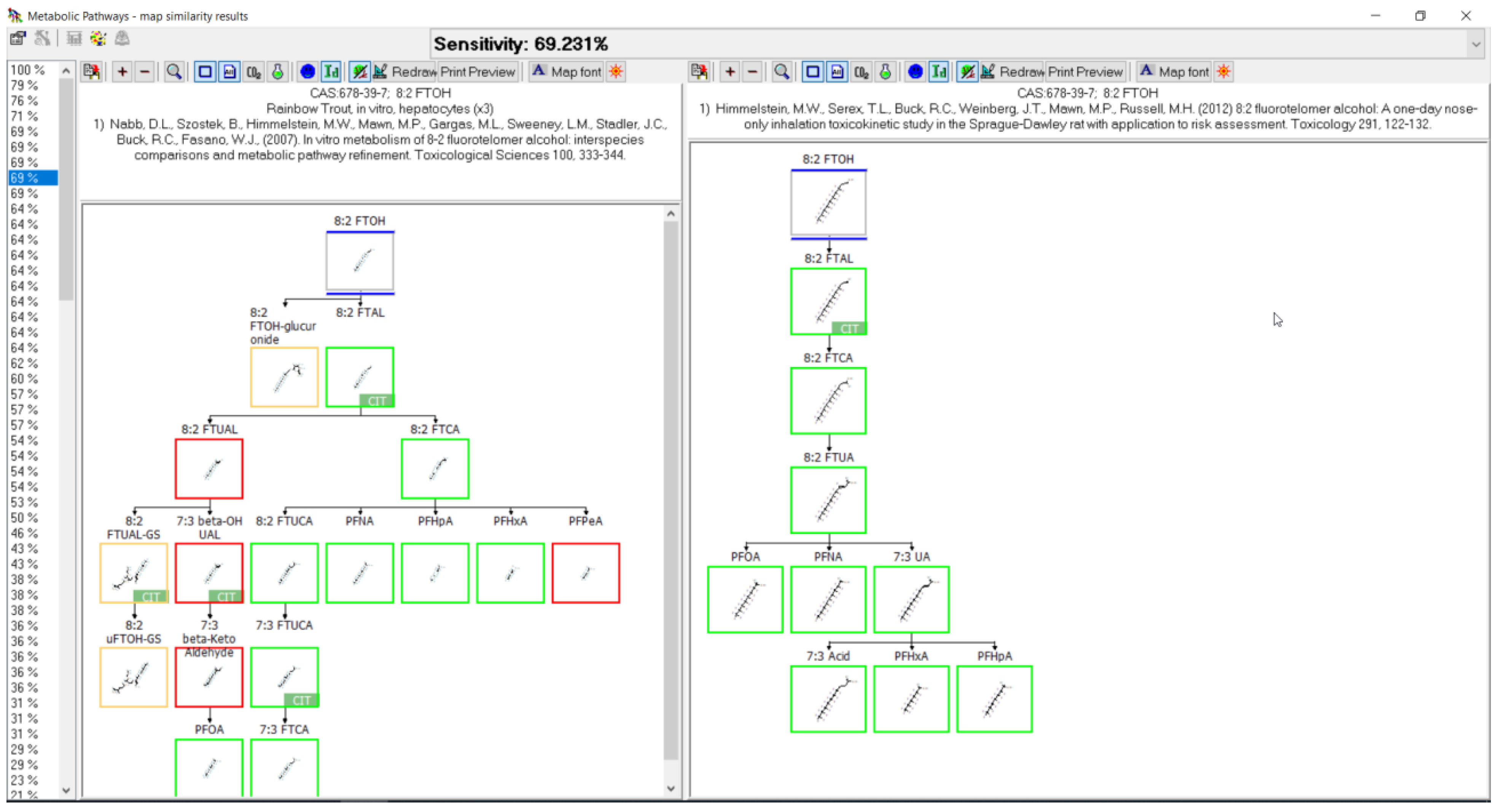
3.3. Evaluation of Metabolic Maps
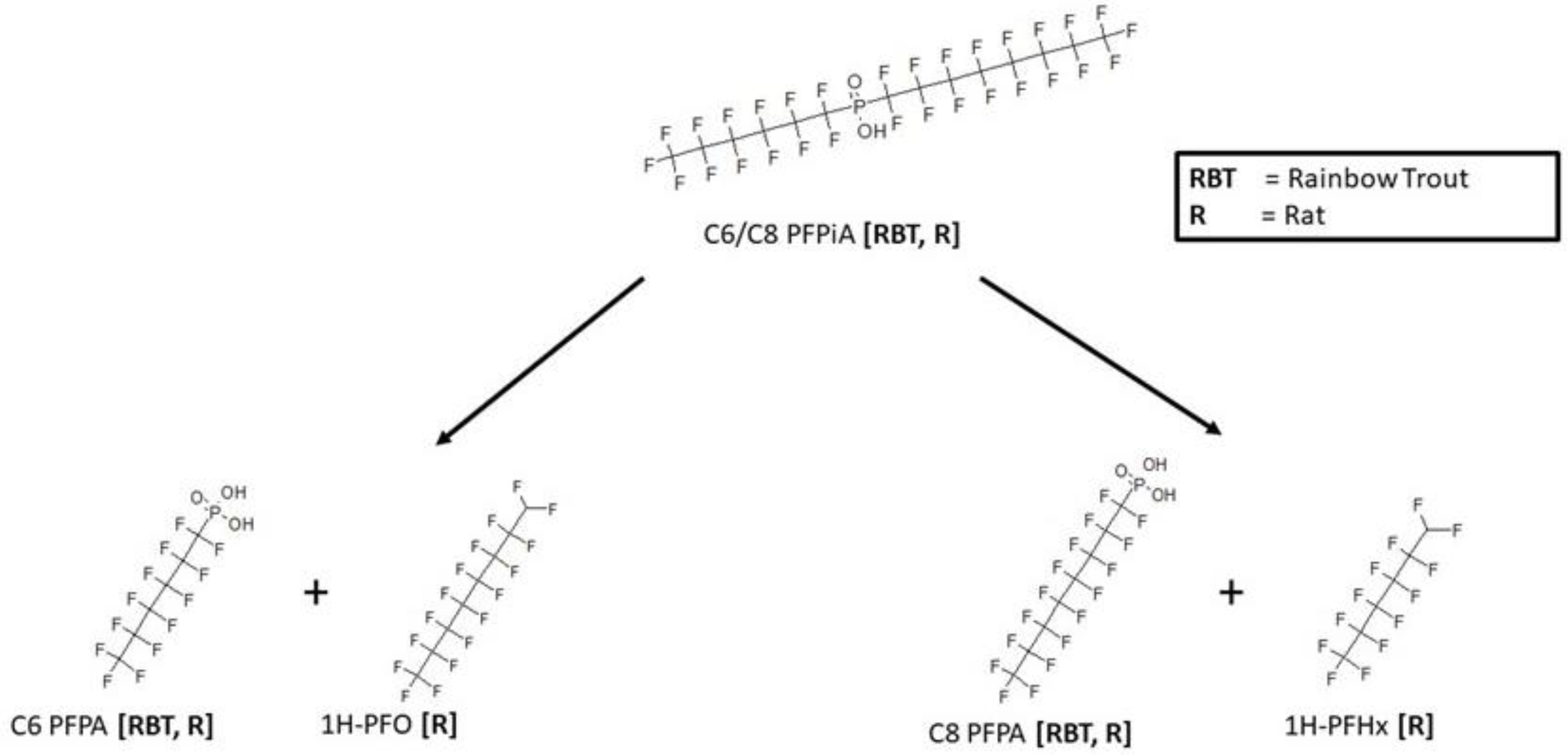
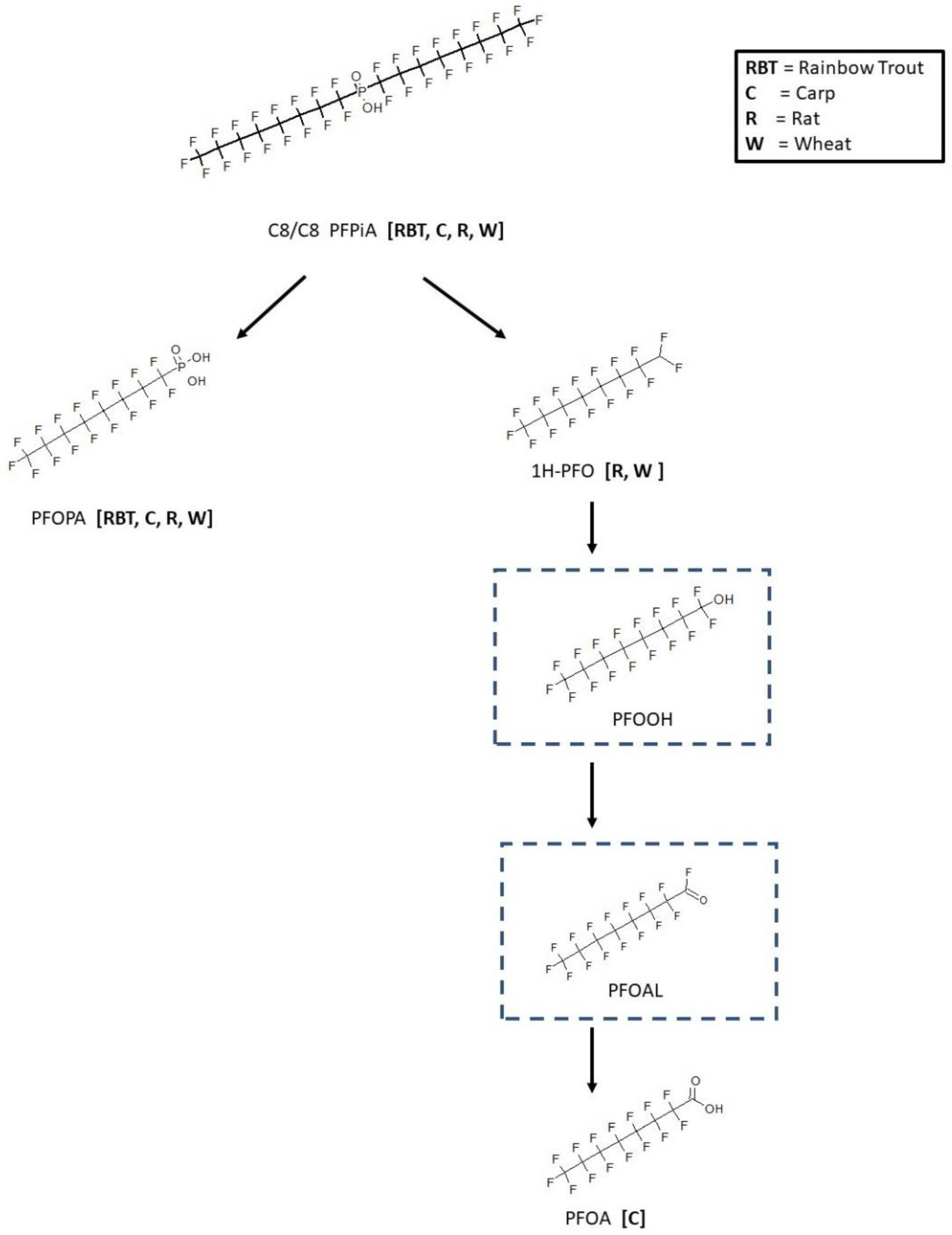
3.4. C6/C8 PFPiA Biotransformation
3.5. C6/C6 PFPiA and C8/C8 PFPiA Biotransformation
3.6. 6:2 Cl-PFESA and 8:2 Cl-PFESA Biotransformation
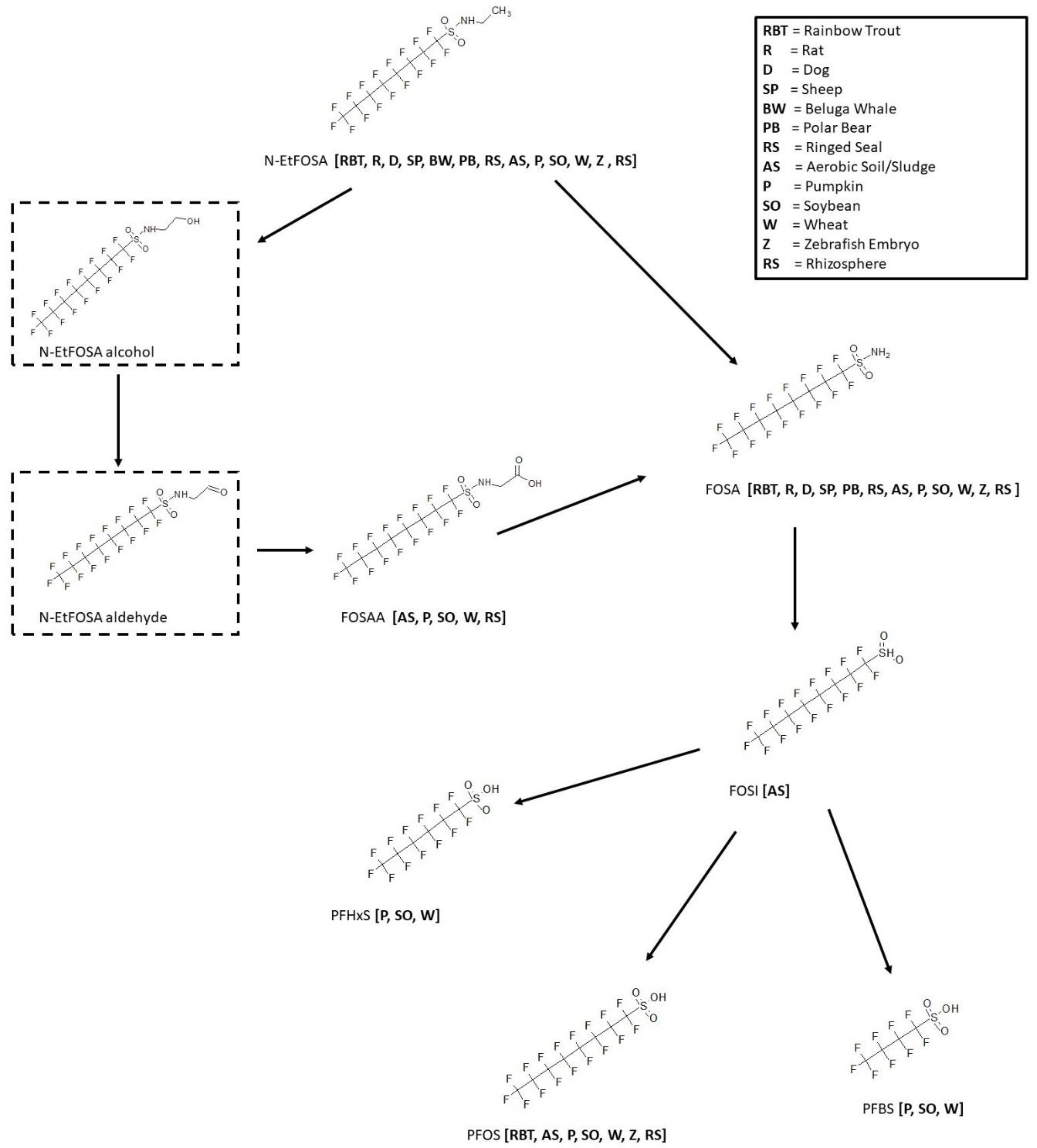
3.7. N-EtFOSA Biotransformation
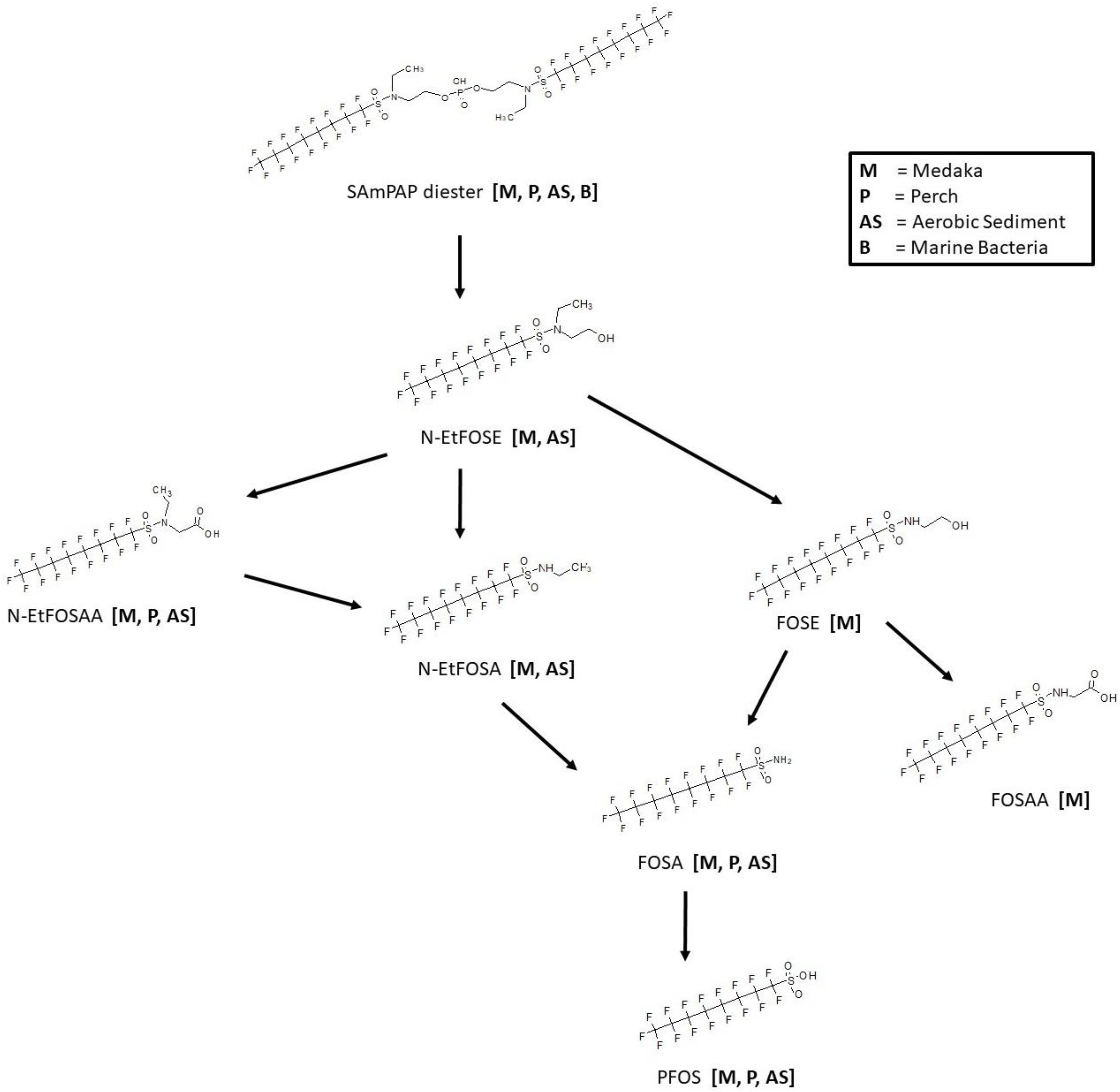
3.8. SAmPAP Diester Biotransformation
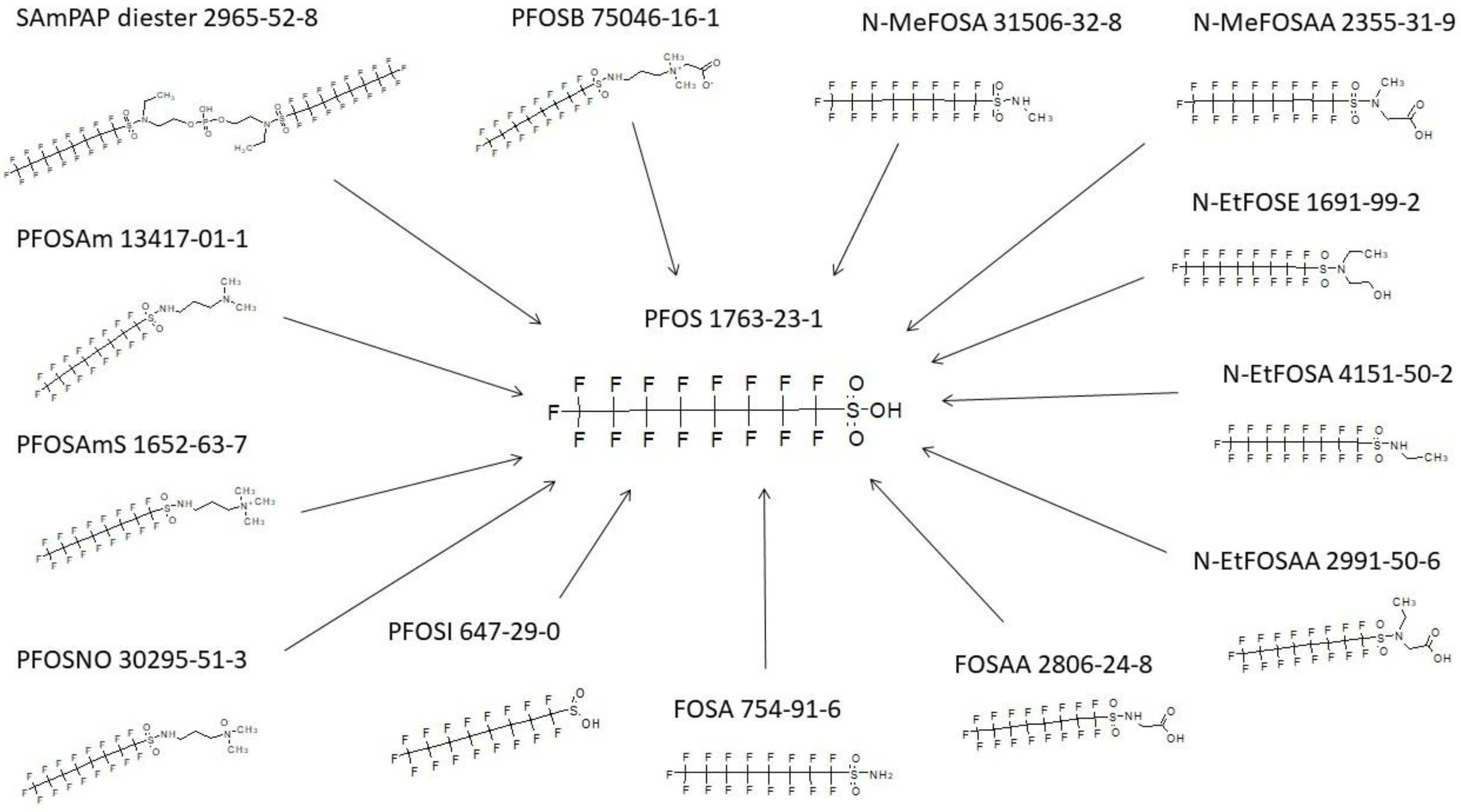
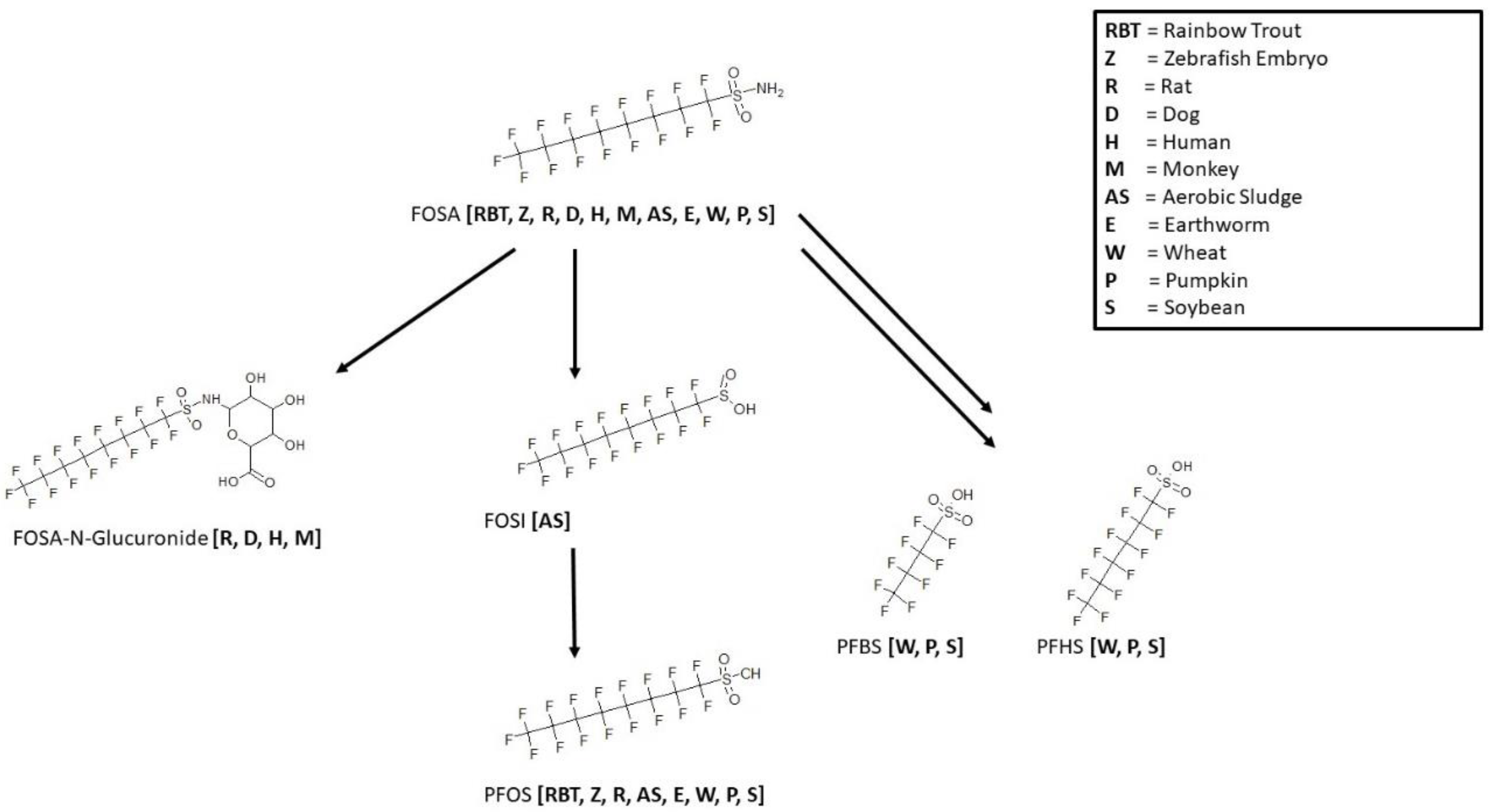
3.9. FOSA Biotransformation
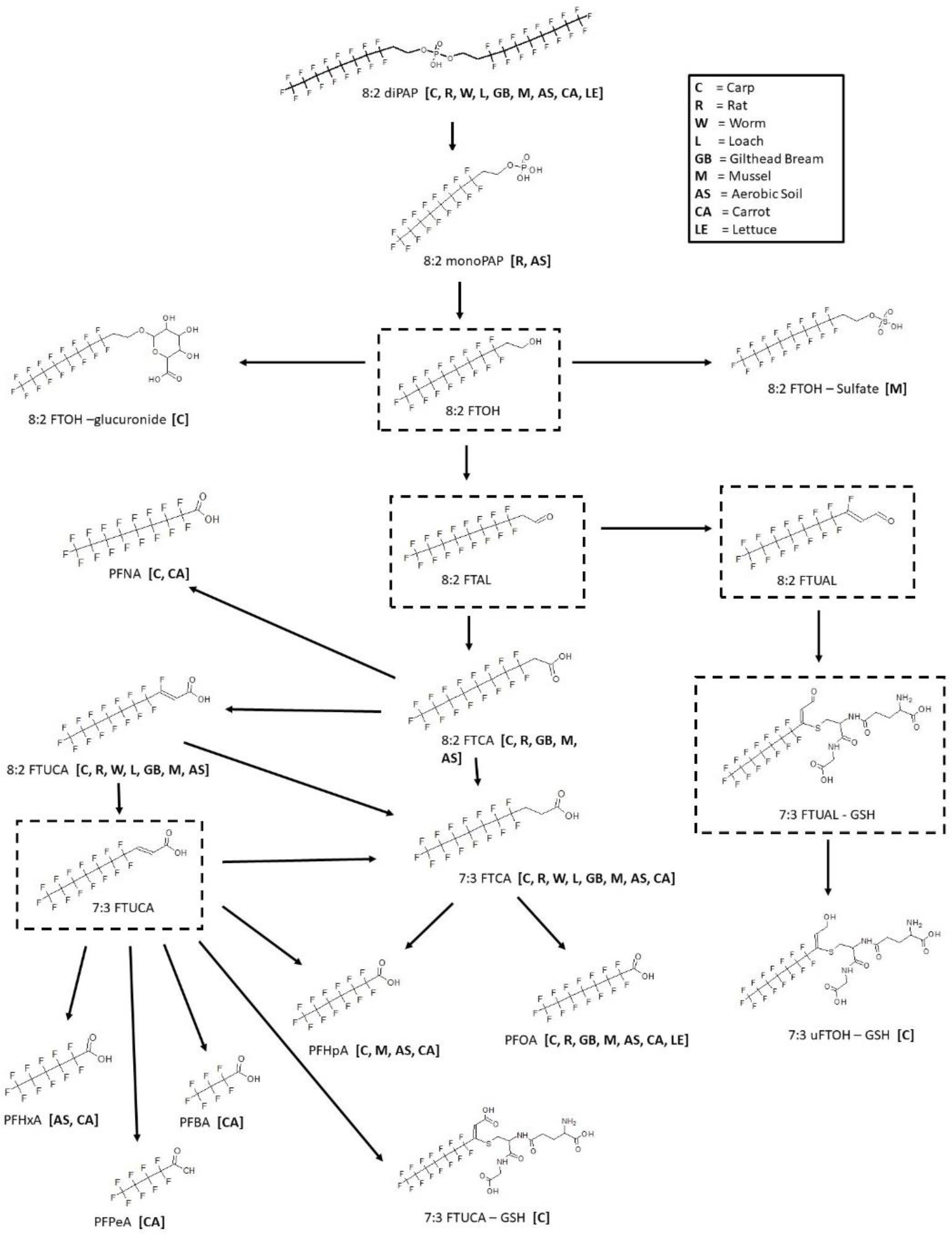
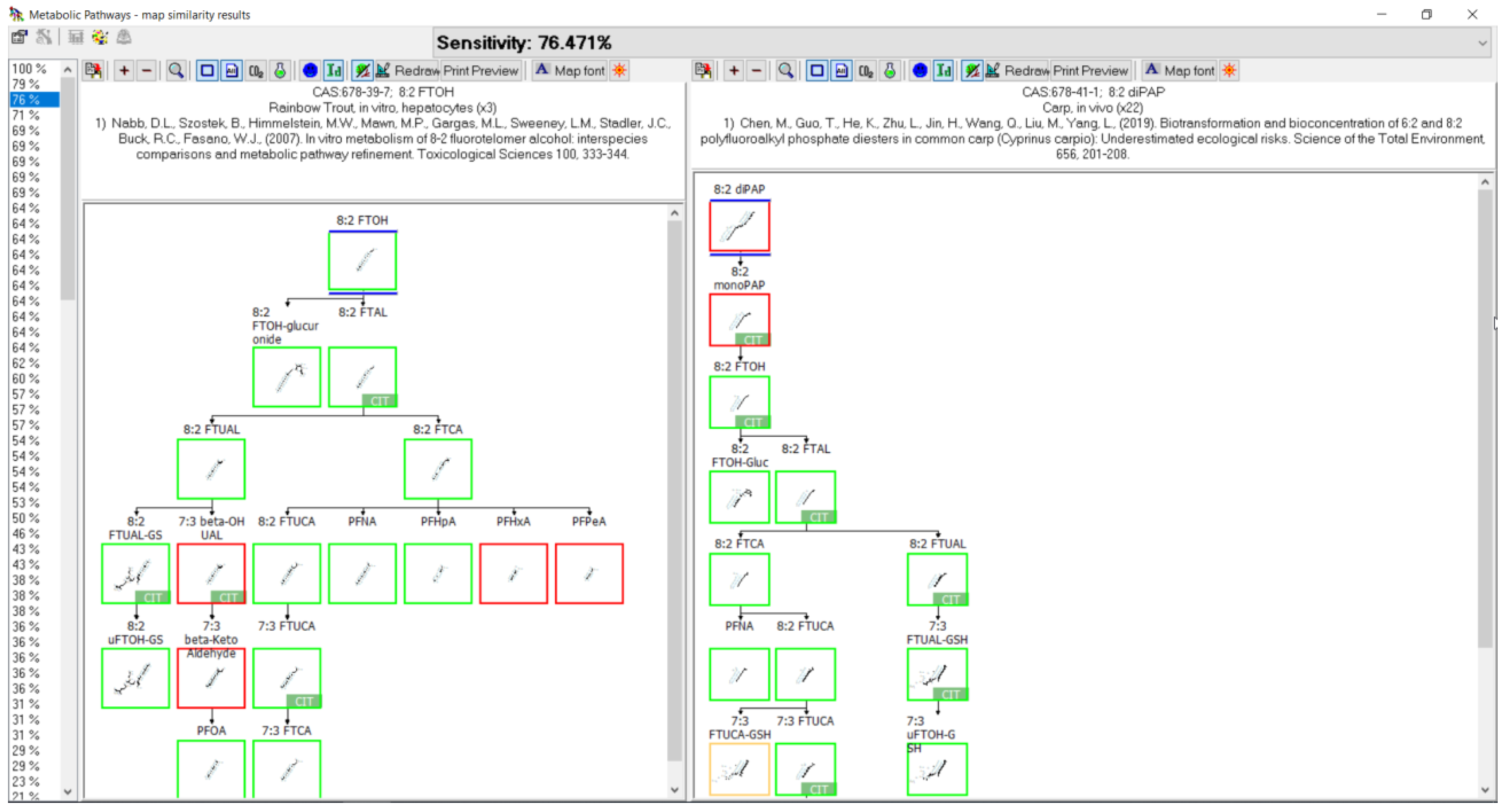
3.10. 8:2 diPAP Biotransformation
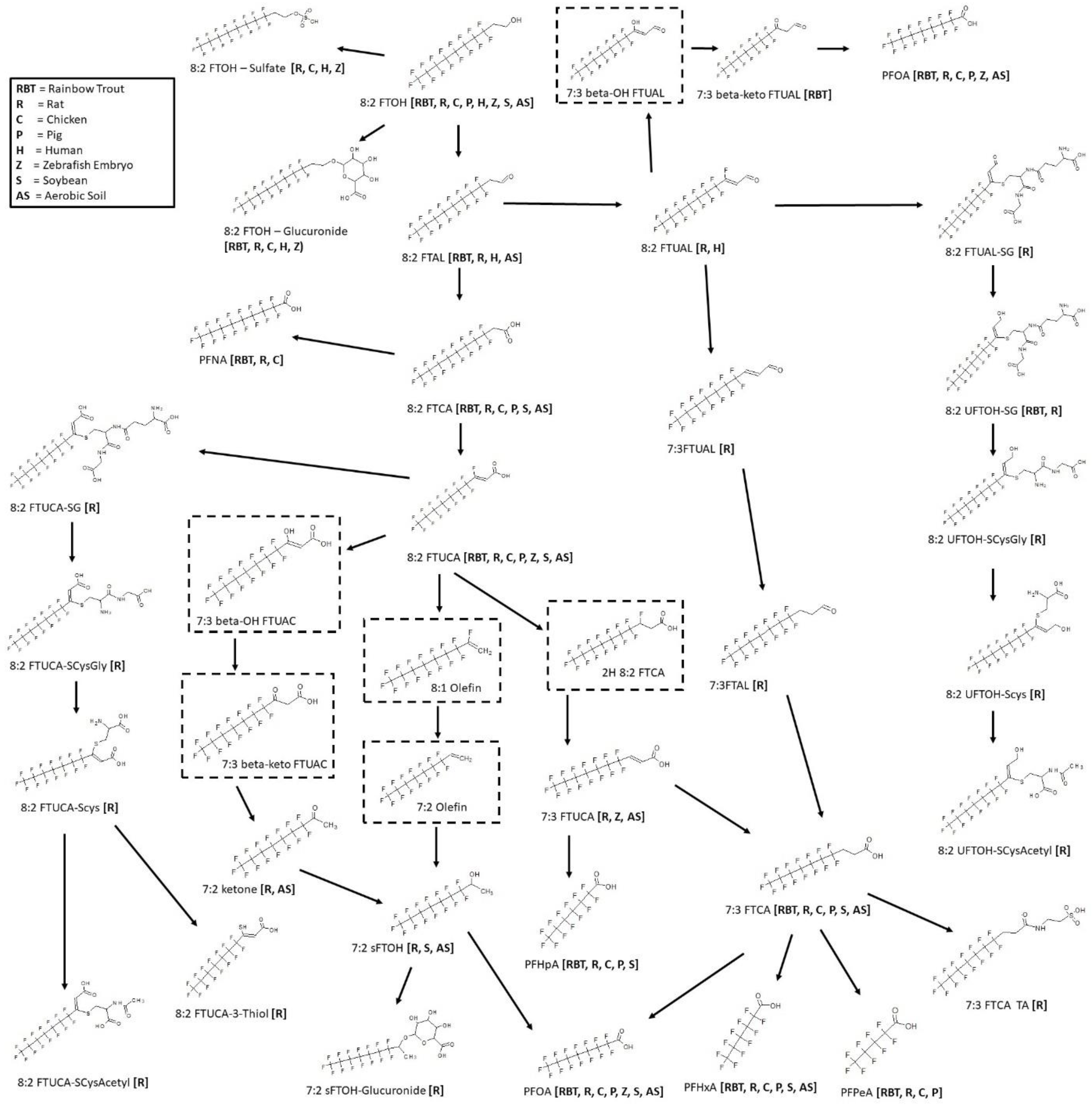
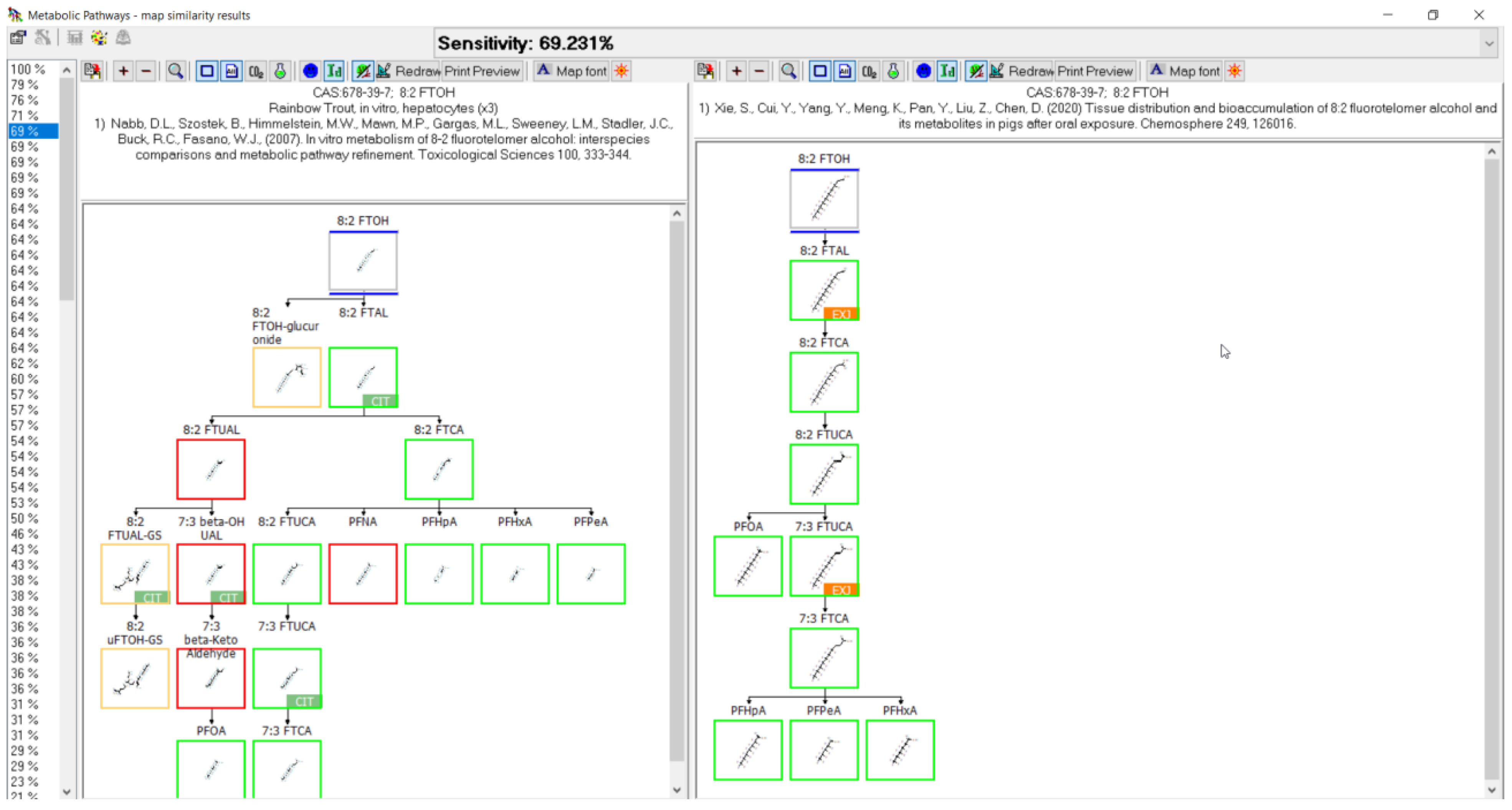
3.11. 8:2 FTOH Biotransformation
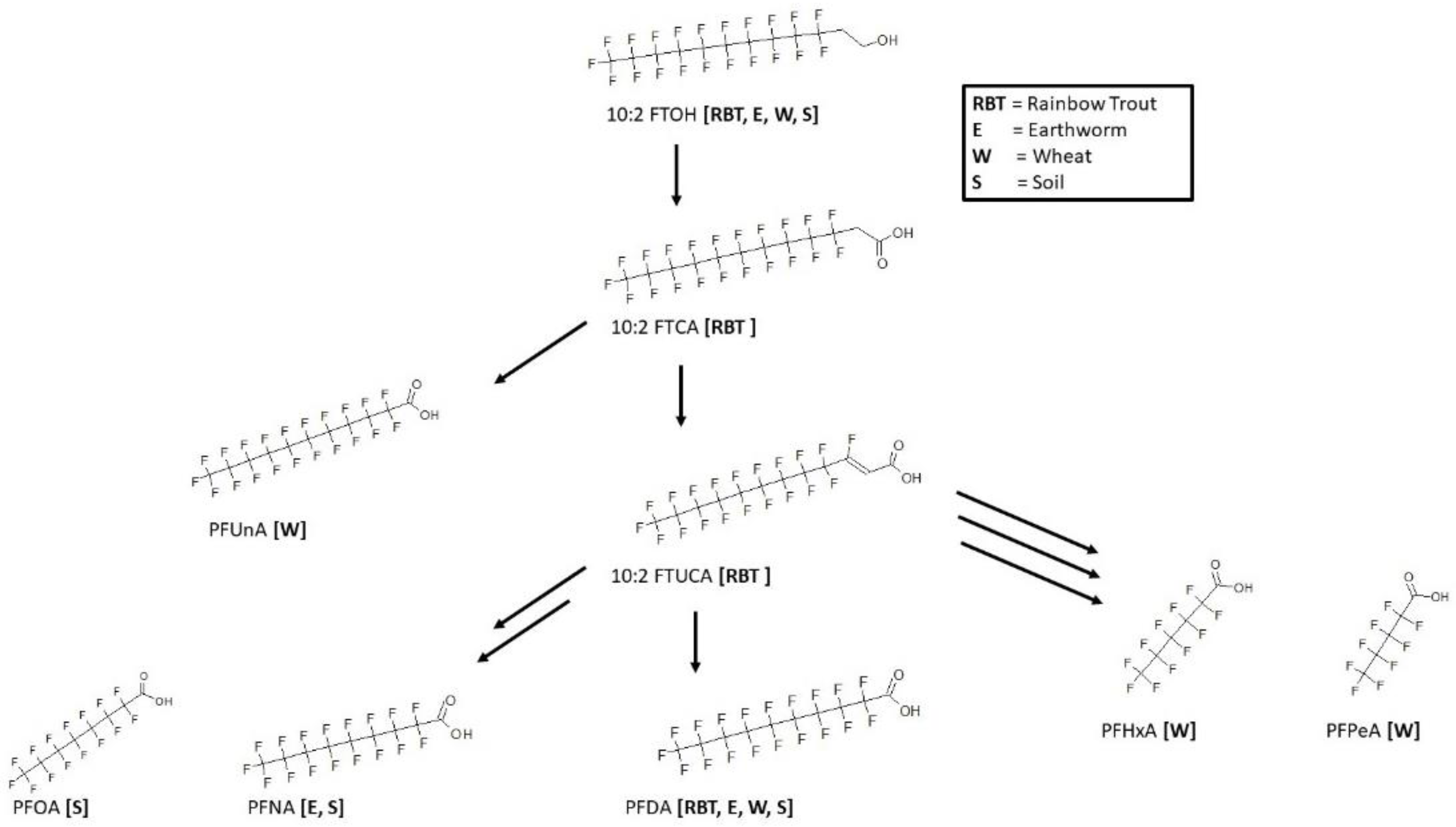
3.12. 10:2 FTOH Biotransformation
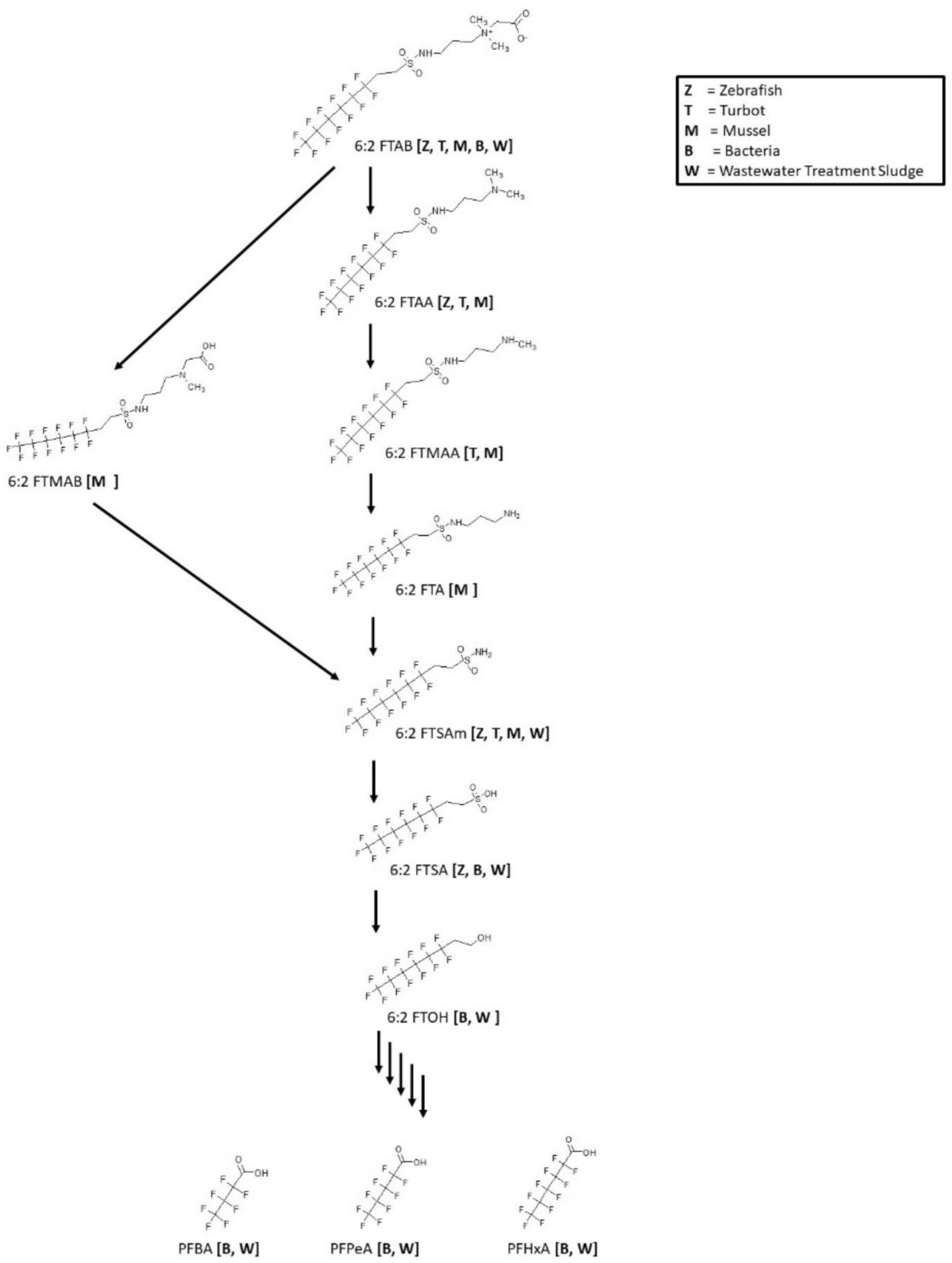
3.13. 6:2 FTAB Biotransformation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Smart, B.E. Characteristics of C-F systems. In Organofluorine Chemistry: Principles and Commercial Applications; Banks, R.E., Smart, B.E., Tatlow, J.C., Eds.; Plenum: New York, NY, USA, 1994; pp. 57–88. [Google Scholar]
- Kissa, E. Fluorinated Surfactants: Synthesis–Properties–Applications (Surfactant Science Series 50); Marcel Dekker: New York, NY, USA, 1994; 469p. [Google Scholar]
- Kissa, E. Fluorinated Surfactants and Repellents (2nd Edition Revised and Expanded) (Surfactant Science Series 97); Marcel Dekker: New York, NY, USA, 2001; 640p. [Google Scholar]
- Rao, N.S.; Baker, B.E. Textile finishes and fluorosurfactants. In Organofluorine Chemistry: Principles and Commercial Applications; Banks, R.E., Smart, B.E., Tatlow, J.C., Eds.; Plenum: New York, NY, USA, 1994; pp. 321–338. [Google Scholar]
- 3M Company. Fluorochemical Use, Distribution and Release Overview. USEPAAdministrative Record AR226-0550. 1999. Available online: https://www.regulations.gov/document/EPA-HQ-OPPT-2002-0051-0003 (accessed on 6 January 2023).
- Taylor, C.K. Fluorinated surfactants in practice. In Design and Selection of Performance Surfactants: Annual Surfactants Review; Karsa, D., Ed.; John Wiley & Sons: New York, NY, USA, 1999; pp. 271–316. [Google Scholar]
- Schaider, L.A.; Balan, S.A.; Blum, A.; Andrews, D.Q.; Strynar, M.J.; Dickinson, M.E.; Lunderberg, D.M.; Lang, J.R.; Peaslee, G.F. Fluorinated compounds in U.S. fast food packaging. Environ. Sci. Technol. Lett. 2017, 4, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Pozo, K.; Moriera, L.B.; Karaskova, P.; Pribylova, P.; Klanova, J.; de Carvalho, M.U.; Maranho, L.A.; de Souza Abessa, D.M. Using large amounts of firefighting foams releases per- and polyfluoroalkyl substances (PFAS) into estuarine environments: A baseline study in Latin America. Mar. Pollut. Bull. 2022, 182, 113938. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Diamond, M.L.; Peaslee, G.F.; Peng, H.; Blum, A.; Wang, Z.; Shalin, A.; Whitehead, H.D.; Green, M.; Schwartz-Narbonne, H.; et al. Per- and polyfluoroalkyl substances in North American school uniforms. Environ. Sci. Technol. 2022, 56, 13845–13857. [Google Scholar] [CrossRef] [PubMed]
- ATSDR. Toxicological Profile for Perfluoroalkyls. Draft for Public Comment; U.S. Dept. of Health & Human Services, Public Health Service, Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2018.
- EPA 822-R-16-002; Health Effects Support Document for Perfluorooctane Sulfonate (PFOS). U.S. Environmental Protection Agency, Office of Water (4304T) Health and Ecological Criteria Division: Washington, DC, USA, 2016.
- EPA 822-R16-003; Health Effects Support Document for Perfluorooctanoic Acid (PFOA). U.S. Environmental Protection Agency, Office of Water (4304T) Health and Ecological Criteria Division: Washington, DC, USA, 2016.
- Olker, J.H.; Elonen, C.M.; Pilli, A.; Anderson, A.; Kinziger, B.; Erickson, S.; Skopinski, M.; Pomplun, A.; LaLone, C.A.; Russom, C.L.; et al. The ECOTOXicology knowledgebase: A curated database of ecologically relevant toxicity tests to support environmental research and risk assessment. Environ. Toxicol. Chem. 2022, 41, 1520–1539. [Google Scholar] [CrossRef]
- Kolanczyk, R.C.; Schmieder, P.K.; Jones, W.J.; Mekenyan, O.G.; Chapkanov, A.; Temelkov, S.; Kotov, S.; Velikova, M.; Kamenska, V.; Vasilev, K.; et al. MetaPath: An electronic knowledge base for collating, exchanging and analyzing case studies of xenobiotic metabolism. Regul. Toxicol. Pharmacol. 2012, 63, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Buck, R.C.; Franklin, J.; Berger, U.; Conder, J.M.; Cousins, I.T.; de Voogt, P.; Jensen, A.A.; Kannan, K.; Mabury, S.A.; van Leeuwen, S.P.J. Perfluoroalkyl and polyfluoroalkyl substances in the environment: Terminology, classification, and origins. Integr. Environ. Assess. Manag. 2011, 7, 513–541. [Google Scholar] [CrossRef]
- Lee, H.; De Silva, A.O.; Mabury, S.A. Dietary bioaccumulation of perfluorophosphonates and perfluorophosphinates in juvenile rainbow trout: Evidence of metabolism of perfluorophosphinates. Environ. Sci. Technol. 2012, 46, 3489–3497. [Google Scholar] [CrossRef]
- Joudan, S.; Yeung, L.; Mabury, S.A. Biological cleavage of the C-P bond in perfluoroalkyl phophonic acids in male Sprague-Dawley rats and the formation of persistent and reactive metabolites. Environ. Health Perspect. 2017, 125, 117001. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Dong, F.; Yi, S.; Zhu, Y.; Zhou, J.; Sun, B.; Shan, G.; Feng, J.; Zhu, L. Probing mechanisms for the tissue-specific distribution and biotransformation of perfluoroalkyl phosphinic acids in common carp (Cyprinus carpio). Environ. Sci. Technol. 2020, 54, 4932–4941. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, Z.; Liu, Q.; Liu, Y.; Liu, M.; Wang, T.; Zhu, L. Insights into uptake, translocation, and transformation mechanisms of perfluorophosphinates and perfluorophosphonates in wheat (Triticum aestivum L.). Environ. Sci. Technol. 2020, 54, 276–285. [Google Scholar] [CrossRef]
- Yi, S.; Zhu, L.; Mabury, S.A. First report on in vivo pharmacokinetics and biotransformation of chlorinated polyfluoroalkyl ether sulfonates in rainbow trout. Environ. Sci. Technol. 2020, 54, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Yang, D.; Zhu, L.; Mabury, S. Significant reductive transformation of 6:2 chlorinated polyfluorooctane ether sulfonate to form hydrogen-substituted polyfluorooctane ether sulfonate and their toxicokinetics in male Sprague-Dawley rats. Environ. Sci. Technol. 2022, 56, 6123–6132. [Google Scholar] [CrossRef]
- Chen, H.; Choi, Y.J.; Lee, L.S. Sorption, aerobic biodegradation, and oxidation potential of PFOS alternatives chlorinated polyfluoroalkyl ether sulfonic acids. Environ. Sci. Technol. 2018, 52, 9827–9834. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Morson, N.; Edwards, E.A.; Yang, D.; Liu, R.; Zhu, L.; Mabury, S.A. Anaerobic microbial dechlorination of 6:2 chlorinated polyfluorooctane ether sulfonate and the underlying mechanisms. Environ. Sci. Technol. 2022, 56, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Tomy, G.T.; Tittlemier, S.A.; Palace, V.P.; Budakowski, W.R.; Braekevelt, E.; Brinkworth, L.; Friesen, K. Biotransformation of N-ethyl perfluorooctanesulfonamide by rainbow trout (Onchorhynchus mykiss) liver microsomes. Environ. Sci. Technol. 2004, 38, 758–762. [Google Scholar] [CrossRef]
- Xu, L.; Krenitsky, D.M.; Seacat, A.M.; Butenhoff, J.L.; Anders, M.W. Biotransformation of N-Ethyl-N-(2-hydroxyethyl)perfluorooctanesulfonamide by rat liver microsomes, cytosol, and slices by expressed rat and human cytochromes P450. Chem. Res. Toxicol. 2004, 17, 767–775. [Google Scholar] [CrossRef]
- Letcher, R.J.; Chu, S.; McKinney, M.A.; Tomy, G.T.; Sonne, C.; Dietz, R. Comparative hepatic in vitro depletion and metabolite formation of major perfluorooctane sulfonate precursors in arctic polar bear, beluga whale and ringed seal. Chemosphere 2014, 112, 225–231. [Google Scholar] [CrossRef]
- Arrendale, R.F.; Stewart, J.T.; Manning, R.; Vitayavirasuk, B. Determination of GX-071 and its major metabolite in rat blood by cold on-column injection capillary GC/ECD. J. Agric. Food Chem. 1989, 37, 1130–1135. [Google Scholar] [CrossRef]
- Manning, R.O.; Bruckner, J.V.; Mispagel, M.E.; Bowen, J.M. Metabolism and disposition of sulfuramid, a unique polyfluorinated insecticide, in the rat. Drug Metab. Dispos. 1991, 19, 205–211. [Google Scholar]
- Vitayavitrasuk, B.; Bowen, J.M. Pharmacokinetics of sulfuramid and its metabolite desethylsulfuramid after intravenous and intraruminal administration of sulfuramid to sheep. Pestic. Sci. 1999, 55, 719–725. [Google Scholar]
- Han, J.; Gu, W.; Barrett, H.; Yang, D.; Tang, S.; Sun, J.; Liu, J.; Krause, H.M.; Houck, K.A.; Peng, H. A roadmap to the structure-related metabolism pathways of per- and polyfluoroalkyl substances in the early life stages of zebrafish (Danio rerio). Environ. Health Perspect. 2021, 129, 77004. [Google Scholar] [CrossRef] [PubMed]
- Avendano, S.M.; Liu, J. Production of PFOS from aerobic soil biotransformation of two perfluoroalkyl sulfonamide derivatives. Chemosphere 2015, 119, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, K.R.; Janssen, E.M.L.; Luthy, R.G.; Criddle, C.S. Aerobic biotransformation and fate of N-ethyl perfluorooctane sulfonamidoethanol (N-EtFOSE) in activated sludge. Environ. Sci. Technol. 2008, 42, 2873–2878. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Harn Te, S.; Reinhard, M.; Yang, Y.; Chen, H.; He, Y.; Yew-Hoong Gin, K. Biotransformation of Sulfluramid (N-ethyl perfluorooctane sulfonamide) and dynamics of associated rhizospheric microbial community in microcosms of wetland plants. Chemosphere 2018, 211, 379–389. [Google Scholar] [CrossRef]
- Zhao, S.; Zhou, T.; Zhu, L.; Wang, B.; Li, Z.; Yang, L.; Liu, L. Uptake, translocation, and biotransformation of N-ethyl perfluorooctanesulfonamide (N-EtFOSA) by hydroponically grown plants. Environ. Pollut. 2018, 235, 404–410. [Google Scholar] [CrossRef]
- Gaillard, J.; Veyrand, B.; Thomas, M.; Dauchy, X.; Boiteux, V.; Marchand, P.; Le Bizec, B.; Banas, D.; Feidt, C. Tissue uptake, distribution, and elimination of perfluoroalkyl substances in juvenile perch through perfluorooctane sulfonamidoethanol based phosphate diester dietary exposure. Environ. Sci. Technol. 2017, 51, 7658–7666. [Google Scholar] [CrossRef]
- Peng, H.; Zhang, S.; Sun, J.; Zhang, Z.; Giesy, J.P.; Hu, J. Isomer-specific accumulation of perfluorooctanesulfonate from (N-Ethyl perfluorooctanesulfonamido)ethanol-based phosphate diester in Japanese Medaka (Oryzias latipes). Environ. Sci. Technol. 2014, 48, 1058–1065. [Google Scholar] [CrossRef]
- Zhang, S.; Peng, H.; Mu, D.; Zhao, H.; Hu, J. simultaneous determination of (N-ethyl perfluorooctanesulfonamido ethanol)-based phosphate diester and triester and their biotransformation to perfluorooctanesulfonate in freshwater sediments. Environ. Pollut. 2018, 234, 821–829. [Google Scholar] [CrossRef]
- Benskin, J.P.; Ikonomou, M.G.; Gobas, F.A.P.C.; Begley, T.H.; Woudneh, M.B.; Cosgrove, J.R. Biodegradation of N-ethyl perfluorooctane sulfonamido ethanol (EtFOSE) and EtFOSE-based phosphate diester (SAmPAP diester) in marine sediments. Environ. Sci. Technol. 2013, 47, 1381–1389. [Google Scholar] [CrossRef]
- Brandsma, S.H.; Smithwick, M.; Solomon, K.; Small, J.; de Boer, J.; Muir, D.C.G. Dietary exposure of rainbow trout to 8:2 and 10:2 fluorotelomer alcohols and perfluorooctanesulfonamide: Uptake, transformation, and elimination. Chemosphere 2011, 82, 253–258. [Google Scholar] [CrossRef]
- Xu, L.; Krenitsky, D.M.; Seacat, A.M.; Butenhoff, J.L.; Tephly, T.R.; Anders, M.W. N-Glucuronidation of perfluorooctanesulfonamide by human, rat, dog, and monkey liver microsomes and by expressed rat and human UDP-glucuronyltransferases. Drug Metab. Dispos. 2006, 34, 1406–1410. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Zhou, T.; Wang, B.; Zhu, L.; Chen, M.; Li, D. Different biotransformation behaviors of perfluorooctane sulfonamide in wheat (Triticum aestivum L.) from earthworms (Eisenia fetida). J. Hazard. Mater. 2018, 346, 191–198. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, B.; Zhong, Z.; Liu, T.; Liang, T.; Zhan, J. Contributions of enzymes and gut microbes to biotransformation of perfluorooctane sulfonamide in earthworms (Eisneia fetida). Chemosphere 2020, 238, 124619. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Liang, T.; Zhou, T.; Li, D.; Wang, B.; Zhan, J.; Liu, L. Biotransformation and responses of antioxidant enzymes in hydroponically cultured soybean and pumpkin exposed to perfluorooctane sulfonamide (FOSA). Ecotoxicol. Environ. Saf. 2018, 161, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, Q.; Zhu, Y.; Zhu, L.; Xiao, B.; Liu, M.; Yang, L. Species dependent accumulation and transformation of 8:2 polyfluoroalkyl phosphate esters in sediment by three benthic organisms. Environ. Int. 2019, 133, 105171. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Guo, T.; He, K.; Zhu, L.; Jin, H.; Wang, Q.; Liu, M.; Yang, L. Biotransformation and bioconcentration of 6:2 and 8:2 polyfluoroalkyl phosphate diesters in common carp (Cyprinus carpio): Underestimated ecological risks. Sci. Total Environ. 2019, 656, 201–208. [Google Scholar] [CrossRef]
- D’eon, J.C.; Mabury, S.A. Production of perfluoronated carboxylic acids (PFCA’s) from the biotransformation of polyfluoroalkyl phosphate surfactants (PAPS): Exploring routes of human contamination. Environ. Sci. Technol. 2007, 41, 4799–4805. [Google Scholar] [CrossRef]
- Zabaleta, I.; Bizkarguenaga, E.; Izagirre, U.; Negreira, N.; Covaci, A.; Benskin, J.P.; Prieto, A.; Zuloaga, O. Biotransformation of 8:2 polyfluoroalkyl phosphate diester in gilthead bream (Sparus aurata). Sci. Total Environ. 2017, 609, 1085–1092. [Google Scholar] [CrossRef]
- Cui, W.; Tan, Z.; Chen, J.; Wu, H.; Geng, Q.; Guo, M.; Zhai, Y. Uptake, tissue distribution and elimination of 8:2 polyfluoroalkyl phosphate diesters in Mytilus galloprovincialis. Environ. Toxicol. 2021, 40, 1990–2002. [Google Scholar] [CrossRef]
- Liu, C.; Liu, J. Aerobic biotransformation of polyfluoroalkyl phosphate esters (PAPS) in soil. Environ. Pollut. 2016, 212, 230–237. [Google Scholar] [CrossRef]
- Bizkarguenaga, E.; Zabaleta, I.; Prieto, A.; Fernandez, L.A.; Zuloaga, O. Uptake of 8:2 perfluoroalkyl phosphate diester and its degradation products by carrot and lettuce from compost-amended soil. Chemosphere 2016, 152, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Nabb, D.L.; Szostek, B.; Himmelstein, M.W.; Mawn, M.P.; Gargas, M.L.; Sweeney, L.M.; Stadler, J.C.; Buck, R.C.; Fasano, W.J. In vitro metabolism of 8-2 fluorotelomer alcohol: Interspecies comparisons and metabolic pathway refinement. Toxicol. Sci. 2007, 100, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.W.; Mabury, S.A.; O’Brien, P.J. Metabolic products and pathways of fluorotelomer alcohols in isolated rat hepatocytes. Chem.-Biol. Interact. 2005, 155, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.W.; Chan, K.; Mabury, S.A.; O’Brien, P.J. Bioactivation of fluorotelomer alcohols in isolated rat hepatocytes. Chem.-Biol. Interact. 2009, 177, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Fasano, W.J.; Carpenter, S.C.; Gannon, S.A.; Snow, T.A.; Stadler, J.C.; Kennedy, G.L.; Buck, R.C.; Korzeniowski, S.H.; Hinderliter, P.M.; Kemper, R.A. Absorption, distribution, metabolism, and elimination of 8-2 fluorotelomer alcohol in the Rat. Toxicol. Sci. 2006, 91, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Fasano, W.J.; Sweeney, L.M.; Mawn, M.P.; Nabb, D.L.; Szostek, B.; Buck, R.C.; Gargas, M.L. Kinetics of 8-2 fluorotelomer alcohol and its metabolites, and liver glutathione status following daily oral dosing for 45 days in male and female rats. Chem. Biol. Interact. 2009, 180, 281–295. [Google Scholar] [CrossRef]
- Himmelstein, M.W.; Serex, T.L.; Buck, R.C.; Weinberg, J.T.; Mawn, M.P.; Russell, M.H. 8:2 fluorotelomer alcohol: A one-day nose-only inhalation toxicokinetic study in the Sprague-Dawley rat with application to risk assessment. Toxicology 2012, 291, 122–132. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, Y.; Xu, W.; Pan, Y.; Wei, Q.; Xie, S. Biotransformation and tissue bioaccumulation of 8:2 fluorotelomer alcohol in broiler by oral exposure. Environ. Pollut. 2020, 267, 115611. [Google Scholar] [CrossRef]
- Xie, S.; Cui, Y.; Yang, Y.; Meng, K.; Pan, Y.; Liu, Z.; Chen, D. Tissue distribution and bioaccumulation of 8:2 fluorotelomer alcohol and its metabolites in pigs after oral exposure. Chemosphere 2020, 249, 126016. [Google Scholar] [CrossRef]
- Li, Z.M.; Guo, L.H.; Ren, X.M. Biotransformation of 8:2 fluorotelomer alcohol by recombinant human cytochrome P450s, human liver microsomes and human liver cytosol. Environ. Sci. Process. Impacts 2016, 18, 538. [Google Scholar] [CrossRef]
- Zhang, H.; Wen, B.; Hu, X.; Wu, Y.; Pan, Y.; Huang, H.; Liu, L.; Zhang, S. Uptake, Translocation, and Metabolism of 8:2 Fluorotelomer Alcohol in Soybean (Glycine max L. Merrill). Environ. Sci. Technol. 2016, 50, 13309–13317. [Google Scholar] [CrossRef] [PubMed]
- Dinglasan, M.J.A.; Ye, Y.; Edwards, E.A.; Mabury, S.A. Fluorotelomer alcohol biodegradation yields poly- and perfluorinated acids. Environ. Sci. Technol. 2004, 38, 2857–2864. [Google Scholar] [CrossRef]
- Wang, N.; Szosek, B.; Buck, R.C.; Folsom, P.W.; Sulecki, L.M.; Gannon, J.T. 8-2 Fluorotelomer alcohol aerobic soil biodegradation: Pathways, metabolites, and metabolic yields. Chemosphere 2009, 75, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhu, L. Uptake and metabolism of 10:2 Fluorotelomer alcohol in soil-earthworm (Eisenia fetida) and soil-wheat (Triticum aestivum L.) systems. Environ. Pollut. 2017, 220, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Cui, Q.; Zhang, H.; Cui, R.; Guo, Y.; Dai, J. Accumulation, Biotransformation, and Endocrine Effects of Fluorotelomer surfactant Mixtures on Zebrafish. Chem. Res. Toxicol. 2019, 32, 1432–1440. [Google Scholar] [CrossRef]
- Moe, M.K.; Huber, S.; Svenson, J.; Hagenaars, A.; Pabon, M.; Trumper, M.; Berger, U.; Knapen, D.; Herzke, D. The structure of the fire-fighting foam surfactant Forafac 1157 and its biological and photolytic transformation products. Chemosphere 2012, 89, 646–875. [Google Scholar] [CrossRef]
- Shaw, D.M.J.; Munoz, G.; Bottos, E.M.; Vo Duy, S.; Sauve, S.; Liu, J.; Van Hamme, J.D. Degradation and defluorination of 6:2 fluorotelomer sulfonamidoalkyl betaine and 6:2 fluorotelomer sulfonate by Gordonia sp. Starin NB4-1Y under sulfur-limiting conditions. Sci. Total Environ. 2019, 647, 690–698. [Google Scholar] [CrossRef]
- D’Agostino, L.A.; Mabury, S.A. Aerobic biodegradation of 2 fluorotelomer sulfonamide-based aqueous film-forming foam components produces perfluoroalkyl carboxylates. Environ. Toxicol. Chem. 2017, 36, 2012–2021. [Google Scholar] [CrossRef] [PubMed]
- Kolanczyk, R.C.; Serrano, J.A.; Tapper, M.A.; Schmieder, P.K. A comparison of fish pesticide metabolic pathways with those of the rat and goat. Regul. Toxicol. Pharmacol. 2018, 94, 124–143. [Google Scholar] [CrossRef] [PubMed]
- Stanley, L.A. Drug metabolism. In Pharmacognosy: Fundamentals, Applications and Strategy; Delgoda, S.B.R., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 527–545. [Google Scholar]
- Sanchez, R.I.; Kauffman, F.C. Regulation of Xenobiotic Metabolism in the Liver. In Comprehensive Toxicology, 2nd ed.; McQueen, C.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 109–128. [Google Scholar]
- Testa, B.; Clement, B. Chapter 24—Biotransformation Reactions and their Enzymes; In The Practice of Medicinal Chemistry, 4th ed.; Wermuth, C., Aldous, D., Raboisson, P., Rognan, D., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 561–584. [Google Scholar]
- Almazroo, O.A.; Miah, M.K.; Venkataramanan, R. Drug metabolism in the liver. Clin. Liver Dis. 2017, 21, 1–20. [Google Scholar] [CrossRef]
- Sandermann, H., Jr. Plant metabolism of xenobiotics. Trends Biochem. Sci. 1992, 17, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Short, C.R.; Hardy, M.L.; Barker, S.A. The in vivo oxidative metabolism of 2,4- and 2,6-dimethylaniline in the dog and rat. Toxicology 1989, 57, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Galatius, A.; Bossi, R.; Sonne, C.; Rigét, F.F.; Kinze, C.C.; Lockyer, C.; Teilmann, J.; Dietz, R. PFAS profiles in three North Sea top predators: Metabolic differences among species? Environ. Sci. Pollut. Res. 2013, 20, 8013–8020. [Google Scholar] [CrossRef] [PubMed]
- Smithwick, M.; Muir, D.C.G.; Mabury, S.A.; Solomon, K.R.; Martin, J.W.; Sonne, C.; Born, E.W.; Letcher, R.J.; Dietz, R. Perfluoroalkyl contaminants in liver tissue from East Greenland polar bears (Ursus maritimus). Environ. Toxicol. Chem. 2005, 24, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Butt, C.M.; Mabury, S.A.; Kwan, M.; Wang, X.W.; Muir, D.C.G. Spatial trends of perfluoroalkyl compounds in ringed seals (Phoca hispida) from the Canadian Arctic. Environ. Toxicol. Chem. 2008, 27, 542–553. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Corsolini, S.; Falandysz, J.; Oehme, G.; Focardi, S.; Giesy, J.P. Perfluorooctanesulfonate and related fluorinated hydrocarbons in marine mammals, fishes, and birds from coasts of the Baltic and the Mediterranean Seas. Environ. Sci. Technol. 2002, 36, 3210–3216. [Google Scholar] [CrossRef]
- Reiner, J.L.; O’Connell, S.G.; Moors, A.J.; Kucklick, J.R.; Becker, P.R.; Keller, J.M. Spatial and temporal trends of perfluorinated compounds in beluga whales (Delphinapterus leucas) from Alaska. Environ. Sci. Technol. 2011, 45, 8129–8136. [Google Scholar] [CrossRef]



















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolanczyk, R.C.; Saley, M.R.; Serrano, J.A.; Daley, S.M.; Tapper, M.A. PFAS Biotransformation Pathways: A Species Comparison Study. Toxics 2023, 11, 74. https://doi.org/10.3390/toxics11010074
Kolanczyk RC, Saley MR, Serrano JA, Daley SM, Tapper MA. PFAS Biotransformation Pathways: A Species Comparison Study. Toxics. 2023; 11(1):74. https://doi.org/10.3390/toxics11010074
Chicago/Turabian StyleKolanczyk, Richard C., Megan R. Saley, Jose A. Serrano, Sara M. Daley, and Mark A. Tapper. 2023. "PFAS Biotransformation Pathways: A Species Comparison Study" Toxics 11, no. 1: 74. https://doi.org/10.3390/toxics11010074