
Cadmium Caused Different Toxicity to Photosystem I and Photosystem II of Freshwater Unicellular Algae Chlorella pyrenoidosa (Chlorophyta)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture of Chlorella pyrenoidosa
2.2. Cd Treatments
2.3. Measurement of Cell Growth
2.4. Measurement of PSI and PSII Activities
2.4.1. Application of the Dual-PAM-100 System
2.4.2. Measurement of the Rapid Light Response Curves
2.4.3. Quantum Yields of Photosystems and CEF
2.4.4. Analysis of RLCs of Electron Transport Rates in PSI and PSII
2.5. Isolation of Photosystems and the Binding with Cd Ions
2.6. Statistics
3. Results
3.1. Effects of Cd on Growth
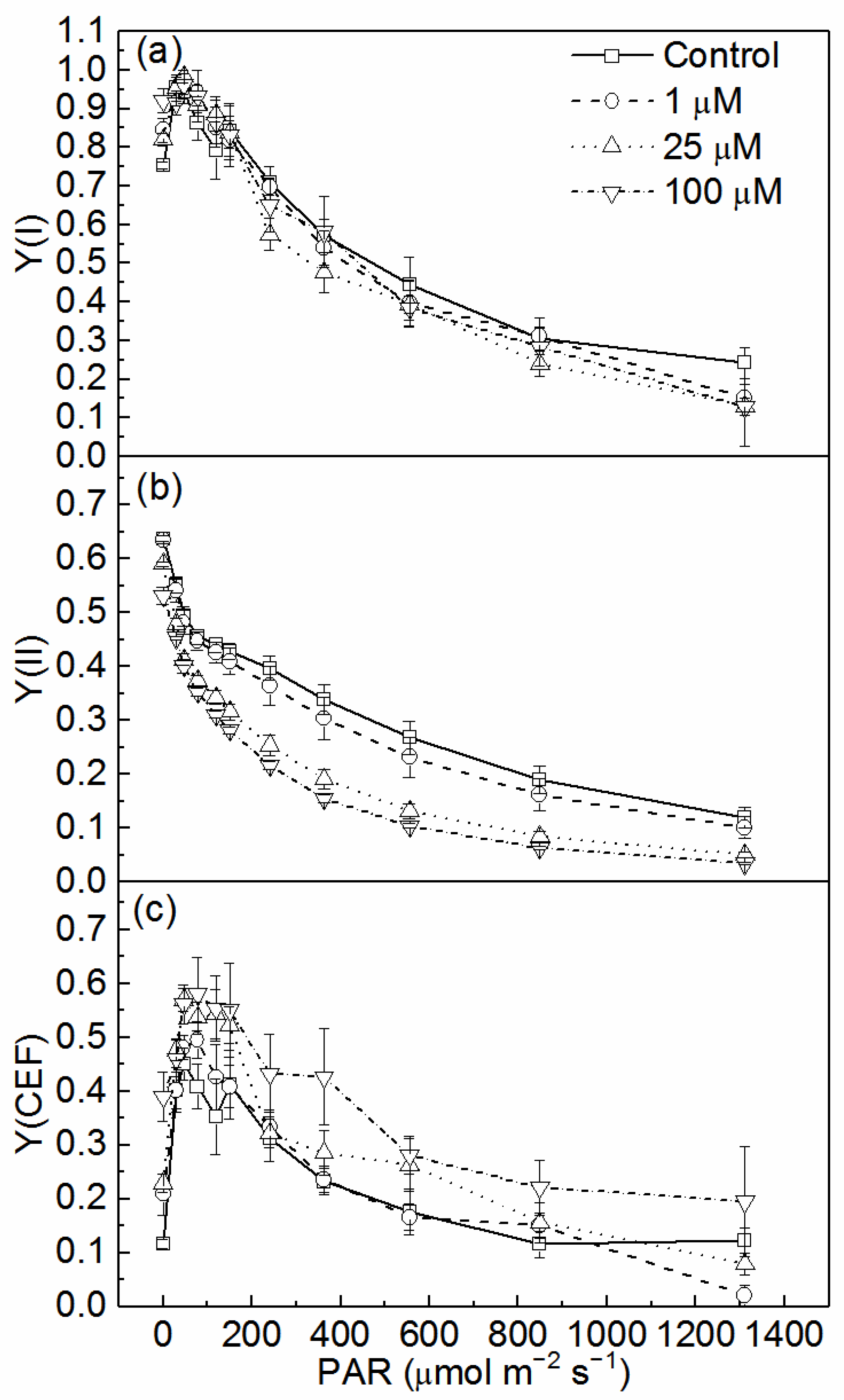
3.2. Effects of Cd on Quantum Yields of Two Photosystems and CEF
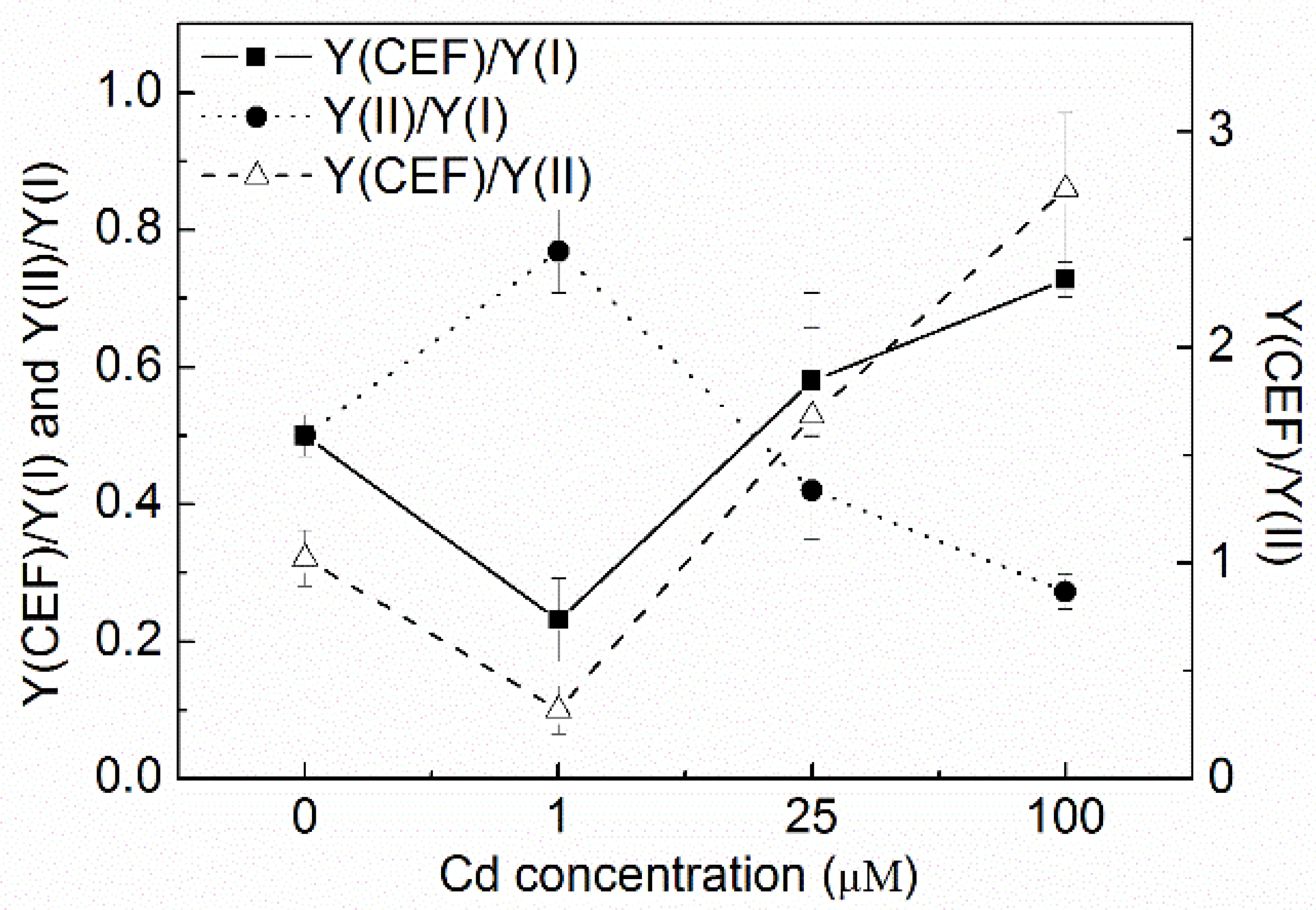
3.3. Changes of Y(CEF)/Y(I), Y(II)/Y(I), and Y(CEF)/Y(II)
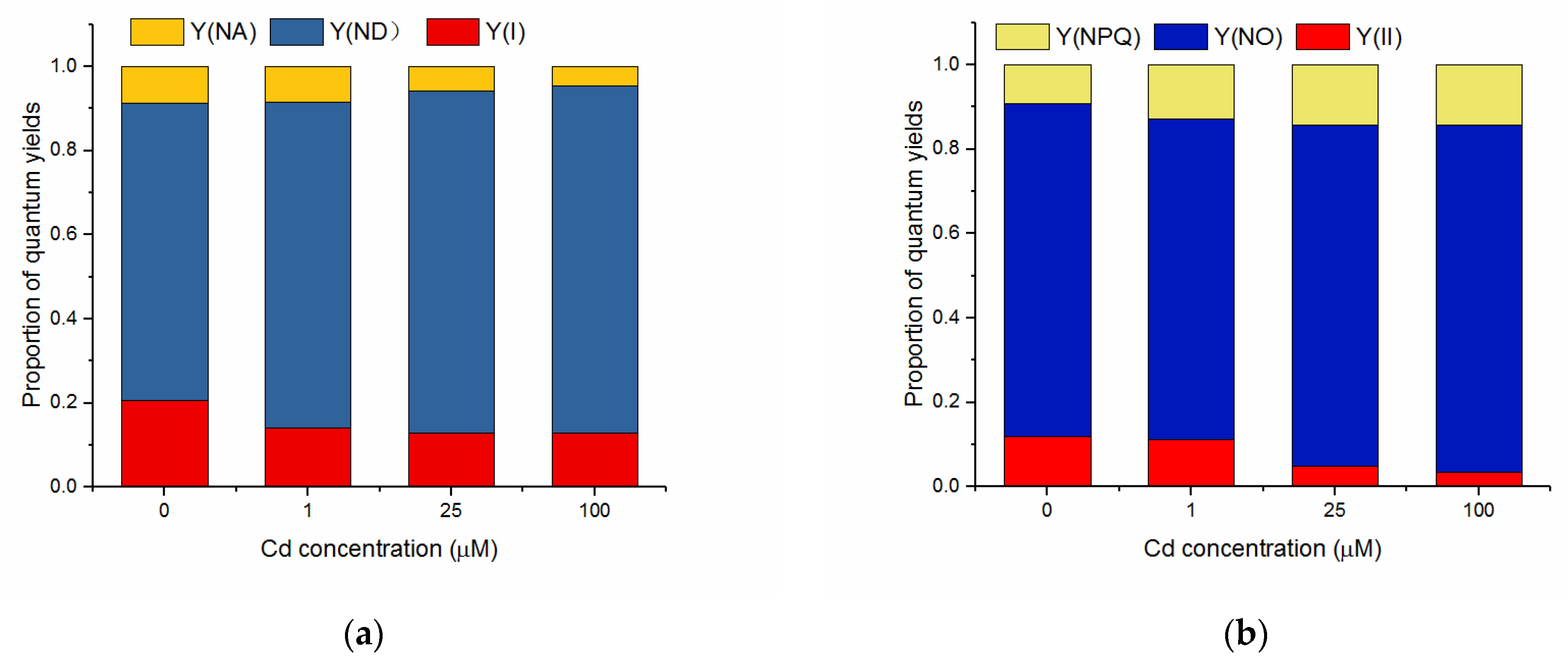
3.4. Effects of Cd on Quantum Yields of Energy Conversion in PSI and PSII
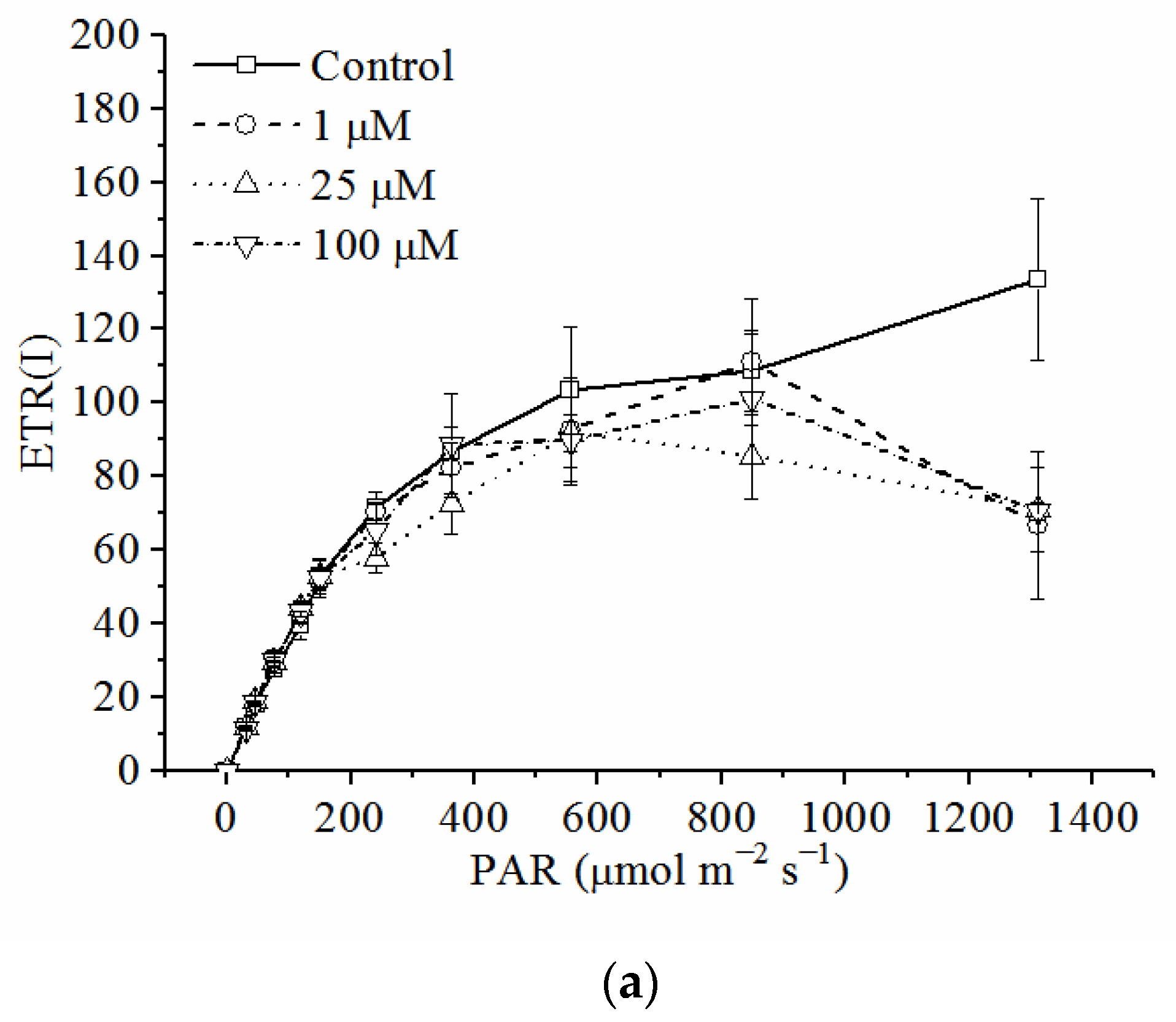
3.5. Effects of Cd on Electron Transport Rates in PSI and PSII
3.6. Binding Ability of Cd Ions with Photosystems
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, F.; Zhang, S.; Cheng, P.; Zhang, S.; Sun, Y. Effects of soil amendments on heavy metal immobilization and accumulation by maize grown in a multiple-metal-contaminated soil and their potential for safe crop production. Toxics 2020, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy metals and pesticides toxicity in agricultural soil and plants: Ecological risks and human health implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.F.; Li, J.J.; Sun, L.W.; Chen, W.; Sheng, G.D.; Liu, W.P.; Fu, Z.W. Combined effect of copper and cadmium on Chlorella vulgaris growth and photosynthesis-related gene transcription. Aquat. Toxicol. 2009, 94, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Alharby, H.F.; Al-Zahrani, H.S.; Abbas, G. Potassium and silicon synergistically increase cadmium and lead tolerance and phytostabilization by quinoa through modulation of physiological and biochemical attributes. Toxics 2022, 10, 169. [Google Scholar] [CrossRef]
- Yang, W.; Dai, H.; Skuza, L.; Wei, S. Enhanced Cd phytoextraction by Solanum nigrum L. from contaminated soils combined with the application of N fertilizers and double harvests. Toxics 2022, 10, 266. [Google Scholar] [CrossRef]
- Sperdouli, I.; Adamakis, I.-D.S.; Dobrikova, A.; Apostolova, E.; Hanć, A.; Moustakas, M. Excess zinc supply reduces cadmium uptake and mitigates cadmium toxicity effects on chloroplast structure, oxidative stress, and Photosystem II photochemical efficiency in Salvia sclarea plants. Toxics 2022, 10, 36. [Google Scholar] [CrossRef]
- Monteiro, C.M.; Fonseca, S.C.; Castro, P.M.L.; Malcata, F.X. Toxicity of cadmium and zinc on two microalgae, Scenedesmus obliquus and Desmodesmus pleiomorphus, from Northern Portugal. J. Appl. Phycol. 2011, 23, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.B.; Juneau, P.; Qiu, B.S. Growth and photosynthetic responses of the bloom-forming cyanobacterium Microcystis aeruginosa to elevated levels of cadmium. Chemosphere 2006, 65, 1738–1746. [Google Scholar] [CrossRef]
- Mohapatra, P.K.; Khillar, R.; Hansdah, B.; Mohanty, R.C. Photosynthetic and fluorescence responses of Solanum melangena L. to field application of dimethoate. Ecotoxicol. Environ. Saf. 2010, 73, 78–83. [Google Scholar] [CrossRef]
- Qian, H.F.; Chen, W.; Sheng, G.D.; Xu, X.Y.; Liu, W.P.; Fu, Z.W. Effects of glufosinate on antioxidant enzymes, subcellular structure, and gene expression in the unicellular green alga Chlorella vulgaris. Aquat. Toxicol. 2008, 88, 301–307. [Google Scholar] [CrossRef]
- Takami, R.; Almeida, J.V.; Vardaris, C.V.; Colepicolo, P.; Barros, M.P. The interplay between thiol-compounds against chromium (VI) in the freshwater green alga Monoraphidium convolutum: Toxicology, photosynthesis, and oxidative stress at a glance. Aquat. Toxicol. 2012, 118–119, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, J.; Guo, Y.N.; Wen, Y.Z.; Liu, J.; Liu, W.P. Evaluation of the role of the glutathione redox cycle in Cu(II) toxicity to green algae by a chiral perturbation approach. Aquat. Toxicol. 2012, 120–121, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zeng, Y.; Qu, J.Y.; Wang, W.-X. Mercury effects on Thalassiosira weissflogii: Applications of two-photon excitation chlorophyll fluorescence lifetime imaging and flow cytometry. Aquat. Toxicol. 2012, 110–111, 133–140. [Google Scholar] [CrossRef]
- Dewez, D.; Geoffroy, L.; Vernet, G.; Popovic, R. Determination of photosynthetic and enzymatic biomarkers sensitivity used to evaluate toxic effects of copper and fludioxonil in alga Scenedesmus obliquus. Aquat. Toxicol. 2005, 74, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.L.; Chen, X.; Zhang, D.Y.; Wang, J.L.; Deng, C.N.; Mu, G.J.; Zhu, H.S. Effect of chromium(VI) on photosystem II activity and heterogeneity of Synechocystis sp. (cyanophyta): Studied with in vivo chlorophyll fluorescence tests. J. Phycol. 2009, 45, 386–394. [Google Scholar] [CrossRef]
- Wang, S.Z.; Pan, X.L. Effects of sb(V) on growth and chlorophyll fluorescence of Microcystis aeruginosa (FACHB-905). Curr. Microbiol. 2012, 65, 733–741. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Pan, X.L.; Mu, G.J.; Wang, J.L. Toxic effects of antimony on photosystem II of Synechocystis sp. as probed by in vivo chlorophyll fluorescence. J. Appl. Phycol. 2010, 22, 479–488. [Google Scholar] [CrossRef]
- Suorsa, M.; Rossi, F.; Tadini, L.; Labs, M.; Colombo, M.; Jahns, P.; Kater, M.M.; Leister, D.; Finazzi, G.; Aro, E.-M.; et al. PGR5-PGRL1-dependent cyclic electron transport modulates linear electron transport rate in Arabidopsis thaliana. Mol. Plant 2016, 9, 271–288. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Niu, J.F.; Chen, W.Z.; Wang, G.C.; Xie, X.J.; Pan, G.H.; Gu, W.H.; Zhu, D.L. The physiological links of the increased photosystem II activity in moderately desiccated Porphyra haitanensis (Bangiales, Rhodophyta) to the cyclic electron flow during desiccation and re-hydration. Photosynth. Res. 2013, 116, 45–54. [Google Scholar] [CrossRef]
- Joliot, P.; Joliot, A. Cyclic electron flow in C3 plants. Biochim. Biophys. Acta 2006, 1757, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Busch, A.; Hippler, M. The structure and function of eukaryotic photosystem I. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 864–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, G.N. Physiology of PSI cyclic electron transport in higher plants. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Siedlecka, A.; Krupa, Z. Interaction between cadmium and iron. Accumulation and distribution of metals and changes in growth parameters of Phaseolus vulgaris L. seedlings. Acta Soc. Bot. Pol. 1996, 65, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Dobrikova, A.G.; Apostolova, E.L.; Hanć, A.; Yotsova, E.; Borisova, P.; Sperdouli, I.; Adamakis, I.-D.S.; Moustakas, M. Cadmium toxicity in Salvia sclarea L.: An integrative response of element uptake, oxidative stress markers, leaf structure and photosynthesis. Ecotoxicol. Environ. Saf. 2021, 209, 111851. [Google Scholar] [CrossRef] [PubMed]
- Neelam, A.; Rai, L.C. Differential responses of three cyanobacteria to UV-B and Cd. J. Microbiol. Biotechnol. 2003, 13, 544–551. [Google Scholar]
- TÓTh, T.; Zsiros, O.; Kis, M.; Garab, G.; KovÁCs, L. Cadmium exerts its toxic effects on photosynthesis via a cascade mechanism in the cyanobacterium, Synechocystis PCC 6803. Plant Cell Environ. 2012, 35, 2075–2086. [Google Scholar] [CrossRef]
- Todorenko, D.; Volgusheva, A.; Timofeev, N.; Kovalenko, I.; Matorin, D.; Antal, T. Multiple in vivo Effects of Cadmium on Photosynthetic Electron Transport in Pea Plants. Photochem. Photobiol. 2021, 97, 1516–1526. [Google Scholar] [CrossRef]
- Pál, M.; Horváth, E.; Janda, T.; Páldi, E.; Szalai, G. Physiological changes and defense mechanisms induced by cadmium stress in maize. J. Plant Nutr. Soil Sci. 2006, 169, 239–246. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. An improved method, using saturating light pulses, for the determination of photosystem I quantum yield via P700+-absorbance changes at 830 nm. Planta 1994, 192, 261–268. [Google Scholar] [CrossRef]
- Coopman, R.E.; Fuentes-Neira, F.P.; Briceño, V.F.; Cabrera, H.M.; Corcuera, L.J.; Bravo, L.A. Light energy partitioning in photosystems I and II during development of Nothofagus nitida growing under different light environments in the Chilean evergreen temperate rain forest. Trees-Struct. Funct. 2010, 24, 247–259. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, S.-B.; Cao, K.-F. Stimulation of cyclic electron flow during recovery after chilling-induced photoinhibition of PSII. Plant Cell Physiol. 2010, 51, 1922–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Ohmori, Y.; Ratel, E. High root temperature blocks both linear and cyclic electron transport in the dark during chilling of the leaves of rice seedlings. Plant Cell Physiol. 2011, 52, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Munekage, Y.; Hashimoto, M.; Miyake, C.; Tomizawa, K.-I.; Endo, T.; Tasaka, M.; Shikanai, T. Cyclic electron flow around photosystem I is essential for photosynthesis. Nature 2004, 429, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yang, S.J.; Zhang, S.B.; Zhang, J.L.; Cao, K.F. Cyclic electron flow plays an important role in photoprotection for the resurrection plant Paraboea rufescens under drought stress. Planta 2012, 235, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Z.; Zhang, D.Y.; Pan, X.L. Effects of cadmium on the activities of photosystems of Chlorella pyrenoidosa and the protective role of cyclic electron flow. Chemosphere 2013, 93, 230–237. [Google Scholar] [CrossRef]
- Wang, S.Z.; Pan, X.L.; Zhang, D.Y. PSI showed higher tolerance to Sb(V) than PSII due to stimulation of cyclic electron flow around PSI. Curr. Microbiol. 2015, 70, 27–34. [Google Scholar] [CrossRef]
- León-Vaz, A.; León, R.; Giráldez, I.; Vega, J.M.; Vigara, J. Impact of heavy metals in the microalga Chlorella sorokiniana and assessment of its potential use in cadmium bioremediation. Aquat. Toxicol. 2021, 239, 105941. [Google Scholar] [CrossRef]
- Yan, C.; Qu, Z.; Wang, J.; Cao, L.; Han, Q. Microalgal bioremediation of heavy metal pollution in water: Recent advances, challenges, and prospects. Chemosphere 2022, 286, 131870. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Pfündel, E.; Klughammer, C.; Schreiber, U. Monitoring the effects of reduced PS II antenna size on quantum yields of photosystems I and II using the Dual-PAM-100 measuring system. PAM Appl. Notes 2008, 1, 21–24. [Google Scholar]
- Platt, T.; Gallegos, C.L.; Harrison, W.G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. J. Mar. Res. 1980, 38, 687–701. [Google Scholar]
- Saroussi, S.; Beer, S. Alpha and quantum yield of aquatic plants derived from PAM fluorometry: Uses and misuses. Aquat. Bot. 2007, 86, 89–92. [Google Scholar] [CrossRef]
- Kühl, M.; Glud, R.N.; Borum, J.; Roberts, R.; Rysgaard, S. Photosynthetic performance of surface-associated algae below sea ice as measured with a pulse-amplitude-modulated (PAM) fluorometer and O2 microsensors. Mar. Ecol. Prog. Ser. 2001, 223, 1–14. [Google Scholar] [CrossRef]
- Fischer, N.; Sétif, P.; Rochaix, J.-D. Targeted mutations in the psaC gene of Chlamydomonas reinhardtii: Preferential reduction of FB at low temperature is not accompanied by altered electron flow from Photosystem I to ferredoxin. Biochemistry 1997, 36, 93–102. [Google Scholar] [CrossRef]
- Cullen, M.; Ray, N.; Husain, S.; Nugent, J.; Nield, J.; Purton, S. A highly active histidine-tagged Chlamydomonas reinhardtii Photosystem II preparation for structural and biophysical analysis. Photochem. Photobiol. Sci. 2007, 6, 1177–1183. [Google Scholar] [CrossRef]
- Marco, P.; Elman, T.; Yacoby, I. Binding of ferredoxin NADP+ oxidoreductase (FNR) to plant photosystem I. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 689–698. [Google Scholar] [CrossRef]
- Bals, T.; Schünemann, D. Isolation of Arabidopsis thylakoid membranes and their use for in vitro protein insertion or transport assays. In Chloroplast Research in Arabidopsis: Methods and Protocols; Jarvis, R.P., Walker, J.M., Eds.; Humana Press: New York, NY, USA, 2011; Volume I, pp. 321–338. [Google Scholar]
- Wu, X.; Zhao, X.; Deng, Z.; Liang, X.; Fang, S. Investigation of interactions between zein and natamycin by fluorescence spectroscopy and molecular dynamics simulation. J. Mol. Liq. 2021, 327, 114873. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Samson, G.; Carpentier, R. Nonphotosynthetic reduction of the intersystem electron transport chain of chloroplasts following heat stress. The Pool Size of Stromal Reductants. Photochem. Photobiol. 2001, 74, 438–443. [Google Scholar] [CrossRef]
- Gao, S.; Shen, S.D.; Wang, G.C.; Niu, J.F.; Lin, A.P.; Pan, G.H. PSI-driven cyclic electron flow allows intertidal macro-algae Ulva sp. (Chlorophyta) to survive in desiccated conditions. Plant Cell Physiol. 2011, 52, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Baścik-Remisiewicz, A.; Tomaszewska, E.; Labuda, K.; Tukaj, Z. The effect of Zn and Mn on the toxicity of Cd to the green microalga Desmodesmus armatus cultured at ambient and elevated (2%) CO2 concentrations. Pol. J. Environ. Stud. 2009, 18, 775–780. [Google Scholar]
- Tukaj, Z.; Baścik-Remisiewicz, A.; Skowroński, T.; Tukaj, C. Cadmium effect on the growth, photosynthesis, ultrastructure and phytochelatin content of green microalga Scenedesmus armatus: A study at low and elevated CO2 concentration. Environmen. Experimen. Bot. 2007, 60, 291–299. [Google Scholar] [CrossRef]
- Mateo, P.; Fernandez-Piñas, F.; Bonilla, I. O2-induced inactivation of nitrogenase as a mechanism for the toxic action of Cd2+ on Nostoc UAM 208. New Phytol. 1994, 126, 267–272. [Google Scholar] [CrossRef]
- Aguilera, A.; Amils, R. Tolerance to cadmium in Chlamydomonas sp. (Chlorophyta) strains isolated from an extreme acidic environment, the Tinto River (SW, Spain). Aquat. Toxicol. 2005, 75, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Sebastian, A.; Prasad, M.N.V. Operative photo assimilation associated proteome modulations are critical for iron-dependent cadmium tolerance in Oryza sativa L. Protoplasma 2015, 252, 1375–1386. [Google Scholar] [CrossRef]
- Monteiro, C.M.; Castro, P.M.L.; Malcata, F.X. Microalga-mediated bioremediation of heavy metal-contaminated surface waters. In Biomanagement of Metal-Contaminated Soils; Khan, M.S., Zaidi, A., Goel, R., Musarrat, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 20, pp. 365–385. [Google Scholar]
- Zhang, H.; Xu, Z.; Guo, K.; Huo, Y.; He, G.; Sun, H.; Guan, Y.; Xu, N.; Yang, W.; Sun, G. Toxic effects of heavy metal Cd and Zn on chlorophyll, carotenoid metabolism and photosynthetic function in tobacco leaves revealed by physiological and proteomics analysis. Ecotoxicol. Environ. Saf. 2020, 202, 110856. [Google Scholar] [CrossRef]
- Lysenko, E.A.; Klaus, A.A.; Kartashov, A.V.; Kusnetsov, V.V. Specificity of Cd, Cu, and Fe effects on barley growth, metal contents in leaves and chloroplasts, and activities of photosystem I and photosystem II. Plant Physiol. Biochem. 2020, 147, 191–204. [Google Scholar] [CrossRef]
- Li, L.; Chen, X.; Zhang, D.; Pan, X. Effects of insecticide acetamiprid on photosystem II (PSII) activity of Synechocystis sp. (FACHB-898). Pestic. Biochem. Physiol. 2010, 98, 300–304. [Google Scholar] [CrossRef]
- Pagter, M.; Liu, F.L.; Jensen, C.R.; Petersen, K.K. Effects of chilling temperatures and short photoperiod on PSII function, sugar concentrations and xylem sap ABA concentrations in two Hydrangea species. Plant Sci. 2008, 175, 547–555. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Xu, Z.; Wang, Y.; Teng, Z.; An, M.; Zhang, Y.; Zhu, W.; Xu, N.; Sun, G. Toxic effects of heavy metals Pb and Cd on mulberry (Morus alba L.) seedling leaves: Photosynthetic function and reactive oxygen species (ROS) metabolism responses. Ecotoxicol. Environ. Saf. 2020, 195, 110469. [Google Scholar] [CrossRef]
- Rocha, G.S.; Lombardi, A.T.; Espíndola, E.L.G. Combination of P-limitation and cadmium in photosynthetic responses of the freshwater microalga Ankistrodesmus densus (Chlorophyceae). Environ. Pollut. 2021, 275, 116673. [Google Scholar] [CrossRef]
- Huang, M.; Zhu, H.; Zhang, J.; Tang, D.; Han, X.; Chen, L.; Du, D.; Yao, J.; Chen, K.; Sun, J. Toxic effects of cadmium on tall fescue and different responses of the photosynthetic activities in the photosystem electron donor and acceptor sides. Sci. Rep. 2017, 7, 14387. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; Krause, G.H.; Winter, K. Responses of photosystem I compared with photosystem II to high-light stress in tropical shade and sun leaves. Plant Cell Environ. 2001, 24, 163–176. [Google Scholar] [CrossRef]
- Perales-Vela, H.V.; Peña-Castro, J.M.; Cañizares-Villanueva, R.O. Heavy metal detoxification in eukaryotic microalgae. Chemosphere 2006, 64, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, B.N.; Mehta, S.K.; Amar, A.; Gaur, J.P. Oxidative stress in Scenedesmus sp. during short- and long-term exposure to Cu2+ and Zn2+. Chemosphere 2006, 62, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Grajek, H.; Rydzyński, D.; Piotrowicz-Cieślak, A.; Herman, A.; Maciejczyk, M.; Wieczorek, Z. Cadmium ion-chlorophyll interaction—Examination of spectral properties and structure of the cadmium-chlorophyll complex and their relevance to photosynthesis inhibition. Chemosphere 2020, 261, 127434. [Google Scholar] [CrossRef]
- Liu, M.; Han, X.; Liu, C.-Q.; Guo, L.; Ding, H.; Lang, Y. Differences in the spectroscopic characteristics of wetland dissolved organic matter binding with Fe3+, Cu2+, Cd2+, Cr3+ and Zn2+. Sci. Total Environ. 2021, 800, 149476. [Google Scholar] [CrossRef]
- Zhang, D.; Pan, X.; Mostofa, K.M.G.; Chen, X.; Mu, G.; Wu, F.; Liu, J.; Song, W.; Yang, J.; Liu, Y.; et al. Complexation between Hg(II) and biofilm extracellular polymeric substances: An application of fluorescence spectroscopy. J. Hazard. Mater. 2010, 175, 359–365. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of CdCl2 (μM) | Parameters of the Light Curve of ETR(I) | Parameters of the Light Curve of ETR(II) | ||||
|---|---|---|---|---|---|---|
| Ik (μmol photon m−2 s−1) | α (e− photon−1) | ETRmax (μmol e− m−2 s−1) | Ik (μmol photon m−2 s−1) | α (e− photon−1) | ETRmax (μmol e− m−2 s−1) | |
| 0 | 257.3 ± 38.1 a | 0.423 ± 0.050 a | 133.0 ± 21.5 a | 304.8 ± 40.3 a | 0.226 ± 0.005 a | 69.3 ± 9.9 a |
| 1 | 240.0 ± 35.9 a | 0.434 ± 0.015 a | 102.9 ± 13.1 a | 256.6 ± 43.9 a | 0.226 ± 0.005 a | 58.4 ± 11.0 a |
| 25 | 209.4 ± 23.2 a | 0.439 ± 0.035 a | 89.8 ± 6.1 a | 150.2 ± 16.0 b | 0.205 ± 0.002 b | 30.9 ± 3.4 b |
| 100 | 202.4 ± 8.8 a | 0.394 ± 0.054 a | 89.1 ± 7.4 a | 123.9 ± 4.0 b | 0.196 ± 0.003 b | 24.3 ± 1.0 b |
| Heavy Metal | PSI Particles | PSII Particles | ||
|---|---|---|---|---|
| Ka (×104 M−1) | n | Ka (×104 M−1) | n | |
| Cd | 2.01 ± 0.18 | 0.52 ± 0.09 | 1.95 ± 0.06 | 0.77 ± 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wufuer, R.; Duo, J.; Li, W.; Pan, X. Cadmium Caused Different Toxicity to Photosystem I and Photosystem II of Freshwater Unicellular Algae Chlorella pyrenoidosa (Chlorophyta). Toxics 2022, 10, 352. https://doi.org/10.3390/toxics10070352
Wang S, Wufuer R, Duo J, Li W, Pan X. Cadmium Caused Different Toxicity to Photosystem I and Photosystem II of Freshwater Unicellular Algae Chlorella pyrenoidosa (Chlorophyta). Toxics. 2022; 10(7):352. https://doi.org/10.3390/toxics10070352
Chicago/Turabian StyleWang, Shuzhi, Rehemanjiang Wufuer, Jia Duo, Wenfeng Li, and Xiangliang Pan. 2022. "Cadmium Caused Different Toxicity to Photosystem I and Photosystem II of Freshwater Unicellular Algae Chlorella pyrenoidosa (Chlorophyta)" Toxics 10, no. 7: 352. https://doi.org/10.3390/toxics10070352