
Ecotoxicological Studies on the Action of Actara 25 WG Insecticide on Prussian Carp (Carassius gibelio) and Marsh Frog (Pelophylax ridibundus)


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The First Experiment Was Carried out with Prussian carp Individuals Separated into Four Lots, Each Lot Being Subdivided in Two Sublots (Ten Fish)
2.2. The Second Experiment Was Carried out with Prussian carp Separated into Four Lots, Each Lot Being Subdivided in Two Sublots (Ten Fish)
2.3. The Third Experiment Was Carried out with Adults of Pelophylax ridibundus, of Both Sexes, Harvested from the Lakes Bordering the City of Piteşti
3. Results and Discussion
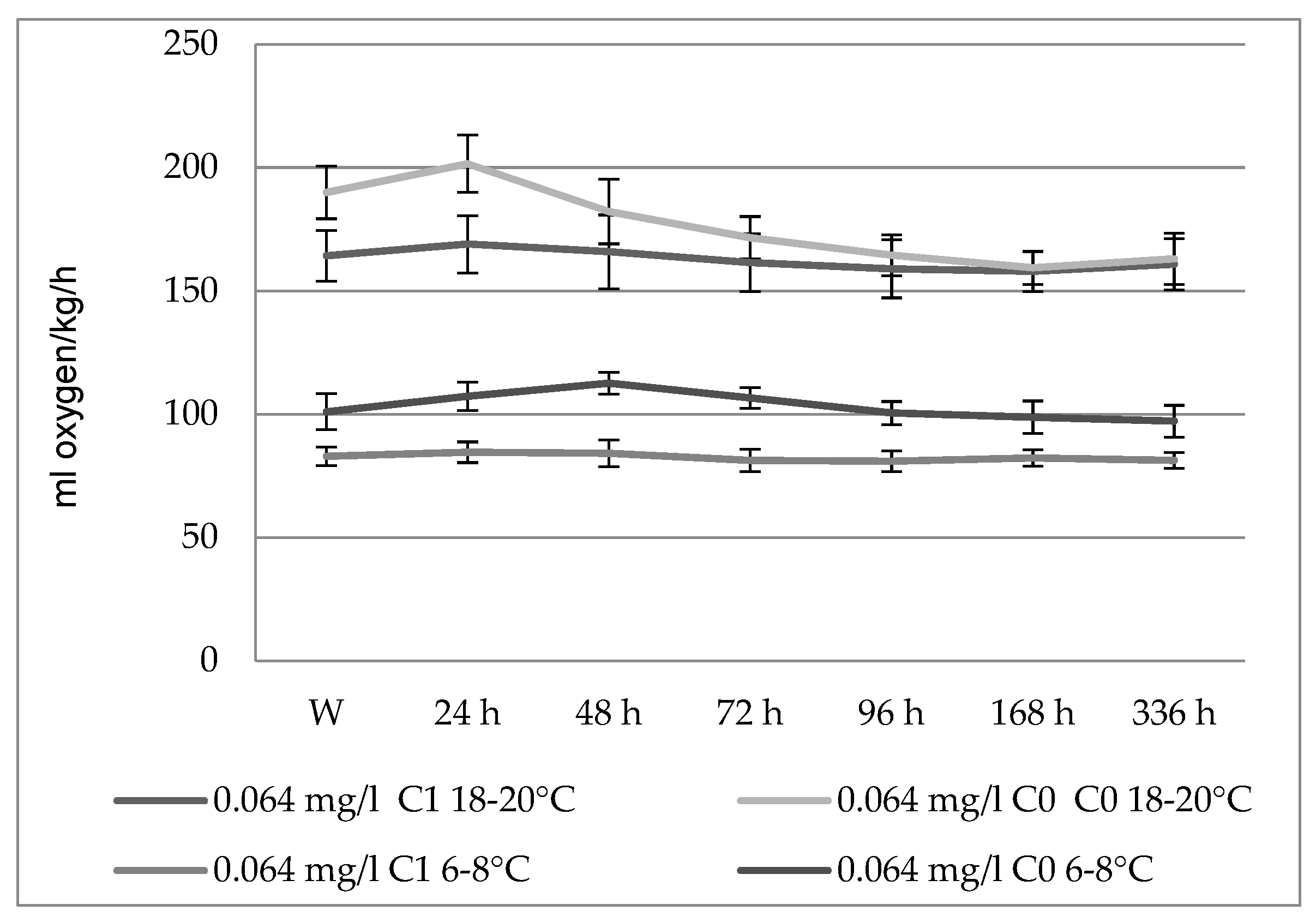
3.1. The Action of Actara 25 WG Insecticide on Oxygen Consumption in Carassius gibelio
3.2. The Action of Actara 25 WG Insecticide on the Respiratory Rate in Carassius gibelio
3.3. Influence of Fish Size on the Toxicity of Actara 25 WG Insecticide in Prussian Carp
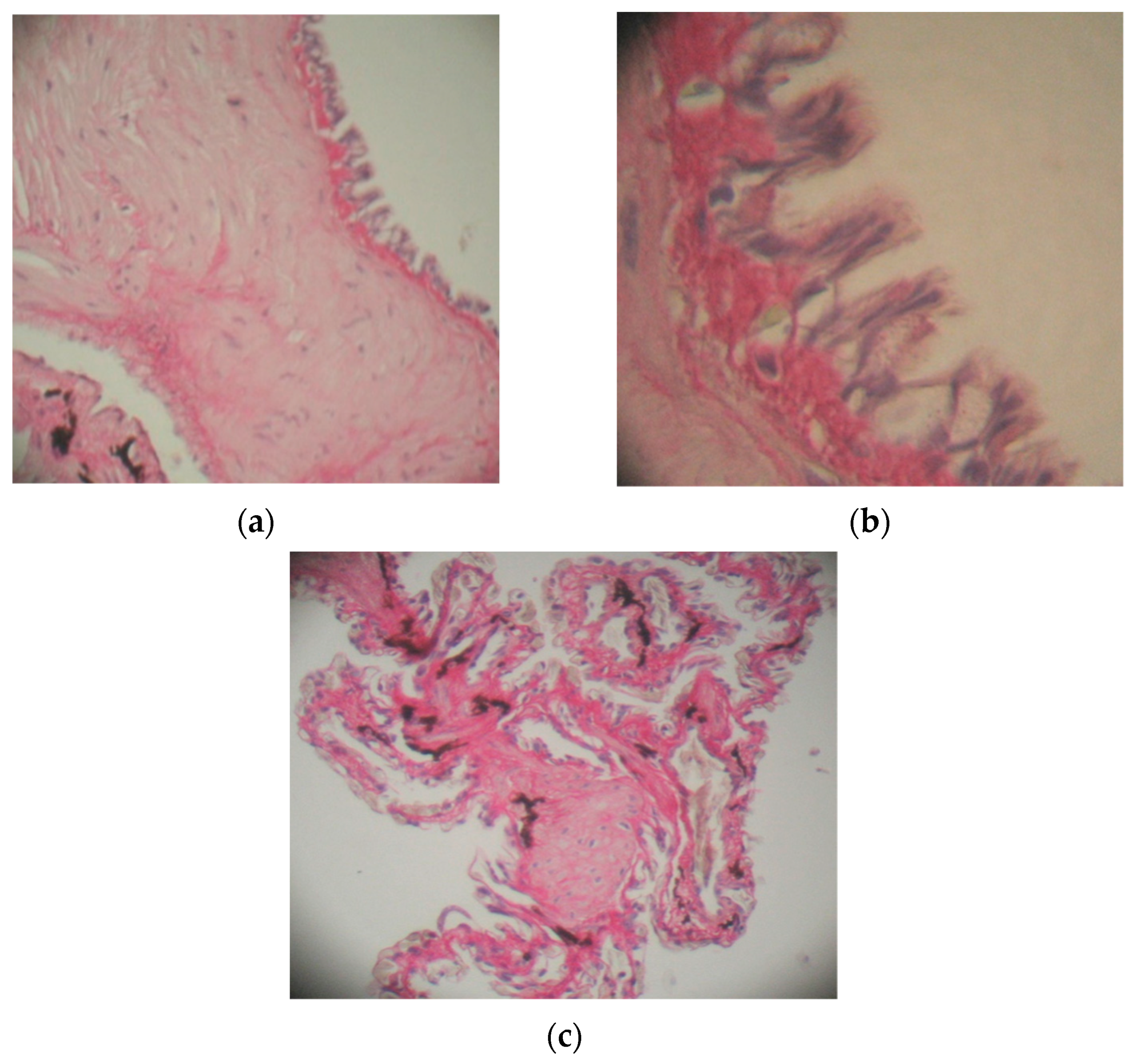
3.4. The Action of Actara 25 WG Insecticide on the Lung in Pelophylax ridibundus
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garczyńska, M.; Pączka, G.; Mazur-Pączka, A.; Kostecka, J. Earthworms in Short-Term Contact with a Low Dose of Neonicotinoid Actara 25WG. Ecol. Eng. 2018, 19, 93–101. [Google Scholar] [CrossRef]
- Wood, T.J.; Goulson, D. The environmental risks of neonicotinoid pesticides: A review of the evidence post 2013. Environ. Sci. Pollut. Res. 2017, 24, 17285–17325. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Conclusions on the peer review of the pesticide risk assessment for bees for active substance thiamethoxam considering the uses as seed treatments and granules. Eur. Fd. Saf. Author J. 2018, 16, 5179. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Sun, Y. Partition distribution of insecticides as a critical factor affecting their rates of absorption from water and relative toxicities to fish. Arch. Environ. Contam. Toxicol. 1997, 6, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Basley, K.; Goulson, D. Neonicotinoids thiamethoxam and clothianidin adversely affect the colonisation of invertebrate populations in aquatic microcosms. Environ. Sci. Pollut. Res. 2018, 25, 9593–9599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huseth, A.S.; Groves, R.L. Environmental fate of soil applied neonicotinoid insecticides in an irrigated potato agroecosystem. PLoS ONE 2014, 9, e97081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.; et al. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. Int. 2015, 22, 35–67. [Google Scholar] [CrossRef]
- Seifert, J. Neonicotinoids. In Encyclopedia of Toxicology, 2nd ed.; Wexler, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 196–200. [Google Scholar] [CrossRef]
- El Hassani, A.K.; Dacher, M.; Gary, V.; Lambin, M.; Gauthier, M.; Armengaud, C. Effects of Sublethal Doses of Acetamiprid and Thiamethoxam on the Behavior of the Honeybee (Apis mellifera). Arch. Environ. Contam. Toxicol. 2008, 54, 653–661. [Google Scholar] [CrossRef]
- Feltham, H.; Park, K.; Goulson, D. Field realistic doses of pesticide imidacloprid reduce bumblebee pollen foraging efficiency. Ecotoxicology 2014, 23, 317–323. [Google Scholar] [CrossRef]
- Gupta, R.C.; Miller Mukherjee, I.R.; Malik, J.K.; Doss, R.B.; Dettbarn, W.D.; Milatovic, D. Insecticides in Biomarkers in Toxicology, 2nd ed.; Academic Press: Oxford, UK, 2019; pp. 455–475. [Google Scholar] [CrossRef]
- Swenson, T.L.; Casida, J.E. Neonicotinoid formaldehyde generators: Possible mechanism of mouse-specific hepatotoxicity/hepatocarcinogenicity of thiamethoxam. Toxicol. Lett. 2013, 216, 139–145. [Google Scholar] [CrossRef]
- Arfat, Y.; Mahmood, N.; Tahir, M.U.; Rashid, M.; Anjum, S.; Zhao, F.; Li, D.J.; Sun, Y.L.; Hu, L.; Zhihao, C. Effect of imidacloprid on hepatotoxicity and nephrotoxicity in male albino mice. Toxicol. Rep. 2014, 1, 554–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandit, A.A.; Choudhary, S.; Singh, B.; Sethi, R.S. Imidacloprid induced histomorphological changes and expression of TLR-4 and TNFa in lung. Pest. Biochem. Physiol. 2016, 131, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Khalil, S.R.; Awad, A.; Mohammed, H.H.; Nassan, M.A. Imidacloprid insecticide exposure induces stress and disrupts glucose homeostasis in male rats. Env. Toxicol. Pharmacol. 2017, 55, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Stivaktakis, P.D.; Kavvalakis, M.P.; Tzatzarakis, M.N.; Alegakis, A.K.; Panagiotakis, M.N.; Fragkiadaki, P.; Vakonaki, E.; Ozcagli, E.; Hayes, W.A.; Rakitskii, V.N. Long-term exposure of rabbits to imidacloprid as quantified in blood induces geneotoxic effect. Chemosphere 2016, 149, 108–113. [Google Scholar] [CrossRef]
- Pandey, S.P.; Mohanty, B. The neonicotinoid pesticide imidacloprid and the dithiocarbamate fungicide mancozeb disrupt the pituitary-thyroid axis of a wildlife bird. Chemosphere 2015, 122, 227–234. [Google Scholar] [CrossRef]
- Stara, A.; Pagano, M.; Capillo, G.; Fabrello, J.; Sandova, M.; Vazzana, I.; Faggio, C. Assessing the effects of neonicotinoid insecticide on the bivalve mollusc Mytilus galloprovincialis. Sci. Total Environ. 2020, 700, 134914. [Google Scholar] [CrossRef]
- Stara, A.; Pagano, M.; Albano, M.; Savoca, S.; Di Bella, G.; Albergamo, A.; Koutkova, Z.; Sandova, M.; Velisek, J.; Fabrello, J.; et al. Effects of long-term exposure of Mytilus galloprovincialis to thiacloprid: A multibiomarker approach. Environ. Pollut. 2021, 289, 117892. [Google Scholar] [CrossRef]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef]
- Pagano, M.; Stara, A.; Aliko, V.; Faggio, C. Impact of neonicotinoids to aquatic invertebrates—In vitro studies on Mytilus galloprovincialis: A review. J. Mar. Sci. Eng. 2020, 8, 801. [Google Scholar] [CrossRef]
- Finnegan, M.C.; Baxter, L.R.; Maul, J.D.; Hanson, M.L.; Hoekstra, P.F. Comprehensive characterization of the acute and chronic toxicity of the neonicotinoid insecticide Thiamethoxam to a suite of aquatic primary producers, invertebrates, and fish. Environ. Toxicol. Chem. 2017, 36, 2838–2848. [Google Scholar] [CrossRef] [Green Version]
- Nistor, N.; Frăsinariu, O.; Ştreangă, V. Acute Poisoning with Neonicotinoid Insecticide In book: Poisoning—From Specific Toxic Agents to Novel Rapid and Simplified Techniques for Analysis. IntechOpen 2017, 6, 107–123. [Google Scholar]
- Mason, R.; Tennekes, H.; Sánchez-Bayo, F.; Jepsen, P.U. Immune suppression by neonicotinoid insecticides at the root of global wildlife declines. J. Environ. Immunol. Toxicol. 2013, 1, 3–12. [Google Scholar] [CrossRef]
- Sinha, S.; Thaker, M.A. Study on the impact of lead acetate pollutant on immunotoxicity produced by thiamethoxam pesticide. Indian J. Pharm. 2014, 46, 596–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Dick, R.A.; Ford, A.K.; Casida, J.E. Enzymes and inhibitors in Neonicotinoid insecticide metabolism. J. Agric. Food Chem. 2009, 97, 4861–4866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maienfisch, P.; Angst, M.; Brandl, F.; Fischer, W.; Hofer, D.; Kayser, H.; Kobel, W.; Rindlisbacher, A.; Senn, R.; Steinemann, A. Chemistry and biology of thiamethoxam: A second-generation neonicotinoid. Pest. Manag. Sci. 2001, 57, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Casida, J.E. Neonicotinoid insecticide toxicology: Mechanisms of Selective Action. Pharmacol. Toxicol. 2005, 45, 247–268. [Google Scholar] [CrossRef] [Green Version]
- Fulton, M.H.; Key, P.B.; DeLorenzo, M.E. Insecticide Toxicity in Fish; Tierney, K.B., Farrell, A.P., Brauner, C.J., Eds.; Fish Physiology; Academic Press: Oxford, UK, 2013; Volume 33, pp. 309–368. ISBN 9780123982544. ISSN 1546-5098. [Google Scholar] [CrossRef]
- Picoş, C.A.; Năstăsescu, G.H. Lucrări Practice de Fiziologie Animală; Tipografia Universităţii din Bucureşti: Bucureşti, Romania, 1988; Volume 107, pp. 122–123; 192–195. [Google Scholar]
- Dumitriu, I.; Fierascu, R.C.; Bunghez, I.R.; Ion, R.M. Application of inductively coupled plasma—atomic emission spectroscopy (ICP-AES) based analysis for water quality control. Environ. Eng. Manag. J. 2009, 8, 347–351. [Google Scholar] [CrossRef]
- Juncueira, L.C.U.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue section. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Directive 2010/63/EU of the European Parliament and of the Council on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, L 276/33, 33–80.
- Lima, L.B.D.; Morais, P.B.; Andrade, R.L.T.; Mattos, L.V.; Moron, S.E. Use of biomarkers to evaluate the ecological risk of xenobiotic associated with agriculture. Environ. Pollut. 2018, 237, 611–624. [Google Scholar] [CrossRef]
- Backthavathsalam, R.; Reddy, Y.S. Changes in bimodal oxygen uptake of an obligate air breather Anabas testudineus (Bloch) exposed to Lindane. Water Res. 1983, 17, 1221–1226. [Google Scholar] [CrossRef]
- Mishra, R.; Shukla, S.P. Endosulfan effects on muscle malate dehydrogenase of the freshwater catfish Claria batrachus. Ecotoxicol. Environ. Saf. 2003, 56, 425–433. [Google Scholar] [CrossRef]
- Wenderlaar Bonga, S.E.; Lock, A.A.C. Toxicants and osmoregulation in Fish. Neth. J. Zool. 1992, 42, 478–493. [Google Scholar]
- Betoulle, C.; Duchiron, C.; De Schaux, P. Lindane increase in vitro respiratory burst activity and intracellular calcium levels in rainbow trout (Oncorhynchus mykiss) head, kidney, phagocytes. J. Aquat. Toxicol. 2000, 48, 211–221. [Google Scholar] [CrossRef]
- Scheil, V.; Köhler, H. Influence of Nickel Chloride, Chlorpyrifos, and Imidacloprid in Combination with Different Temperatures on the Embryogenesis of the Zebrafish Danio rerio. Arch. Environ. Contam. Toxicol. 2009, 56, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, E.; Stoyanova, S.; Velcheva, I.; Yancheva, V. Histopathological Alterations in Common Carp (Cyprinus carpio L.) Gills Caused by Thiamethoxam. Braz. Arch. Biol. Technol. 2014, 57, 991–996. [Google Scholar] [CrossRef] [Green Version]
- Stoyanova, S.; Nyeste, K.; Georgieva, E.; Uchikov, P.; Velcheva, I.; Yancheva, V. Toxicological impact of a neonicotinoid insecticide and an organophosphorus fungicide on bighead carp (Hypophthalmichthys nobilis Richardson, 1845) gills: A comparative study. North-West. J. Zool. 2020, 16, 64–73. [Google Scholar]
- Roy, B.; Nath, S. Some haematological investigations on Oreochromis niloticus (Trewavas) following exposure to Thiamethoxam. Acta Zool. Litu. 2011, 21, 301–305. [Google Scholar] [CrossRef]
- Gul, S.T.; Khan, A.; Ahmad, M.; Anwar, M.F.; Khatoon, A.; Saleemi, M.K.; Akram, M.N. Effect of sublethal doses of thiamethoxam (a neonicotinoid) on hemato-biochemical parameters in broiler chicks. Toxin Rev. 2018, 37, 144–148. [Google Scholar] [CrossRef]
- Kumar, V.A.; Janaiah, C.; Venkateshwarlu, P. Effect of thiamethoxam alters serum biochemical parameters in Channa punctatus (Bloch). Asian. J. Bio Sci. 2010, 5, 106–110. [Google Scholar]
- Elhamalawy, O.H.; Al-Anany, F.S.; El Makawy, A.I. Thiamethoxam-induced hematological, biochemical, and genetic alterations and the ameliorated effect of Moringa oleifera in male mice. Toxicol. Rep. 2022, 9, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Al-Sharqi, S.; Alwan, M.J.; Al-Bideri, A.W. Histological changes induced by the action of Actara 25 WG insecticides in mice. Int. J. Adv. Biol. Res. 2012, 2, 760–765. [Google Scholar]
- Green, T.; Toghill, A.; Lee, R.; Waechter, F.; Weber, E.; Noakes, J. Thiamethoxam induced mouse liver tumors and their relevance to humans. Toxicol. Sci. 2005, 86, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.H.; Wang, J.H.; Zhu, L.S.; Chen, A.M.; Wnag, J. Thiametoxam induces oxidative stress and antioxidant response in zebreafish (Dario rerio) livers. Environ. Toxicol. 2015, 31, 2006–2015. [Google Scholar] [CrossRef] [PubMed]
- Velisek, J.; Stara, A. Effect of thiacloprid on early stages of common carp (Cyprinus carpio). Chemosphere 2018, 194, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.E.D.; Perez, M.R.; Acayaba, R.D.; Raimundo, C.C.M.; Martines, C.B.R. DNA damage and oxidative stress induced by imidacloprid exposure in different tissue of the Neotropical fish Prochilodus lineatus. Chemosphere 2018, 195, 125–134. [Google Scholar] [CrossRef]
- Stara, A.; Bellinvia, R.; Velisek, J.; Strouhova, A.; Kouba, A.; Faggio, C. Acute exposure of common yabby (Cherax destructor) to the neonicotinoid pesticide. Sci. Total Environ. 2019, 665, 718–723. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T (°C) | Actara 25 WG Concentration (mg/L) | Before Exposure | 24 h | 48 h | 72 h | 96 h | 168 h | 336 h |
|---|---|---|---|---|---|---|---|---|
| 18–20 | 0.064 | 164.3 ± 10.65 | 169 ± 11.58 | 165.9 ± 13.05 | 161.5 ± 8.64 | 159 ± 8.28 | 158 ± 6.64 * | 160.9 ± 10.38 * |
| 0.128 | 150 ± 9.42 | 128.9 ± 7.76 * | 115.9 ± 4.48 * | 110.88 ± 3.14 * | 102.77 ± 5.95 * | 90.88 ± 4.8 * | 92.66 ± 7.56 * | |
| 0.256 | 160.8 ± 3.45 | 131.1 ± 5.56 * | 113.44 ± 4.95 * | 104.77 ± 4.91 * | 94.87 ± 7.39 * | 88 ± 4.16 * | 86.14 ± 9.8 * | |
| 0.512 | 157.7 ± 5.25 | 115.88 ± 3.78 | 103.42 ± 5.22 | 94.4 ± 5.5 | 90.25 ± 10.21 * | 90.5 ± 16.52 * | 89.33 ± 4.16 * | |
| 0 (control lot) | 165 ± 9.54 | 167 ± 2.42 | 166 ± 2.55 | 164 ± 3.65 | 166 ± 1.56 | 168 ± 4.82 | 166 ± 3.46 | |
| 6–8 | 0.064 | 82.9 ± 3.78 | 84.6 ± 4.19 | 84.1 ± 5.42 | 81.3 ± 4.52 | 80.9 ± 4.17 | 82.3 ± 3.33 | 81.3 ± 3.26 |
| 0.128 | 81.4 ± 4.03 | 73.6 ± 3.47 * | 70.7 ± 3.94 * | 69.9 ± 2.99 * | 68.3 ± 3.94 * | 67.3 ± 4.47 * | 66.2 ± 3.6 * | |
| 0.256 | 84.9 ± 2.92 | 72.4 ± 2.67 * | 67.4 ± 2.79 * | 64.88 ± 3.51 * | 62.33 ± 4.18 * | 60.62 ± 4.27 * | 61.25 ± 3.25 * | |
| 0.512 | 88.3 ± 3.19 | 71.3 ± 2.75 * | 64.22 ± 3.52 * | 58.37 ± 4.8 * | 52.28 ± 2.98 * | 53.28 ± 2.92 * | 52.66 ± 1.63 * | |
| 0 (control lot) | 84 ± 2.54 | 86 ± 3.25 | 84 ± 4.82 | 86 ± 3.64 | 85 ± 3.25 | 82 ± 2.45 | 83 ± 2.55 |
| T (°C) | Actara 25 WG Concentration (mg/L) | Before Exposure | 24 h | 48 h | 72 h | 96 h | 168 h | 336 h |
|---|---|---|---|---|---|---|---|---|
| 18–20 | 0.064 | 71.5 ± 4.08 | 72.2 ± 3.04 | 72.2 ± 2.04 | 70.8 ± 1.75 | 69.3 ± 1.76 | 68.2 ± 1.68 | 70.4 ± 3.73 |
| 0.128 | 67.6 ± 3.65 | 79.3 ± 5.29 | 80.6 ± 5.44 * | 72.44 ± 5.91 | 64.55 ± 3.97 * | 61.11 ± 4.22 * | 59.88 ± 4.67 * | |
| 0.256 | 71.2 ± 1.75 | 83.1 ± 4.01 * | 82.11 ± 4.13 * | 73.55 ± 6.36 | 68.25 ± 5.7 * | 52.828 ± 2.82 * | 59.57 ± 2.99 * | |
| 0.512 | 72.2 ± 4.93 | 75.66 ± 1.22 | 70.85 ± 2.73 | 65.6 ± 2.19 | 62 ± 2.82 * | 58 ± 4.24 * | 59.33 ± 5.77 * | |
| 0 (control lot) | 72 ± 2.45 | 73 ± 1.25 | 73 ± 064 | 75 ± 2.52 | 71 ± 3.55 | 73 ± 2.45 | 73 ± 3.45 | |
| 6–8 | 0.064 | 50.3 ± 2.86 | 52 ± 3.74 | 51.8 ± 3.25 | 51.6 ± 3.33 | 49.3 ± 3.26 | 49.6 ± 2.67 | 49.4 ± 2.95 |
| 0.128 | 47.9 ± 1.91 | 53.7 ± 1.82 | 52.9 ± 1.72 * | 50.3 ± 2.05 | 47.3 ± 2.49 | 44.7 ± 1.63 * | 43.11 ± 2.97 * | |
| 0.256 | 48.9 ± 2.23 | 55.1 ± 1.44 | 54.4 ± 1.5 * | 50.88 ± 2.36 | 47.66 ± 2.29 | 46 ± 2.87 | 44.12 ± 2.53 | |
| 0.512 | 47.6 ± 2.45 | 57.8 ± 4.58 * | 54.55 ± 3.24 * | 45.37 ± 2.26 | 42.28 ± 2.42 * | 39.85 ± 4.09 * | 41.66 ± 1.86 | |
| 0 (control lot) | 51 ± 2.45 | 52 ± 1.25 | 50 ± 3.25 | 51 ± 2.65 | 50 ± 4.15 | 51 ± 2.67 | 50 ± 3.25 |
| Batch | Number of Measurements | Average and Standard Deviation |
|---|---|---|
| C 6–8 °C | 200 | 0.58 ± 0.029 |
| Lot I | 200 | 0.67 ± 0.019 |
| C 18–20 °C | 200 | 0.75 ± 0.022 |
| Lot II | 200 | 0.88 ± 0.018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paunescu, A.; Soare, L.C.; Fierascu, I.; Fierascu, R.C.; Mihaescu, C.F.; Tofan, L.; Ponepal, C.M. Ecotoxicological Studies on the Action of Actara 25 WG Insecticide on Prussian Carp (Carassius gibelio) and Marsh Frog (Pelophylax ridibundus). Toxics 2022, 10, 114. https://doi.org/10.3390/toxics10030114
Paunescu A, Soare LC, Fierascu I, Fierascu RC, Mihaescu CF, Tofan L, Ponepal CM. Ecotoxicological Studies on the Action of Actara 25 WG Insecticide on Prussian Carp (Carassius gibelio) and Marsh Frog (Pelophylax ridibundus). Toxics. 2022; 10(3):114. https://doi.org/10.3390/toxics10030114
Chicago/Turabian StylePaunescu, Alina, Liliana Cristina Soare, Irina Fierascu, Radu Claudiu Fierascu, Cristina Florina Mihaescu, Lucica Tofan, and Cristina Maria Ponepal. 2022. "Ecotoxicological Studies on the Action of Actara 25 WG Insecticide on Prussian Carp (Carassius gibelio) and Marsh Frog (Pelophylax ridibundus)" Toxics 10, no. 3: 114. https://doi.org/10.3390/toxics10030114