
Deltamethrin-Induced Respiratory and Behavioral Effects and Adverse Outcome Pathways (AOP) in Short-Term Exposed Mozambique Tilapia, Oreochromis mossambicus
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Procedures
2.2. Sublethal Toxicity Endpoints
2.2.1. Behavioral Toxicity
2.2.2. Respiratory Distress
2.3. AOP Development
2.4. Statistical Analysis
3. Results and Discussion
3.1. 96-h Acute Toxicity Studies
3.2. General Unified Threshold Model of Survival (GUTS) Analysis
3.3. Toxicity Factor and Safe Level Assessment
3.4. Respiratory Distress
3.5. Behavioral Toxicity
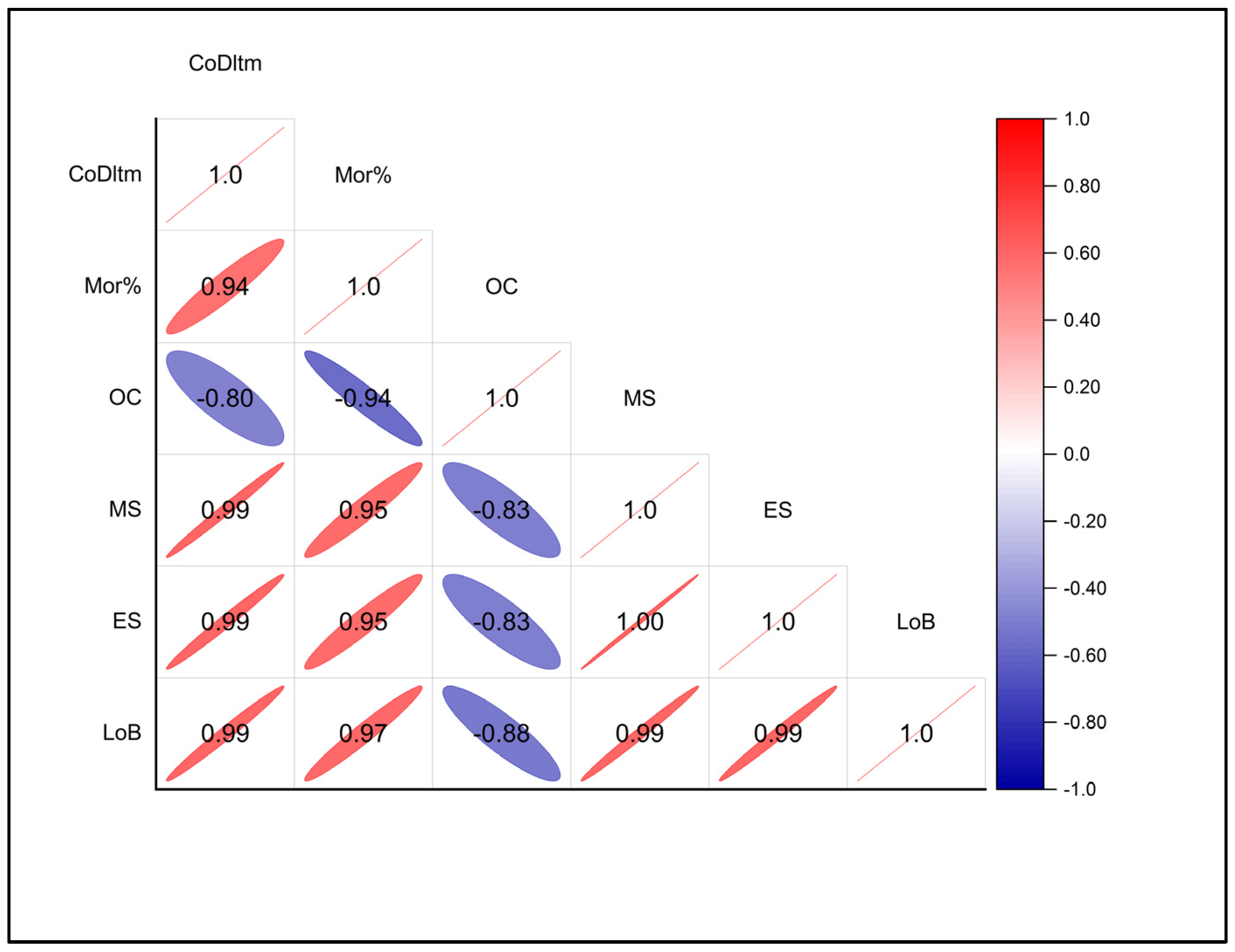
3.6. Correlation Analysis
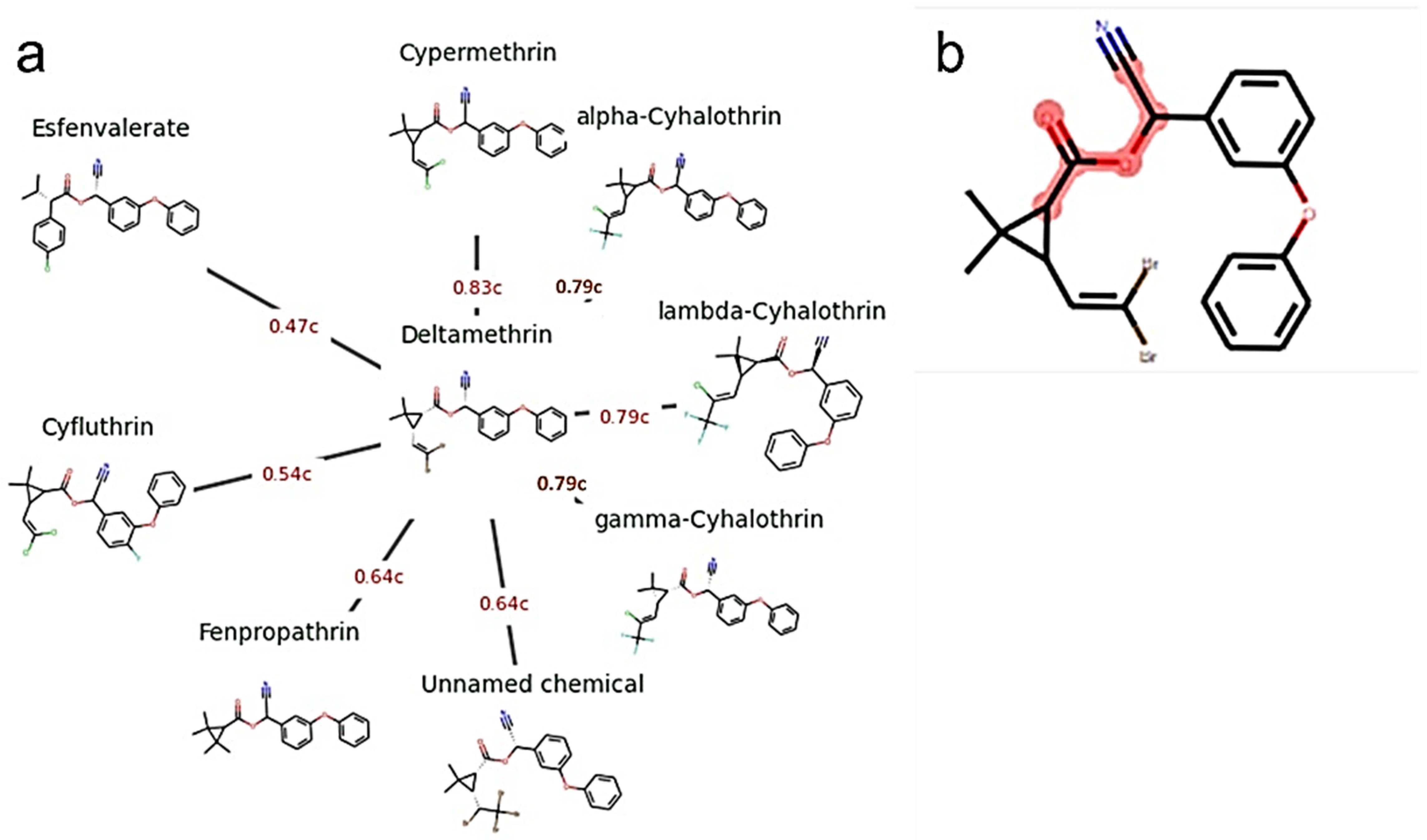
3.7. In Silico Read-Across Analysis
AOP for Behavioral and Respiratory Toxicity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Esteve-Llorens, X.; Ita-Nagy, D.; Parodi, E.; González-García, S.; Moreira, M.T.; Feijoo, G.; Vázquez-Rowe, I. Environmental footprint of critical agro-export products in the Peruvian hyper-arid coast: A case study for green asparagus and avocado. Sci. Total Environ. 2022, 818, 151686. [Google Scholar] [CrossRef]
- Fu, H.; Tan, P.; Wang, R.; Li, S.; Liu, H.; Yang, Y.; Wu, Z. Advances in organophosphorus pesticides pollution: Current status and challenges in ecotoxicological, sustainable agriculture, and degradation strategies. J. Hazard. Mater. 2022, 424, 127494. [Google Scholar] [CrossRef]
- Persello, C.; Wegner, J.D.; Hänsch, R.; Tuia, D.; Ghamisi, P.; Koeva, M.; Camps-Valls, G. Deep learning and earth observation to support the sustainable development goals: Current approaches, open challenges, and future opportunities. IEEE Geosci. Remotse Sens. Mag. 2022, 10, 172–200. [Google Scholar] [CrossRef]
- Fenibo, E.O.; Ijoma, G.N.; Matambo, T. Biopesticides in sustainable agriculture: Current status and future prospects. In New and Future Development in Biopesticide Research: Biotechnological Exploration; Springer: Berlin/Heidelberg, Germany, 2022; pp. 1–53. [Google Scholar]
- Caquet, T.; Hanson, M.L.; Roucaute, M.; Graham, D.W.; Lagadic, L. Influence of isolation on the recovery of pond mesocosms from the application of an insecticide. II. Benthic macroinvertebrate responses. Environ. Toxicol. Chem. Int. J. 2007, 26, 1280–1290. [Google Scholar] [CrossRef]
- Hanson, R.; Dodoo, D.; Essumang, D.; Blay, J.; Yankson, K. The effect of some selected pesticides on the growth and reproduction of fresh water Oreochromis niloticus, Chrysicthys nigrodigitatus and Clarias gariepinus. Bull. Environ. Contam. Toxicol. 2007, 79, 544–547. [Google Scholar] [CrossRef]
- Farag, M.R.; Alagawany, M.; Bilal, R.M.; Gewida, A.G.; Dhama, K.; Abdel-Latif, H.M.; Amer, M.S.; Rivero-Perez, N.; Zaragoza-Bastida, A.; Binnaser, Y.S. An overview on the potential hazards of pyrethroid insecticides in fish, with special emphasis on cypermethrin toxicity. Animals 2021, 11, 1880. [Google Scholar] [CrossRef]
- Datta, M.; Kaviraj, A. Acute toxicity of the synthetic pyrethroid deltamethrin to freshwater catfish Clarias gariepinus. Bull. Environ. Contam. Toxicol. 2003, 70, 296–299. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef]
- Sokolova, I.M. Energy-limited tolerance to stress as a conceptual framework to integrate the effects of multiple stressors. Integr. Comp. Biol. 2013, 53, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, S.P.; Coats, J.R. Comparative toxicology of the pyrethroid insecticides. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 1989; pp. 133–177. [Google Scholar] [CrossRef]
- Eni, G.; Ibor, O.R.; Andem, A.B.; Oku, E.E.; Chukwuka, A.V.; Adeogun, A.O.; Arukwe, A. Biochemical and endocrine-disrupting effects in Clarias gariepinus exposed to the synthetic pyrethroids, cypermethrin and deltamethrin. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 225, 108584. [Google Scholar] [CrossRef]
- Pimpao, C.T.; Zampronio, A.R.; de Assis, H.C.S. Exposure of Ancistrus multispinis (Regan, 1912, Pisces, Teleostei) to deltamethrin: Effects on cellular immunity. Fish Shellfish Immunol. 2008, 25, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Sun, Y.; Ares, I.; Anadón, A.; Martínez, M.; Martínez-Larrañaga, M.R.; Yuan, Z.; Wang, X.; Martínez, M.A. Deltamethrin toxicity: A review of oxidative stress and metabolism. Environ. Res. 2019, 170, 260–281. [Google Scholar] [CrossRef] [PubMed]
- Montanha, F.P.; Galeb, L.A.; Mikos, J.D.; Ganeco, L.N.; Pereira, T.P.; Tanaka, A.; Kirschnik, P.G.; Pimpão, C.T. Pyrethroid toxicity in silver catfish, Rhamdia quelen. Pesqui. Veterinária Bras. 2012, 32, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Chukwuka, A.V.; Mukherjee, D.; Dhara, K.; Saha, N.C.; Faggio, C. Behavioral and physiological toxicity thresholds of a freshwater vertebrate (Heteropneustes fossilis) and invertebrate (Branchiura sowerbyi), exposed to zinc oxide nanoparticles (nZnO): A General Unified Threshold model of Survival (GUTS). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 262, 109450. [Google Scholar] [CrossRef]
- Kishore, D.; Shubhajit, S.; Chukwuka, A.V.; Chandra, S.N. Behavioural toxicity and respiratory distress in early life and adult stage of walking catfish Clarias batrachus (Linnaeus) under acute fluoride exposures. Toxicol. Environ. Health Sci. 2022, 14, 33–46. [Google Scholar] [CrossRef]
- Ogbeide, O.; Uhunamure, G.; Uwagboe, L.; Osakpamwan, T.; Glory, M.; Chukwuka, A. Comparative gill and liver pathology of Tilapia zilli, Clarias gariepinus and Neochanna diversus in owan river (Nigeria): Relative ecological risks of species in a pesticide-impacted river. Chemosphere 2019, 234, 1–13. [Google Scholar] [CrossRef]
- Stine, K.E.; Brown, T.M. Principles of Toxicology; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Méjanelle, L.; Jara, B.; Dachs, J. Fate of pyrethroids in freshwater and marine environments. In Pyrethroid Insecticides; Springer: Berlin/Heidelberg, Germany, 2020; pp. 81–107. [Google Scholar]
- Pawlisz, A.; Busnarda, J.; McLauchlin, A.; Caux, P.Y.; Kent, R. Canadian water quality guidelines for deltamethrin. Environ. Toxicol. Water Qual. Int. J. 1998, 13, 175–210. [Google Scholar] [CrossRef]
- Amin, K.A.; Hashem, K.S. Deltamethrin-induced oxidative stress and biochemical changes in tissues and blood of catfish (Clarias gariepinus): Antioxidant defense and role of alpha-tocopherol. BMC Vet. Res. 2012, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Chukwuka, A.; Ogbeide, O.; Uhunamure, G. Gonad pathology and intersex severity in pelagic (Tilapia zilli) and benthic (Neochanna diversus and Clarias gariepinus) species from a pesticide-impacted agrarian catchment, south-south Nigeria. Chemosphere 2019, 225, 535–547. [Google Scholar] [CrossRef]
- Kukkonen, J.V.; Landrum, P.F. Effect of particle-xenobiotic contact time on bioavailability of sediment-associated benzo (a) pyrene to benthic amphipod, Diporeia spp. Aquat. Toxicol. 1998, 42, 229–242. [Google Scholar] [CrossRef]
- Ortego, L.S.; Benson, W.H. Effects of dissolved humic material on the toxicity of selected pyrethroid insecticides. Environ. Toxicol. Chem. Int. J. 1992, 11, 261–265. [Google Scholar] [CrossRef]
- Chukwuka, A.V.; Adebowale, A.H.; Adekolurejo, O.A. Morphological differences and trophic niche breadth of Sarotherodon galilaeus and Oreochromis niloticus from the freshwater Lake Geriyo, North-Eastern Nigeria. Croat. J. Fish. Ribar. 2019, 77, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Ibor, O.R.; Eni, G.; Andem, A.B.; Joseph, A.; Chukwuka, A.V. Sexual Dimorphism in Black Jaw Tiliapia () and Banded Jewelfish () from the Great KWA River, Calabar, Nigeria. Croat. J. Fish. 2017, 75, 122–131. [Google Scholar] [CrossRef]
- Giddings, J.M.; Wirtz, J.; Campana, D.; Dobbs, M. Derivation of combined species sensitivity distributions for acute toxicity of pyrethroids to aquatic animals. Ecotoxicology 2019, 28, 242–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawood, M.A.; AbdEl-Kader, M.F.; Moustafa, E.M.; Gewaily, M.S.; Abdo, S.E. Growth performance and hemato-immunological responses of Nile tilapia (Oreochromis niloticus) exposed to deltamethrin and fed immunobiotics. Environ. Sci. Pollut. Res. 2020, 27, 11608–11617. [Google Scholar] [CrossRef] [PubMed]
- Amrutha, M.; Nampoothiri, K.M. In silico analysis of nitrilase-3 protein from Corynebacterium glutamicum for bioremediation of nitrile herbicides. J. Genet. Eng. Biotechnol. 2022, 20, 51. [Google Scholar] [CrossRef]
- Raies, A.B.; Bajic, V.B. In silico toxicology: Computational methods for the prediction of chemical toxicity. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2016, 6, 147–172. [Google Scholar] [CrossRef] [Green Version]
- Grasso, E.J.; Sottile, A.E.; Coronel, C.E. Structural prediction and in silico physicochemical characterization for mouse caltrin I and bovine caltrin proteins. Bioinform. Biol. Insights 2016, 10, 225–236. [Google Scholar] [CrossRef] [Green Version]
- U.S. Environmental Protection Agency. Integrated Risk Information System; U.S. Environmental Protection Agency: Washington, DC, USA, 2011; p. 20460.
- Reish, D.L.; Oshida, P.S. Manual of Methods in Aquatic Environment Research; Food & Agriculture Organization: Rome, Italy, 1987; Volume 247. [Google Scholar]
- Dhara, K.; Saha, S.; Panigrahi, A.K.; Saha, N.C. Sensitivity of the freshwater tropical oligochaete, Branchiura sowerbyi (Beddard, 1892) to the grey list metal, Zinc. Int. J. Life Sci. 2020, 8, 93–101. [Google Scholar]
- Saha, S.; Saha, N.C. Study on Acute Toxicity of Bifenthrin to (Clarias batrachus Linn.). Indian J. Ecol. 2021, 48, 545–548. [Google Scholar]
- Dhara, K.; Chukwuka, A.V.; Saha, S.; Saha, N.C.; Faggio, C. Effects of short-term selenium exposure on respiratory activity and proximate body composition of early-life stages of Catla catla, Labeo rohita and Cirrhinus mrigala. Environ. Toxicol. Pharmacol. 2021, 90, 103805. [Google Scholar] [CrossRef] [PubMed]
- Pandit, D.N.; Sinha, A. Ecotoxicity of silver nanoparticles in an Indian air-breathing catfish, Clarias batrachus (Linnaeus). J. Entomol. Zool. Stud. 2019, 7, 906–911. [Google Scholar]
- Dhara, K.; Saha, S.; Chukwuka, A.V.; Pal, P.; Saha, N.C.; Faggio, C. Fluoride sensitivity in freshwater snail, Bellamya bengalensis (Lamarck, 1882): An integrative biomarker response assessment of behavioral indices, oxygen consumption, haemocyte and tissue protein levels under environmentally relevant exposure concentrations. Environ. Toxicol. Pharmacol. 2021, 89, 103789. [Google Scholar] [PubMed]
- Dhara, K.; Saha, S.; Mukherjee, D.; Saha, N.C. Comparative acute toxicity of mercury to air breathing fish, Channa gachua (Ham.) and non-air breathing fish Cyprinus carpio (Linn.): Ethological and Haematological Consideration. Indian J. Ecol. 2021, 48, 1243–1253. [Google Scholar]
- Villeneuve, D.L.; Crump, D.; Garcia-Reyero, N.; Hecker, M.; Hutchinson, T.H.; LaLone, C.A.; Landesmann, B.; Lettieri, T.; Munn, S.; Nepelska, M. Adverse outcome pathway (AOP) development I: Strategies and principles. Toxicol. Sci. 2014, 142, 312–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.J.; Grulke, C.M.; Edwards, J.; McEachran, A.D.; Mansouri, K.; Baker, N.C.; Patlewicz, G.; Shah, I.; Wambaugh, J.F.; Judson, R.S.; et al. The CompTox Chemistry Dashboard: A community data resource for environmental chemistry. J. Cheminform. 2017, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finney, D. Statistical logic in the monitoring of reactions to therapeutic drugs. Methods Inf. Med. 1971, 10, 237–245. [Google Scholar] [CrossRef]
- Gheorghe, S.; Stoica, C.; Vasile, G.G.; Nita-Lazar, M.; Stanescu, E.; Lucaciu, I.E. Metals toxic effects in aquatic ecosystems: Modulators of water quality. In Water Quality; InTech: Rijeka, Croatia, 2017; pp. 60–89. [Google Scholar]
- Adeogun, A.O.; Ogidan, I.M.; Ibor, O.R.; Chukwuka, A.V.; Adedara, I.A.; Farombi, E.O. Long-term exposure to industrial effluent induces oxidative stress and affects growth in Clarias gariepinus. Res. J. Environ. Earth Sci. 2012, 4, 738–746. [Google Scholar]
- Focks, A.; Belgers, D.; Boerwinkel, M.-C.; Buijse, L.; Roessink, I.; Van den Brink, P.J. Calibration and validation of toxicokinetic-toxicodynamic models for three neonicotinoids and some aquatic macroinvertebrates. Ecotoxicology 2018, 27, 992–1007. [Google Scholar] [CrossRef] [Green Version]
- Ashauer, R.; O’Connor, I.; Hintermeister, A.; Escher, B.I. Death dilemma and organism recovery in ecotoxicology. Environ. Sci. Technol. 2015, 49, 10136–10146. [Google Scholar] [CrossRef]
- Leist, M.; Ghallab, A.; Graepel, R.; Marchan, R.; Hassan, R.; Bennekou, S.H.; Limonciel, A.; Vinken, M.; Schildknecht, S.; Waldmann, T.; et al. Adverse outcome pathways: Opportunities, limitations and open questions. Arch. Toxicol. 2017, 91, 3477–3505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willming, M.M.; Lilavois, C.R.; Barron, M.G.; Raimondo, S. Acute toxicity prediction to threatened and endangered species using Interspecies Correlation Estimation (ICE) models. Environ. Sci. Technol. 2016, 50, 10700–10707. [Google Scholar] [CrossRef] [PubMed]
- Moermans, R.; Van Hecke, P. Effect of sample size and number of doses for the determination of LD50. J. Appl. Entomol. 1995, 119, 637–640. [Google Scholar] [CrossRef]
- Gunderson, A.R.; Stillman, J.H. Plasticity in thermal tolerance has limited potential to buffer ectotherms from global warming. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150401. [Google Scholar] [CrossRef] [Green Version]
- Magnhagen, C.; Braithwaite, V.; Forsgren, E.; Kapoor, B. Fish Behaviour; Science Publishers: Enfield, NH, USA, 2008. [Google Scholar]
- Hart, W.; Weston, R.F.; Demann, J. An apparatus for oxygenating test solutions in which fish are used as test animals for evaluating toxicity. Trans. Am. Fish. Soc. 1948, 75, 228–236. [Google Scholar] [CrossRef]
- Saha, S.; Mukherjee, D.; Saha, N.C. Evaluation of acute toxicity and behavioral responses of Heteropneustes fossilis (Linn.) exposed to Captan. Int. J. Life Sci. 2018, 6, 205–208. [Google Scholar]
- Saha, S.; Mukherjee, D.; Dhara, K.; Saha, N.C. Acute Toxicity Bioassay of a Pyrethroid Pesticide Bifenthrin to the Asian Stinging Catfish, Heteropneustes fossilis (Bloch). Curr. World Environ. 2021, 16, 250–258. [Google Scholar] [CrossRef]
- Velisek, J.; Svobodova, Z.; Piackova, V. Effects of acute exposure to bifenthrin on some haematological, biochemical and histopathological parameters of rainbow trout (Oncorhynchus mykiss). Vet. Med. 2009, 54, 131–137. [Google Scholar] [CrossRef]
- Aldridge, W. Toxicology of pyrethroids. In Mode of Action, Metabolism and Toxicology; Elsevier: Amsterdam, The Netherlands, 1983; pp. 485–490. [Google Scholar]
- Rand, G.M.; Petrocelli, S.R. Fundamentals of Aquatic Toxicology; Hemisphere Publishing: London, UK, 1985. [Google Scholar]
- Schulz, R.; Dabrowski, J.M. Combined effects of predatory fish and sublethal pesticide contamination on the behavior and mortality of mayfly nymphs. Environ. Toxicol. Chem. Int. J. 2001, 20, 2537–2543. [Google Scholar] [CrossRef]
- Saglio, P.; Olsén, K.H.; Bretaud, S. Behavioral and olfactory responses to prochloraz, bentazone, and nicosulfuron-contaminated flows in goldfish. Arch. Environ. Contam. Toxicol. 2001, 41, 192–200. [Google Scholar] [CrossRef]
- Legradi, J.B.; Di Paolo, C.; Kraak, M.H.S.; Van der Geest, H.G.; Schymanski, E.L.; Williams, A.J.; Dingemans, M.M.L.; Massei, R.; Brack, W.; Cousin, X.a.; et al. An ecotoxicological view on neurotoxicity assessment. Environ. Sci. Eur. 2018, 30, 46. [Google Scholar] [CrossRef] [PubMed]
- Paparella, M.; Bennekou, S.H.; Bal-Price, A. An analysis of the limitations and uncertainties of in vivo developmental neurotoxicity testing and assessment to identify the potential for alternative approaches. Reprod. Toxicol. 2020, 96, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Weeks Santos, S.; Cachot, J.; Cormier, B.; Mazzella, N.; Gourves, P.-Y.; Clérandeau, C.; Morin, B.; Gonzalez, P. Environmentally Relevant Mixture of Pesticides Affect Mobility and DNA Integrity of Early Life Stages of Rainbow Trout (Oncorhynchus mykiss). Toxics 2021, 9, 174. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Herrera, N.; Mena, F.; Espinoza, M.; Romero, A. Neurotoxicity of organophosphate pesticides could reduce the ability of fish to escape predation under low doses of exposure. Sci. Rep. 2019, 9, 10530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slaninova, A.; Smutna, M.; Modra, H.; Svobodova, Z. REVIEWS Oxidative stress in fish induced by pesticides. Neuroendocrinol. Lett. 2009, 30, 2. [Google Scholar] [PubMed]
- Ullah, S.; Zorriehzahra, M.J. Ecotoxicology: A review of pesticides induced toxicity in fish. Adv. Anim. Vet. Sci. 2015, 3, 40–57. [Google Scholar] [CrossRef]
- Saha, S.; Chukwuka, A.V.; Mukherjee, D.; Dhara, K.; Adeogun, A.O.; Saha, N.C. Effects of short-term sub-lethal diazinon® exposure on behavioural patterns and respiratory function in Clarias batrachus: Inferences for adaptive capacity in the wild. Chem. Ecol. 2022, 38, 180–194. [Google Scholar] [CrossRef]
- Sharma, P.; Firdous, S.; Singh, R. Neurotoxic effect of cypermethrin and protective role of resveratrol in Wistar rats. Int. J. Nutr. Pharmacol. Neurol. Dis. 2014, 4, 104. [Google Scholar]
- Groh, K.J.; Carvalho, R.N.; Chipman, J.K.; Denslow, N.D.; Halder, M.; Murphy, C.A.; Roelofs, D.; Rolaki, A.; Schirmer, K.; Watanabe, K.H. Development and application of the adverse outcome pathway framework for understanding and predicting chronic toxicity: II. A focus on growth impairment in fish. Chemosphere 2015, 120, 778–792. [Google Scholar] [CrossRef]
- Congdon, J.D.; Dunham, A.E.; Hopkins, W.A.; Rowe, C.L.; Hinton, T.G. Resource allocation-based life histories: A conceptual basis for studies of ecological toxicology. Environ. Toxicol. Chem. Int. J. 2001, 20, 1698–1703. [Google Scholar] [CrossRef]
- Masud, S.; Singh, I. Effect of Cypermethrin on some hematological parameters and prediction of their recovery in a freshwater Teleost, Cyprinus carpio. Afr. J. Environ. Sci. Technol. 2013, 7, 852–856. [Google Scholar]
- Singh, S.; Tiwari, R.K.; Pandey, R.S. Evaluation of acute toxicity of triazophos and deltamethrin and their inhibitory effect on AChE activity in Channa punctatus. Toxicol. Rep. 2018, 5, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, Y.; Tomé, H.V.; Guedes, R.N.; Oliveira, E.E. Deltamethrin toxicity and impaired swimming behavior of two backswimmer species. Environ. Toxicol. Chem. 2017, 36, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.; Philip, G. Tissues of Freshwater Fish, Cyprinus carpio Exposed to Technical Grade Cypermethrin. Bull. Environ. Contam. Toxicol. 1994, 52, 619–626. [Google Scholar] [CrossRef]
- Köprücü, K.; Seker, E. Acute toxicity of deltamethrin for freshwater mussel, Unio elongatulus eucirrus bourguignat. Bull. Environ. Contam. Toxicol. 2008, 80, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Strungaru, S.-A.; Plavan, G.; Ciobica, A.; Nicoara, M.; Robea, M.A.; Solcan, C.; Petrovici, A. Toxicity and chronic effects of deltamethrin exposure on zebrafish (Danio rerio) as a reference model for freshwater fish community. Ecotoxicol. Environ. Saf. 2019, 171, 854–862. [Google Scholar] [CrossRef]
- Ullah, S.; Li, Z.; Arifeen, M.Z.U.; Khan, S.U.; Fahad, S. Multiple biomarkers based appraisal of deltamethrin induced toxicity in silver carp (Hypophthalmichthys molitrix). Chemosphere 2019, 214, 519–533. [Google Scholar] [CrossRef]
- Abbassy, M.; Eldefrawi, M.; Eldefrawi, A. Allethrin interactions with the nicotinic acetylcholine receptor channel. Life Sci. 1982, 31, 1547–1552. [Google Scholar] [CrossRef]
- Stara, A.; Zuskova, E.; Machova, J.; Priborsky, J.; Velisek, J. Effects of acute exposure to deltamethrin and recovery time on common carp (Cyprinus carpio L.). Neuroendocrinol. Lett. 2015, 36, 133–140. [Google Scholar]
- Suvetha, L.; Ramesh, M.; Saravanan, M. Influence of cypermethrin toxicity on ionic regulation and gill Na+/K+-ATPase activity of a freshwater teleost fish Cyprinus carpio. Environ. Toxicol. Pharmacol. 2010, 29, 44–49. [Google Scholar] [CrossRef]
- Olufayo, M.; Alade, O. Acute toxicity and histological changes in gills, liver and kidney of catfish, Heterobranchus bidorsalis exposed to cypermethrin concentration. Afr. J. Agric. Res. 2012, 7, 4453–4459. [Google Scholar] [CrossRef]
- Shaluei, F.; Hedayati, A.; Kolangi, H.; Jahanbakhshi, A.; Baghfalaki, M. Evaluation of the acute toxicity of cypermethrin and its effect on behavioral responses of Caspian roach (Rutilus rutilus caspicus) and silver carp (Hypophthalmicthys molitrix). Glob. Vet. 2012, 9, 215–219. [Google Scholar]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. Int. J. 2010, 29, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.M.; Coughlan, M.T.; Cooper, M.E. Oxidative stress as a major culprit in kidney disease in diabetes. Diabetes 2008, 57, 1446–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beketov, M.A.; Liess, M. Ecotoxicology and macroecology–time for integration. Environ. Pollut. 2012, 162, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Crane, M.; Newman, M.C. What level of effect is a no observed effect? Environ. Toxicol. Chem. Int. J. 2000, 19, 516–519. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lethal Concentration | Concentration with 95% Confidence Intervals (µg/L) | ||||
|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | ||
| Oreochromis mossambicus | LC10 | 8.319 [6.442–9.347] | 8.049 [6.041–9.154] | 7.035 [5.658–7.914] | 6.052 [4.894–6.834] |
| LC20 | 9.511 [8.063–10.468] | 9.406 [7.876–10.476] | 7.985 [6.808–8.789] | 6.839 [5.809–7.563] | |
| LC30 | 10.476 [9.315–11.560] | 10.524 [9.288–11.853] | 8.748 [7.731–9.540] | 7.468 [6.547–8.169] | |
| LC40 | 11.377 [10.324–12.842] | 11.584 [10.398–13.547] | 9.457 [8.551–10.313] | 8.052 [7.217–8.765] | |
| LC50 | 12.290 [11.174–14.411] | 12.671 [11.334–15.649] | 10.172 [9.310–11.193] | 8.639 [7.860–9.417] | |
| sn | Compound | Endpoint | Mechanism | Reference |
|---|---|---|---|---|
| 1 | Cypermethrin | Respiratory distress | Reduction in red blood cells (RBCs) count and hemoglobin (Hb) value | [71] |
| 2 | Deltamethrin | Behavioral toxicity; Respiratory distress (increased opercular movements) | Nerve innervations in organs via acetylcholinesterase (AChE) inhibition in brain, muscle, and gills in fish | [72] |
| 3 | Deltamethrin | Neurotoxicity | Impairing the functions of the voltage-gated sodium channels in excitable cells and inhibiting ATPase enzymes | [73] |
| 4 | Cypermethrin | Neurotoxicity | Low activities of ATPases in brain of the fish exposed to cypermethrin suggests a decrease in the transmission of nerve impulse | [74] |
| 5 | Deltamethrin | Oxidative stress | Enhanced LPO while decreasing GSH and CAT levels, leading to tissue pathologies | [75] |
| 6 | Deltamethrin | Behavioral toxicity | Severe brain pathology, i.e., lesions of the telecenphalon in zebrafish correlated with increased rates of aggression behavior | [76] |
| 7 | Deltamethrin | Behavioral toxicity Oxidative stress | Behavioral inconsistencies via hematoxicity; i.e., changes in hematological parameters Induced oxidative stress (measured as an increase in ROS production, LPO, and antioxidant enzyme activities including SOD, POD, CAT, and GR), and increased metabolic enzyme activities (AAT, LDH, and GDH) in silver carp brain, gills, liver, and muscle tissues | [77] |
| 8 | Deltamethrin | Neurotoxicity | Altered binding receptor of nicotinic acetylcholine receptors in electroplax membranes of fish | [78] |
| 9 | Deltamethrin | Oxidative stress | Inhibited enzymatic activity of SOD, CAT, and GR, as well as the oxidative damage to the gills, liver, and kidney in fish. | [79] |
| 10 | Cypermethrin | Disrupted gill integrity | (a) Decreased ion levels in blood, as measured by specific ion concentrations (Na+, K+, and Cl−) and changes in gill Na+/K+-ATPase activity in fish | [80] |
| (b) Gill pathology | [81] | |||
| 11 | Cypermethrin | Behavioral toxicity | Abnormal swimming behavior attributed to Ache inhibition | [82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chukwuka, A.V.; Saha, S.; Mukherjee, D.; Banerjee, P.; Dhara, K.; Saha, N.C. Deltamethrin-Induced Respiratory and Behavioral Effects and Adverse Outcome Pathways (AOP) in Short-Term Exposed Mozambique Tilapia, Oreochromis mossambicus. Toxics 2022, 10, 701. https://doi.org/10.3390/toxics10110701
Chukwuka AV, Saha S, Mukherjee D, Banerjee P, Dhara K, Saha NC. Deltamethrin-Induced Respiratory and Behavioral Effects and Adverse Outcome Pathways (AOP) in Short-Term Exposed Mozambique Tilapia, Oreochromis mossambicus. Toxics. 2022; 10(11):701. https://doi.org/10.3390/toxics10110701
Chicago/Turabian StyleChukwuka, Azubuike V., Shubhajit Saha, Dip Mukherjee, Priyajit Banerjee, Kishore Dhara, and Nimai Chandra Saha. 2022. "Deltamethrin-Induced Respiratory and Behavioral Effects and Adverse Outcome Pathways (AOP) in Short-Term Exposed Mozambique Tilapia, Oreochromis mossambicus" Toxics 10, no. 11: 701. https://doi.org/10.3390/toxics10110701