

Effects of Different Concentrations of Oil Mist Particulate Matter on Pulmonary Fibrosis In Vivo and In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Highlights
- OMPM exposure induced lung lesions and PF in rats.
- OMPM exposure induced PF via TGF-β1/Smad3 and TGF-β1/p38 MAPK signaling pathways.
- OMPM exposure induced fibrotic phenotypes in the BEAS-2B cell line.
- OMPM exposure activated the differentiation of the HFL-1 cell line.
- OMPM exposure led to the BEAS-2B cell line being able to induce the differentiation of the HFL-1 cell line.
Abstract
1. Introduction
2. Materials and Methods
2.1. OMPM Collection
2.2. Animal Model and Grouping
2.3. Cell Culture and Treatment
2.4. Histopathology
2.5. Immunofluorescence
2.6. Western Blotting
2.7. ELISA Assay
2.8. Statistical Analysis
3. Results and Discussion
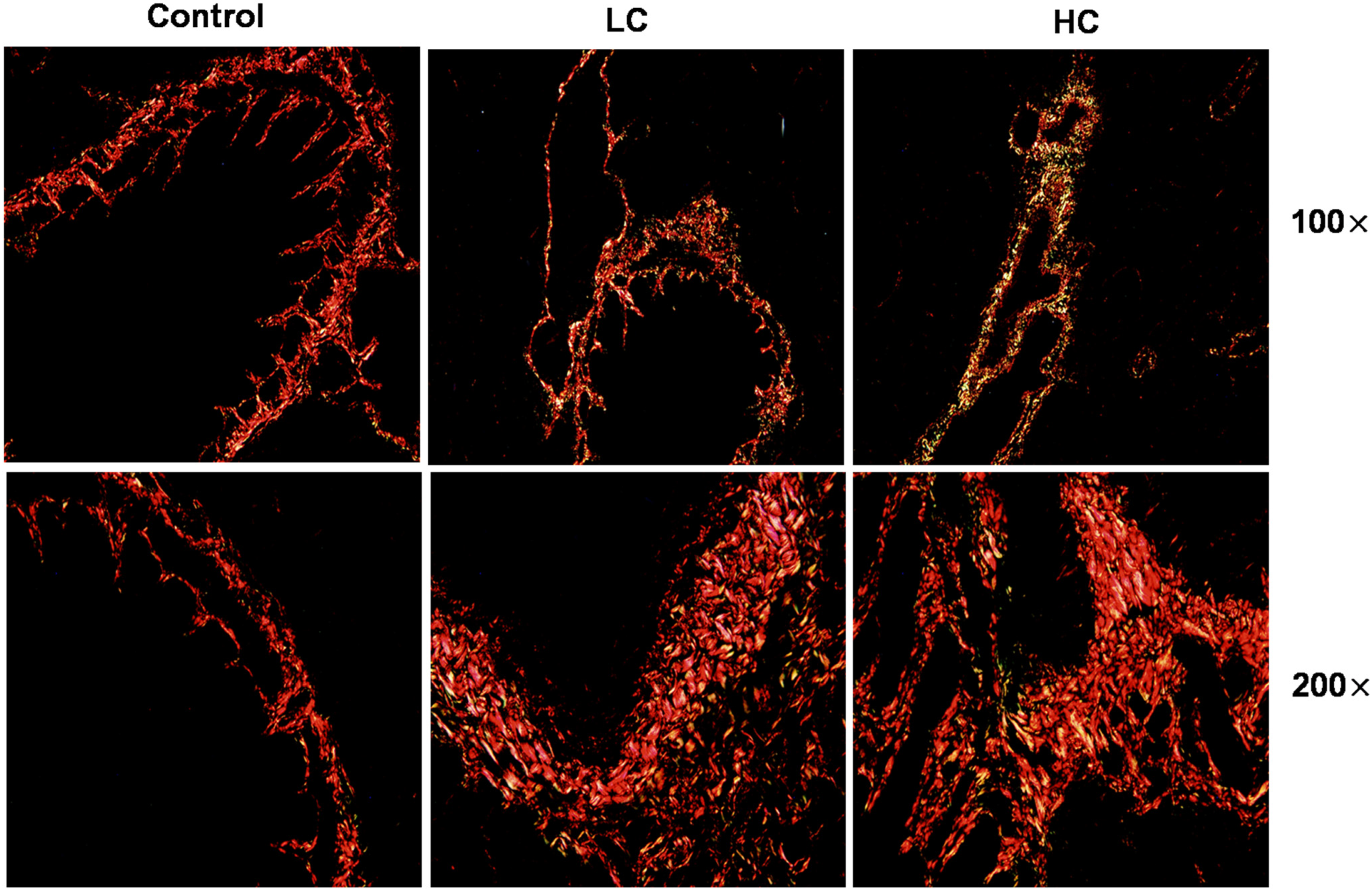
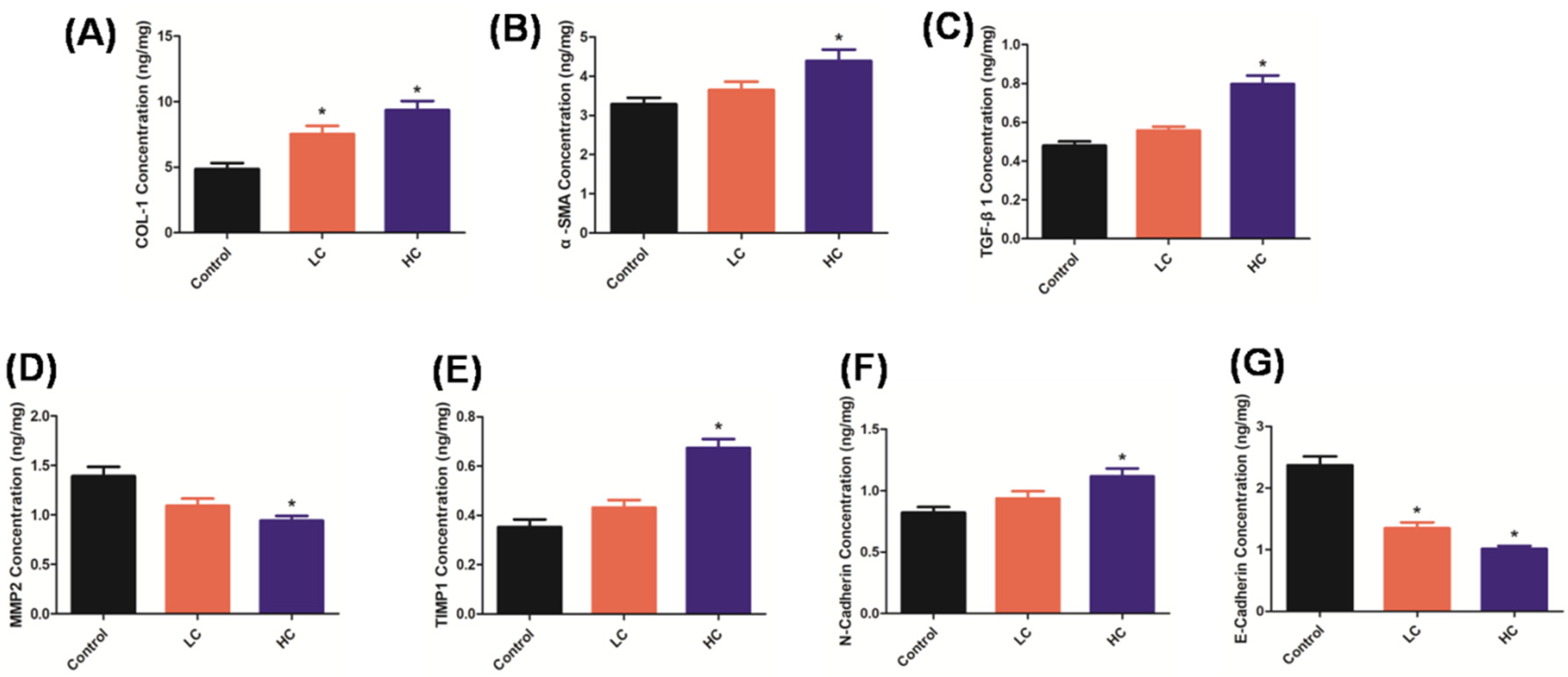
3.1. OMPM Exposure Induced Lung Lesions and PF in Rats
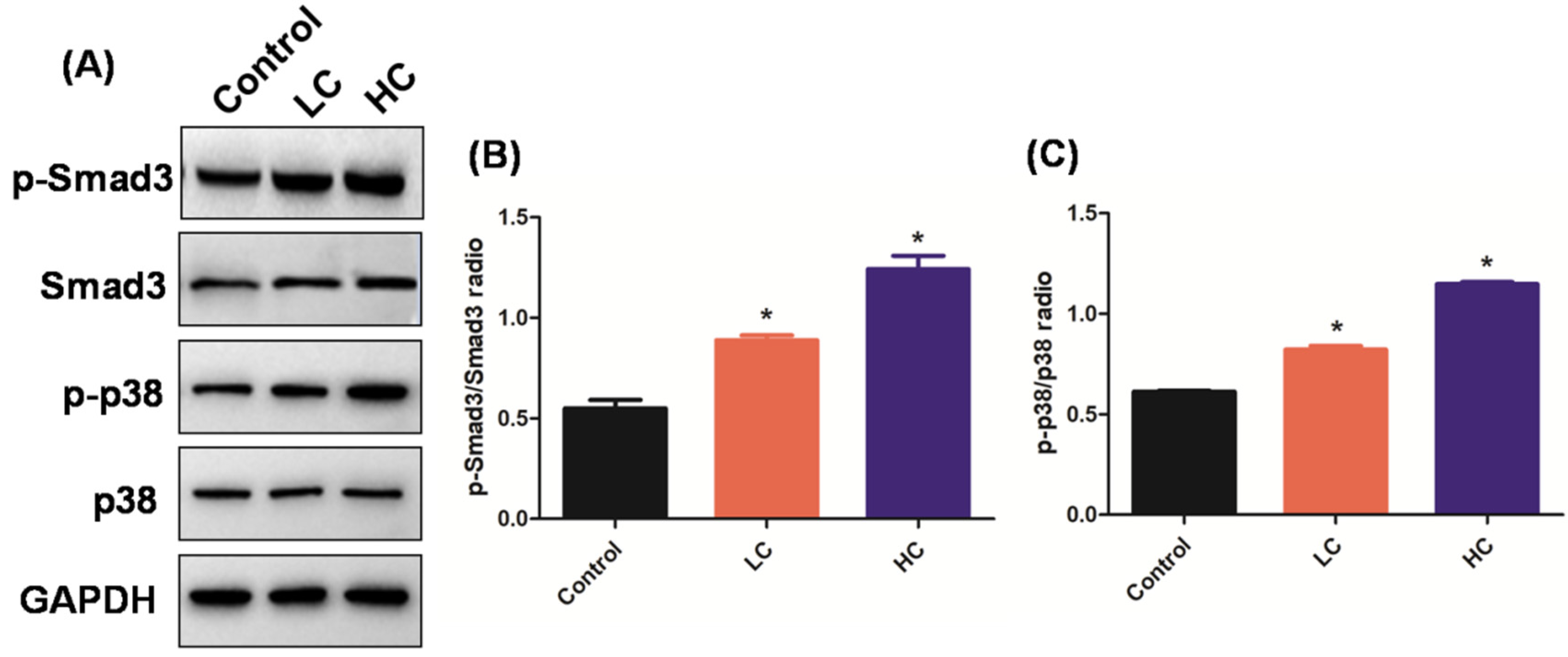
3.2. OMPM Exposure Induced PF via the TGF-β1/Smad3 and TGF-β1/p38 MAPK Signaling Pathways
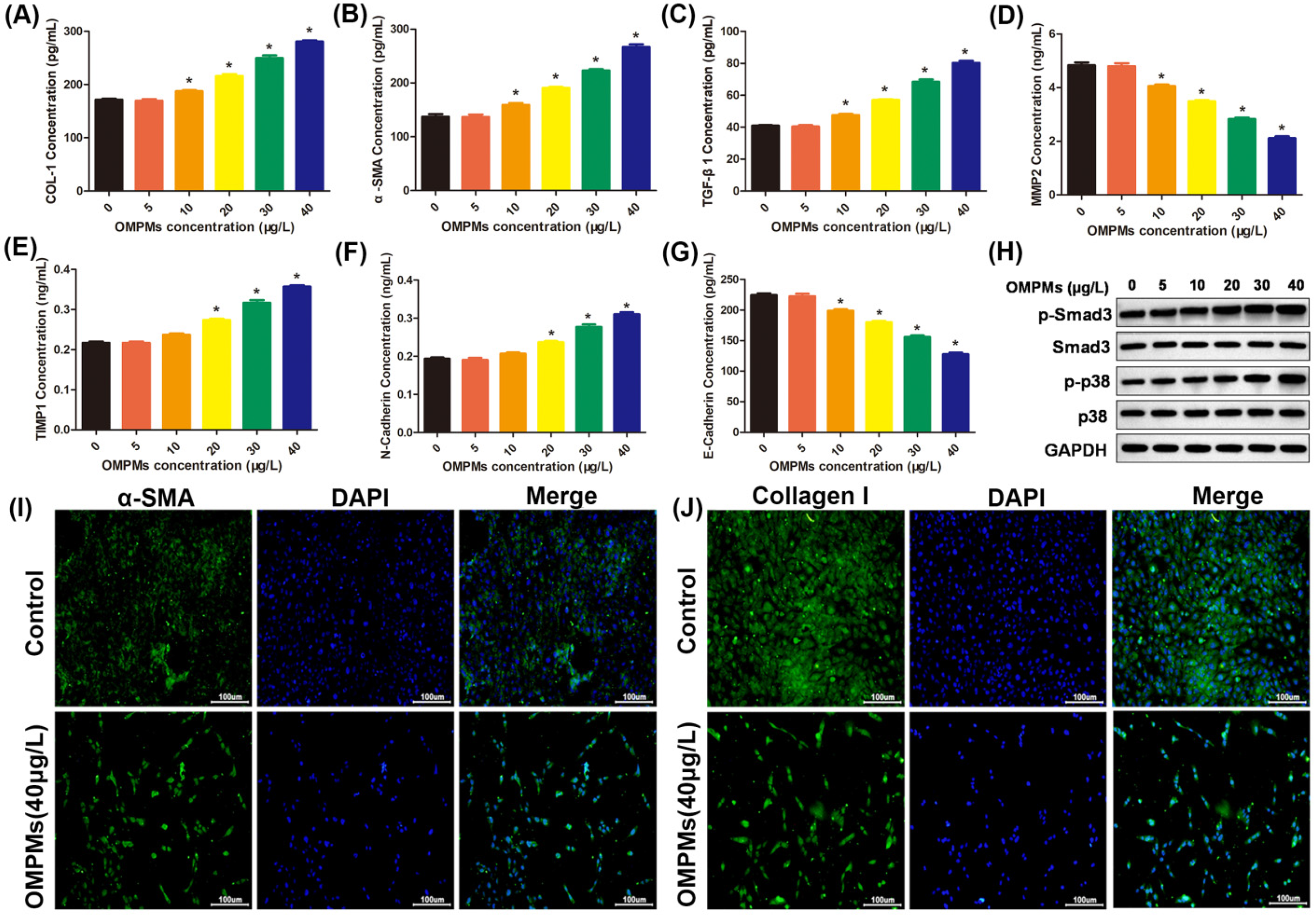
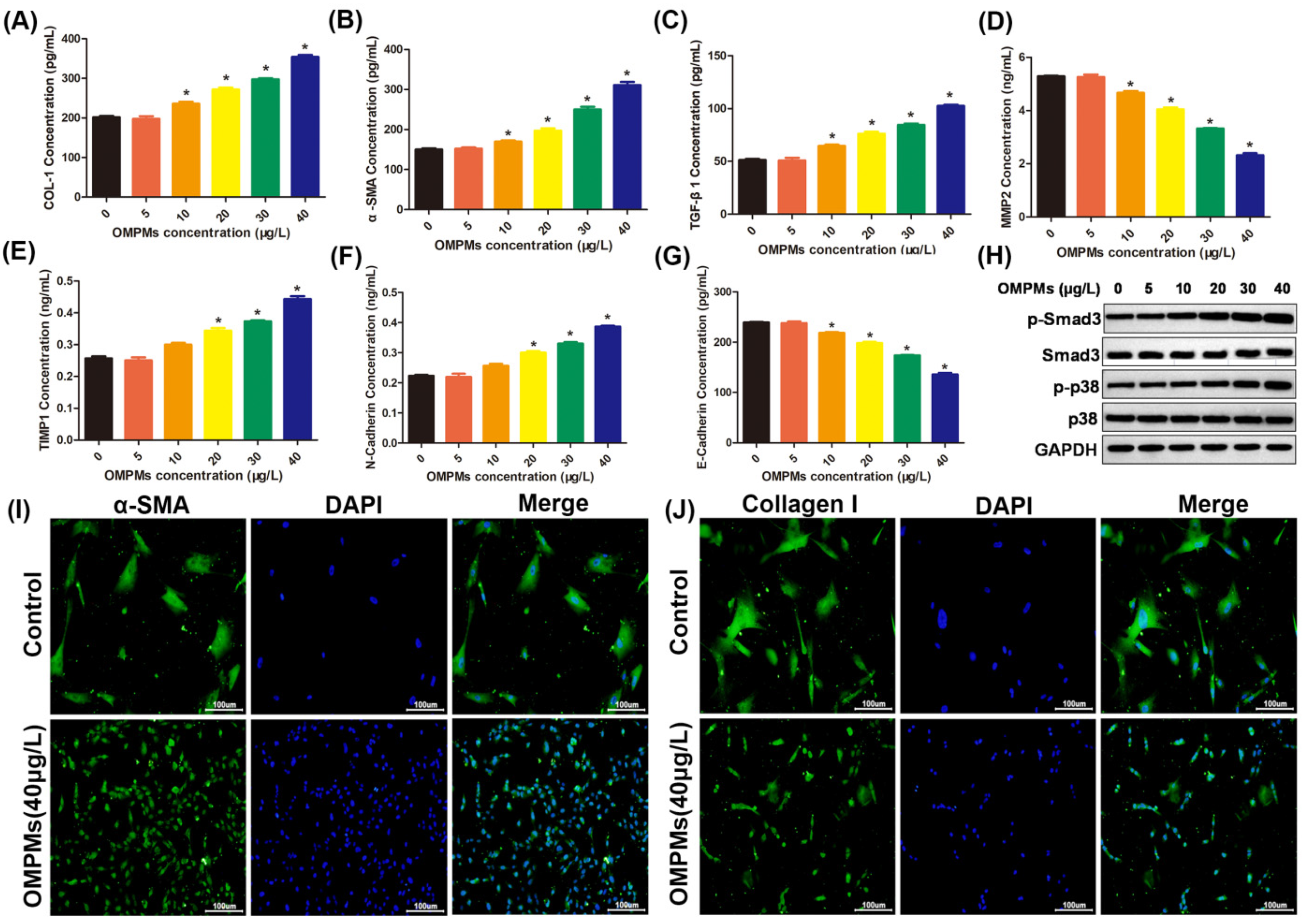
3.3. OMPM Exposure Induced Fibrotic Phenotypes in the BEAS-2B Cell Line
3.4. OMPM Exposure Activated the Differentiation of the HFL-1 Cell Line
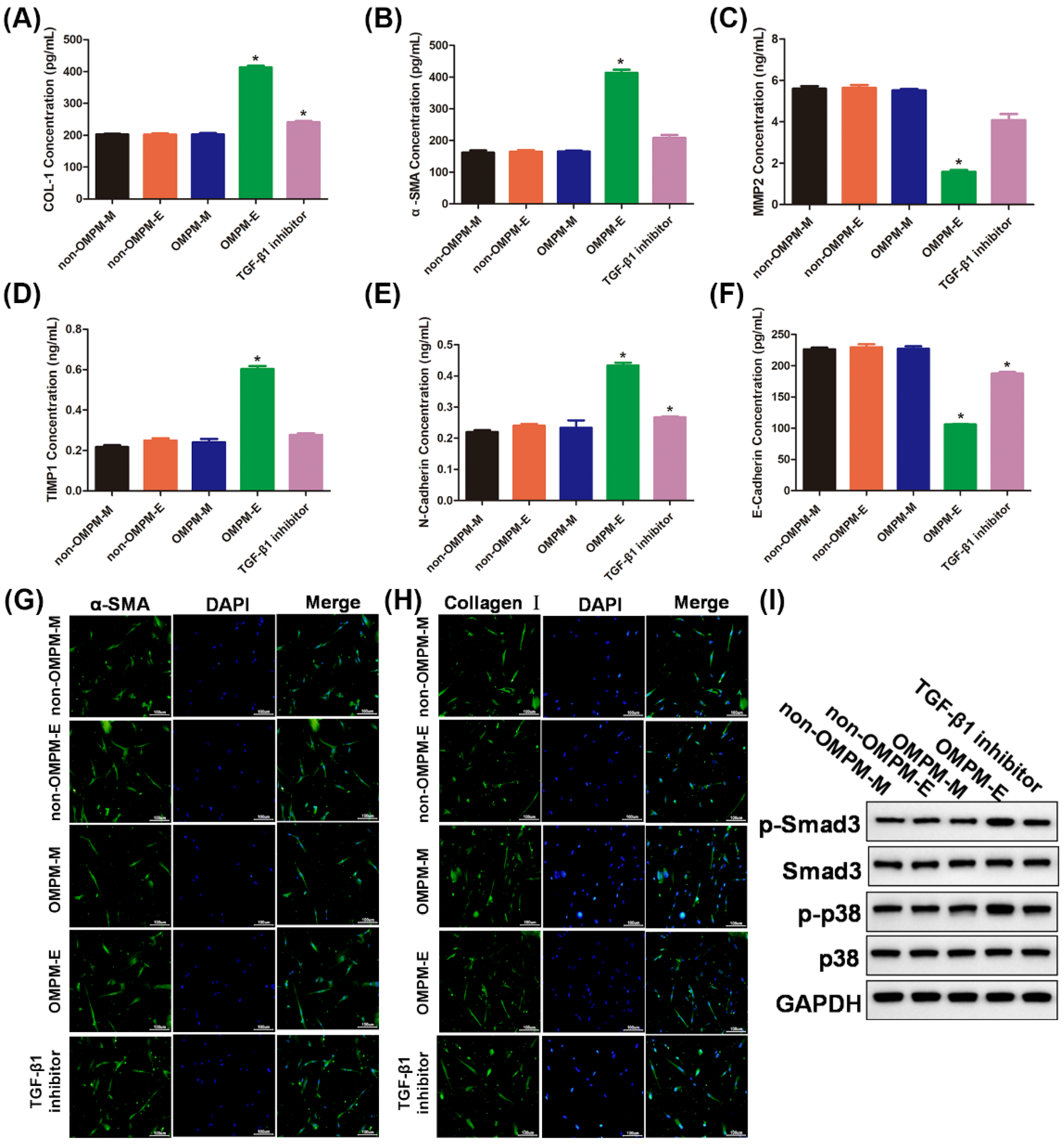
3.5. OMPM Exposed BEAS-2B Cell Line Could Induce Differentiation of HFL-1 Cell Line
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, K.F.; Tsai, Y.P.; Lai, C.H.; Xiang, Y.K.; Chuang, K.Y.; Zhu, Z.H. Human health-risk assessment based on chronic exposure to the carbonyl compounds and metals emitted by burning incense at temples. Environ. Sci. Pollut. Res. Int. 2021, 28, 40640–40652. [Google Scholar] [CrossRef] [PubMed]
- Fois, A.G.; Paliogiannis, P.; Sotgia, S.; Mangoni, A.A.; Zinellu, E.; Pirina, P.; Carru, C.; Zinellu, A. Evaluation of oxidative stress biomarkers in idiopathic pulmonary fibrosis and therapeutic applications: A systematic review. Respir. Res. 2018, 19, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, A.; Lunding, L.P.; Ehlers, J.C.; Weckmann, M.; Zissler, U.M.; Wegmann, M. More Than Just a Barrier: The Immune Functions of the Airway Epithelium in Asthma Pathogenesis. Front. Immunol. 2020, 11, 761. [Google Scholar] [CrossRef]
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Organ and tissue fibrosis: Molecular signals, cellular mechanisms and translational implications. Mol. Asp. Med. 2019, 65, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.H.; Chen, Y.C.; Lin, K.A.; Lin, Y.X.; Lee, T.H.; Lin, C.H. Adverse pulmonary impacts of environmental concentrations of oil mist particulate matter in normal human bronchial epithelial cell. Sci. Total Environ. 2022, 809, 151119. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Feng, J.; Lu, X.; Yao, Z.; Liu, Q.; Lv, Y.; Han, Y.; Deng, J.; Zhou, Y. Isorhamnetin Inhibits Liver Fibrosis by Reducing Autophagy and Inhibiting Extracellular Matrix Formation via the TGF-beta1/Smad3 and TGF-beta1/p38 MAPK Pathways. Mediat. Inflamm. 2019, 2019, 6175091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Ren, L.; Shan, K.; Guo, X.; Wang, J.; Cui, B.; An, J. Serum Inflammatory Cytokines Comparison in Gastric Cancer Therapy. Open Med.-Warsaw. 2019, 14, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, W.; Zhang, F.; Roepstorff, P.; Yang, F.; Lu, Z.; Ding, W. TMT-Based Quantitative Proteomics Analysis Reveals Airborne PM2.5-Induced Pulmonary Fibrosis. Int. J. Environ. Res. Public Health 2018, 16, 98. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.P.; McEnery, T.; McQuillan, K.; McElvaney, O.F.; McElvaney, O.J.; Landers, S.; Coleman, O.; Bussayajirapong, A.; Hawkins, P.; Henry, M.; et al. alpha1 Antitrypsin therapy modulates the neutrophil membrane proteome and secretome. Eur. Respir. J. 2020, 55, 1901678. [Google Scholar] [CrossRef]
- Deng, X.; Zhang, F.; Wang, L.; Rui, W.; Long, F.; Zhao, Y.; Chen, D.; Ding, W. Airborne fine particulate matter induces multiple cell death pathways in human lung epithelial cells. Apoptosis 2014, 19, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, S.; Dales, R.; Kauri, L.M.; Mahmud, M.; Van Ryswyk, K.; Vanos, J.; Liu, L.; Kumarathasan, P.; Thomson, E.; Vincent, R.; et al. Metal composition of fine particulate air pollution and acute changes in cardiorespiratory physiology. Environ. Pollut. 2014, 189, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Li, Z.; Liao, Z.; Gao, S.; Hua, L.; Ye, X.; Wang, Y.; Jiang, S.; Wang, N.; Zhou, D.; et al. PM2.5 induced pulmonary fibrosis in vivo and in vitro. Ecotoxicol. Environ. Saf. 2019, 171, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Pan, W.; Long, Z.; Wang, C.; Feng, Z. Study of the Oil Mist Filtration Performance: Pressure Drop Characteristics and Filter Efficiency Model. Aerosol Air Qual. Res. 2017, 17, 1063–1072. [Google Scholar] [CrossRef]
- Yin, P.; Guo, J.; Wang, L.; Fan, W.; Lu, F.; Guo, M.; Moreno, S.B.R.; Wang, Y.; Wang, H.; Zhou, M.; et al. Higher Risk of Cardiovascular Disease Associated with Smaller Size-Fractioned Particulate Matter. Environ. Sci. Technol. Lett. 2020, 7, 95–101. [Google Scholar] [CrossRef]
- Chu, H.; Xin, J.; Yuan, Q.; Zhang, X.; Pan, W.; Zeng, X.; Chen, Y.; Ma, G.; Ge, Y.; Du, M.; et al. Evaluation of vulnerable PM2.5-exposure individuals: A repeated-measure study in an elderly population. Environ. Sci. Pollut. Res. 2018, 25, 11833–11840. [Google Scholar] [CrossRef]
- Bergstra, A.D.; Brunekreef, B.; Burdorf, A. The effect of industry-related air pollution on lung function and respiratory symptoms in school children. Environ. Health 2018, 17, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camelo, A.; Dunmore, R.; Sleeman, M.A.; Clarke, D.L. The epithelium in idiopathic pulmonary fibrosis: Breaking the barrier. Front. Pharmacol. 2014, 4, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Wang, X.; Xu, L.; Zhang, H.; Li, C.; Liu, Q.; Chen, Y.; Chung, K.F.; Adcock, I.M.; Li, F. Chronic lung inflammation and pulmonary fibrosis after multiple intranasal instillation of PM2.5 in mice. Environ. Toxicol. 2021, 36, 1434–1446. [Google Scholar] [CrossRef] [PubMed]
- Gaston, B.; May, W.J.; Sullivan, S.; Yemen, S.; Marozkina, N.V.; Palmer, L.A.; Bates, J.N.; Lewis, S.J. Essential role of hemoglobin beta-93-cysteine in posthypoxia facilitation of breathing in conscious mice. J. Appl. Physiol. 2014, 116, 1290–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghatak, S.; Hascall, V.C.; Markwald, R.R.; Feghali-Bostwick, C.; Artlett, C.M.; Gooz, M.; Bogatkevich, G.S.; Atanelishvili, I.; Silver, R.M.; Wood, J.; et al. Transforming growth factor beta1 (TGFbeta1)-induced CD44V6-NOX4 signaling in pathogenesis of idiopathic pulmonary fibrosis. J. Biol. Chem. 2017, 292, 10490–10519. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Saito, S.; Sanchez, C.G.; Zhuang, Y.; Gongora Rosero, R.E.; Shan, B.; Luo, F.; Lasky, J.A. Translational Research in Acute Lung Injury and Pulmonary Fibrosis: TGF-beta1 stimulates HDAC4 nucleus-to-cytoplasm translocation and NADPH oxidase 4-derived reactive oxygen species in normal human lung fibroblasts. Am. J. Physiol. Lung Cell Mol. Physiol. 2017, 312, L936–L944. [Google Scholar] [CrossRef] [Green Version]
- de Brito, J.M.; Mauad, T.; Cavalheiro, G.F.; Yoshizaki, K.; de André, P.A.; Lichtenfels, A.J.F.C.; Guimarães, E.T.; Rivero, D.H.R.F.; Antonangelo, L.; Oliveira, L.B.; et al. Acute exposure to diesel and sewage biodiesel exhaust causes pulmonary and systemic inflammation in mice. Sci. Total Environ. 2018, 628–629, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Hewlett, J.C.; Kropski, J.A.; Blackwell, T.S. Idiopathic pulmonary fibrosis: Epithelial-mesenchymal interactions and emerging therapeutic targets. Matrix Biol. 2018, 71–72, 112–127. [Google Scholar] [CrossRef] [PubMed]
- Asker, S.; Asker, M.; Yeltekin, A.C.; Aslan, M.; Ozbay, B.; Demir, H.; Turan, H. Serum levels of trace minerals and heavy metals in severe COPD patients with and without pulmonary hypertension. Int. J. Chronic Obs. Pulm. Dis. 2018, 13, 1803–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, H.; Liu, H.; Shi, Y.; Lai, W.; Liu, X.; Xi, Z.; Lin, B. Combined multi-omics analysis reveals oil mist particulate matter-induced lung injury in rats: Pathological damage, proteomics, metabolic disturbances, and lung dysbiosis. Ecotoxicol. Environ. Saf. 2022, 241, 113759. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.; Allen, D.; Phoenix, V.R.; Le Roux, G.; Durántez Jiménez, P.; Simonneau, A.; Binet, S.; Galop, D. Atmospheric transport and deposition of microplastics in a remote mountain catchment. Nat. Geosci. 2019, 12, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Martins, M.A.; Soares, A.M.V.; Cuesta, A.; Oliveira, M. Polystyrene nanoplastics alter the cytotoxicity of human pharmaceuticals on marine fish cell lines. Environ. Toxicol. Pharmacol. 2019, 69, 57–65. [Google Scholar] [CrossRef]
- Ornatowski, W.; Lu, Q.; Yegambaram, M.; Garcia, A.E.; Zemskov, E.A.; Maltepe, E.; Fineman, J.R.; Wang, T.; Black, S.M. Complex interplay between autophagy and oxidative stress in the development of pulmonary disease. Redox Biol. 2020, 36, 101679. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Zhuang, Y.; Shan, B.; Danchuk, S.; Luo, F.; Korfei, M.; Guenther, A.; Lasky, J.A. Tubastatin ameliorates pulmonary fibrosis by targeting the TGFbeta-PI3K-Akt pathway. PLoS ONE 2017, 12, e0186615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, M.; Coppock, R.; Lindeque, P.K.; Altin, D.; Reed, S.; Pond, D.W.; Sørensen, L.; Galloway, T.S.; Booth, A.M. Effects of Nylon Microplastic on Feeding, Lipid Accumulation, and Moulting in a Coldwater Copepod. Environ. Sci. Technol. 2019, 53, 7075–7082. [Google Scholar] [CrossRef] [PubMed]
- Dawson, A.L.; Kawaguchi, S.; King, C.K.; Townsend, K.A.; King, R.; Huston, W.M.; Bengtson Nash, S.M. Turning microplastics into nanoplastics through digestive fragmentation by Antarctic krill. Nat. Commun. 2018, 9, 1001. [Google Scholar] [CrossRef]
- Zychowski, K.E.; Kodali, V.; Harmon, M.; Tyler, C.R.; Sanchez, B.; Ordonez Suarez, Y.; Herbert, G.; Wheeler, A.; Avasarala, S.; Cerrato, J.M.; et al. Respirable Uranyl-Vanadate-Containing Particulate Matter Derived From a Legacy Uranium Mine Site Exhibits Potentiated Cardiopulmonary Toxicity. Toxicol. Sci. 2018, 164, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Q.; Zhang, C.Z.; Shao, B.; Pang, D.H.; Han, G.Z.; Lin, L. Effects of abnormal expression of fusion and fission genes on the morphology and function of lung macrophage mitochondria in SiO2-induced silicosis fibrosis in rats in vivo. Toxicol. Lett. 2019, 312, 181–187. [Google Scholar] [CrossRef]
- Hahladakis, J.N.; Velis, C.A.; Weber, R.; Iacovidou, E.; Purnell, P. An overview of chemical additives present in plastics: Migration, release, fate and environmental impact during their use, disposal and recycling. J. Hazard. Mater. 2018, 344, 179–199. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.S.; Jimenez-Martinez, J.; Mitrano, D.M. Transport of Nano- and Microplastic through Unsaturated Porous Media from Sewage Sludge Application. Environ. Sci. Technol. 2020, 54, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.; Huang, Q.; Lin, Y.; Ye, G.; Zhu, H.; Dong, S. Epithelial-mesenchymal transition effect of fine particulate matter from the Yangtze River Delta region in China on human bronchial epithelial cells. J. Environ. Sci. 2018, 66, 155–164. [Google Scholar] [CrossRef]
- Dysart, M.M.; Galvis, B.R.; Russell, A.G.; Barker, T.H. Environmental particulate (PM2.5) augments stiffness-induced alveolar epithelial cell mechanoactivation of transforming growth factor beta. PLoS ONE 2014, 9, e106821. [Google Scholar] [CrossRef] [Green Version]
- Leppänen, I.; Lappalainen, T.; Lohtander, T.; Jonkergouw, C.; Arola, S.; Tammelin, T. Capturing colloidal nano- and microplastics with plant-based nanocellulose networks. Nat. Commun. 2022, 13, 1814. [Google Scholar] [CrossRef]
- Malafaia, G.; de Souza, A.M.; Pereira, A.C.; Gonçalves, S.; da Costa Araújo, A.P.; Ribeiro, R.X.; Rocha, T.L. Developmental toxicity in zebrafish exposed to polyethylene microplastics under static and semi-static aquatic systems. Sci. Total Environ. 2020, 700, 134867. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, H.; Liu, H.; Shi, Y.; Lai, W.; Liu, X.; Xi, Z.; Lin, B. Effects of Different Concentrations of Oil Mist Particulate Matter on Pulmonary Fibrosis In Vivo and In Vitro. Toxics 2022, 10, 647. https://doi.org/10.3390/toxics10110647
Nie H, Liu H, Shi Y, Lai W, Liu X, Xi Z, Lin B. Effects of Different Concentrations of Oil Mist Particulate Matter on Pulmonary Fibrosis In Vivo and In Vitro. Toxics. 2022; 10(11):647. https://doi.org/10.3390/toxics10110647
Chicago/Turabian StyleNie, Huipeng, Huanliang Liu, Yue Shi, Wenqing Lai, Xuan Liu, Zhuge Xi, and Bencheng Lin. 2022. "Effects of Different Concentrations of Oil Mist Particulate Matter on Pulmonary Fibrosis In Vivo and In Vitro" Toxics 10, no. 11: 647. https://doi.org/10.3390/toxics10110647