Use of a Selected Leuconostoc Citreum Strain as a Starter for Making a “Yeast-Free” Bread

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Selection of Lactic Acid Bacteria
2.3. Molecular Characterization of LAB Strains
2.4. Preparation of Liquid Sourdoughs and Bread-Making at Laboratory Scale
2.5. Biochemical Characterization of Laboratory Sourdoughs and Bread Doughs
2.6. Microbiological Analyses and Monitoring of Selected Strains
2.7. Identification of LAB and Yeasts
2.8. Characterization of the Laboratory Breads
2.9. Effect of Salt on Starter Performances
2.10. Bread-Making at a Pilot Plant Scale
2.11. Sensory Analysis of Puccia Bread
2.12. Statistical Analysis
3. Results
3.1. Selection of the Starters
3.2. Genotypic Characterization of LAB Strains
3.3. Bread-Making at Laboratory Scale
3.3.1. Characterization of Liquid Sourdoughs and Bread Doughs
3.3.2. Microbiological and Molecular Characterization of LAB and Yeasts
3.3.3. Bread characterization
3.4. Effect of Salt Addition
3.5. Bread-Making at the Industrial Pilot Scale
3.5.1. Characterization of Sourdough and Puccia Doughs
3.5.2. Microbiological Analyses
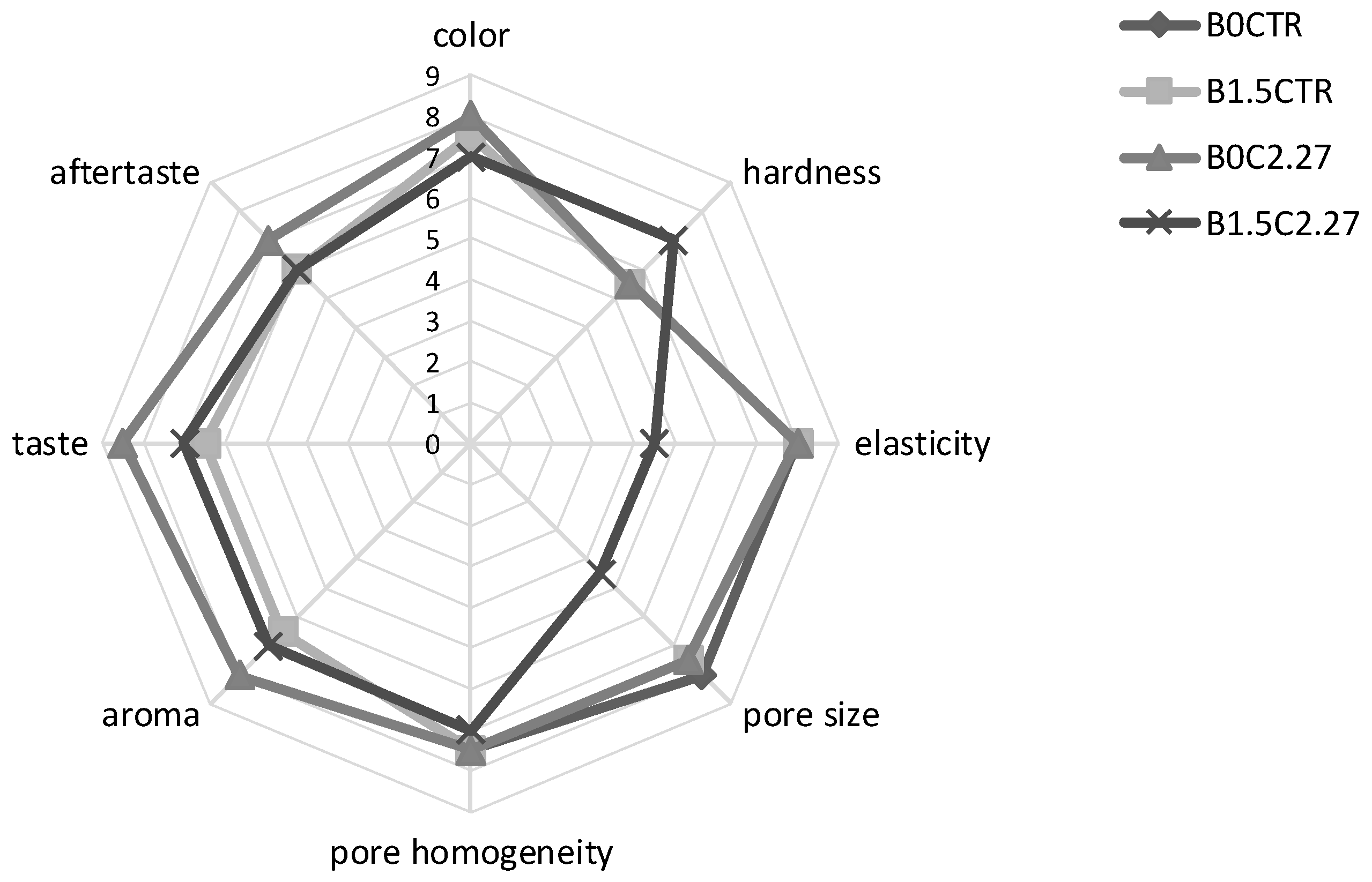
3.5.3. Puccia Bread Characterization
4. Discussion
4.1. Selection of the Starter Based on Technological and Microbiological Properties
4.2. Bread-Making at Industrial Pilot Scale
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shakoor, Z.; Al Faifi, A.; Al Amro, B.; Al Tawil, L.M.; Al Ohaly, R.Y. Prevalence of IgG-mediated food intolerance among patients with allergic symptoms. Ann. Saudi Med. 2016, 36, 386–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi, M.; Perricone, R.; Blank, M.; Perricone, C.; Shoenfeld, Y. Anti-Saccharomyces cerevisiae autoantibodies in autoimmune diseases: From bread baking to autoimmunity. Clin. Rev. Allergy Immunol. 2013, 45, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Lied, G.A.; Lillestøl, K.; Valeur, J.; Berstad, A. Intestinal B cell-activating factor: An indicator of non-IgE-mediated hypersensitivity reactions to food? Aliment. Pharmacol. Ther. 2010, 32, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.; Krause, I. Anti-Saccharomyces cerevisiae Antibodies Autoimmune Diseases. In Infection and Autoimmunity, 2nd ed.; Shoenfeld, Y., Agmon-Levin, N., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2015; pp. 671–679. [Google Scholar]
- Triggs, C.M.; Munday, K.; Hu, R.; Fraser, A.G.; Gearry, R.B.; Barclay, M.L.; Ferguson, L.R. Dietary factors in chronic inflammation: Food tolerances and intolerances of a New Zealand Caucasian Crohn’s disease population. Mutat. Res. 2010, 690, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Muratori, P.; Muratori, L.; Guidi, M.; Maccariello, S.; Pappas, G.; Ferrari, R.; Bianchi, F.B. Anti-Saccharomyces cerevisiae antibodies (ASCA) and autoimmune liver diseases. Clin. Exp. Immunol. 2003, 132, 473–476. [Google Scholar] [CrossRef]
- Salamati, S.; Martins, C.; Kulseng, B. Baker’s yeast (Saccharomyces cerevisiae) antigen in obese and normal weight subjects. Clin. Obes. 2014, 5, 42–47. [Google Scholar] [CrossRef]
- Musatti, A.; Mapelli, C.; Foschino, R.; Picozzi, C.; Rollini, M. Unconventional bacterial association for dough leavening. Int. J. Food Microbiol. 2016, 237, 28–34. [Google Scholar] [CrossRef]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Technol. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Corsetti, A. Technology of sourdough fermentation and sourdough applications. In Handbook on Sourdough Biotechnology; Gänzle, M., Gobbetti, M., Eds.; Springer: New York, NY, USA, 2013; pp. 85–103. [Google Scholar]
- Choi, H.; Kim, Y.W.; Hwang, I.; Kim, J.; Yoon, S. Evaluation of Leuconostoc citreum HO12 and Weissella koreensis HO20 isolated from kimchi as a starter culture for whole wheat sourdough. Food Chem. 2012, 134, 2208–2216. [Google Scholar] [CrossRef]
- Corona, O.; Alfonzo, A.; Ventimiglia, G.; Nasca, A.; Francesca, N.; Martorana, A.; Moschetti, G.; Settani, L. Industrial application of selected lactic acid bacteria isolated from local semolinas for typical sourdoughs bread production. Food Microbiol. 2016, 59, 43–56. [Google Scholar] [CrossRef]
- Mamhoud, A.; Nionelli, L.; Bouzaine, T.; Hamdi, M.; Gobbetti, M.; Rizzello, C.G. Selection of lactic acid bacteria isolated from Tunisian cereals and exploitation of the use as starters for sourdough fermentation. Int. J. Food Microbiol. 2016, 225, 9–19. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Lorusso, A.; Montemurro, M.; Gobbetti, M. Use of sourdough made with quinoa (Chenopodium quinoa) flour and autochthonous selected lactic acid bacteria for enhancing the nutritional, textural and sensory features of white bread. Food Microbiol. 2016, 56, 1–13. [Google Scholar] [CrossRef]
- Ventimiglia, G.; Alfonzo, A.; Galluzzo, P.; Corona, O.; Francesca, N.; Caracappa, S.; Moschetti, G.; Settanni, L. Codominance of Lactobacillus plantarum and obligate heterofermentative lactic acid bacteria during sourdough fermentation. Food Microbiol. 2015, 51, 57–68. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Van Kerrebroeck, S.; Harth, H.; Huys, G.; Daniel, H.M.; Weckx, S. Microbial ecology of sourdough fermentations: Diverse or uniform? Food Microbiol. 2014, 37, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, P.; Ciati, R.; Leporati, A.; Paese, M. Liquid sourdough fermentation: Industrial application perspectives. Food Microbiol. 2007, 24, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, A.; Lavermicocca, P.; Morea, M.; Baruzzi, F.; Tosti, N.; Gobbetti, M. Phenotypic and molecular identification and clustering of lactic acid bacteria and yeasts from wheat (species Triticum durum and Triticum aestivum) sourdoughs of Southern Italy. Int. J. Food Microbiol. 2001, 64, 95–104. [Google Scholar] [CrossRef]
- Valerio, F.; Favilla, M.; De Bellis, P.; Sisto, A.; De Candia, S.; Lavermicocca, P. Antifungal activity of strains of lactic acid bacteria isolated from a semolina ecosystem against Penicillium roqueforti, Aspergillus niger and Endomyces fibuliger contaminating bakery products. Syst. Appl. Microbiol. 2009, 32, 438–448. [Google Scholar] [CrossRef]
- Minervini, F.; Pinto, D.; Di Cagno, R.; De Angelis, M.; Gobbetti, M. Scouting the application of sourdough to frozen dough bread technology. J. Cereal Sci. 2011, 54, 296–304. [Google Scholar] [CrossRef]
- De Bellis, P.; Valerio, F.; Sisto, A.; Lonigro, S.L.; Lavermicocca, P. Probiotic table olives: Microbial populations adhering on olive surface in fermentation sets inoculated with the probiotic strain Lactobacillus paracasei IMPC2.1 in an industrial plant. Int. J. Food Microbiol. 2010, 140, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Valerio, F.; Conte, A.; Di Biase, M.; Lattanzio, V.M.T.; Lonigro, L.S.; Padalino, L.; Pontonio, E.; Lavermicocca, P. Formulation of yeast-leavened bread with reduced salt content by using a Lactobacillus plantarum fermentation product. Food Chem. 2017, 221, 582–589. [Google Scholar] [CrossRef]
- Weiss, W.; Vogelmeier, C.; Gorg, A. Electrophoretic characterization of wheat grain allergens from different cultivars involved in bakers’asthma. Electrophoresis 1993, 14, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for determination of Proteolysis in Milk and Isolated Milk Proteins. J. Diary Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; De Angelis, M.; Gobbetti, M. Effect of sourdough fermentation on stabilisation, and chemical and nutritional characteristics of wheat germ. Food Chem. 2010, 119, 1079–1089. [Google Scholar] [CrossRef]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA gene sequence analysis and multiplex PCR assay with recA gene-derived primers. Appl. Environ. Microbiol. 2001, 67, 3450–3454. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.; Canchaya, C.; Meylan, V.; Klaenhammer, T.R.; Zink, R. Analysis, characterization, and loci of the tuf genes in Lactobacillus and Bifidobacterium species and their direct application for species identification. Appl. Environ. Microbiol. 2003, 69, 6908–6922. [Google Scholar] [CrossRef]
- Gadanho, M.; Almeida, J.M.G.C.F.; Sampaio, J.P. Assessment of yeast diversity in a marine environment in the south of Portugal by microsatellite-primed PCR. Antonie Leeuwenhoek 2003, 84, 217–227. [Google Scholar] [CrossRef]
- Valerio, F.; De Bellis, P.; Di Biase, M.; Lonigro, S.L.; Giussani, B.; Visconti, A.; Lavermicocca, P.; Sisto, A. Diversity of spore-forming bacteria and identification of Bacillus amyloliquefaciens as a species frequently associated with the ropy spoilage of bread. Int. J. Food Microbiol. 2012, 156, 278–285. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Crowley, P.; Grau, H.; Arendt, E.K. Influence of additives and mixing time on crumb grain characteristics of wheat bread. Cereal Chem. 2000, 77, 370–375. [Google Scholar] [CrossRef]
- Corsetti, A.; Settani, L. Lactobacilli in sourdough fermentation. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- Spicher, G. Baked goods. In Biotechnology; Rehm, H.J., Reed, G., Eds.; Verlag Chemie: Weinheim, Germany, 1983; pp. 1–80. [Google Scholar]
- Settanni, L.; Ventimiglia, G.; Alfonzo, A.; Corona, O.; Miceli, A.; Moschetti, G. An integrated technological approach to the selection of lactic acid bacteria of flour origin for sourdough production. Food Res. Int. 2013, 54, 1569–1578. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; Harth, H.; Kerrebroeck, S.V.; Leroy, F. Yeast diversity of sourdoughs and associated metabolic properties and functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Lattanzi, A.; De Angelis, M.; Celano, G.; Gobbetti, M. House microbiotas as sources of lactic acid bacteria and yeasts in traditional Italian sourdoughs. Food Microbiol. 2015, 52, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; De Angelis, M.; Di Cagno, R.; Gobbetti, M. Ecological parameters influencing microbial diversity and stability of traditional sourdough. Int. J. Food Microbiol. 2014, 171, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Carr, F.J.; Chill, D.; Maida, N. The Lactic Acid Bacteria: A Literature Survey. Crit. Rev. Microbiol. 2002, 28, 281–370. [Google Scholar] [CrossRef] [PubMed]
- Pétel, C.; Onno, B.; Prost, C. Sourdough volatile compounds and their contribution to bread: A review. Trends Food Sci. Technol. 2017, 59, 105–123. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Type of Fermentation | Culture Collection | Isolation Source | Culture Medium |
|---|---|---|---|---|---|
| Lb. plantarum | 21B [18] | FHe | ISPA-CNR | Sourdough | MRS |
| Lc. lactis | C5.6 [19] | OHo | ISPA-CNR | Durum wheat semolina | mMRS [18] |
| W. confusa | C5.7 [19] | OHe | ISPA-CNR | Durum wheat semolina | MRS |
| L. citreum | C2.27 [19] | OHe | ISPA-CNR | Durum wheat semolina | MRS |
| W. cibaria | C21.4 [19] | OHe | ISPA-CNR | Durum wheat semolina | mMRS |
| Lb. brevis | 18F | OHe | ISPA-CNR | Sourdough | MRS |
| P. pentosaceus | LM | FHe | ISPA-CNR | Durum wheat semolina | MRS |
| Lb. plantarum | DC400 | FHe | DiSSPA | Sourdough | MRS |
| Lc. lactis | 10γ | OHo | DiSSPA | Sourdough | mMRS |
| W. cibaria | 5S | OHe | DiSSPA | Sourdough | mMRS |
| Lb. rossiae | DSM15814 | OHe | DSMZ | Sourdough | mMRS |
| W. confusa | DSM20196 | OHe | DSMZ | Sugar cane | MRS |
| Species | Strain | ΔpH * | ΔV | TTA | ΔpH | ΔV | TTA | ΔpH | ΔV | TTA |
|---|---|---|---|---|---|---|---|---|---|---|
| 8 h | 16 h | 24 h | ||||||||
| Lb. plantarum | 21B | 2.10 ± 0.01 e | 0 a | 4.33 ± 0.17 cde | 2.32 ± 0.01 d | 0 a | 7.87 ± 0.13 ef | 2.44 ± 0.04 de | 0 a | 9 ± 0.3 fg |
| Lc. lactis | C5.6 | 1.72 ± 0.01 d | 0 a | 3.50 ± 0.29 abc | 1.84 ± 0.00 a | 0 a | 3.33 ± 0.33 a | 1.97 ± 0.01 a | 0 a | 4.67 ± 0.33 a |
| W. confusa | C5.7 | 1.68 ± 0.02 cd | 7.50 ± 0.00 d | 4.17 ± 0.17 bcde | 1.95 ± 0.03 ab | 8.75 ± 0.72 b | 6.00 ± 0.00 bc | 2.02 ± 0.02 a | 10.75 ± 1.01 b | 5.00 ± 0.58 ab |
| L. citreum | C2.27 | 2.06 ± 0.01 e | 9.83 ± 0.17 e | 4.67 ± 0.33 de | 2.28 ± 0.02 d | 13.90 ± 2.31 cd | 9.00 ± 0.00 f | 2.37 ± 0.03 cde | 17.17 ± 1.67 c | 9.67 ± 0.17 g |
| W. cibaria | C21.4 | 1.73 ± 0.10 d | 6.50 ± 0.00 d | 4.83 ± 0.44 de | 2.10 ± 0.05 bc | 8.00 ± 0.58 b | 6.50 ± 0.29 bcd | 2.05 ± 0.02 a | 7.75 ± 0.14 b | 6.33 ± 0.33 bcd |
| Lb. brevis | 18F | 0.80 ± 0.07 a | 1.33 ± 0.17 b | 3.77 ± 0.27 abcd | 1.84 ± 0.08 a | 11.10 ± 0.74 bc | 5.63 ± 0.20 bc | 2.24 ± 0.02 b | 17.33 ± 0.88 c | 7.27 ± 0.14 de |
| P. pentosaceus | LM | 1.76 ± 0.00 d | 0 a | 2.83 ± 0.17 a | 2.26 ± 0.01 d | 0 a | 7.33 ± 0.33 de | 2.34 ± 0.01 bcd | 0 a | 7.17 ± 0.17 de |
| Lb. plantarum | DC400 | 1.63 ± 0.02 cd | 0 a | 3.00 ± 0.00 a | 2.34 ± 0.01 d | 0 a | 5.33 ± 0.33 b | 2.40 ± 0.01 cde | 0 a | 8.00 ± 0.58 ef |
| Lc. lactis | 10γ | 1.68 ± 0.01 cd | 0 a | 3.33 ± 0.17 abc | 2.22 ± 0.01 cd | 0 a | 6.50 ± 0.29 bcd | 2.31 ± 0.01 bc | 0 a | 6.83 ± 0.16 cde |
| W. cibaria | 5S | 1.29 ± 0.05 b | 2.50 ± 0.00 c | 3.17 ± 0.17 ab | 2.33 ± 0.01 d | 17.50 ± 0.00 de | 8.17 ± 0.17 ef | 2.45 ± 0.03 e | 19.25 ± 0.14 c | 9.00 ± 0.00 fg |
| Lb. rossiae | DSM 15814 | 1.48 ± 0.10 bc | 2.50 ± 0.00 c | 3.00 ± 0.00 a | 2.28 ± 0.01 d | 18.25 ± 0.72 e | 6.67 ± 0.17 cd | 2.47 ± 0.02 e | 20.00 ± 0.00 c | 9.00 ± 0.00 fg |
| W. confusa | DSM 20196 | 1.61 ± 0.01 cd | 8.75 ± 0.72 e | 5.17 ± 0.17 e | 2.09 ± 0.02 bc | 11.00 ± 0.29 bc | 5.83 ± 0.17 bc | 2.07 ± 0.02 a | 10.00 ± 0.00 b | 5.33 ± 0.33 abc |
| Characteristics | Liquid Sourdough | Bread Dough | |||
|---|---|---|---|---|---|
| SC2.27 | SC5.7 | DC2.27 | DC5.7 | DCTR | |
| pH | 3.48 ± 0.05 a | 3.56 ± 0.05 a | 4.29 ± 0.05 b | 4.30 ± 0.04 b | 5.18 ± 0.22 a |
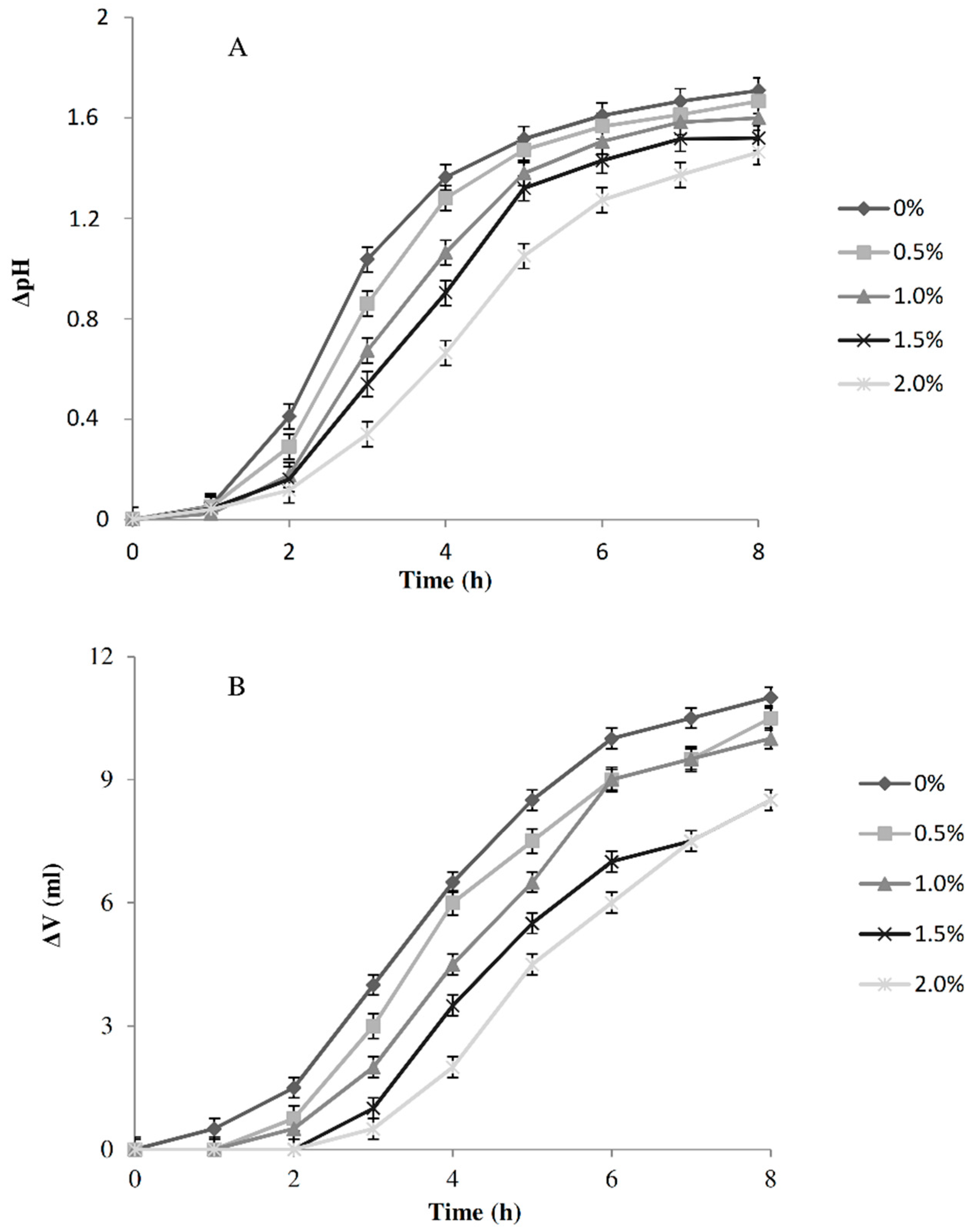
| ΔpH | 2.69 ± 0.07 a | 2.60 ± 0.09 a | 0.89 ± 0.05 a | 0.92 ± 0.07 a | 0.30 ± 0.07 b |
| ΔV (mL) | - | - | 8.10 ± 0.24 b | 4.80 ± 0.59 b | 20.42 ± 2.45 a |
| TTA (mL) | 4.10 ± 0.10 a | 3.60 ± 0.10 a | 5.80 ± 0.43 a | 4.70 ± 0.44 a | 1.75 ± 0.02 b |
| Lactic acid (mmol/kg) | 6.53 ± 0.16 a | 4.27 ± 0.34 b | 13.68 ± 0.42 a | 10.79 ± 0.24 b | ND |
| Acetic acid (mmol/kg) | 2.17 ± 0.04 a | 0.92 ± 0.07 b | 6.48 ± 0.24 | ND | ND |
| QF | 3.01 ± 0.10 a | 4.77 ± 0.77 a | 2.11 ± 0.11 | - | - |
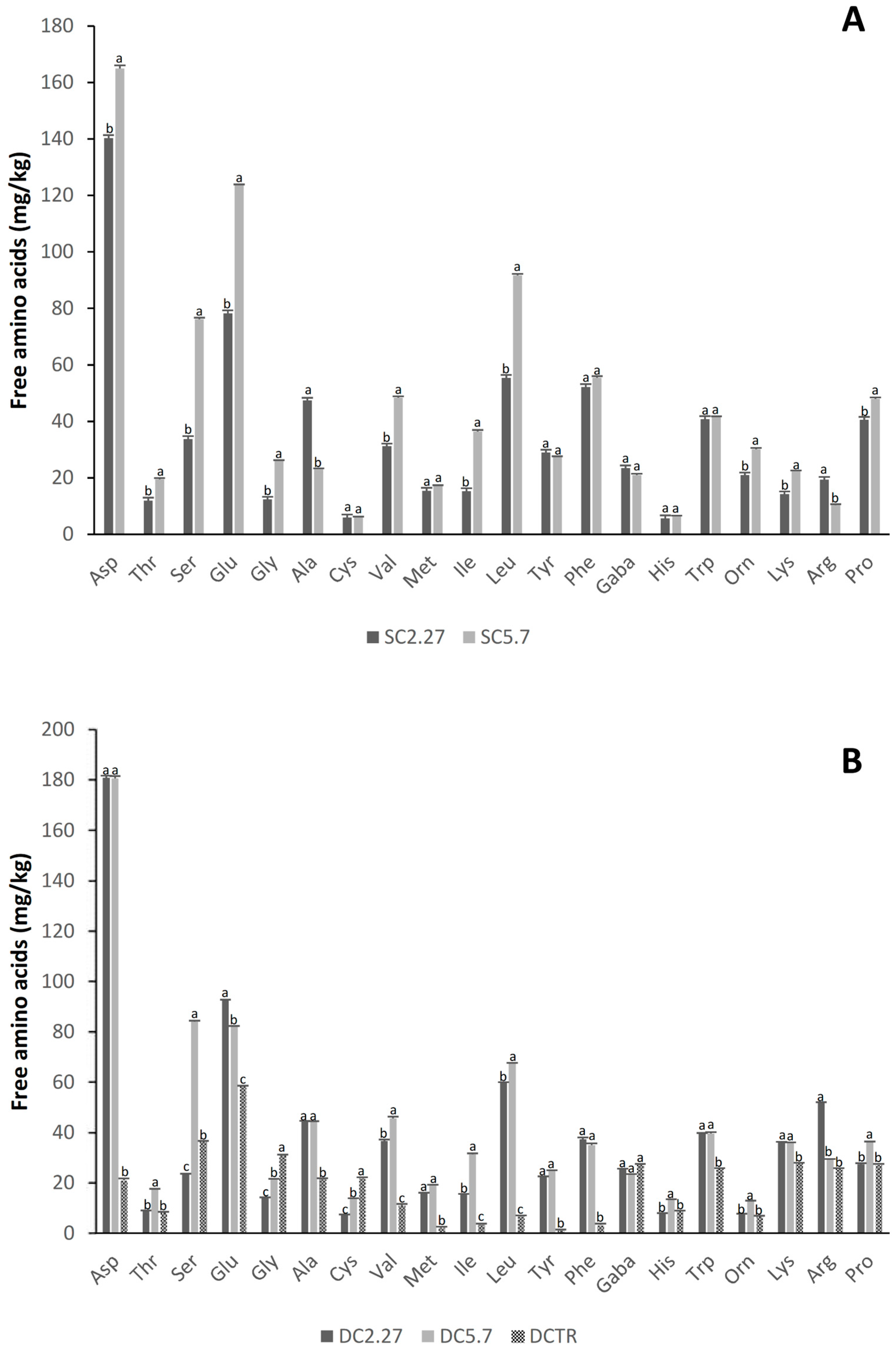
| Peptides (g/kg) | 7.36 ± 0.31 b | 7.92 ± 0.11 a | 9.46 ± 0.11 a | 9.76 ± 0.68 a | 4.25 ± 0.54 b |
| Total Free Amino Acids (mg/kg) | 693 ± 16 b | 894 ± 17 a | 753 ± 7 b | 856 ± 8 a | 428 ± 6 c |
| Sample | LAB (log cfu/g) | Starter Strain (log cfu/g) | Other LAB Species § | Yeasts (log cfu/g) | Moulds (log cfu/g) | Yeast Species |
|---|---|---|---|---|---|---|
| SC2.27 | 8.94 ± 0.28 a | 8.94 ± 0.28 a | - | - | 0.60 ± 0.09 a | - |
| SC5.7 | 8.43 ± 0.24 a | 8.27 ± 0.28 a | Lb. plantarum (1) * | - | 0.90 ± 0.21 a | - |
| DC2.27 | 8.83 ± 0.17 a | 8.83 ± 0.17 a | - | - | - | - |
| DC5.7 | 8.63 ± 0.17 a | 6.88 ± 1.83 a | Lb. plantarum (1) * | 1.80 ± 0.40 b | 1.79 ± 0.79 | F. magnum/F. floriforme (1) |
| DCTR | 5.57 ± 0.23 b | - | Lb. sakei (1) | 7.99 ± 0.23 a | - | S. cerevisiae (1) |
| Lb. curvatus (2) | ||||||
| L. gelidum (1) | ||||||
| L. pseudomesenteroides (1) | ||||||
| Lb. brevis (1) | ||||||
| Lb. paraplantarum (2) | ||||||
| Lb. paracasei (1) |
| Characteristics of Breads | Bread | ||
|---|---|---|---|
| BC2.27 | BC5.7 | BCTR | |
| pH | 4.48 ± 0.04 b | 4.61 ± 0.001 b | 5.52 ± 0.15 a |
| TTA (mL) | 5.80 ± 0.42 a | 3.06 ± 0.15 b | 1.15 ± 0.01 c |
| Lactic Acid (mmol/kg) | 15.52 ± 0.25 a | 16.30 ± 0.81 a | ND |
| Acetic Acid (mmol/kg) | 4.30 ± 0.01 | ND | ND |
| QF | 3.61 ± 0.08 | - | - |
| aw | 0.979 ± 0.002 a | 0.975 ± 0.002 a | 0.977 ± 0.000 a |
| Specific volume (cm3/g) | 2.21 ± 0.1 b | 2.20 ± 0.2 b | 2.46 ± 0.2 a |
| Textural profile analysis | |||
| Hardness (g) | 7050 ± 63 b | 7564 ± 66 a | 6034 ± 52 c |
| Resilience | 0.67 ± 0.04 b | 0.61 ± 0.04 c | 0.76 ± 0.05 a |
| Fracturability | 5579 ± 36 b | 5210 ± 49 c | 5745 ± 51 a |
| Image analysis | |||
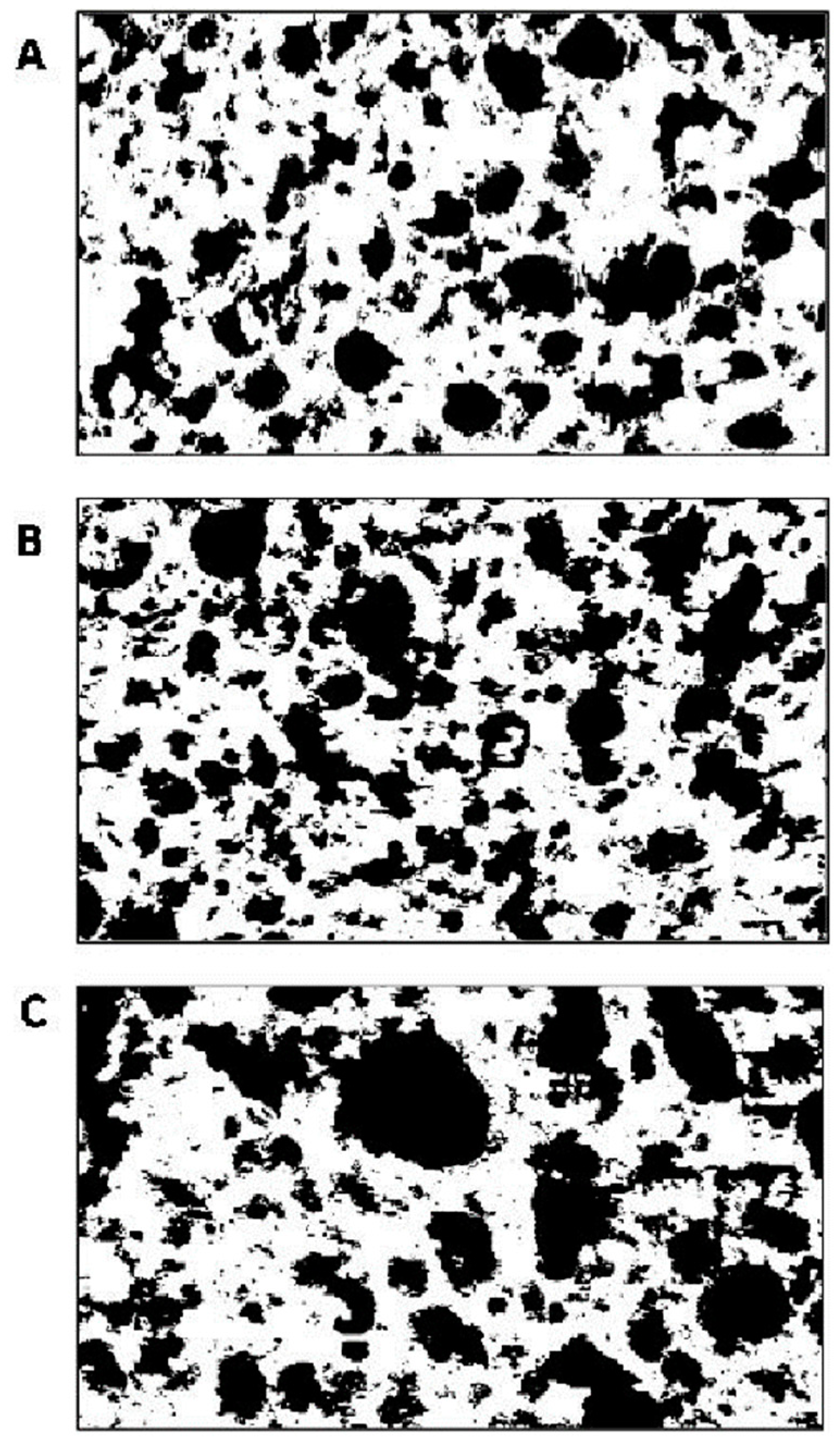
| Black pixel area * (%) | 34.8 ± 0.3 b | 34.9 ± 0.1 b | 38.9 ± 0.1 a |
| Color analysis | |||
| L | 51.9 ± 1.3 b | 47.4 ± 8.8 c | 60.1 ± 2.2 a |
| a | 9.9 ± 1.2 a | 8.3 ± 1.4 a | 5.5 ± 1.3 b |
| b | 32.6 ± 1.6 a | 31.3 ± 0.8 a | 31.1 ± 0.5 a |
| ΔE | 52.3 ± 1.2 b | 54.7 ± 1.9 a | 44.1 ± 1.3 c |
| Chemical Characteristics | Sourdough | Bread Dough | Puccia Bread | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SC2.27 | D0C2.27 | D1.5C2.27 | D0CTR | D1.5CTR | B0C2.27 | B1.5C2.27 | B0CTR | B1.5CTR | |
| pH | 3.57 ± 0.05 | 4.58 ± 0.15 a | 4.78 ± 0.28 ac | 5.48 ± 0.05 b | 5.44 ± 0.06 bc | 4.98 ± 0.08 a | 5.53 ± 0.07 a | 5.94 ± 0.34 a | 5.72 ± 0.02 a |
| TTA (mL) | 5.53 ± 0.24 | 6.55 ± 0.81 a | 5.55 ± 0.04 a | 2.90 ± 0.03 b | 3.03 ± 0.04 b | 4.60 ± 0.81 a | 3.05 ± 0.32 a | 1.85 ± 0.37 b | 1.80 ± 0.30 b |
| Lactic acid (mmol/Kg) | 15.82 ± 0.02 | 23.04 ± 0.25 a | 19.02 ± 0.39 b | 2.31 ± 0.18 c | ND | 14.29 ± 0.91 a | 8.42 ± 0.45 b | ND | ND |
| Acetic acid (mmol/Kg) | 8.23 ± 0.15 | 7.05 ± 0.95 a | 5.73 ± 0.12 a | ND | ND | 5.67 ± 0.63 a | 2.52 ± 0.05 b | ND | ND |
| QF | 1.92 ± 0.03 | 3.31 ± 0.02 a | 3.39 ± 0.46 a | - | - | 2.60 ± 0.04 b | 3.34 ± 0.87 a | - | - |
| Sample | LAB (log cfu/g) | Starter Strain (log cfu/g) | Other LAB Species § | Yeasts (log cfu/g) | Moulds (log cfu/g) | Yeast Species |
|---|---|---|---|---|---|---|
| SC2.27 | 8.90 ± 0.04 | 8.72 ± 0.02 | Lb. rossiae (1) * | 4.22 ± 0.06 | - | W. anomalus (1) C. pararugosa (1) C. parapsilosis (1) ** |
| Lb. plantarum (1) | ||||||
| D0C2.27 | 8.91 ± 0.04 a | 8.76 ± 0.08 a | Lb. rossiae (1) * | 3.82 ± 0.01 b | - | S. cerevisiae (1) *** C. pararugosa (1) R. mucillaginosa (1) C. parapsilosis (1) ** H. burtonii (1) |
| D1.5C2.27 | 8.91 ± 0.09 a | 8.85 ± 0.07 a | 3.95 ± 0.06 b | - | ||
| D0CTR | 6.68 ± 0.02 b | - | L. mesenteroides (1) Lb. plantarum (2) Lb. rossiae (1) * | 7.97 ± 0.01 a | S. cerevisiae (1) *** | |
| D1.5CTR | 6.82 ± 0.18 b | - | 8.03 ± 0.14 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Bellis, P.; Rizzello, C.G.; Sisto, A.; Valerio, F.; Lonigro, S.L.; Conte, A.; Lorusso, V.; Lavermicocca, P. Use of a Selected Leuconostoc Citreum Strain as a Starter for Making a “Yeast-Free” Bread. Foods 2019, 8, 70. https://doi.org/10.3390/foods8020070
De Bellis P, Rizzello CG, Sisto A, Valerio F, Lonigro SL, Conte A, Lorusso V, Lavermicocca P. Use of a Selected Leuconostoc Citreum Strain as a Starter for Making a “Yeast-Free” Bread. Foods. 2019; 8(2):70. https://doi.org/10.3390/foods8020070
Chicago/Turabian StyleDe Bellis, Palmira, Carlo Giuseppe Rizzello, Angelo Sisto, Francesca Valerio, Stella Lisa Lonigro, Amalia Conte, Valeria Lorusso, and Paola Lavermicocca. 2019. "Use of a Selected Leuconostoc Citreum Strain as a Starter for Making a “Yeast-Free” Bread" Foods 8, no. 2: 70. https://doi.org/10.3390/foods8020070