Feeding of 1-Kestose Induces Glutathione-S-Transferase Expression in Mouse Liver

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Analysis for Blood Components
2.3. Preparation of Total RNA
2.4. Microarray Analyses
2.5. Quantification of the Gene Expression Level Using qRT-PCR
2.6. Hepatic GST Activity Assay
2.7. Statistics Analysis
3. Results
3.1. Body Weight, Food Intake and Tissue Weights
3.2. Concentrations of Blood Components
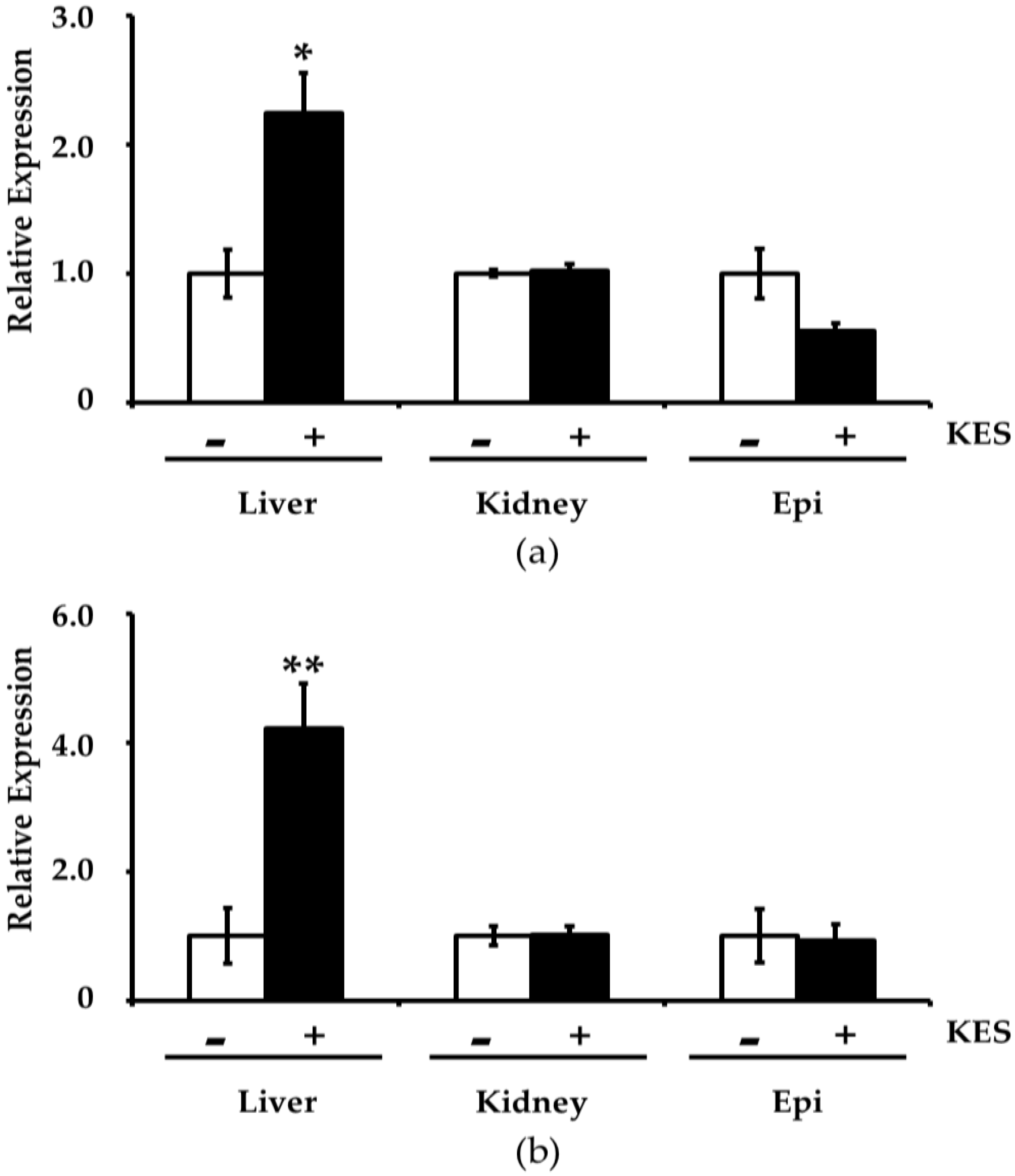
3.3. Effects of Dietary Intake of 1-Kestose on Gene Expression Related to Oxidative Stress in the Tissues
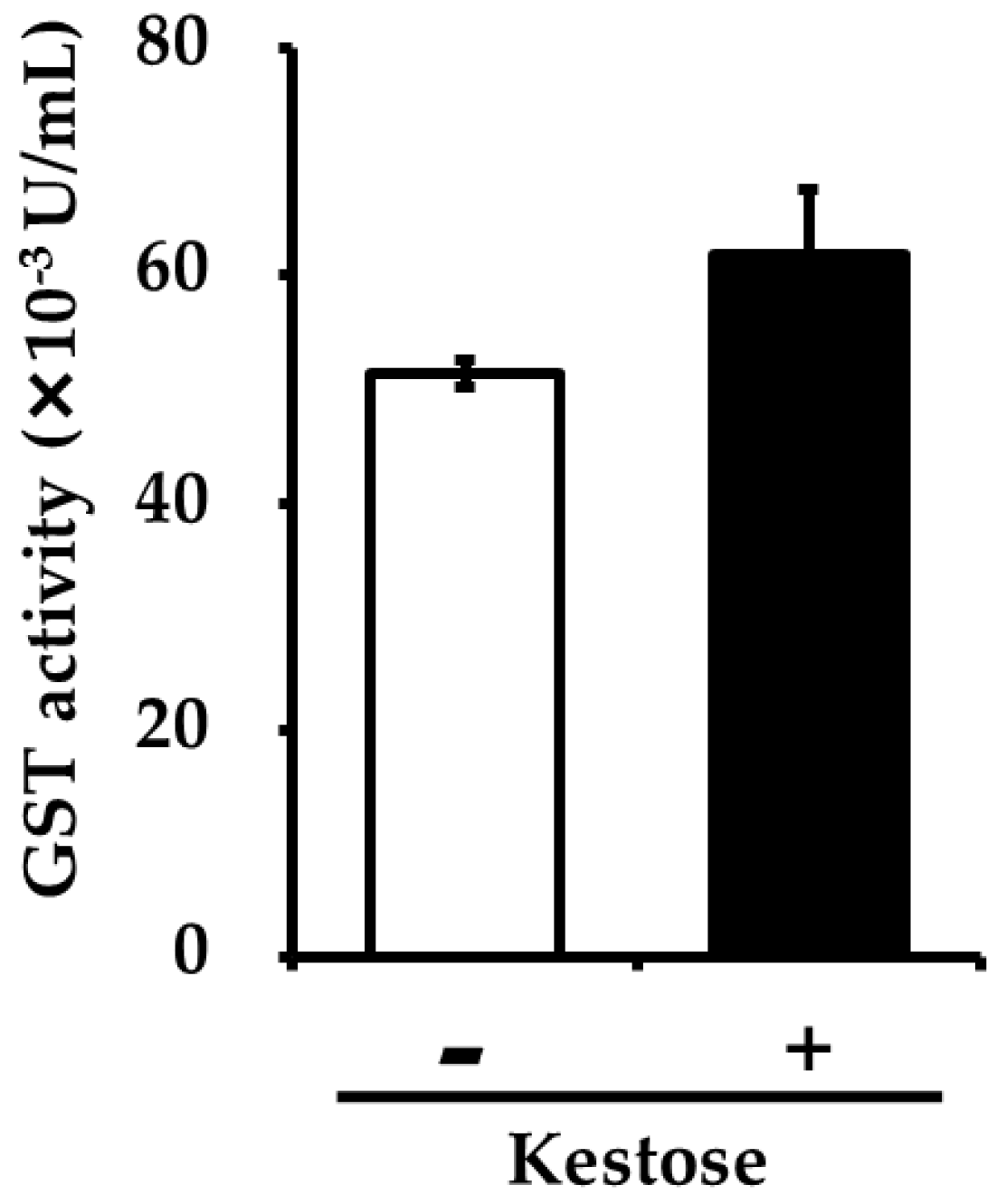
3.4. Hepatic GST Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [PubMed]
- Paineau, D.; Respondek, F.; Menet, V.; Sauvage, R.; Bornet, F.; Wagner, A. Effects of short-chain fructooligosaccharideson faecal bifidobacteria and specific immune response in formula-fed term infants: A randomized, double-blind, placebo-controlled trial. J. Nutr. Sci. Vitaminol. 2014, 60, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Kondo, N.; Yamaguchi, Y.; Hashiguchi, M.; Tanabe, K.; Ushiroda, C.; Kawahashi-Tokuhisa, M.; Yui, K.; Miyakoda, M.; Oku, T. Daily feeding of fructooligosaccharide or glucomannan delays onset of senescence in SAMP8 mice. Gastroenterol. Res. Pract. 2014, 2014, 303184. [Google Scholar] [CrossRef] [PubMed]
- Gomides, A.F.F.; de Paula, S.O.; GoncËalves, R.V.; de Oliveira, L.L.; Ferreira, C.L.; Comastri, D.S.; Peluzio Mdo, C. Prebiotics prevent the appearance of aberrant crypt foci (ACF) in the colon of BALB/c mice for increasing the gene expression of p16 protein. Nutr Hosp. 2014, 30, 883–890. [Google Scholar] [CrossRef] [PubMed]
- CapitaÂn-Cañadas, F.; Ortega-Gonzalez, M.; Guadix, E.; Zarzuelo, A.; Suárez, M.D.; de Medina, F.S.; Martínez-Augustin, O. Prebiotic oligosaccharides directly modulate proinflammatory cytokine production in monocytes via activation of TLR4. Mol. Nutr. Food Res. 2014, 58, 1098–1110. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Bauer, L.L.; Fahey, G.C.; Hogarth, A.J.C.L.; Wolf, B.W.; Diane, E.; Hunter, D.E. Selected fructooligosaccharide (1-kestose, nystose, and 1F-β-fructofuranosylnystose) composition of foods and feeds. J. Agric. Food Chem. 1997, 45, 3076–3082. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Vahedi, K.; Achour, L.; Attar, A.; Salfati, J.; Pochart, P.; Marteau, P.; Flourié, B.; Bornet, F.; Rambaud, J.C. Short-chain fructo-oligosaccharide administration dose-dependently increases fecal bifidobacteria in healthy humans. J. Nutr. 1999, 129, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Nakamura, S.; Konishi, K.; Nakagawa, J.; Tochio, T. Variations in prebiotic oligosaccharide fermentation by intestinal lactic acid bacteria. Int. J. Food Sci. Nutr. 2016, 67, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Tochio, T.; Kitaura, Y.; Nakamura, S.; Sugawa, C.; Takahashi, M.; Endo, A.; Shimomura, Y. An Alteration in the Cecal Microbiota Composition by Feeding of 1-Kestose Results in a Marked Increase in the Cecal Butyrate Content in Rats. PLoS ONE 2016, 11, e0166850. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Advances in mechanisms of anti-oxidation. Discov. Med. 2014, 17, 121–130. [Google Scholar] [PubMed]
- Halliwell, B.; Aeschbach, R.; Löliger, J.; Aruoma, O.I. The characterization of antioxidants. Food Chem. Toxicol. 1995, 33, 601–617. [Google Scholar] [CrossRef]
- Westhuyzen, J. The oxidation hypothesis of atherosclerosis: An update. Ann. Clin. Lab. Sci. 1997, 27, 1–10. [Google Scholar] [PubMed]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson’s disease. J. Parkinsons Dis. 2013, 3, 461–491. [Google Scholar] [PubMed]
- Kostner, K.; Hornykewycz, S.; Yang, P.; Neunteufl, T.; Glogar, D.; Weidinger, F.; Maurer, G.; Huber, K. Is oxidative stress causally linked to unstable angina pectoris? A study in 100 CAD patients and matched controls. Cardiovasc. Res. 1997, 36, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Ide, T.; Tsutsui, H.; Hayashidani, S.; Kang, D.; Suematsu, N.; Nakamura, K.; Utsumi, H.; Hamasaki, N.; Takeshita, A. Mitochondrial DNA damage and dysfunction associated with oxidative stress in failing hearts after myocardial infarction. Circ. Res. 2001, 88, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.J.; Zhang, X.; Chen, W.W. Role of oxidative stress in Alzheimer’s disease. Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Salehi-Abargouei, A.; Ghiasvand, R.; Hariri, M. Prebiotics, Prosynbiotics and Synbiotics: Can They Reduce Plasma Oxidative Stress Parameters? A Systematic Review. Probiotics Antimicrob. Proteins 2017, 9, 1–11. [Google Scholar] [CrossRef]
- Ebert, M.N.; Beyer-Sehlmeyer, G.; Liegibel, U.M.; Kautenburger, T.; Becker, T.W.; Pool-Zobel, B.L. Butyrate induces glutathione S-transferase in human colon cells and protects from genetic damage by 4-hydroxy-2-nonenal. Nutr. Cancer 2001, 41, 156–164. [Google Scholar] [CrossRef]
- De Vos, W.M.; de Vos, E.A. Role of the intestinal microbiome in health and disease: From correlation to causation. Nutr. Rev. 2012, 70, S45–S56. [Google Scholar] [CrossRef]
- De Goffau, M.C.; Fuentes, S.; van den Bogert, B.; Honkanen, H.; de Vos, W.M.; Welling, G.W.; Hyöty, H.; Harmsen, H.J. Aberrant gut microbiota composition at the onset of type 1 diabetes in young children. Diabetologia 2014, 57, 1569–1577. [Google Scholar] [CrossRef]
- Pineiro, M.; Asp, N.G.; Reid, G.; Macfarlane, S.; Morelli, L.; Brunser, O.; Tuohy, K. FAO Technical meeting on prebiotics. J. Clin. Gastroenterol. 2008, 42, S156–S159. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ingredient | Control Diet | 1-Kestose Diet |
|---|---|---|
| (g/100 g Diet) | ||
| Corn starch | 39.7 | 39.7 |
| Maltodextrin 10 | 13.2 | 13.2 |
| Sucrose | 10.0 | 5.0 |
| Casein | 20.0 | 20.0 |
| Soybean oil | 7.0 | 7.0 |
| Cellulose | 5.0 | 5.0 |
| Mineral mix | 3.5 | 3.5 |
| Vitamin mix | 1.0 | 1.0 |
| L-Cysteine | 0.3 | 0.3 |
| Choline bitartrate | 0.3 | 0.3 |
| 1-Kestose | 0 | 5.0 |
| Total | 100.0 | 100.0 |
| Target | GenBank Acc | Oligonucleotide Sequence |
|---|---|---|
| β-Actin | NM007393.4 | CATCCGTAAAGACCTCTATGCCAAC |
| ATGGAGCCACCGATCCACA | ||
| Gstp1 | NM_0103541.1 | CGGCAAATATGTCACCCTCATCTA |
| TCTGGGACAGCAGGGTCTCA | ||
| Gsta4 | NM_010357.3 | TGACACAGACCAGGGCCATC |
| ATCAGGTCCTGGGTGCCATC |
| KES (−) | KES (+) | |
|---|---|---|
| Means ± SE | ||
| Body weight (g) | 33.7 ± 1.4 | 32.0 ± 1.3 |
| Food intake (g/day) | 3.30 ± 0.09 | 3.53 ± 0.12 |
| Liver (g) | 1.45 ± 0.09 | 1.31 ± 0.08 |
| Kidney (g) | 0.32 ± 0.01 | 0.31 ± 0.01 |
| Epidermal adipose tissue (g) | 1.52 ± 0.14 | 1.23 ± 0.16 |
| Cecum (g) | 0.09 ± 0.00 | 0.16 ± 0.01 * |
| Cecum content (g) | 0.27 ± 0.02 | 0.43 ± 0.03 * |
| Serum triglycerides (mg/dL) | 155 ± 23 | 133 ± 13 |
| Serum total cholesterol (mg/dL) | 155 ± 1 | 138 ± 4 |
| Serum free fatty acids (mEq/L) | 0.57 ± 0.01 | 0.51 ± 0.02 |
| Gene | Expression Ratio KES (+)/KES (−) |
|---|---|
| Gstp1 (glutathione S-transferase pi 1) | 2.13 |
| Tgfb3 (transforming growth factor beta 3) | 1.61 |
| Gsta4 (glutathione S-transferase alpha 4) | 1.50 |
| Cyp4a10 (cytochrome P450, family 4, subfamily a, polypeptide 10) | 0.75 |
| Srebf1 (sterol regulatory element-binding transcription factor 1) | 0.75 |
| Col18a1 (collagen type XVIII alpha 1 chain) | 0.70 |
| Cyp4a12a (cytochrome P450, family 4, subfamily a, polypeptide 12a) | 0.70 |
| Ldlr (low-density lipoprotein receptor) | 0.69 |
| Keap1 (Kelch-like ECH-associated protein 1) | 0.69 |
| Gck (glucokinase) | 0.69 |
| Gstt3 (glutathione S-transferase theta 3) | 0.64 |
| Fasn (fatty acid synthase) | 0.47 |
| Mt3 (metallothionein 3) | 0.45 |
| Apoa4 (apolipoprotein A4) | 0.40 |
| Cyp7a1 (cytochrome P450, family 7, subfamily a, polypeptide 1) | 0.34 |
| Mt1 (metallothionein 1) | 0.28 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tochio, T.; Ueno, Y.; Kitaura, Y.; Shinohara, M.; Kadota, Y.; Minoda, K.; Shimomura, Y.; Osawa, T. Feeding of 1-Kestose Induces Glutathione-S-Transferase Expression in Mouse Liver. Foods 2019, 8, 69. https://doi.org/10.3390/foods8020069
Tochio T, Ueno Y, Kitaura Y, Shinohara M, Kadota Y, Minoda K, Shimomura Y, Osawa T. Feeding of 1-Kestose Induces Glutathione-S-Transferase Expression in Mouse Liver. Foods. 2019; 8(2):69. https://doi.org/10.3390/foods8020069
Chicago/Turabian StyleTochio, Takumi, Yuki Ueno, Yasuyuki Kitaura, Mikako Shinohara, Yoshihiro Kadota, Kanako Minoda, Yoshiharu Shimomura, and Toshihiko Osawa. 2019. "Feeding of 1-Kestose Induces Glutathione-S-Transferase Expression in Mouse Liver" Foods 8, no. 2: 69. https://doi.org/10.3390/foods8020069