
Effect of Mechanical Damage in Green-Making Process on Aroma of Rougui Tea
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Collection
2.2. Analysis of Aroma Components
2.3. Transcriptomic Analysis on RGT Samples
2.4. Proteome Analysis of Tea Samples
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Statistical Analysis
3. Results
3.1. Analysis of Content of Volatile Compounds in Tea Leaves
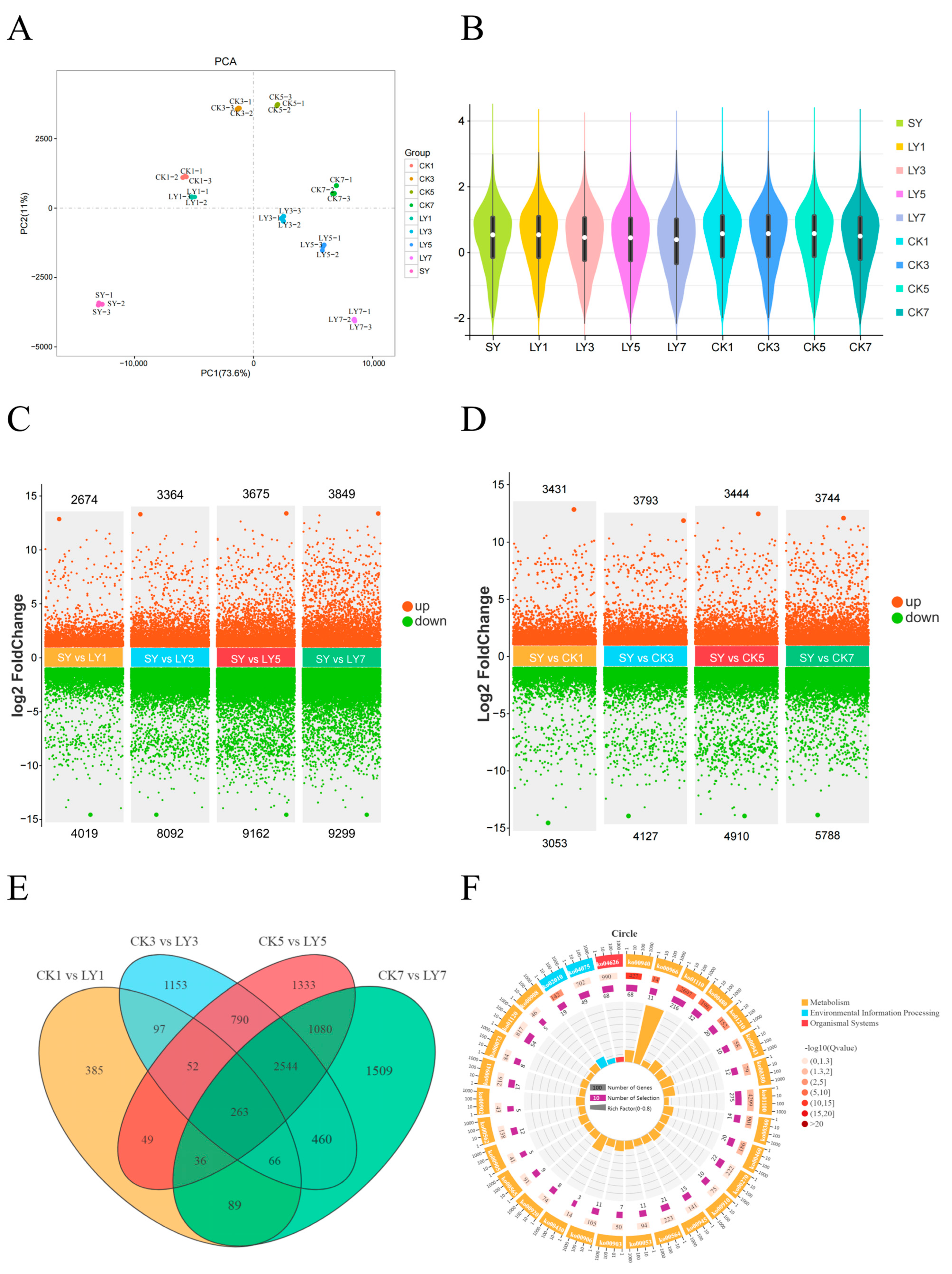
3.2. Analysis of Transcriptomic Profiles of Tea Samples
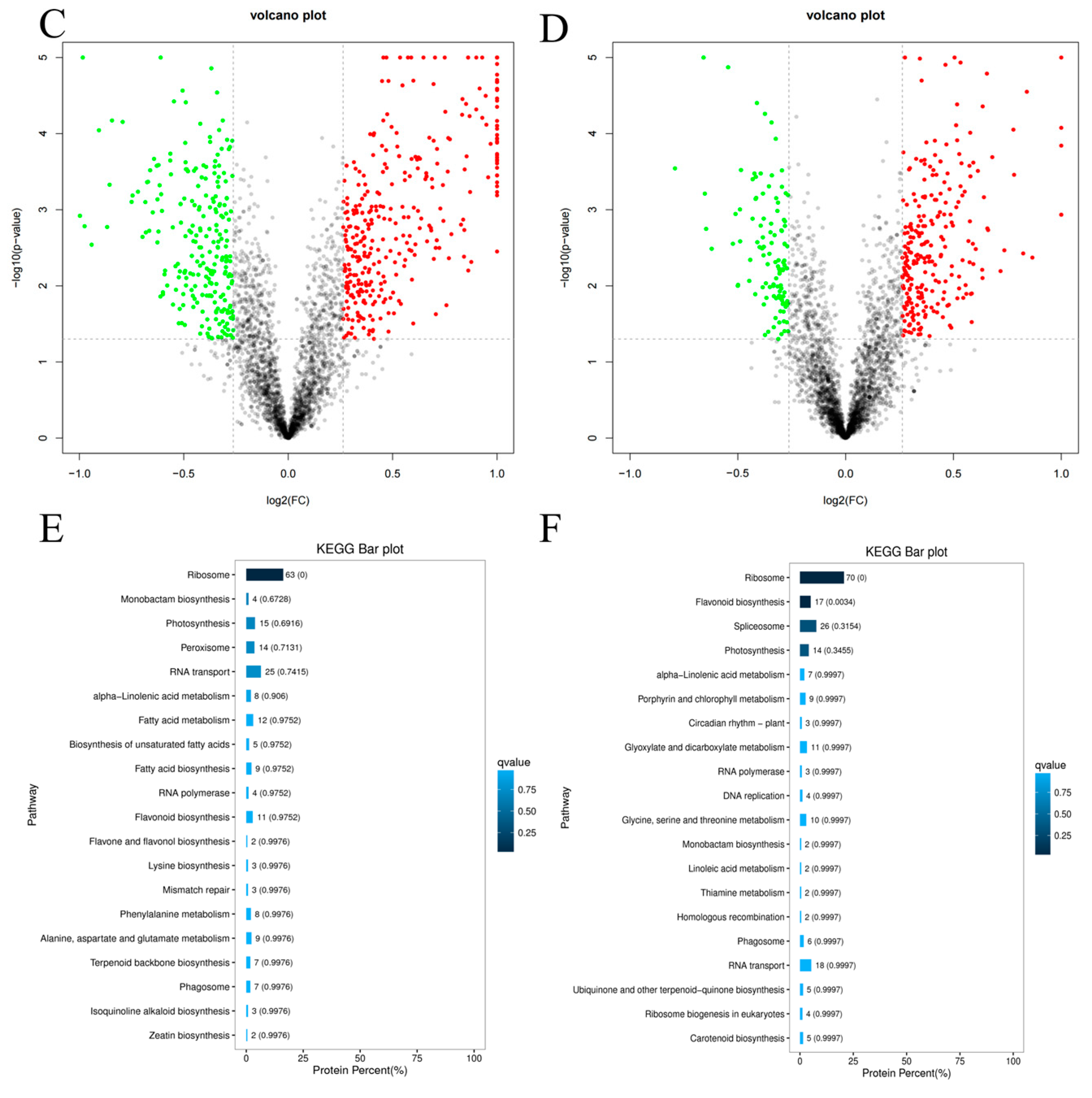
3.3. Proteome Profiling of Proteins Differentially Expressed in Response to Green-Making Process
3.4. Correlation Analysis of Proteome and Transcriptome
3.5. Analysis of DEGs and DEPs Related to Biosynthesis of Aroma Components
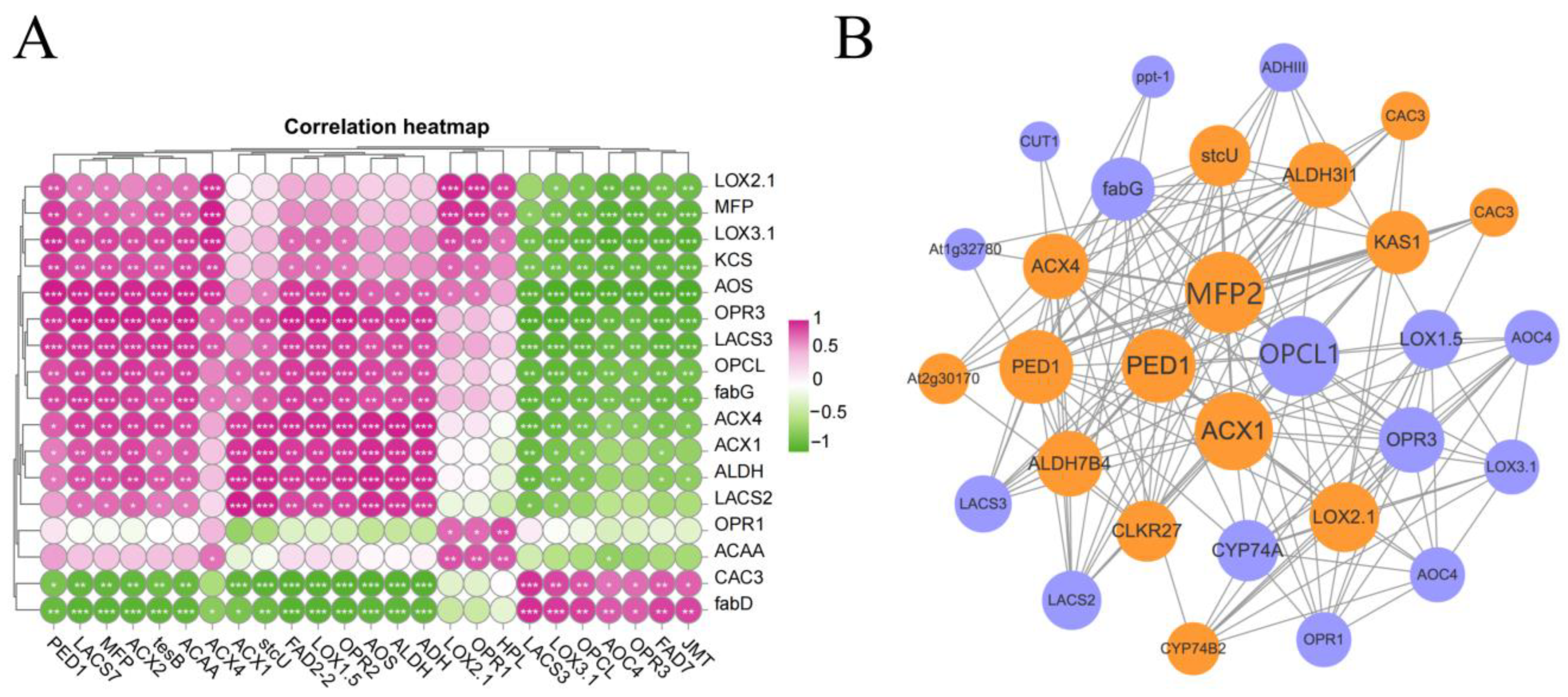
3.6. Correlation Analysis of DEGs and DEPs Related to Fatty Acid Metabolism and α-Linolenic Acid Metabolism
3.7. Protein–Protein Interaction Network Analysis
3.8. Quantitative Real-Time PCR (qRT-PCR) Validation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, X.; Ho, C.T.; Wan, X.; Zhu, H.; Liu, Q.; Wen, Z. Changes of volatile compounds and odor profiles in Wuyi rock tea during processing. Food Chem. 2021, 341, 128230. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Wang, H.; Cao, Q.; Song, H.; Xu, Y.; Lin, Y. Aroma-active compounds related to Maillard reaction during roasting in Wuyi rock tea. J. Food Compos. Anal. 2023, 115, 104954. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, F.; Sun, J.; Ni, L. Dynamic changes of volatile and phenolic components during the whole manufacturing process of Wuyi Rock tea (Rougui). Food Chem. 2022, 367, 130624. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Chen, F.; Liu, G.; Zhang, Q.; Ye, J.; Lei, W.; Jia, X.; He, H. Comparative analysis on the quality of Wuyi Rougui (Camellia sinensis) tea with different grades. Food Sci. Technol. 2022, 42, 115321. [Google Scholar] [CrossRef]
- Jia, X.; Wang, Y.; Li, Q.; Zhang, Q.; Zhang, Y.; Lin, S.; Cheng, P.; Chen, M.; Du, M.; Ye, J.; et al. Contribution of traditional deep fermentation to volatile metabolites and odor characteristics of Wuyi rock tea. Front. Bioeng. Biotechnol. 2023, 11, 1193095. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Chen, Y.; Lee, C.; Cheng, C.; Roan, S.; Chen, I. Monitoring volatile compound profiles and chemical compositions during the process of manufacturing semi-fermented oolong tea. J. Hortic. Sci. Biotech. 2013, 88, 159–164. [Google Scholar] [CrossRef]
- Lin, S.; Lo, L.; Chen, I.; Chen, P. Effect of shaking process on correlations between catechins and volatiles in oolong tea. J. Food Drug Anal. 2016, 24, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Hu, Q.; Wu, Z.; Bi, W.; Chen, B.; Hao, Z.; Wu, L.; Ye, N.; Sun, Y. Volatile metabolomics and coexpression network analyses provide insight into the formation of the characteristic cultivar aroma of oolong tea (Camellia sinensis). Food Sci. Technol. 2022, 164, 113666. [Google Scholar] [CrossRef]
- Tseng, T.S.; Hsiao, M.H.; Chen, P.A.; Lin, S.Y.; Chiu, S.W.; Yao, D.J. Utilization of a gas-sensing system to discriminate smell and to monitor fermentation during the manufacture of oolong tea leaves. Micromachines 2021, 12, 93. [Google Scholar] [CrossRef]
- Jia, X.; Zhang, Q.; Chen, M.; Wang, Y.; Lin, S.; Pan, Y.; Cheng, P.; Li, M.; Zhang, Y.; Ye, J.; et al. Analysis of the effect of different withering methods on tea quality based on transcriptomics and metabolomics. Front. Plant Sci. 2023, 14, 123568. [Google Scholar] [CrossRef]
- Zhang, Q.; Jia, X.; Chen, M.; Wang, Y.; Lin, S.; Pan, Y.; Cheng, P.; Li, M.; Zhang, Y.; Luo, Z.; et al. Effect of different degrees of withering on gene expression and metabolite content of Wuyi rock tea leaves. LWT Food Sci. Technol. 2023, 189, 115462. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Jin, S.; Xu, Y.; Granato, D.; Fu, Y.; Sun, W.; Yin, J.; Xu, Y. Exogenous stimulation-induced biosynthesis of volatile compounds: Aroma formation of oolong tea at postharvest stage. Crit. Rev. Food Sci. Nutr. 2024, 64, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.; Fu, X.; Zhou, Y.; Katsuno, T.; Mei, X.; Deng, R.; Xu, X.; Zhang, L.; Dong, F.; Watanabe, N. Does enzymatic hydrolysis of glycosidically bound volatile compounds really con-tribute to the formation of volatile compounds during the oolong tea manufacturing process? J. Agric. Food Chem. 2015, 63, 6905–6914. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Li, D.; Ma, Y.; Zhang, W.; Lin, C.; Zheng, X.; Liang, Y.; Lu, J. Formation mechanism of the oolong tea characteristic aroma during bruising and withering treatment. Food Chem. 2018, 269, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Watanabe, N.; Yang, Z. Understanding the biosyntheses and stress response mechanisms of aroma compounds in tea (Camellia sinensis) to safely and effectively improve tea aroma. Crit. Rev. Food Sci. Nutr. 2019, 59, 2321–2334. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, Y.; Li, L.; Li, F.; He, Y.; Wu, J.; Wei, C. Transcriptomic and phytochemical analyses reveal root-mediated resource-based defense response to leaf herbivory by ectropis oblique in tea plant (Camellia sinensis). J. Agric. Food Chem. 2019, 67, 5465–5476. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zeng, L.; Hou, X.; Liao, Y.; Yang, Z. Low temperature synergistically promotes wounding-induced indole accumulation by INDUCER OF CBF EXPRESSION-mediated alterations of jasmonic acid signaling in Camellia sinensis. J. Exp. Bot. 2020, 71, 2172–2185. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Liu, X.; Zhou, Y.; Wang, X.; Zeng, L.; Fu, X.; Li, J.; Tang, J.; Dong, F.; Yang, Z. Formation and emission of linalool in tea (Camellia sinensis) leaves infested by tea green leafhopper (Empoasca (Matsumurasca) onukii Matsuda). Food Chem. 2017, 237, 356–363. [Google Scholar] [CrossRef]
- Li, J.; Zeng, L.; Liao, Y.; Gu, D.; Tang, J.; Yang, Z. Influence of chloroplast defects on formation of jasmonic acid and characteristic aroma compounds in tea (Camellia sinensis) leaves exposed to postharvest stresses. Int. J. Mol. Sci. 2019, 20, 1044. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, B.; Wu, Q.; Bi, W.; Ni, Z.; Lai, Z.; Sun, Y. Formation and Regulation of Aroma-Related Volatiles During the Manufacturing Process of Wuyi Rougui Tea via LOX-HPL Pathway. J. Food Sci. Biotech. 2019, 40, 100–110. [Google Scholar]
- Zheng, Y.; Hu, Q.; Yang, Y.; Wu, Z.; Wu, L.; Wang, P.; Deng, H.; Ye, N.; Sun, Y. Architecture and Dynamics of the Wounding-Induced Gene Regulatory Network During the Oolong Tea Manufacturing Process (Camellia sinensis). Front. Plant Sci. 2022, 12, 788469. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Huang, X.; Liu, S.; Liu, J.; Guo, Y.; Sun, Y.; Lin, J.; Guo, Y.; Wei, S. Understanding the formation mechanism of oolong tea characteristic non-volatile chemical constitutes during manufacturing processes by using integrated widely-targeted metabolome and DIA proteome analysis. Food Chem. 2020, 310, 125941. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, C.; Lin, J.; Sun, Y.; Wei, S.; Wu, L. The Impact of Different Withering Approaches on the Metabolism of Flavor Compounds in Oolong Tea Leaves. Foods 2022, 11, 3601. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, J.; Lv, H.; Peng, Q.; Schreiner, M.; Baldermann, S.; Lin, Z. Integrated proteomic and metabolomic analyses reveal the importance of aroma precursor accumulation and storage in methyl jasmonate-primed tea leaves. Hortic. Res. 2021, 8, 95. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Z.; Chen, W.; Lan, X.; Zhan, S.; Sun, Y.; Ni, L. Comparative study of the volatile fingerprints of roasted and unroasted oolong tea by sensory profiling and HS-SPME-GC-MS. Curr. Res. Food Sci. 2023, 6, 100442. [Google Scholar] [CrossRef] [PubMed]
- Zhan, S.; Su, W.; Wang, D.; Chen, W.; Lan, X.; Sun, Y.; Ni, L. Extraction of volatile components of Rougui tea using the headspace-solid-phase microextraction and simultaneous distillation extraction. J. Food Sci. Technol. 2022, 22, 181–190. [Google Scholar]
- Wu, Q.; Zhou, Z.; Ni, Z.; Lai, Z.; Sun, Y. Screening and expression analysis of key regulatory genes for α-farnene formation during oolong tea processing. J. Food Sci. Technol. 2020, 41, 135–142. [Google Scholar]
- Wang, D.; Yoshimura, T.; Kubota, K.; Kobayashi, A. Analysis of glycosidically bound aroma precursors in tea leaves: 1. Qualitative and quantitative analyses of glycosides with aglycons as aroma compounds. J. Agric. Food Chem. 2000, 48, 5411–5418. [Google Scholar] [CrossRef]
- Yuan, Q.P.; Yue, H.Y. Studies on PAL enzyme activity and acid-producing factors in L-phenylalanine production. J. B. Univ. Chem. Technol. 2003, 30, 8. [Google Scholar]
- Ma, C.; Li, J.; Chen, W.; Wang, W.; Qi, D.; Pang, S.; Miao, A. Study of the aroma formation and transformation during the manufacturing process of oolong tea by solid-phase micro-extraction and gas chromatography-mass spectrometry combined with chemometrics. Food Res. Int. 2018, 108, 413–422. [Google Scholar] [CrossRef]
- Mosblech, A.; Feussner, I.; Heilmann, I. Oxylipins: Structurally diverse metabolites from fatty acid oxidation. Plant Physiol. Biochem. 2009, 47, 511–517. [Google Scholar] [CrossRef]
- Lu, L.; Zhan, D.; Zhou, C.; Zhu, C.; Xie, S.; Xu, K.; Tian, C.; Lai, Z.; Guo, Y. Effects of key genes of jasmonic acid synthesis and transduction pathway in tea plant on terpenoids during oolong tea processing. Tea Sci. 2023, 43, 91–108. [Google Scholar]
- Li, C.X.; Lin, J.Q.; Hu, Q.C.; Sun, Y.; Wu, L.Y. An integrated metabolomic and transcriptomic analysis reveals the dynamic changes of key metabolites and flavor formation over Tieguanyin oolong tea production. Food Chem. 2023, 20, 100952. [Google Scholar] [CrossRef]
- Zeng, L.; Fu, Y.Q.; Liu, Y.Y.; Huang, J.S.; Chen, J.X.; Yin, J.F.; Jin, S.; Sun, W.J.; Xu, Y.Q. Comparative analysis of different grades of Tieguanyin oolong tea based on metabolomics and sensory evaluation. LWT 2023, 174, 114423. [Google Scholar] [CrossRef]
- Chen, M.H.; Zhang, Y.; Wang, Y.H.; Cheng, P.Y.; Zhang, Q.; Li, M.Z.; Jia, X.L.; Pan, Y.B.; Lin, S.X.; Luo, Z.W.; et al. Transcriptomic analysis of the effect of shaking and tumbling degree on quality formation of Wuyi rock tea. J. Food Sci. 2023, 89, 81–95. [Google Scholar] [CrossRef]
- Ye, C.; Xie, B.; Li, T.; Xu, X.; Xu, X.; Wang, H.; Wu, Z.; Heng, Z. Progress on aroma components and synthetic regulation mechanism of horticultural plants. J. Guangdong Agric. Sci. 2023, 50, 98–112. [Google Scholar]
- Xing, J.Y. Study on Key Genes Regulating Ovary Abortion in Hazelnut by Combining Proteome and Transcriptome Analysis. Master’s Thesis, Jilin Normal University, Jilin, China, 2019. [Google Scholar]
- Zhou, Z.W.; You, F.N.; Liu, B.B.; Deng, T.T.; Lai, Z.X.; Sun, Y. Effect of mechanical force during turning-over on the formation of aliphatic aroma in oolong tea. J. Food Sci. 2019, 40, 52–59. [Google Scholar]
- Cheng, Q.; Zhang, B.; Qiang, Z. Expression profiles of two novel lipoxygenase genes in Populus deltoides. Plant Sci. 2006, 170, 1027–1035. [Google Scholar] [CrossRef]
- Noordermeer, M.A.; Veldink, G.A.; Vliegenthart, J.F. Fatty acid hydroperoxide lyase: A plant cytochrome P450 enzyme involved in wound healing and pest resistance. ChemBioChem 2001, 2, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Vogt, J.; Schiller, D.; Ulrich, D. Identification of lipoxygenase (LOX) genes putatively involved in fruit flavour formation in apple (Malus × domestica). Tree Genet. Genomes 2013, 9, 1493–1511. [Google Scholar] [CrossRef]
- Cho, J.Y.; Mizutani, M.; Shimizu, B.I.; Kinoshita, T.; Ogura, M.; Tokoro, K.; Lin, M.L.; Sakata, K. Chemical profiling and gene expression profiling during the manufacturing process of Taiwan oolong tea “Oriental Beauty”. Biosci. Biotechnol. Biochem. 2007, 71, 1476–1486. [Google Scholar] [CrossRef]
- Zeng, L.T.; Zhou, Y.; Fu, X.M.; Liao, Y.Y.; Yuan, Y.F.; Jia, Y.X.; Dong, F.; Yang, Z.Y. Biosynthesis of jasmine lactone in tea (Camellia sinensis) leaves and its formation in response to multiple stresses. J. Agric. Food Chem. 2018, 66, 3899–3909. [Google Scholar] [CrossRef]
- Savchenko, T.; Pearse, I.S.; Ignatia, L.; Karban, R.; Dehesh, K. Insect herbivores selectively suppress the HPL branch of the oxylipin pathway in host plants. Plant J. 2013, 73, 653–662. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, X.Q.; Yu, K.J.; Zhu, B.Q.; Lan, Y.B.; Duan, C.Q.; Pan, Q.H. Varietal dependence of GLVs accumulation and LOX-HPL pathway gene expression in Four Vitis vinifera wine grapes. Int. J. Mol. Sci. 2016, 17, 1924. [Google Scholar] [CrossRef]
- Hui, X.; Liu, H.; Tian, F.L.; Li, F.F.; Li, H.; Gao, W.Y. Inhibition of green tea and the catechins against 1-deoxy-d-xylulose 5-phosphate reductoisomerase, the key enzyme of the MEP terpenoid biosynthetic pathway. Fitoterapia 2016, 113, 80–84. [Google Scholar] [CrossRef]
- Chen, L.B.; Liu, B.Y.; Wang, Y.G.; Xia, L.F.; Li, Y.Y.; Tian, Y.P. Cloning and sequence analysis of HMGS gene in tea tree. J. Northwest Agric. Sci. 2013, 22, 72–76. [Google Scholar]
- Zhang, Q.J.; Zhang, A.H.; Sun, J.B.; Zhang, L.X. Research progress on allelopathy of terpenoids in plants. Chin. J. Ecol. 2012, 21, 187–193. [Google Scholar] [CrossRef]
- Wang, X.W.; Zeng, L.T.; Liao, Y.Y.; Li, J.L.; Tang, J.C.; Yang, Z.Y. Formation of α-Farnesene in Tea (Camellia sinensis) Leaves Induced by Herbivore-Derived Wounding and Its Effect on Neighboring Tea Plants. Int. J. Mol. Sci. 2019, 20, 4151. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, L.; Liu, X.; Gui, J.; Mei, X.; Fu, X.; Dong, F.; Tang, J.; Zhang, L.; Yang, Z.Y. Formation of (E)-nerolidol in tea (Camellia sinensis) leaves exposed to multiple stresses during tea manufacturing. Food Chem. 2017, 231, 78–86. [Google Scholar] [CrossRef]
- Tieman, D.; Taylor, M.; Schauer, N.; Fernie, A.R.; Hanson, A.D.; Klee, H.J. Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde. Proc. Natl. Acad. Sci. USA 2006, 103, 87–92. [Google Scholar] [CrossRef]
- Zeng, L.T.; Zhou, Y.; Gui, J.D.; Fu, X.M.; Mei, X.; Zhen, Y.P.; Yang, Z.Y. Formation of volatile tea constituent indole during the oolong tea manufacturing process. J. Agric. Food Chem. 2016, 64, 5011–5019. [Google Scholar] [CrossRef]
- Sohn, S.I.; Pandian, S.; Rakkammal, K.; Largia, M.J.V.; Thamilarasan, S.K.; Balaji, S.; Rames, M. Jasmonates in plant growth and development and elicitation of secondary metabolites: An updated overview. Front. Plant Sci. 2022, 13, 942789. [Google Scholar] [CrossRef]
- Guo, L.; Chen, M.J.; Guo, Y.L.; Lin, Z. Variations in Fatty Acids Affected Their Derivative Volatiles during Tieguanyin Tea Processing. Foods. 2022, 11, 1563. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Y.; Wang, Y.; Zou, J.; Lin, S.; Chen, M.; Miao, P.; Jia, X.; Cheng, P.; Pang, X.; et al. Transcriptomic Analysis of the Effect of Pruning on Growth, Quality, and Yield of Wuyi Rock Tea. Plants 2023, 12, 3625. [Google Scholar] [CrossRef]
- Charbonnier, S.; Gallego, O.; Gavin, A.C. The social network of a cell: Recent advances in interactome mapping. Biotechnol. Annu. Rev. 2008, 14, 1–28. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P. STRING v10: Protein-proteininteraction networks, integrated over the tree of life. Nucleic. Acids Res. 2015, 43, 447–452. [Google Scholar] [CrossRef]
- Chen, S.; Lu, X.; Ge, L.; Sun, X.; Xin, Z. Wound- and pathogen-activated de novo JA synthesis using different ACX isozymes in tea plant (Camellia sinensis). J. Plant Physiol. 2019, 243, 153047. [Google Scholar] [CrossRef]
- Xin, Z.; Chen, S.; Ge, L.; Li, X.; Sun, X. The involvement of a herbivore-induced acyl-CoA oxidase gene, CsACX1, in the synthesis of jasmonic acid and its expression in flower opening in tea plant (Camellia sinensis). Plant Physiol. Biochem. 2019, 135, 132–140. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, F.; Wu, H.; Li, Z.; Huang, Y.; Lin, X.; Gao, C.; Wang, Z.; Yu, W.; Sun, W. Effect of Mechanical Damage in Green-Making Process on Aroma of Rougui Tea. Foods 2024, 13, 1315. https://doi.org/10.3390/foods13091315
Lin F, Wu H, Li Z, Huang Y, Lin X, Gao C, Wang Z, Yu W, Sun W. Effect of Mechanical Damage in Green-Making Process on Aroma of Rougui Tea. Foods. 2024; 13(9):1315. https://doi.org/10.3390/foods13091315
Chicago/Turabian StyleLin, Fuming, Huini Wu, Zhaolong Li, Yan Huang, Xiying Lin, Chenxi Gao, Zhihui Wang, Wenquan Yu, and Weijiang Sun. 2024. "Effect of Mechanical Damage in Green-Making Process on Aroma of Rougui Tea" Foods 13, no. 9: 1315. https://doi.org/10.3390/foods13091315