
A New Duplex Recombinase Polymerase Amplification (D-RPA) Method for the Simultaneous and Rapid Detection of Shigella and Bacillus cereus in Food
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. DNA Extraction
2.3. Primer Design and Screening
2.4. D-RPA Reaction
2.5. Condition Optimization
2.6. Specificity and Sensitivity Analysis
2.7. Application of D-RPA in Food Samples
3. Results
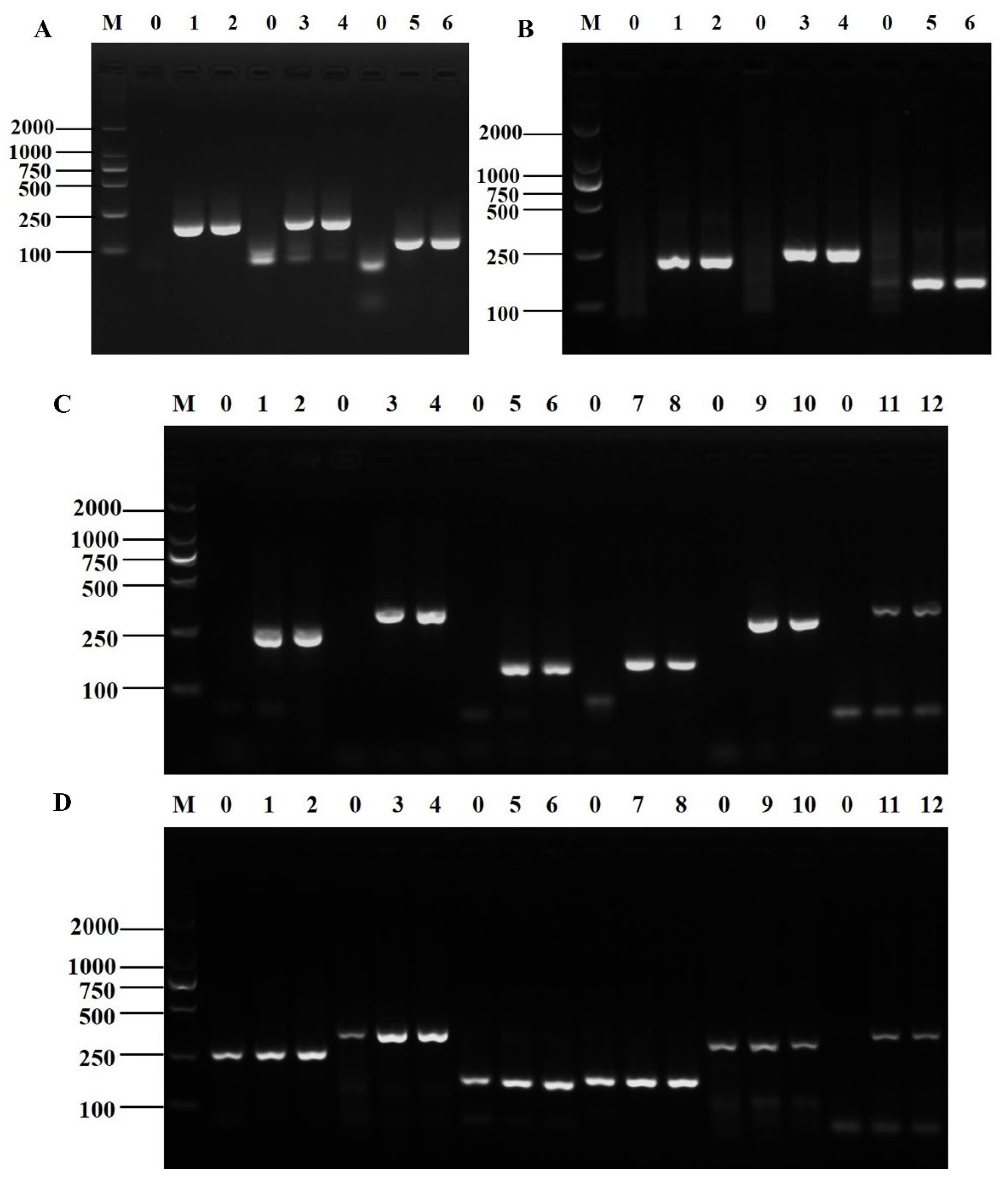
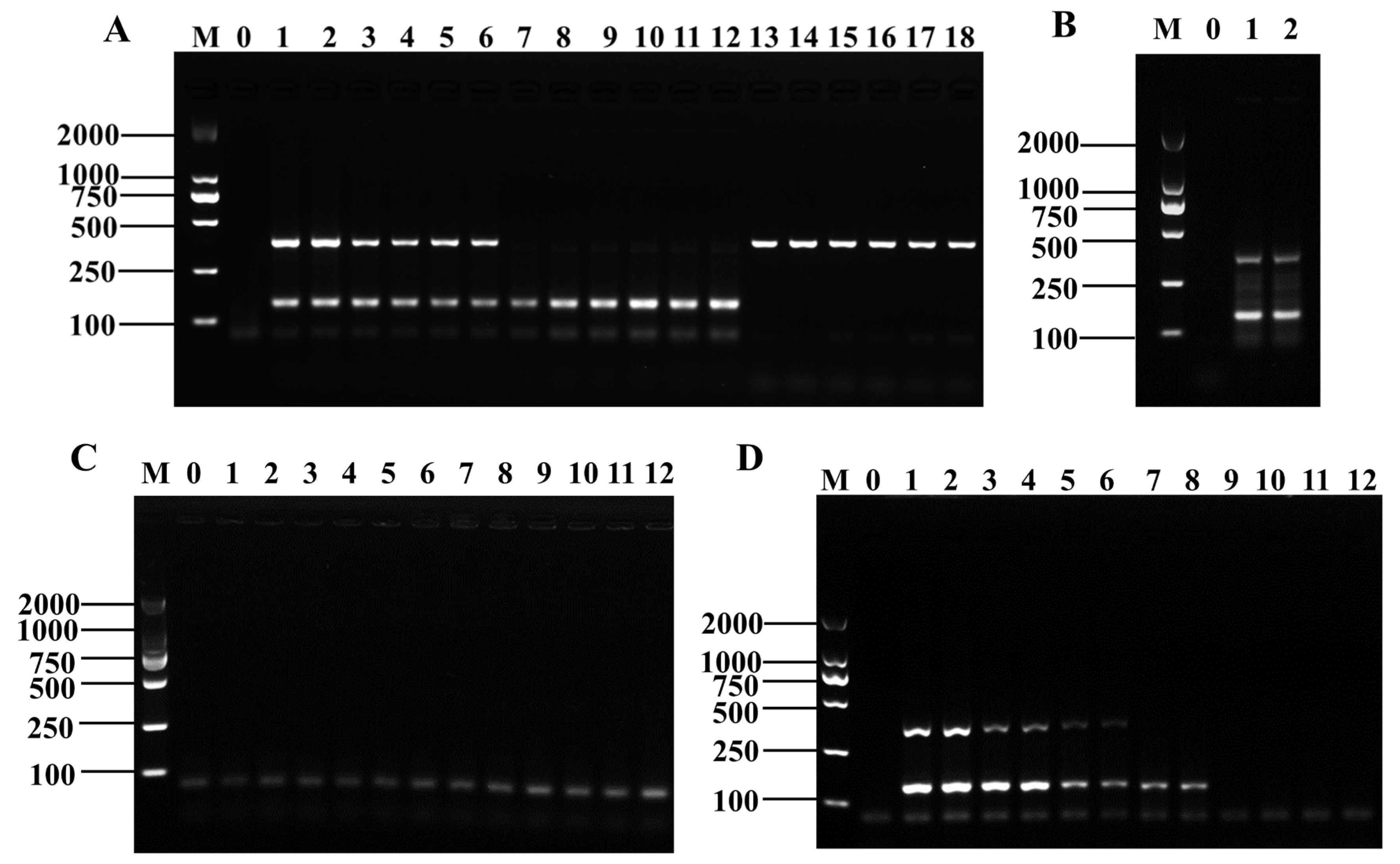
3.1. Primer Screening for Shigella and B. cereus
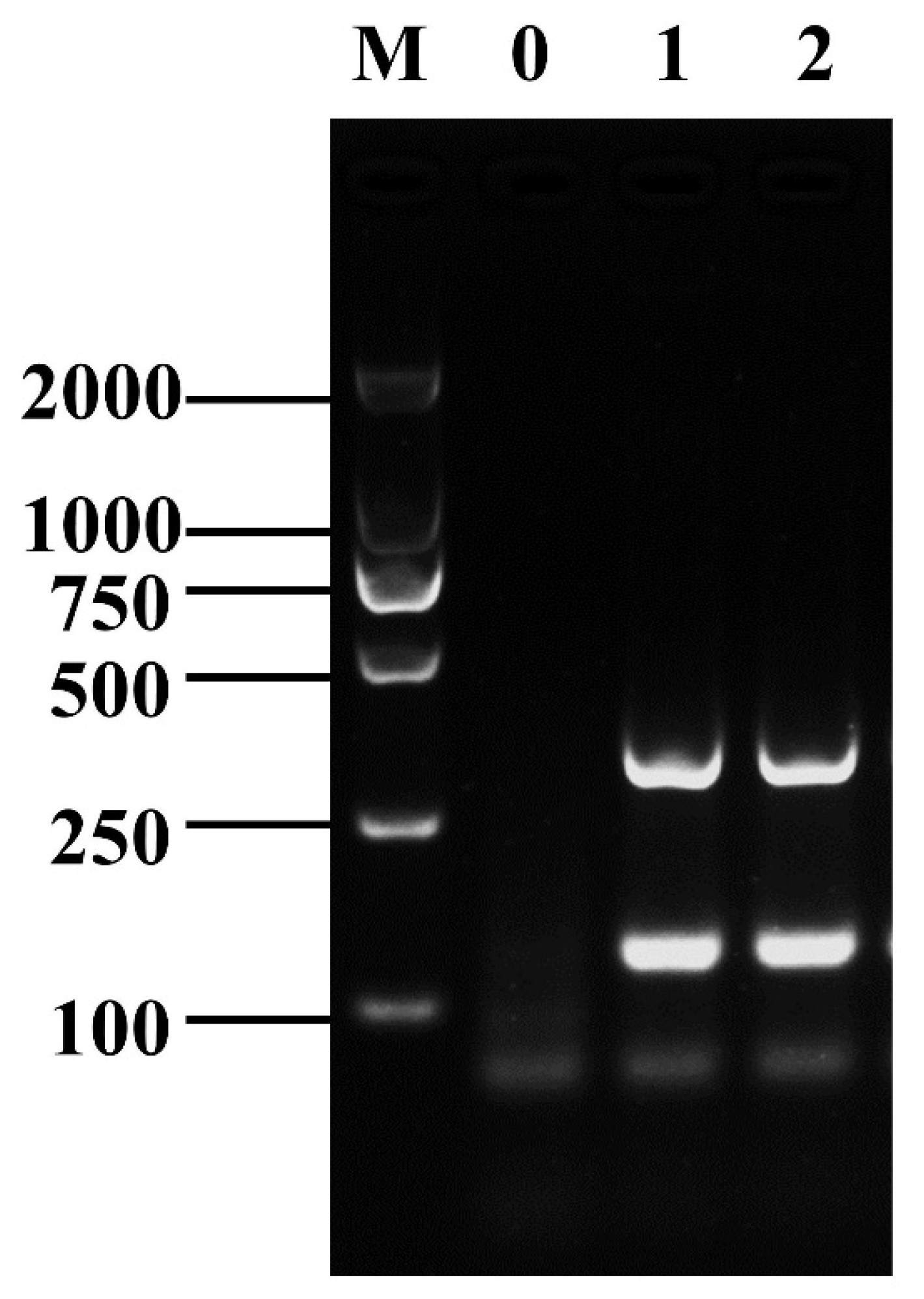
3.2. Establishment of D-RPA Reaction System for Shigella and B. cereus
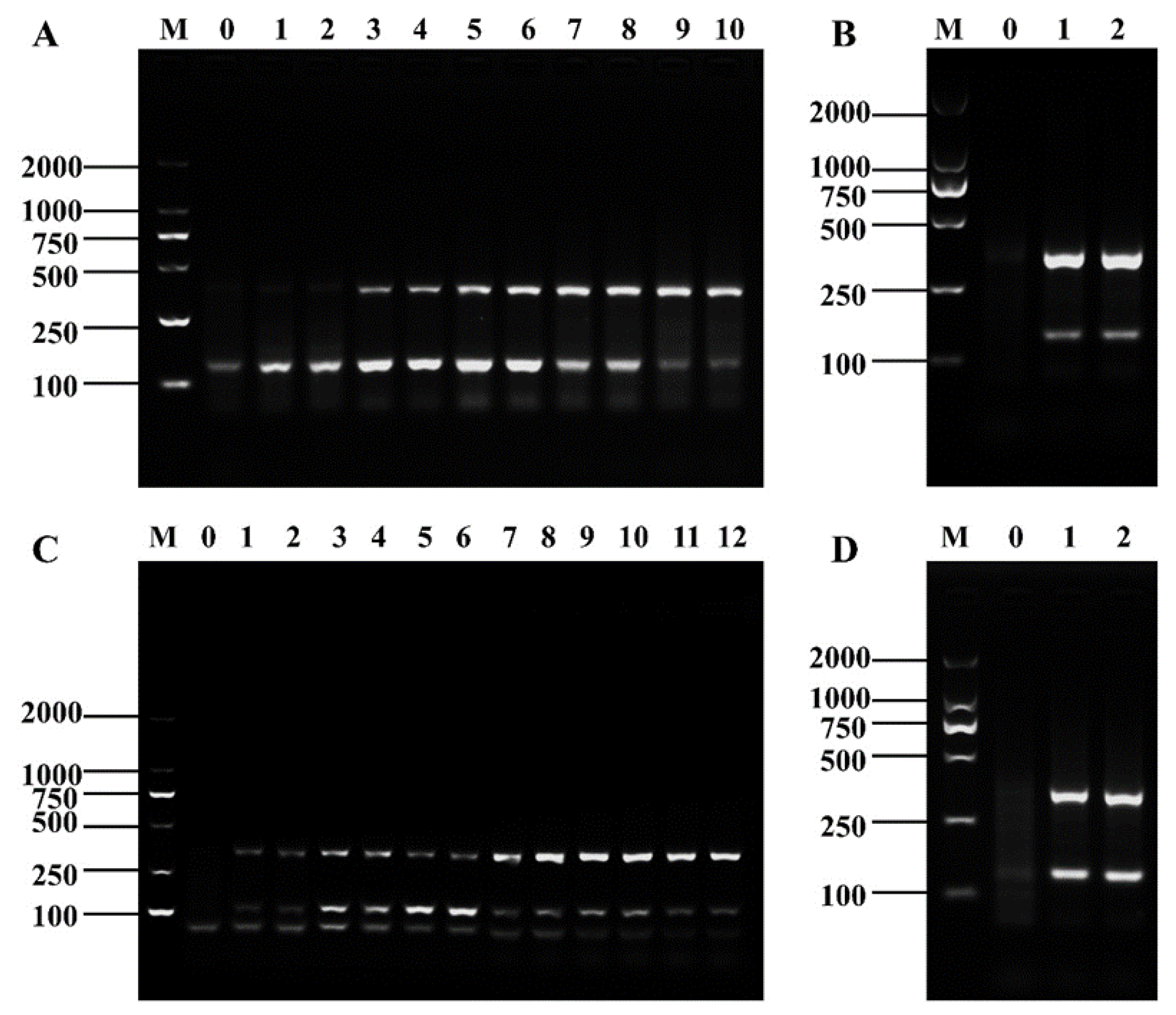
3.3. Optimization of D-RPA Reaction Conditions
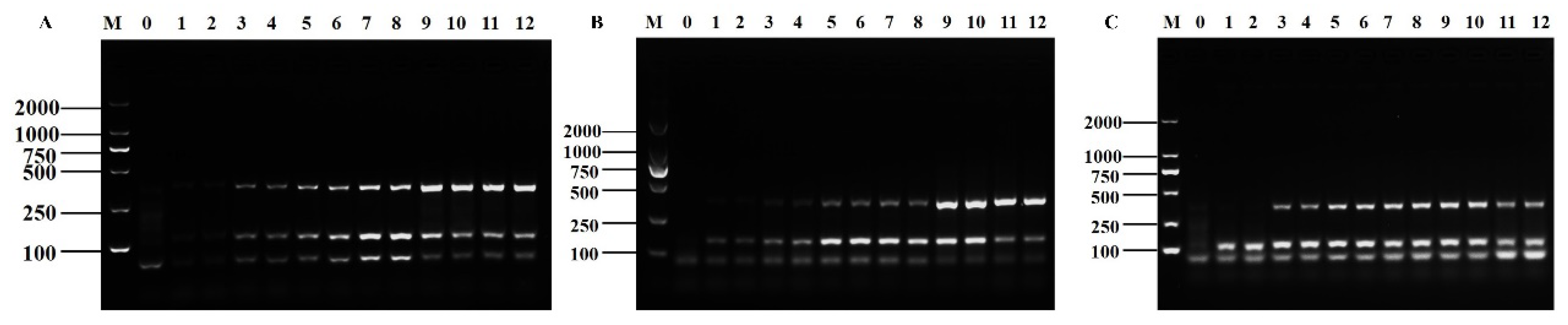
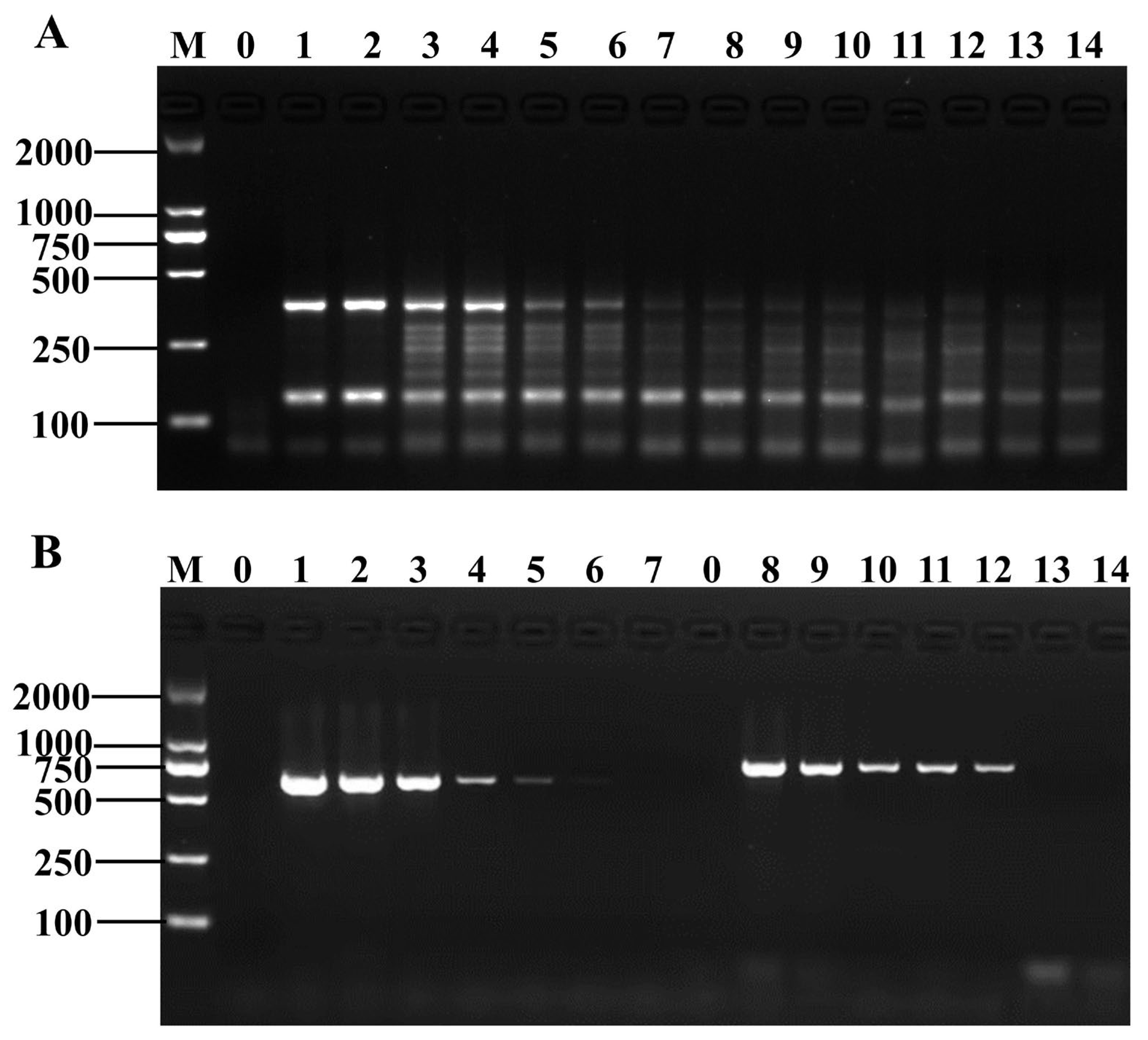
3.4. Specificity and Sensitivity of the Method
3.5. Application of D-RPA in Actual Sample Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ang, B.; Xu, X.; Liu, L.; Xu, L.; Kuang, H.; Xu, C. A colloidal gold immunochromatographic strip assay for the rapid detection of Shigella in milk and meat products. New J. Chem. 2021, 46, 103–109. [Google Scholar] [CrossRef]
- Lin, X.; Lai, W.; Zhang, L. Recent Advances in Detection of Shigella Species in Food. Food Sci. 2009, 30, 271–275. [Google Scholar]
- Kotloff, K.L.; Winickoff, J.P.; Ivanoff, B.; Clemens, J.D.; Swerdlow, D.L.; Sansonetti, P.J.; Adak, G.K.; Levine, M.M. Global burden of Shigella infections: Implications for vaccine development and implementation of control strategies. Bull. WHO 1999, 77, 651–666. [Google Scholar]
- Arnesen, L.P.S.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Grutsch, A.A.; Nimmer, P.S.; Pittsley, R.H.; Kornilow, K.G.; McKillip, J.L. Molecular Pathogenesis of Bacillus spp., with Emphasis on the Dairy Industry. Fine Focus 2018, 4, 203–222. [Google Scholar] [CrossRef]
- Cao, F.; Wang, P.; Jiang, L.; Chen, Y. Advances in Methods for Bacillus cereus Typing. Food Sci. 2017, 38, 286–290. [Google Scholar]
- Zhang, Z.; Feng, L.; Xu, H.; Liu, C.; Shah, N.P.; Wei, H. Detection of viable enterotoxin-producing Bacillus cereus and analysis of toxigenicity from ready-to-eat foods and infant formula milk powder by multiplex PCR. J. Dairy Sci. 2016, 99, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Cetin-Karaca, H.; Newman, M.C. Antimicrobial efficacy of phytochemicals against Bacillus cereus in reconstituted infant rice cereal. Food Microbiol. 2018, 69, 189–195. [Google Scholar] [CrossRef]
- RamaRao, N.; Tran, S.-L.; Marin, M.; Vidic, J. Advanced Methods for Detection of Bacillus cereus and Its Pathogenic Factors. Sensors 2020, 20, 2667. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.; Su, H.; Ding, H.; Sun, X.; Gao, H.; Geng, Y.; Wang, Z. Rapid analysis of Escherichia coli O157:H7 using isothermal recombinase polymerase amplification combined with triple-labeled nucleotide probes. Mol. Cell. Probes 2020, 50, 101501. [Google Scholar] [CrossRef]
- Yao, L.; Lu, J.; Qu, M.; Jiang, Y.; Li, F.; Guo, Y.; Wang, L.; Zhai, Y. Methodology and application of PCR-RFLP for species identification in tuna sashimi. Food Sci. Nutr. 2020, 8, 3138–3146. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Lee, J.-L. Development of a multiplex real-time recombinase polymerase amplification (RPA) assay for rapid quantitative detection of Campylobacter coli and jejuni from eggs and chicken products. Food Control. 2017, 73, 1247–1255. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Zeng, H.; Liu, X.; Jiang, W.; Wang, Y.; Ouyang, W.; Tang, X. RPA-Cas12a-FS: A frontline nucleic acid rapid detection system for food safety based on CRISPR-Cas12a combined with recombinase polymerase amplification. Food Chem. 2021, 334, 127608. [Google Scholar] [CrossRef]
- Chang, C.-C.; Chen, C.-C.; Wei, S.-C.; Lu, H.-H.; Liang, Y.-H.; Lin, C.-W. Diagnostic Devices for Isothermal Nucleic Acid Amplification. Sensors 2012, 12, 8319–8337. [Google Scholar] [CrossRef] [PubMed]
- Crannell, Z.A.; Rohrman, B.; Richards-Kortum, R. Equipment-Free Incubation of Recombinase Polymerase Amplification Reactions Using Body Heat. PLoS ONE 2014, 9, e112146. [Google Scholar] [CrossRef]
- Deng, H.; Gao, Z. Bioanalytical applications of isothermal nucleic acid amplification techniques. Anal. Chim. Acta 2015, 853, 30–45. [Google Scholar] [CrossRef]
- Li, J.; Ma, B.; Fang, J.; Zhi, A.; Chen, E.; Xu, Y.; Yu, X.; Sun, C.; Zhang, M. Recombinase Polymerase Amplification (RPA) Combined with Lateral Flow Immunoassay for Rapid Detection of Salmonella in Food. Foods 2020, 9, 27. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, X.; Wang, Y.; Shen, H.; Jiang, G.; Dong, J.; Zhao, P.; Gao, S. A Real-Time Recombinase Polymerase Amplification Method for Rapid Detection of Vibrio vulnificus in Seafood. Front. Microbiol. 2020, 11, 586981. [Google Scholar] [CrossRef]
- Du, X.-J.; Zang, Y.-X.; Liu, H.-B.; Li, P.; Wang, S. Rapid Detection of Staphylococcus aureus Via Recombinase Polymerase Amplification Combined with Lateral Flow Strip. Food Anal. Methods 2018, 11, 2296–2306. [Google Scholar] [CrossRef]
- Gao, W.; Huang, H.; Zhang, Y.; Zhu, P.; Yan, X.; Fan, J.; Chen, X. Recombinase Polymerase Amplification-Based Assay for Rapid Detection of Listeria monocytogenes in Food Samples. Food Anal. Methods 2017, 10, 1972–1981. [Google Scholar] [CrossRef]
- Choi, G.; Jung, J.H.; Park, B.H.; Oh, S.J.; Seo, J.H.; Choi, J.S.; Kim, D.H.; Seo, T.S. A centrifugal direct recombinase polymerase amplification (direct-RPA) microdevice for multiplex and real-time identification of food poisoning bacteria. Lab Chip 2016, 16, 2309–2316. [Google Scholar] [CrossRef]
- Ahn, H.; Batule, B.S.; Seok, Y.; Kim, M.-G. Single-Step Recombinase Polymerase Amplification Assay Based on a Paper Chip for Simultaneous Detection of Multiple Foodborne Pathogens. Anal. Chem. 2018, 90, 10211–10216. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Li, J.; Chen, K.; Yu, X.; Sun, C.; Zhang, M. Multiplex Recombinase Polymerase Amplification Assay for the Simultaneous Detection of Three Foodborne Pathogens in Seafood. Foods 2020, 9, 278. [Google Scholar] [CrossRef]
- Li, K.; Luo, Y.; Huang, K.; Yang, Z.; Wan, Y.; Xu, W. Single universal primer recombinase polymerase amplification-based lateral flow biosensor (SUP-RPA-LFB) for multiplex detection of genetically modified maize. Anal. Chim. Acta 2020, 1127, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wei, X.; Huang, R.; Sun, X.; Jing, J.; Gao, H.; Geng, Y.; Dong, C.; Jiang, C. Advance in RPA detection technologies of foodborne pathogenic bacteria. Sci. Technol. Food Ind. 2018, 39, 329–334. [Google Scholar]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical Evaluation of Two Primers Commonly Used for Amplification of Bacterial 16S rRNA Genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef]
- Ran, D.D.; Yu, X.-H.; Song, D.Z.; Jian, L.I. Advances in research on virulence-related factors in Shigella. J. Southwest Univ. Natl. (Nat. Sci. Ed.) 2012, 38, 390–395. [Google Scholar]
- Guinebretière, M.-H.; Broussolle, V.; Nguyen-The, C. Enterotoxigenic Profiles of Food-Poisoning and Food-Borne Bacillus cereus Strains. J. Clin. Microbiol. 2002, 40, 3053–3056. [Google Scholar] [CrossRef]
- General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China. Rapid Detection of Many Pathogenic Bacteria in Food PCR Method. Available online: https://hbba.sacinfo.org.cn/attachment/onlineRead/a904658c962953ee3986d2a1be3c08e1 (accessed on 6 March 2023).
- Hansen, B.M.; Hendriksen, N.B. Detection of enterotoxic Bacillus cereus and Bacillus thuringiensis strains by PCR analysis. Appl. Environ. Microbiol. 2001, 67, 185–189. [Google Scholar] [CrossRef]
- Bian, Z.; Liu, W.; Jin, J.; Hao, Y.; Jiang, L.; Xie, Y.; Zhang, H. Development of a recombinase polymerase amplification assay with lateral flow dipstick (RPA-LFD) for rapid detection of Shigella spp. and enteroinvasive Escherichia coli. PLoS ONE 2022, 17, e0278869. [Google Scholar] [CrossRef]
- Liu, L.; Nan, H.; Sun, X.; Jiang, Y.; Wang, J.; Wang, J. Development and Application of Real-Time Recombinase Polymerase Amplification Assay for Detection of Bacillus cereus in Food. J. Food Sci. Technol. 2018, 36, 89–94. [Google Scholar]
- Shi, Y.; Xu, C.; Yu, B.; Lu, Y.; Mei, L. Research Progress in Recombinase Polymerase Amplification(RPA). Chin. J. Virol. 2020, 36, 522–532. [Google Scholar]
- Cheng, N.; Song, Y.; Shi, Q.; Du, D.; Liu, D.; Luo, Y.; Xu, W.; Lin, Y. Au@Pd Nanopopcorn and Aptamer Nanoflower Assisted Lateral Flow Strip for Thermal Detection of Exosomes. Anal. Chem. 2019, 91, 13986–13993. [Google Scholar] [CrossRef]
- Wei, Y.; Li, L.; Liu, Y.; Xiang, S.; Zhang, H.; Yi, L.; Shang, Y.; Xu, W. Identification techniques and detection methods of edible fungi species. Food Chem. 2022, 374, 131803. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ID of Strains |
|---|---|
| Shigella | CMCC[B] 51105 |
| 72 * | |
| 92 * | |
| Bacillus cereus | CMCC[B] 63303 |
| 121 * | |
| 124 * | |
| Escherichia coli O157:H7 | NCTC 12900 |
| Salmonella | CMCC[B] 50094 |
| Vibrio parahaemolyticus | ATCC 17802 |
| Enterobacter sakazakii | ATCC 29544 |
| Listeria monocytogenes | CMCC[B] 54002 |
| Staphylococcus aureus | CMCC[B] 26003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, S.; Zhang, H.; Cha, X.; Lin, Y.; Shang, Y. A New Duplex Recombinase Polymerase Amplification (D-RPA) Method for the Simultaneous and Rapid Detection of Shigella and Bacillus cereus in Food. Foods 2023, 12, 1889. https://doi.org/10.3390/foods12091889
Xiang S, Zhang H, Cha X, Lin Y, Shang Y. A New Duplex Recombinase Polymerase Amplification (D-RPA) Method for the Simultaneous and Rapid Detection of Shigella and Bacillus cereus in Food. Foods. 2023; 12(9):1889. https://doi.org/10.3390/foods12091889
Chicago/Turabian StyleXiang, Shuna, Hanyue Zhang, Xiaoyan Cha, Yingting Lin, and Ying Shang. 2023. "A New Duplex Recombinase Polymerase Amplification (D-RPA) Method for the Simultaneous and Rapid Detection of Shigella and Bacillus cereus in Food" Foods 12, no. 9: 1889. https://doi.org/10.3390/foods12091889