
The Impact of In Vitro Digestion on the Polyphenol Content and Antioxidant Activity of Spanish Ciders

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Samples
2.3. In Vitro Digestion
- Gastro-intestinal digestion (GID): simulating the digestive process within the stomach and in the intestinal lumen and preparing GID samples with additions of pepsin, pancreatin, and bile salts.
- Oral, Gastric, and Intestinal Digestion (OGID): simulating the oral, gastric, and intestinal digestive process and preparing the OGID samples with additions of α-amylase, pepsin, pancreatin, and bile salts. The enzymatic digestive process was prepared following the protocol of Miller et al. [25], adapted by Rufián-Henares and Delgado-Andrade [26], and modified by Pastoriza et al. [27] to include the preliminary oral digestion phase. Briefly, the physiological extraction was performed in 3 stages: oral, gastric, and intestinal. In the first place, oral digestion was simulated. Therefore, salivary α-amylase was added to 10 mL of cider, which was incubated and continuously stirred in a vertical rotary shaker (STUART ROTATOR SB3) inside an incubator (Heratherm THERMO SCIENTIFIC) for half an hour at 37 °C. Subsequently, the pH was adjusted to 2, and a pepsin solution was added in the phase simulating gastric digestion. Once again, the sample was incubated and continuously stirred for 2 h at 37 °C. The last phase simulated the digestive processes of the small intestine. The pH was adjusted to 5.3, then the pancreatin solution and bile salts were added, and the pH was once again adjusted to 7.5. The samples were incubated and stirred for 2 h at 37 °C. After completing digestion, the samples were placed in a bath at a temperature of 100 °C for 4 min and then ice-cooled. Finally, samples were centrifuged (HERAEUS Megafuge 16R) at 5500× g for 1 h at 4 °C. Afterward, a soluble, bio-accessible fraction was obtained that was stored at −30 °C until its analysis. No insoluble fraction remained; therefore, the subsequent fermentation phase was not carried out.
2.4. Determination of Total Polyphenols
2.5. Chemical Methods to Determine Antioxidant Capacity
2.5.1. DPPH Assay
2.5.2. ABTS Assay
2.5.3. FRAP Assay
2.5.4. ORAC Assay
2.6. Percentage Recovery
2.7. Statistical Analysis
3. Results and Discussion
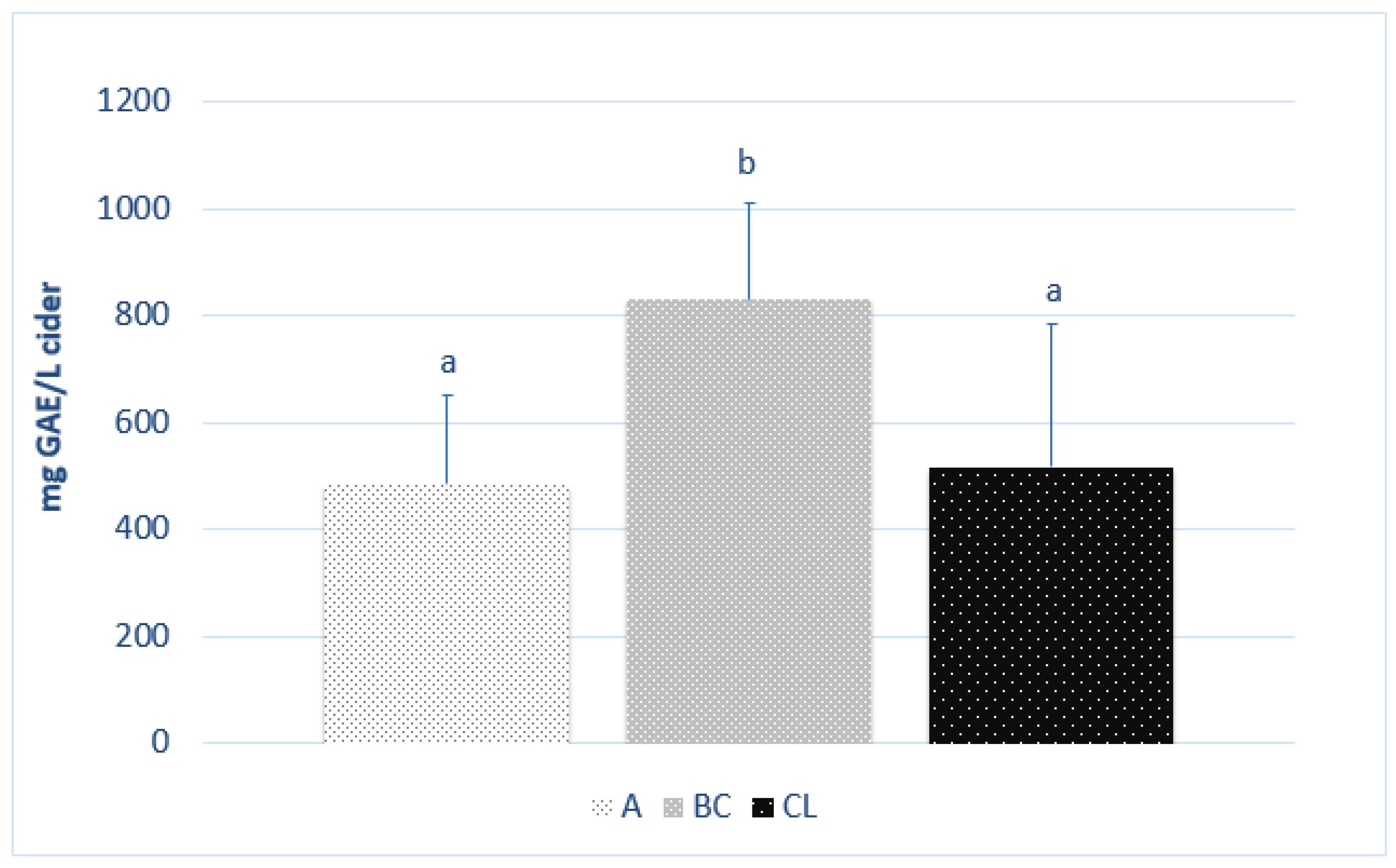
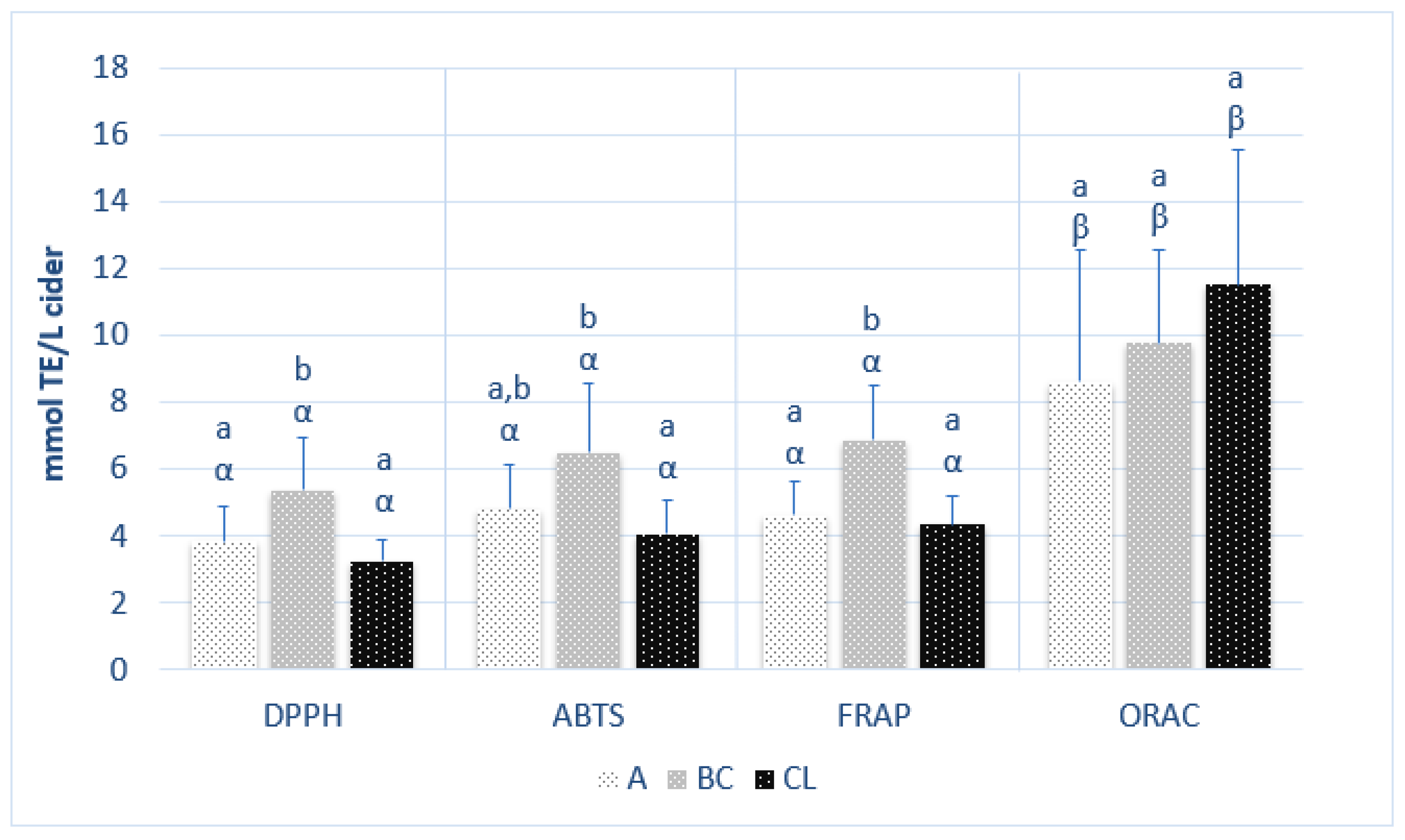
3.1. Polyphenol Content and Antioxidant Activity in Ciders from Different Origins
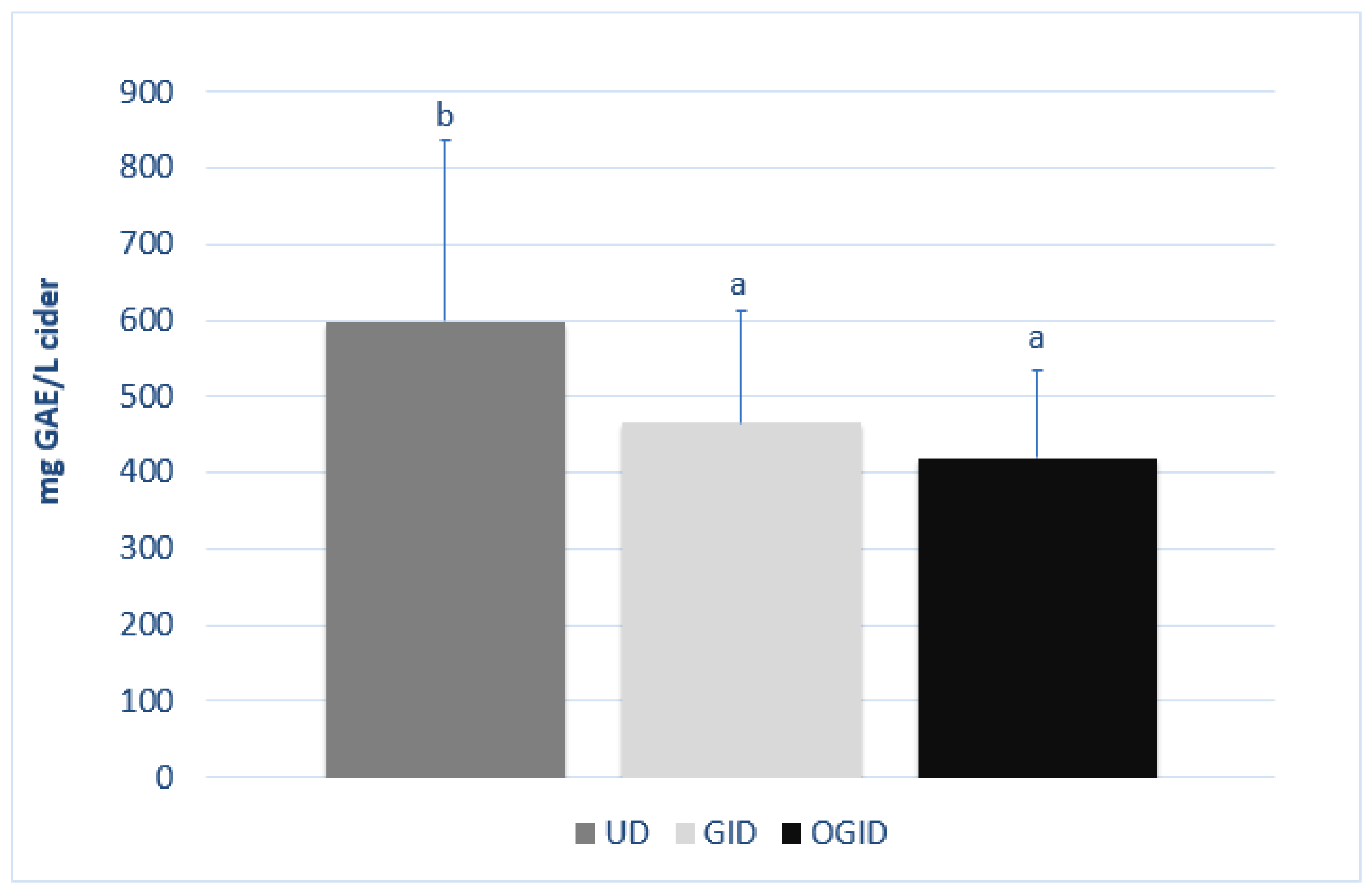
3.2. Effect of In Vitro Digestion on the Phenolic Content of Ciders
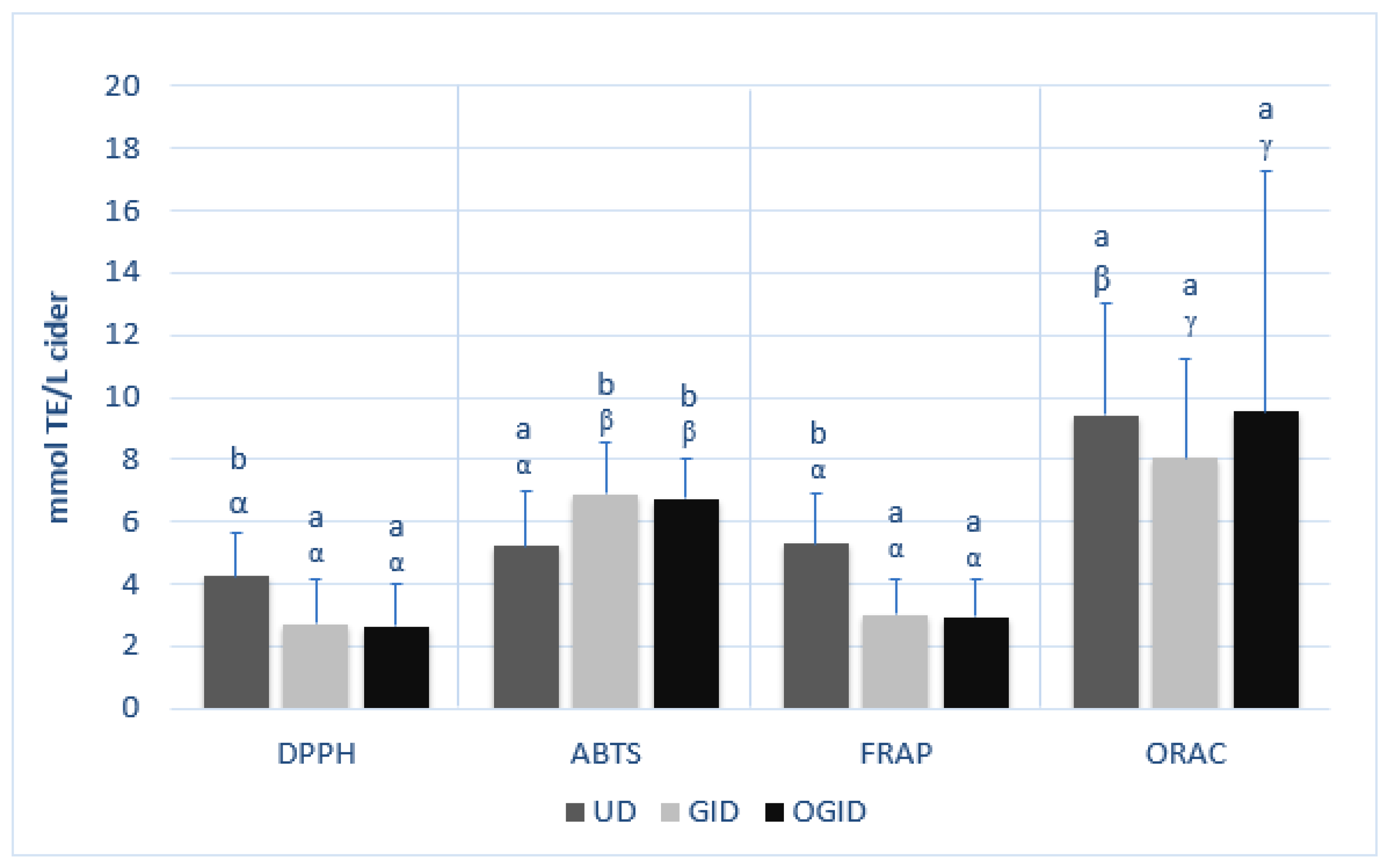
3.3. Effect of In Vitro Digestion on Antioxidant Activity of Ciders
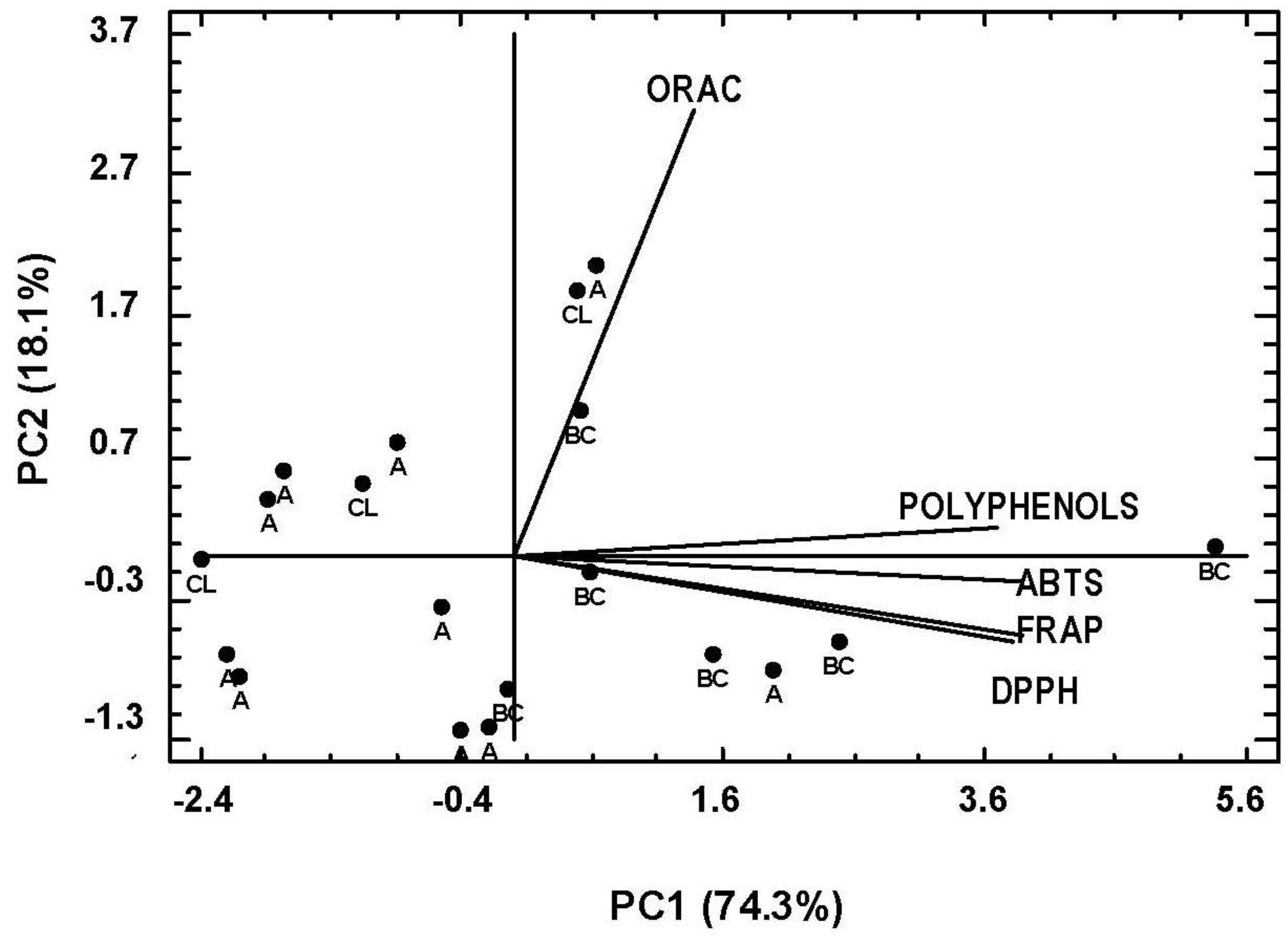
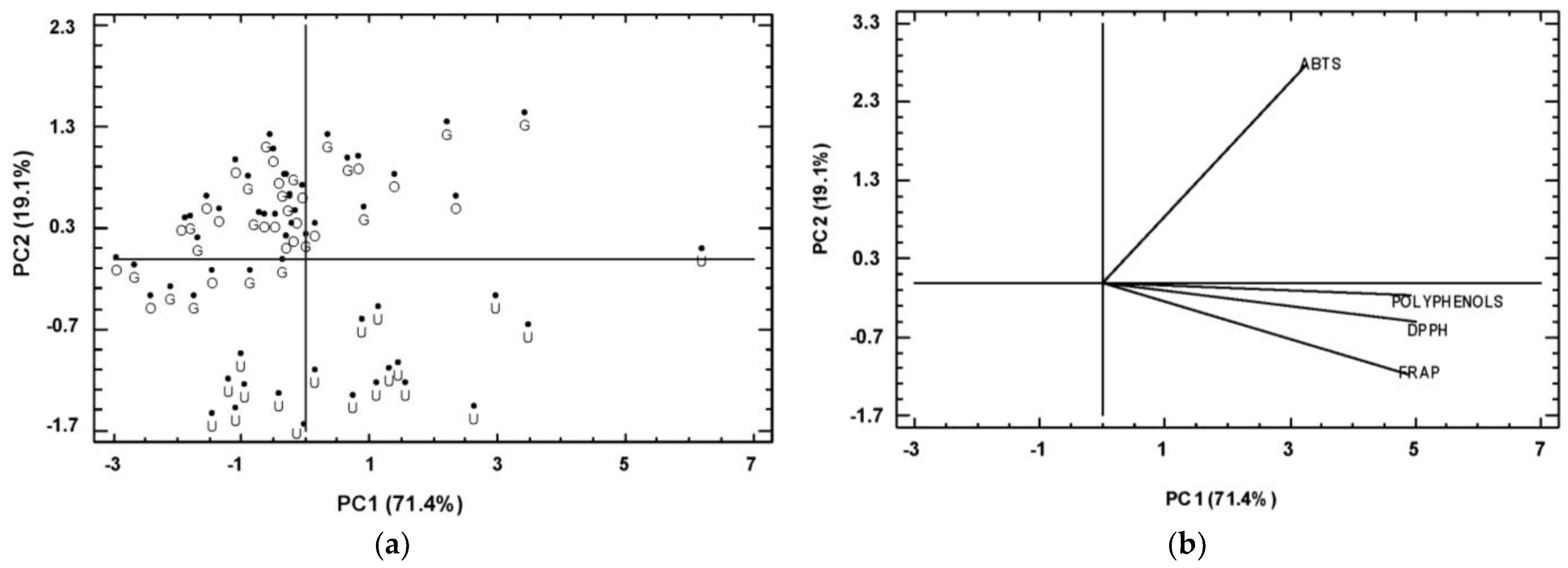
3.4. Correlation Analysis
3.5. Limitations
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Real Decreto 72/2017; de 10 de Febrero, por el que se Aprueba la Norma de Calidad de las Diferentes Categorías de la Sidra Natural y de la Sidra. Boletín Oficial del Estado n° 44, de 21 de Febrero de 2017; Ministerio de la Presidencia, Relaciones con las Cortes y Memoria Democrática: Madrid, España, 2017; pp. 11416–11423.
- Moreiras, O.; Carbajal, A.; Cabrera, L.; Cuadrado, C. Tablas de Composición de Alimentos, 18th ed.; Pirámide: Madrid, Spain, 2016; pp. 113–115. [Google Scholar]
- Picinelli, A.; Diñero, Y.; Mangas, J.; Rodríguez, R.; Suárez, B. Phenolic and antioxidant composition of cider. J. Food Compos. Anal. 2009, 22, 644–648. [Google Scholar] [CrossRef]
- Kumar, S.; Krishna Chaitanya, R.; Preedy, V.R. Assessment of antioxidant potential of dietary components. In HIV/AIDS: Oxidative Stress and Dietary Antioxidants; Preedy, V.R., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 239–253. [Google Scholar]
- Shahidi, F.; Zhong, Y.; Chandrasekara, A. Antioxidants and human health. In Cereals and Pulses: Nutraceutical Properties and Health Benefits, 1st ed.; Yu, L.L., Tsao, R., Shahidi, F., Eds.; John Wiley & Sons, Inc.: Chichester, UK, 2012; pp. 273–308. [Google Scholar]
- Lea, A.G.H.; Beech, F. The phenolics of ciders: Effect of cultural conditions. J. Sci. Food Agric. 1978, 29, 493–496. [Google Scholar] [CrossRef]
- Laaksonen, O.; Kuldjärv, R.; Paalme, T.; Virkki, M.; Yang, B. Impact of apple cultivar, ripening stage, fermentation type and yeast strain on phenolic composition of apple ciders. Food Chem. 2017, 233, 29–37. [Google Scholar] [CrossRef]
- Marks, S.C.; Mullen, W.; Crozier, A. Flavonoid and Hydroxycinnamate Profiles of English Apple Ciders. J. Agric. Food Chem. 2007, 55, 8723–8730. [Google Scholar] [CrossRef]
- Alonso-Salces, R.M.; Guyot, S.; Herrero, C.; Berrueta, L.A.; Drilleau, J.F.; Gallo, B.; Vicente, F. Chemometric characterisation of Basque and French ciders according to their polyphenolic profiles. Anal. Bioanal. Chem. 2004, 379, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Sanoner, P.; Guyot, S.; Marnet, N.; Molle, D.; Drilleau, J.P. Polyphenol profiles of French cider apple varieties (Malus domestica sp.). J. Agric. Food Chem. 1999, 47, 4847–4853. [Google Scholar] [CrossRef] [PubMed]
- Budak, H.N.; Ozçelik, F.; Güzel-Seydim, Z.B. Antioxidant activity and phenolic content of apple cider. Turk. J. Agric.-Food Sci. Technol. 2015, 3, 356–360. [Google Scholar] [CrossRef]
- Zuriarrain-Ocio, A.; Zuriarrain, J.; Vidal, M.; Dueñas, M.T.; Berregi, I. Antioxidant activity and phenolic profiles of ciders from the Basque Country. Food Biosci. 2021, 41, 100887. [Google Scholar] [CrossRef]
- Carrillo, C.; Barrio, A.; Cavia, M.M.; Alonso-Torre, S.R. Global antioxidant response of meat. J. Sci. Food Agric. 2017, 97, 2358–2365. [Google Scholar] [CrossRef]
- Carrillo, C.; Rey, R.; Hendrickx, M.; Cavia, M.M.; Alonso-Torre, S.R. Antioxidant capacity of beetroot: Traditional vs novel approaches. Plant Foods Hum. Nutr. 2017, 72, 266–273. [Google Scholar] [CrossRef]
- Zoubiri, L.; Bakir, S.; Barkat, M.; Carrillo, C.; Capanoglu, E. Changes in the phenolic profile, antioxidant capacity and in vitro bioaccsessibility of two Algerian grape varieties, Cardinal and Dabouki (Sabel), during the production of traditional sun-dried raisins and homemade jam. J. Berry Res. 2019, 9, 709–724. [Google Scholar] [CrossRef]
- Guo, R.; Chang, X.; Guo, X.; Brennan, C.B.; Li, T.; Fu, X.; Liu, R.H. Phenolic compounds, antioxidant activity, antiproliferative activity and bioaccessibility of Sea buckthorn (Hippophaë rhamnoides L.) berries as affected by in vitro digestion. Food Funct. 2017, 8, 4229–4240. [Google Scholar] [CrossRef] [PubMed]
- Jara-Palacios, M.J.; Gonçalves, S.; Hernanz, D.; Heredia, F.J.; Romano, A. Effects of in vitro gastrointestinal digestion on phenolic compounds and antioxidant activity of different white winemaking by products extracts. Food Res. Int. 2018, 109, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Barak, T.H.; Celep, E.; Inan, Y.; Yesilada, E. Influence of in vitro human digestion on the bioavailability of phenolic content and antioxidant activity of Viburnum opulus L. (European cranberry) fruit extracts. Ind. Crops Prod. 2019, 131, 62–69. [Google Scholar] [CrossRef]
- Czubinski, J.; Wroblewska, K.; Czyzniejewski, M.; Górnaś, P.; Kachlicki, P.; Sigera, A. Bioaccessibility of defatted lupin seed phenolic compounds in a standardized static in vitro digestion system. Food Res. Int. 2019, 116, 1126–1134. [Google Scholar] [CrossRef]
- Qin, W.; Ketnawa, S.; Ogawa, Y. Effect of digestive enzymes and pH on variation of bioavailability of green tea during simulated in vitro gastrointestinal digestion. Food Sci. Hum. Wellness 2022, 11, 669–675. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Balance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—an international consensus. Food Funct. 2014, 5, 1113. [Google Scholar] [CrossRef]
- Álvarez, J.; Pastoriza, S.; Alonso-Olalla, R.; Delgado-Andrade, C.; Rufián-Henares, L.A. Nutritional and physicochemical characteristic of commercial Spanish citrus juices. Food Chem. 2014, 164, 396–405. [Google Scholar] [CrossRef]
- European Cider and Fruit Wine Association. Available online: https://aicv.org/en/publications (accessed on 17 April 2023).
- Rodríguez, R.; Pinicelli, A.; Suárez, B. Phenolic profile of asturian (Spain) natural cider. J. Agric. Food Chem. 2006, 54, 120–124. [Google Scholar] [CrossRef]
- Miller, D.D.; Schricker, B.R.; Rasmussen, R.R.; Van Campen, D. An in vitro method for estimation of iron availability from meals. Am. J. Clin. Nutr. 1981, 34, 2248–2256. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; Delgado-Andrade, C. Effect of digestive process on Maillard reaction indexes and antioxidant properties of breakfast cereals. Food Res. Int. 2009, 42, 394–400. [Google Scholar] [CrossRef]
- Pastoriza, S.; Delgado-Andrade, C.; Haro, A.; Rufián-Henares, J. A physiologic approach to test the global antioxidant response of foods. The GAR method. Food Chem. 2011, 129, 1926–1932. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteau reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Miller, N.J.; Diplock, A.T.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Strain, J.J. Ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Cao, G.; Alessio, H.M.; Cutler, R.G. Oxygen-Radical absorbance Capacity assay for antioxidants. Free Radic. Biol. Med. 1993, 14, 303311. [Google Scholar] [CrossRef]
- Ljevar, A.; Ćurko, N.; Tomašević, M.; Radošević, K.; Srček, V.G.; Ganić, K.K. Phenolic composition, antioxidant capacity and in vitro cytotoxicity assessment of fruit wines. Food Technol. Biotechnol. 2016, 54, 145–155. [Google Scholar] [CrossRef]
- Ye, M.; Yue, T.; Yuan, Y. Evolution if polyphenols and organic acids during the fermentation of apple cider. J. Sci. Food Agric. 2014, 94, 2951–2957. [Google Scholar] [CrossRef]
- Van der Sluis, A.A.; Dekker, M.; Skrede, G.; Jongen, W.M.F. Activity and concentration of polyphenolic antioxidants in apple juice. 1. Effect of existing methods. J. Agric. Food Chem. 2002, 50, 7211–7219. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Goñi, I. Antioxidant capacity of the Spanish Mediterranean diet. Food Chem. 2006, 94, 442–444. [Google Scholar] [CrossRef]
- Seeram, N.P.; Aviram, M.; Yanjun, Z.; Henning, S.M.; Feng, L.; Dreher, M.; Hber, D. Comparison of antioxidant potency of commonly consumes polyphenol-rich beverages in the United States. J. Agric. Food Chem. 2008, 56, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.J.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin–Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. J. Agric. Food Chem. 2004, 52, 4026–4037. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutr. J. 2010, 9, 3. [Google Scholar] [CrossRef]
- Friedman, M.; Jürgens, H.S. Effect of pH on the stability of plant phenolic compounds. J. Agric. Food Chem. 2000, 48, 2101–2110. [Google Scholar] [CrossRef]
- Oliveira, D.; Laitmer, C.; Parpot, P.; Gill, C.I.; Oliveira, R. Antioxidant and antigenotoxic activities of Ginkgo biloba L. leaf extract are retained after in vitro gastrointestinal digestive conditions. Eur. J. Nutr. 2020, 59, 465–476. [Google Scholar] [CrossRef]
- Liu, X.; Shi, J.; Yi, J.; Zhang, X.; Ma, Q.; Cai, S. The effect of in vitro simulated gastrointestinal digestion on phenolic bioaccessibility and bioactivities of Prinsepia utilis Royle fruits. LWT-Food Sci. Technol. 2021, 138, 110782. [Google Scholar] [CrossRef]
- Celep, E.; Charehsaz, M.; Akyüz, S.; Acar, E.T.; Yeselida, E. Effect of in vitro gastrointestinal digestion on the bioavailability of phenolic components and the antioxidant potentials of some Turkish fruit wines. Food Res. Int. 2015, 78, 209–2015. [Google Scholar] [CrossRef] [PubMed]
- Wootton-Beard, P.C.; Ryan, L. A beetroot juice shot is a significant and convenient source of bioaccessible antioxidants. J. Funct. Foods 2011, 3, 329–334. [Google Scholar] [CrossRef]
- Bhatt, A.; Patel, V. Evaluation of Actual Antioxidant Capacity of Papaya (Carica papaya) Using an In Vitro Gastrointestinal Model. Int. J. Fruit Sci. 2015, 15, 26–35. [Google Scholar] [CrossRef]
- He, M.; Zeng, J.; Zhai, L.; Liu, Y.; Wu, H.; Zhang, R.; Li, Z.; Xia, E. Effect of in vitro simulated gastrointestinal digestion on polyphenol and polysaccharide content and their biological activities among 22 fruit juices. Food Res. Int. 2017, 102, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Gao, J.; Wei, Z.; Shahidi, F. Effect of in vitro digestion on phenolics and antioxidant activity of red and yellow colored pea hulls. Food Chem. 2021, 337, 127606. [Google Scholar] [CrossRef]
- Sun, D.; Huang, S.Q.; Cai, S.B.; Cao, J.X.; Han, P. Digestion property and synergistic effect on biological activity of purple rice (Oryza sativa L.) anthocyanins subjected to a simulated gastrointestinal digestion in vitro. Food Res. Int. 2015, 78, 114–123. [Google Scholar] [CrossRef]
- Bermúdez-Soto, M.J.; Tomás-Barberán, F.A.; García-Conesa, M.T. Stability of polyphenols in chokeberry (Aronia melanocarpa) subjected to in vitro gastric and pancreatic digestion. Food Chem. 2007, 102, 865–874. [Google Scholar] [CrossRef]
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Złotek, U.; Zielińska, E. Digestion and bioavailability of bioactive phytochemicals. Int. J. Food Sci. Technol. 2017, 52, 291–305. [Google Scholar] [CrossRef]
- Moyo, S.M.; Serem, J.C.; Bester, M.J.; Mavumengwana, V.; Kayitesi, E. The impact of boling and in vitro human digestion of Solanum nigrum complex (Black nightshade) on phenolic compounds bioactivity and bioaccessibility. Food Res. Int. 2020, 137, 109720. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.H. Degradation kinetics of fisetin and quercetin in solutions affected by medium pH, temperature and co-existing proteins. J. Serb. Chem. Soc. 2016, 81, 243–253. [Google Scholar] [CrossRef]
- Carrillo, C.; Kamiloglu, S.; Grootaert, C.; Van Camp, J.; Hendrickx, M. Co-ingestion of black carrot and strawberry. Effects on anthocyanin stability, bioaccessibility and uptake. Foods. 2020, 9, 1595. [Google Scholar] [CrossRef] [PubMed]
- Caicedo-Lopez, L.H.; Luzardo-Ocampo, I.; Cuellar-Nuñez, M.L.; Campos-Vega, R.; Mendoza, S.; Loarca-Piña, G. Effect of the in vitro gastrointestinal digestion on free-phenolic compounds and mono/oligosaccharides from Moringa oleifera leaves: Bioaccessibility, intestinal permeability and antioxidant capacity. Food Res. Int. 2019, 120, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Pavan, V.; Soriano Sancho, R.A.; Pastore, G.M. The effect of in vitro digestion on the antioxidant activity of fruit extracts (Carica papaya, Artocarpus heterophillus and Annona marcgravii). LWT—Food Sci. Technol. 2014, 59, 1247–1251. [Google Scholar] [CrossRef]
- Ryan, L.; Prescott, S.L. Stability of the antioxidant capacity of twenty-five commercially available fruit juices subjected to an in vitro digestion. Int. J. Food Sci. Technol. 2010, 45, 1191–1197. [Google Scholar] [CrossRef]
- Baker, I.; Chohan, M.; Opara, E.I. Impact of Cooking and Digestion, In Vitro, on the Antioxidant Capacity and Anti-Inflammatory Activity of Cinnamon, Clove and Nutmeg. Plant Foods Hum. Nutr. 2013, 68, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, S.; Moreira, E.; Andrade, P.B.; Valentao, P.; Romano, A. Effect of in vitro gastrointestinal digestion on the total phenolic contents and antioxidant activity of wild Mediterranean edible plant extracts. Eur. Food Res. Technol. 2019, 245, 753–762. [Google Scholar] [CrossRef]
- Spínola, V.; Llorent-Martínez, E.J.; Castilho, P.C. Antioxidant polyphenols of Madeira sorrel (Rumex maderensis): How do they survive to in vitro simulated gastrointestinal digestion? Food Chem. 2018, 259, 105–112. [Google Scholar] [CrossRef]
- Bayliak, M.M.; Burdyliuk, N.I.; Lushchak, V.I. Effects of pH on antioxidant and prooxidant properties of common medicinal herbs. Open Life Sci. 2016, 11, 298–307. [Google Scholar] [CrossRef]
- Serrano, J.; Goñi, I.; Saura-Calixto, F. Food antioxidant capacity determined by chemical methods may underestimate the physiological antioxidant capacity. Food Res. Int. 2007, 40, 15–21. [Google Scholar] [CrossRef]
- Albishi, T.; John, J.A.; Al-Khalifa, A.S.; Shahidi, F. Phenolic content and antioxidant activities of selected potato varieties and their processing by-products. J. Funct. Foods. 2013, 5, 590–600. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khahnizadeh, S. Which polyphenolic compounds contribute to the total antioxidant activities of apple? J. Agric. Food Chem. 2005, 53, 4989–4995. [Google Scholar] [CrossRef] [PubMed]
- Vanzani, P.; Rossetto, M.; Rigo, A.; Vrhovsek, U.; Mattivi, F.; D’ Amato, E.; Scarpa, M. Major phytochemicals in apple cultivars: Contribution to peroxyl radical trapping efficiency. J. Agric. Food Chem. 2005, 53, 3377–3382. [Google Scholar] [CrossRef] [PubMed]
- Gullon, B.; Pintado, M.E.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. In vitro gastrointestinal digestion of pomegranate peel (Punica granatum) flour obtained from co-products: Changes in the antioxidant potential and bioactive compounds stability. J. Funct. Foods 2015, 19, 617–628. [Google Scholar] [CrossRef]
- Lucas-Gonzalez, R.; Navarro-Coves, S.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Assessment of polyphenolic profile stability and changes in the antioxidant potential of maqui berry (Aristotelia chilensis (Molina) Stuntz) during in vitro gastrointestinal digestion. Ind. Crops Prod. 2016, 94, 774–782. [Google Scholar] [CrossRef]
- Gorinstein, S.; Haruenkit, R.; Poovarodom, S.; Vearasilp, S.; Ruamsuke, P.; Namiesnik, J.; Leontowicz, M.; Leontowicz, H.; Suhaj, M.; Sheng, G.P. Some analytical assays for the determination of bioactivity of exotic fruits. Phytochem. Anal. 2010, 21, 355–362. [Google Scholar] [CrossRef]
- Chun, O.K.; Kim, D.O.; Moon, H.Y.; Kang, H.G.; Lee, C.Y. Contribution of individual polyphenolics to total antioxidant capacity of plums. J. Agric. Food Chem. 2003, 51, 7240–7245. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.O.; Chun, O.K.; Kim, Y.J.; Moon, H.Y.; Lee, C.Y. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Ciders | Total Polyphenol Content UD Samples (mg GAE/L Cider) | % Recovery Polyphenols | |
|---|---|---|---|---|
| GID | OGID | |||
| Asturias | A1 | 433 ± 38 | 119 | 122 |
| A2 | 383 ± 28 | 141 | 126 | |
| A3 | 568 ± 22 | 65 | 66 | |
| A4 | 585 ± 95 | 59 | 59 | |
| A5 | 384 ± 17 | 73 | 68 | |
| A6 | 794 ± 81 | 59 | 67 | |
| A7 | 520 ± 3 | 75 | 68 | |
| A8 | 248 ± 39 | 140 | 126 | |
| A9 | 296 ± 48 | 121 | 136 | |
| A10 | 632 ± 16 | 98 | 74 | |
| Basque Country | BC1 | 1153 ± 177 | 58 | 58 |
| BC2 | 655 ± 9 | 80 | 68 | |
| BC3 | 861 ± 13 | 44 | 45 | |
| BC4 | 802 ± 46 | 56 | 56 | |
| BC5 | 679 ± 26 | 75 | 74 | |
| BC6 | 830 ± 46 | 85 | 42 | |
| Castile-and-Leon | CL1 | 338 ± 25 | 76 | 66 |
| CL2 | 826 ± 27 | 93 | 72 | |
| CL3 | 383 ± 19 | 92 | 78 | |
| Average | 598 ± 239 | 85 ± 28 a | 77 ± 28 a | |
| DPPH (mmol TE/L) | % Recovery Antioxidant Activity by DPPH | ABTS (mmol TE/L) | % Recovery Antioxidant Activity by ABTS | FRAP (mmol TE/L) | % Recovery Antioxidant Activity by FRAP | ORAC (mmol TE/L) | % Recovery Antioxidant Activity by ORAC | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Origin | Ciders | UD | GID | OGID | UD | GID | OGID | UD | GID | OGID | UD | GID | OGID |
| Asturias | A1 | 2.35 ± 0.14 | 126 | 126 | 3.75 ± 0.52 | 157 | 167 | 3.82 ± 0.30 | 76 | 72 | 4.84 ± 0.63 | 158 | 95 |
| A2 | 2.69 ± 0.27 | 153 | 97 | 4.22 ± 0.41 | 151 | 178 | 3.47 ± 0.48 | 67 | 62 | 10.36 ± 0.27 | 80 | 85 | |
| A3 | 4.39 ± 0.47 | 33 | 26 | 4.64 ± 0.27 | 99 | 136 | 5.24 ± 0.08 | 45 | 41 | 4.96 ± 2.23 | 93 | 77 | |
| A4 | 4.25 ± 0.36 | 34 | 31 | 5.88 ± 0.38 | 98 | 97 | 4.88 ± 0.09 | 34 | 32 | 4.89 ± 2.64 | 76 | 79 | |
| A5 | 2.95 ± 0.29 | 24 | 14 | 3.25 ± 0.51 | 137 | 134 | 3.48 ± 0.09 | 32 | 21 | 5.53 ± 0.20 | 78 | 81 | |
| A6 | 5.73 ± 0.72 | 37 | 33 | 7.58 ± 0.25 | 86 | 85 | 6.84 ± 0.08 | 40 | 39 | 7.87 ± 0.82 | 80 | 99 | |
| A7 | 3.61 ± 0.13 | 58 | 52 | 3.67 ± 0.33 | 203 | 170 | 4.68 ± 0.42 | 94 | 47 | 11.85 ± 0.24 | 72 | 42 | |
| A8 | 3.22 ± 0.17 | 124 | 139 | 3.87 ± 0.02 | 192 | 181 | 3.72 ± 0.34 | 68 | 80 | 10.08 ± 0.30 | 90 | 61 | |
| A9 | 4.83 ± 0.16 | 40 | 65 | 4.97 ± 0.16 | 164 | 152 | 5.12 ± 0.43 | 66 | 70 | 8.66 ± 0.65 | 81 | 64 | |
| A10 | 4.42 ± 0.19 | 93 | 68 | 6.16 ± 0.26 | 122 | 108 | 4.85 ± 0.10 | 76 | 66 | 17.08 ± 0.88 | 77 | 202 | |
| Basque Country | BC1 | 7.78 ± 0.05 | 63 | 50 | 10.32 ± 0.41 | 95 | 81 | 9.75 ± 0.19 | 51 | 45 | 13.13 ± 0.69 | 71 | 60 |
| BC2 | 4.11 ± 0.46 | 57 | 47 | 5.02 ± 0.31 | 141 | 146 | 5.63 ± 0.07 | 54 | 94 | 6.00 ± 2.61 | 99 | 87 | |
| BC3 | 6.67 ± 0.30 | 20 | 23 | 7.34 ± 0.44 | 75 | 98 | 7.01 ± 0.08 | 28 | 35 | 9.06 ± 0.44 | 55 | 82 | |
| BC4 | 5.46 ± 0.19 | 35 | 48 | 5.65 ± 0.39 | 123 | 131 | 7.30 ± 0.03 | 31 | 41 | 8.15 ± 0.51 | 70 | 124 | |
| BC5 | 4.45 ± 0.29 | 82 | 87 | 5.37 ± 0.54 | 155 | 158 | 6.26 ± 0.31 | 61 | 65 | 9.43 ± 0.52 | 110 | 81 | |
| BC6 | 3.74 ± 0.04 | 64 | 85 | 5.31 ± 0.09 | 153 | 128 | 5.35 ± 0.40 | 49 | 61 | 12.86 ± 0.70 | 111 | 93 | |
| Castile and Leon | CL1 | 2.67 ± 0.34 | 61 | 53 | 2.98 ± 0.02 | 153 | 143 | 3.30 ± 0.29 | 61 | 55 | 7.84 ± 0.18 | 132 | 93 |
| CL2 | 3.86 ± 0.47 | 166 | 167 | 5.06 ± 0.62 | 211 | 173 | 4.92 ± 0.20 | 117 | 104 | 15.87 ± 0.84 | 85 | 93 | |
| CL3 | 3.27 ± 0.32 | 67 | 69 | 4.14 ± 0.02 | 140 | 128 | 4.77 ± 0.23 | 77 | 55 | 10.82 ± 0.64 | 54 | 224 | |
| Average | 4.23 ± 1.40 | 70 ± 43 a | 67 ± 41 a | 5.22 ± 1.76 | 140 ± 38 a | 137 ± 31 a | 5.28 ± 1.61 | 59 ± 23 a | 57 ± 21 a | 9.44 ± 3.58 | 88 ± 26 a | 96 ± 45 a | |
| Correlation Coefficient | DPPH | ABTS | FRAP | ORAC |
|---|---|---|---|---|
| UD | 0.7627 ** | 0.7956 ** | 0.8149 ** | 0.3889 ns |
| GID | 0.7436 ** | 0.7584 ** | 0.6168 * | 0.5995 * |
| OGID | 0.5577 * | 0.7241 ** | 0.6102 * | 0.0701 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavia, M.M.; Arlanzón, N.; Busto, N.; Carrillo, C.; Alonso-Torre, S.R. The Impact of In Vitro Digestion on the Polyphenol Content and Antioxidant Activity of Spanish Ciders. Foods 2023, 12, 1861. https://doi.org/10.3390/foods12091861
Cavia MM, Arlanzón N, Busto N, Carrillo C, Alonso-Torre SR. The Impact of In Vitro Digestion on the Polyphenol Content and Antioxidant Activity of Spanish Ciders. Foods. 2023; 12(9):1861. https://doi.org/10.3390/foods12091861
Chicago/Turabian StyleCavia, Mari Mar, Nerea Arlanzón, Natalia Busto, Celia Carrillo, and Sara R. Alonso-Torre. 2023. "The Impact of In Vitro Digestion on the Polyphenol Content and Antioxidant Activity of Spanish Ciders" Foods 12, no. 9: 1861. https://doi.org/10.3390/foods12091861