


Impact of Pre-Storage Melatonin Application on the Standard, Sensory, and Bioactive Quality of Early Sweet Cherry

 ,
,  ,
,  and
and
Abstract
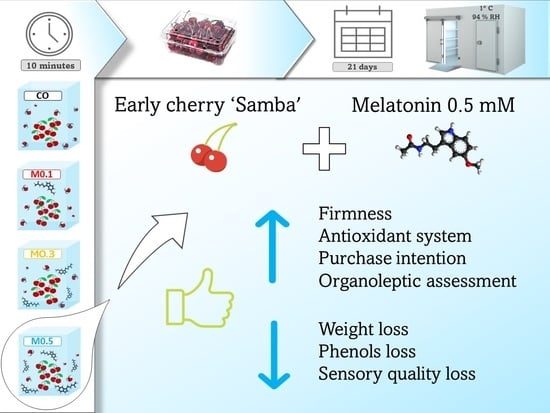
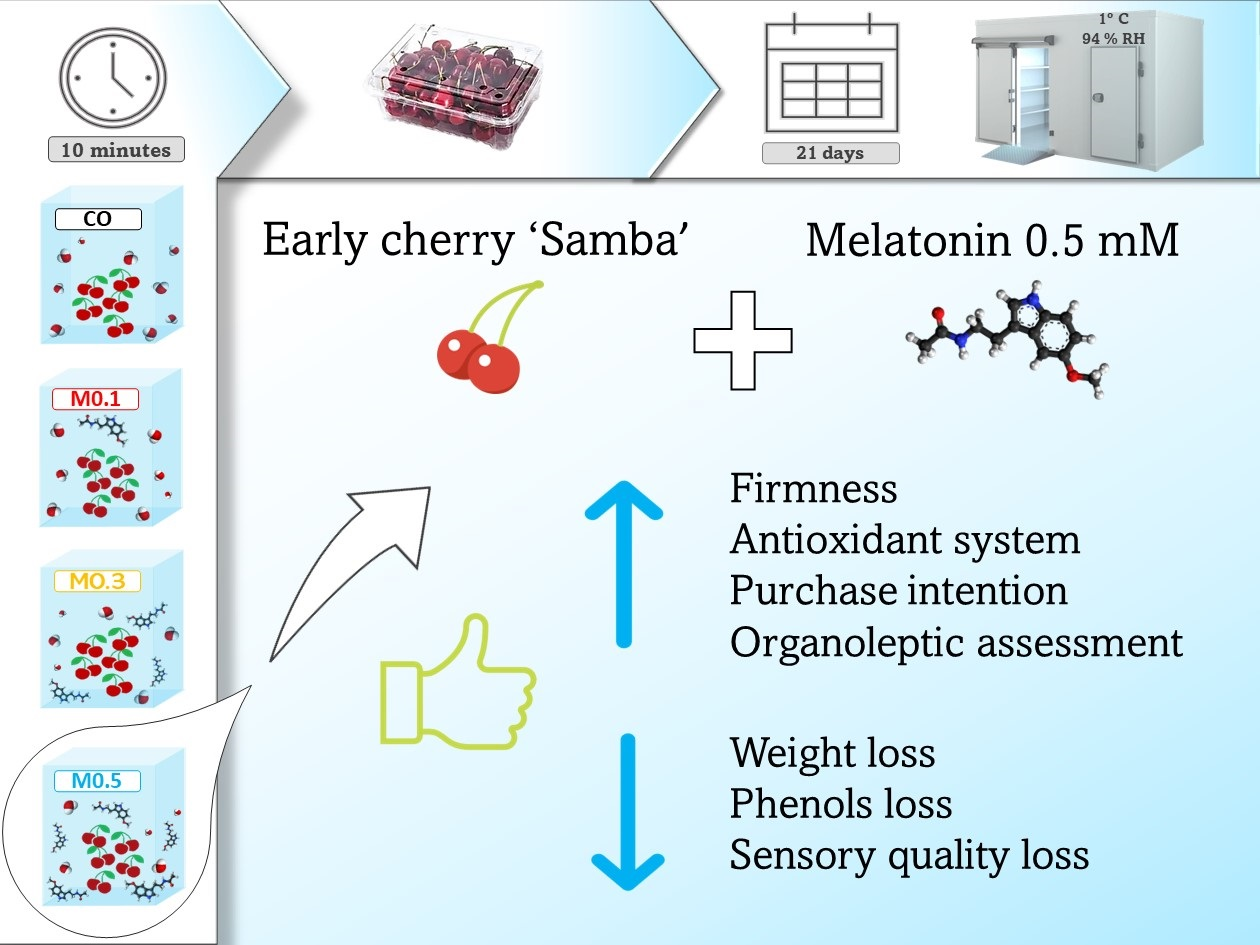
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Fruit Quality Parameters
2.3. Respiration Rate
2.4. Weight Loss and Disorder Evaluation
2.5. Sensory Analysis and Consumer Satisfaction
2.6. Bioactive Compounds
2.6.1. Identification and Quantification of Phenolic Compounds
2.6.2. Antioxidant Activity
2.6.3. Enzymatic Antioxidant Activity
2.7. Statistical Analysis
3. Results
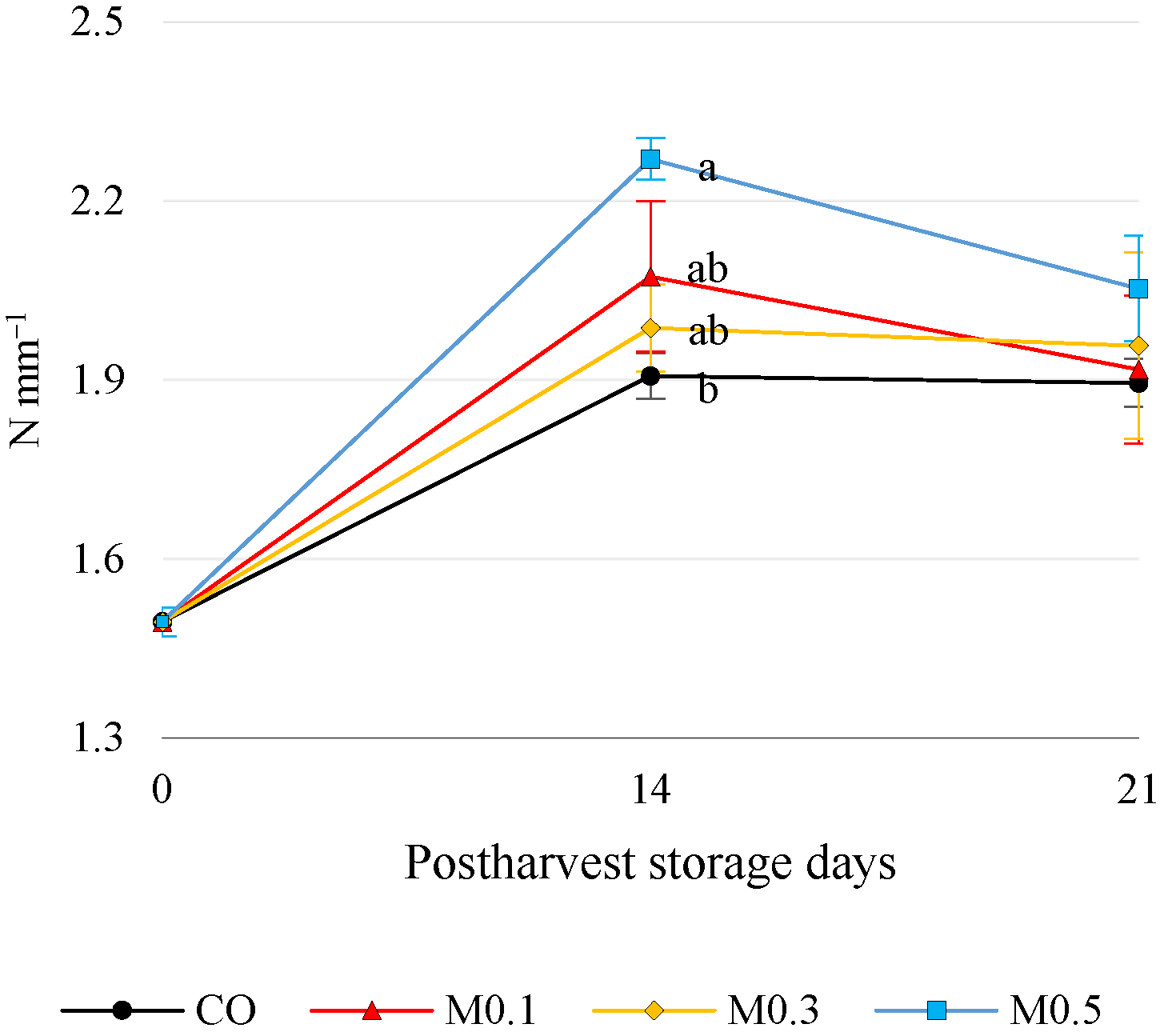
3.1. Quality Parameters
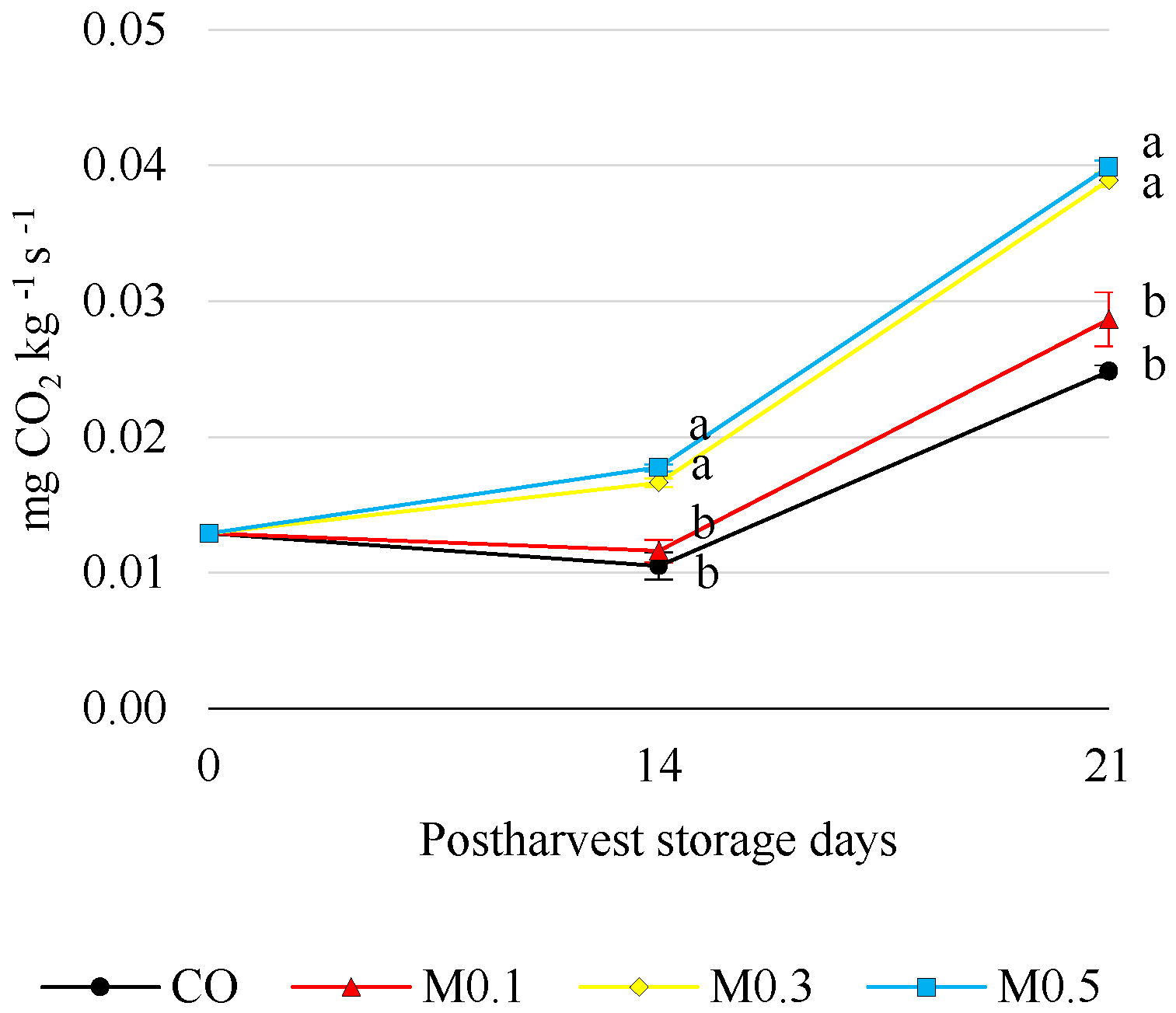
3.2. Respiration Rate
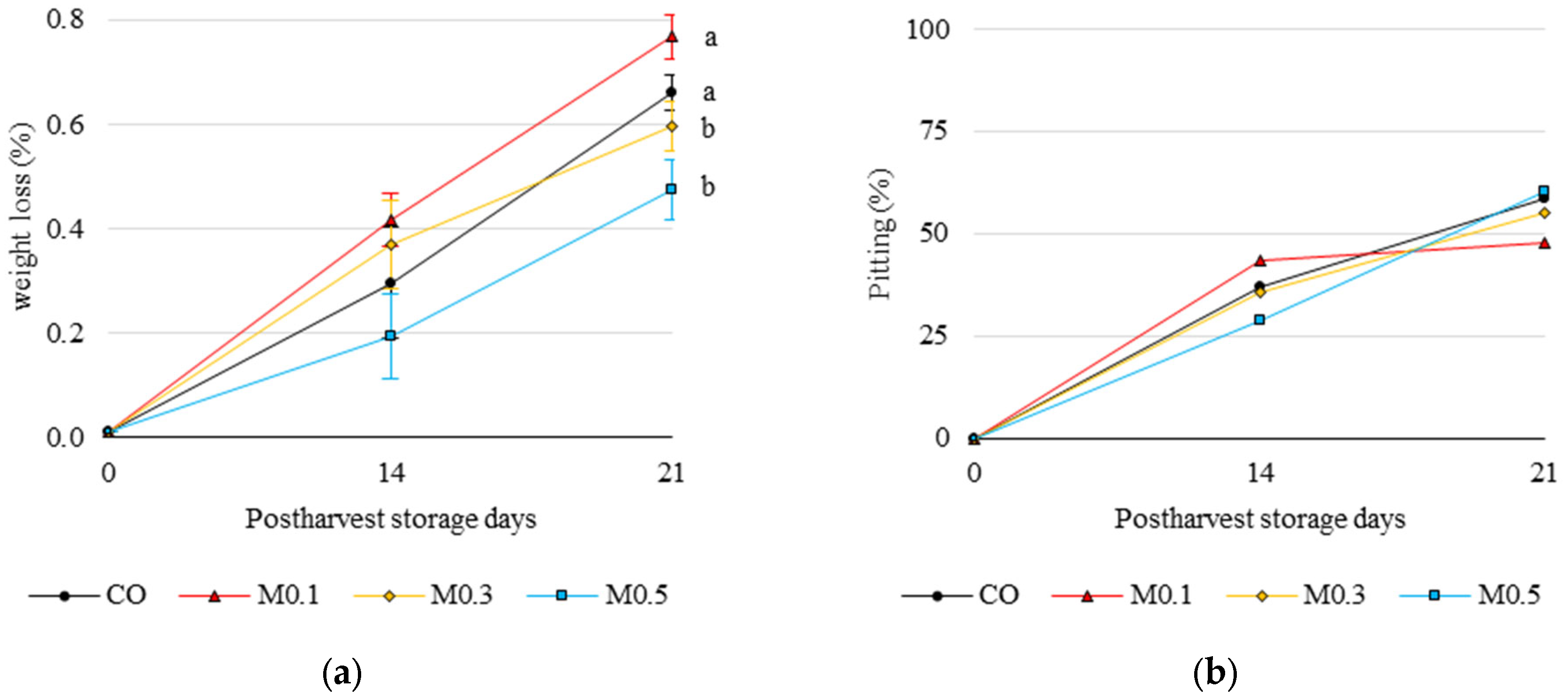
3.3. Weight Loss and Disorder Evaluation
3.4. Sensory Analysis and Consumer Satisfaction
3.5. Bioactive Quality
3.5.1. Identification and Quantification of Phenolic Compounds
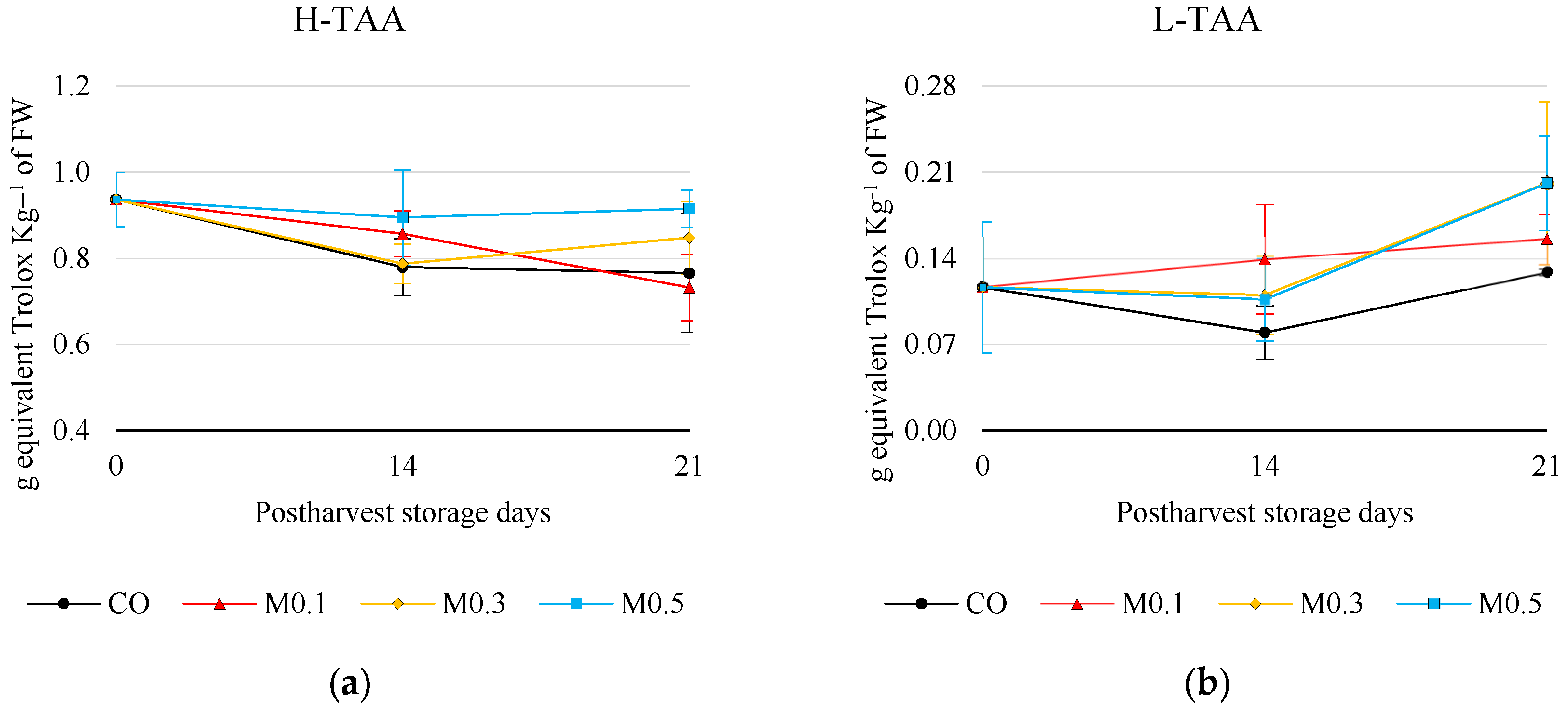
3.5.2. Antioxidant Activity
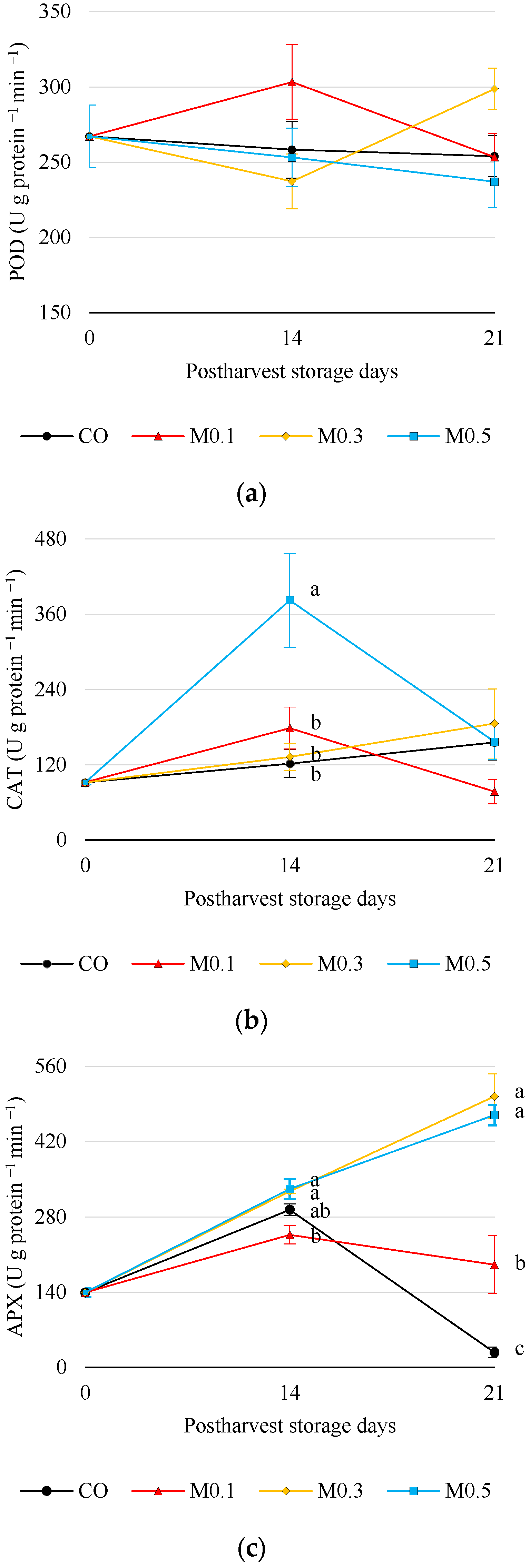
3.5.3. Enzymatic Antioxidant Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 13 April 2023).
- Federación Española de Asociaciones de Productores Exportadores de Frutas; Hortalizas Flores y Plantas Vivas (FEPEX). Available online: https://www.fepex.es/datos-del-sector/comercio-intra-extra-ue-frutas-hortalizas (accessed on 16 February 2023).
- Serradilla, M.J.; Akšić, M.F.; Manganaris, G.A.; Ercisli, S.; González-Gómez, D.; Valero, D. Fruit Chemistry, Nutritional Benefits and Social Aspects of Cherries. In Cherries: Botany, Production and Uses; Quero-García, J., Lezzoni, A., Puławska, J., Lang, G., Eds.; CABI: Wallingford, UK, 2017; pp. 420–441. [Google Scholar]
- Wang, F.; Zhang, X.; Yang, Q.; Zhao, Q. Exogenous Melatonin Delays Postharvest Fruit Senescence and Maintains the Quality of Sweet Cherries. Food Chem. 2019, 301, 125311. [Google Scholar] [CrossRef]
- Sharafi, Y.; Jannatizadeh, A.; Fard, J.R.; Aghdam, M.S. Melatonin Treatment Delays Senescence and Improves Antioxidant Potential of Sweet Cherry Fruits during Cold Storage. Sci. Hortic. 2021, 288, 110304. [Google Scholar] [CrossRef]
- Zoffoli, J.P.; Toivonen, P.; Wang Yan, W.Y. Postharvest Biology and Handling for Fresh Markets. In Cherries: Botany, Production and Uses; Quero-García, J., Lezzoni, A., Puławska, J., Lang, G., Eds.; CABI: Wallingford, UK, 2017; pp. 460–484. [Google Scholar]
- Michailidis, M.; Tanou, G.; Sarrou, E.; Karagiannis, E.; Ganopoulos, I.; Martens, S.; Molassiotis, A. Pre- and Post-Harvest Melatonin Application Boosted Phenolic Compounds Accumulation and Altered Respiratory Characters in Sweet Cherry Fruit. Front. Nutr. 2021, 8, 695061. [Google Scholar] [CrossRef]
- López, L.; Larrigaudière, C.; Giné-Bordonaba, J.; Echeverria, G. Defining Key Parameters and Predictive Markers of ‘Early Bigi’ Cherry Consumer Satisfaction by Means of Differential Storage Scenarios. Postharvest Biol. Technol. 2023, 195, 112117. [Google Scholar] [CrossRef]
- Cabañas, C.M.; Hernández, A.; Serradilla, M.J.; Moraga, C.; Martín, A.; Córdoba, M.d.G.; Ruiz-Moyano, S. Improvement of Shelf-life of Cherry (Prunus avium L.) by Combined Application of Modified-atmosphere Packaging and Antagonistic Yeast for Long-distance Export. J. Sci. Food Agric. 2023. [Google Scholar] [CrossRef] [PubMed]
- Valverde, J.M.; Giménez, M.J.; Guillén, F.; Valero, D.; Martínez-Romero, D.; Serrano, M. Methyl Salicylate Treatments of Sweet Cherry Trees Increase Antioxidant Systems in Fruit at Harvest and during Storage. Postharvest Biol. Technol. 2015, 109, 106–113. [Google Scholar] [CrossRef]
- Pan, L.; Chen, X.; Xu, W.; Fan, S.; Wan, T.; Zhang, J.; Cai, Y. Methyl Jasmonate Induces Postharvest Disease Resistance to Decay Caused by Alternaria Alternata in Sweet Cherry Fruit. Sci. Hortic. 2022, 292, 110624. [Google Scholar] [CrossRef]
- Giménez, M.J.; Valverde, J.M.; Valero, D.; Guillén, F.; Martínez-Romero, D.; Serrano, M.; Castillo, S. Quality and Antioxidant Properties on Sweet Cherries as Affected by Preharvest Salicylic and Acetylsalicylic Acids Treatments. Food Chem. 2014, 160, 226–232. [Google Scholar] [CrossRef]
- Wang, L.; Jin, P.; Wang, J.; Jiang, L.; Shan, T.; Zheng, Y. Effect of β-Aminobutyric Acid on Cell Wall Modification and Senescence in Sweet Cherry during Storage at 20 °C. Food Chem. 2015, 175, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Valero, D.; Serrano, M. Postharvest Biology and Technology for Preserving Fruit Quality; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9781439802670. [Google Scholar]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in Edible Plants Identified by Radioimmunoassay and by High Performance Liquid Chromatography-Mass Spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A New Plant Hormone and/or a Plant Master Regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ren, J.; Huang, X.; Zheng, X.; Tian, Y.; Shi, L.; Dong, P.; Li, Z. Melatonin: Biosynthesis, Content, and Function in Horticultural Plants and Potential Application. Sci. Hortic. 2021, 288, 110392. [Google Scholar] [CrossRef]
- Hu, W.; Kong, H.; Guo, Y.; Zhang, Y.; Ding, Z.; Tie, W.; Yan, Y.; Huang, Q.; Peng, M.; Shi, H.; et al. Comparative Physiological and Transcriptomic Analyses Reveal the Actions of Melatonin in the Delay of Postharvest Physiological Deterioration of Cassava. Front. Plant Sci. 2016, 7, 736. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Z.K.; Chai, H.K.; Cheng, N.; Yang, Y.; Wang, D.N.; Yang, T.; Cao, W. Melatonin Treatment Delays Postharvest Senescence and Regulates Reactive Oxygen Species Metabolism in Peach Fruit. Postharvest Biol. Technol. 2016, 118, 103–110. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of Melatonin Treatment on the Postharvest Quality of Strawberry Fruit. Postharvest Biol. Technol. 2018, 139, 47–55. [Google Scholar] [CrossRef]
- Miranda, S.; Vilches, P.; Suazo, M.; Pavez, L.; García, K.; Méndez, M.A.; González, M.; Meisel, L.A.; Defilippi, B.G.; del Pozo, T. Melatonin Triggers Metabolic and Gene Expression Changes Leading to Improved Quality Traits of Two Sweet Cherry Cultivars during Cold Storage. Food Chem. 2020, 319, 126360. [Google Scholar] [CrossRef]
- Xu, T.; Chen, Y.; Kang, H. Melatonin Is a Potential Target for Improving Post-Harvest Preservation of Fruits and Vegetables. Front. Plant Sci. 2019, 10, 1388. [Google Scholar] [CrossRef]
- Quero-García, J.; Schuster, M.; López-Ortega, G.; Charlot, G. Sweet Cherry Varieties and Improvement. In Cherries: Botany, Production and Uses; CABI, Ed.; CABI: Wallingford, UK, 2017; Volume 4, pp. 60–94. [Google Scholar]
- Carrión-Antolí, A.; Lorente-Mento, J.M.; Valverde, J.M.; Castillo, S.; Valero, D.; Serrano, M. Effects of Melatonin Treatment on Sweet Cherry Tree Yield and Fruit Quality. Agronomy 2022, 12, 3. [Google Scholar] [CrossRef]
- Saltveit, M.E. Measuring Respiration. Available online: https://ucanr.edu/datastoreFiles/234-20.pdf (accessed on 16 February 2023).
- Serradilla, M.J.; Falagán, N.; Bohmer, B.; Terry, L.A.; Alamar, M.C. The Role of Ethylene and 1-MCP in Early-Season Sweet Cherry ‘Burlat’ Storage Life. Sci. Hortic. 2019, 258, 108787. [Google Scholar] [CrossRef]
- Candan, A.P.; Raffo Teófilo, D.; Colodner, A. Pautas Para El Mantenimiento de La Calidad de Cerezas Frescas; Ediciones, I.N.T.A., Ed.; Estación Experimental Agropecuaria Alto Valle: Río Negro, Argentina, 2017; ISBN 978-987-521-852-9. [Google Scholar]
- UNE-EN ISO 8589:2010 Análisis sensorial. Guía general para el diseño de salas de cata. In Análisis Sensorial, 2nd ed.; AENOR: Madrid, Spain, 2010; pp. 53–74.
- Serradilla, M.J.; Lozano, M.; Bernalte, M.J.; Ayuso, M.C.; López-Corrales, M.; González-Gómez, D. Physicochemical and Bioactive Properties Evolution during Ripening of ‘Ambrunés’ Sweet Cherry Cultivar. LWT-Food Sci. Technol. 2011, 44, 199–205. [Google Scholar] [CrossRef]
- Manzano Durán, R.; Sánchez, J.E.F.; Velardo-Micharet, B.; Gómez, M.J.R. Multivariate Optimization of Ultrasound-Assisted Extraction for the Determination of Phenolic Compounds in Plums (Prunus Salicina Lindl.) by High-Performance Liquid Chromatography (HPLC). Instrum. Sci. Technol. 2020, 48, 113–127. [Google Scholar] [CrossRef]
- Serrano, M.; Díaz-Mula, H.M.; Zapata, P.J.; Castillo, S.; Guillén, F.; Martínez-Romero, D.; Valverde, J.M.; Valero, D. Maturity Stage at Harvest Determines the Fruit Quality and Antioxidant Potential after Storage of Sweet Cherry Cultivars. J. Agric. Food Chem. 2009, 57, 3240–3246. [Google Scholar] [CrossRef] [PubMed]
- Carrión-Antolí, A.; Martínez-Romero, D.; Guillén, F.; Zapata, P.J.; Serrano, M.; Valero, D. Melatonin Pre-Harvest Treatments Leads to Maintenance of Sweet Cherry Quality During Storage by Increasing Antioxidant Systems. Front. Plant Sci. 2022, 13, 863467. [Google Scholar] [CrossRef] [PubMed]
- Bal, E.; Torçuk, A.İ.; Özer, C. Influence of Melatonin Treatments on Fruit Quality and Storage Life of Sweet Cherry Cv. ‘Sweetheart.’ Erwerbs-Obstbau 2022, 64, 127–133. [Google Scholar] [CrossRef]
- Xia, H.; Shen, Y.; Shen, T.; Wang, X.; Zhang, X.; Hu, P.; Liang, D.; Lin, L.; Deng, H.; Wang, J.; et al. Melatonin Accumulation in Sweet Cherry and Its Influence on Fruit Quality and Antioxidant Properties. Molecules 2020, 25, 753. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant Growth Regulator and/or Biostimulator during Stress? Trends Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and Its Relationship to Plant Hormones. Ann. Bot. 2018, 121, 195–207. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Growth Activity, Rooting Capacity, and Tropism: Three Auxinic Precepts Fulfilled by Melatonin. Acta Physiol. Plant 2017, 39, 127. [Google Scholar] [CrossRef]
- Jaime, P.; Salvador, M.L.; Oria, R. Respiration Rate of Sweet Cherries: ‘Burlat’, ‘Sunburst’ and ‘Sweetheart’ Cultivars. J. Food Sci. 2001, 66, 43–47. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, X.; Long, L.E. The Effect of Postharvest Calcium Application in Hydro-Cooling Water on Tissue Calcium Content, Biochemical Changes, and Quality Attributes of Sweet Cherry Fruit. Food Chem. 2014, 160, 22–30. [Google Scholar] [CrossRef]
- Fan, S.; Xiong, T.; Lei, Q.; Tan, Q.; Cai, J.; Song, Z.; Yang, M.; Chen, W.; Li, X.; Zhu, X. Melatonin Treatment Improves Postharvest Preservation and Resistance of Guava Fruit (Psidium Guajava L.). Foods 2022, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Njie, A.; Zhang, W.; Dong, X.; Lu, C.; Pan, X.; Liu, Q. Effect of Melatonin on Fruit Quality via Decay Inhibition and Enhancement of Antioxidative Enzyme Activities and Genes Expression of Two Mango Cultivars during Cold Storage. Foods 2022, 11, 3209. [Google Scholar] [CrossRef]
- Lin, X.; Huang, S.; Huber, D.J.; Zhang, Q.; Wan, X.; Peng, J.; Luo, D.; Dong, X.; Zhu, S. Melatonin Treatment Affects Wax Composition and Maintains Storage Quality in ‘Kongxin’ Plum (Prunus Salicina L. Cv) during Postharvest. Foods 2022, 11, 3972. [Google Scholar] [CrossRef]
- Crisosto, C.H.; Crisosto, G.M.; Metheney, P. Consumer Acceptance of ‘Brooks’ and ‘Bing’ Cherries Is Mainly Dependent on Fruit SSC and Visual Skin Color. Postharvest Biol. Technol. 2003, 28, 159–167. [Google Scholar] [CrossRef]
- Tijero, V.; Muñoz, P.; Munné-Bosch, S. Melatonin as an Inhibitor of Sweet Cherries Ripening in Orchard Trees. Plant Physiol. Biochem. 2019, 140, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Pasquariello, M.S.; di Patre, D.; Mastrobuoni, F.; Zampella, L.; Scortichini, M.; Petriccione, M. Influence of Postharvest Chitosan Treatment on Enzymatic Browning and Antioxidant Enzyme Activity in Sweet Cherry Fruit. Postharvest Biol. Technol. 2015, 109, 45–56. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L* Skin | a*/b* Skin | L* Flesh | a*/b* Flesh | TSS (%) | TA (%) | |
|---|---|---|---|---|---|---|
| Storage time | ||||||
| 0 | 28.24 ± 0.11 b1 | 3.92 ± 0.09 | 42.16 ± 0.71 | 1.83 ± 0.03 b | 15.73 ± 0.19 a | 0.88 ± 0.01 a |
| 14 | 28.69 ± 0.17 b | 3.77 ± 0.06 | 42.23 ± 1.03 | 1.84 ± 0.06 b | 15.83 ± 0.23 a | 0.61 ± 0.04 b |
| 21 | 29.98 ± 0.12 a | 3.72 ± 0.05 | 40.16 ± 0.50 | 2.03 ± 0.02 a | 14.94 ± 0.15 b | 0.63 ± 0.01 b |
| Treatment | ||||||
| CO | 28.77 ± 0.31 | 3.85 ± 0.77 | 40.29 ± 0.91 | 1.98 ± 0.05 | 15.61 ± 0.29 | 0.73 ± 0.04 |
| M0.1 | 28.96 ± 0.33 | 3.82 ± 0.09 | 41.74 ± 0.86 | 1.89 ± 0.05 | 15.54 ± 0.88 | 0.72 ± 0.05 |
| M0.3 | 29.16 ± 0.30 | 3.77 ± 0.09 | 42.30 ± 0.74 | 1.85 ± 0.04 | 15.32 ± 0.64 | 0.71 ± 0.04 |
| M0.5 | 29.00 ± 0.27 | 3.77 ± 0.09 | 41.74 ± 1.18 | 1.87 ± 0.08 | 15.52 ± 0.78 | 0.66 ± 0.06 |
| p date 2 | *** | ns | ns | *** | * | *** |
| p treatment | ns | ns | ns | ns | ns | ns |
| p date * treatment | ns | ns | ns | * | ns | ns |
| Sensory Attributes | 0 Days | 14 Days | 21 Days | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CO | CO | M0.1 | M0.3 | M0.5 | CO | M0.1 | M0.3 | M0.5 | |
| Colour uniformity | 8.6 | 7.5 b* | 7.7 ab | 8.5 a | 8.5 a | 8.8 | 8.2 | 9.1 | 9.2 |
| Skin colour | 7.2 | 7.0 | 6.6 | 7.6 | 7.6 | 7.4 ab | 7.2 b | 8.4 a | 7.8 ab |
| Sourness flavour | 5.2 | 3.9 b | 4.8 ab | 4.0 b | 5.0 a | 3.7 | 3.8 | 3.9 | 4.3 |
| Sweetness flavour | 4.6 | 5.6 a | 4.1 b | 4.7 ab | 4.5 ab | 3.8 | 4.2 | 4.0 | 4.1 |
| Consumer Satisfaction | 0 Days | 14 Days | 21 Days | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CO | CO | M0.1 | M0.3 | M0.5 | CO | M0.1 | M0.3 | M0.5 | |
| I quite dislike it | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8.3 | 0 |
| I dislike it slightly | 18.2 | 16.7 | 8.3 | 8.3 | 0 | 8.3 | 8.3 | 8.3 | 16.7 |
| I neither like it nor dislike it | 0 | 16.7 | 16.7 | 8.3 | 25 | 25 | 50 | 16.7 | 8.3 |
| I like it slightly | 18.2 | 25 | 58.3 | 50 | 50 | 50 | 16.7 | 50 | 41.7 |
| I quite like it | 36.3 | 16.7 | 16.7 | 16.7 | 0 | 16.7 | 16.7 | 8.3 | 8.3 |
| I like it very much | 18.2 | 25 | 0 | 16.7 | 25 | 0 | 8.3 | 8.3 | 8.3 |
| Think it’s wonderful | 9.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Purchase intention | 72.7 | 75 | 83.3 | 91.7 | 91.7 | 65 | 50 | 66.7 | 75 |
| Polyphenols | Storage Time | Treatment | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 14 | 21 | p # | CO | M0.1 | M0.3 | M0.5 | p ## | p ### | ||
| Anthocyanins | C3G | 2.23 ± 0.05 a1 | 1.04 ± 0.25 b | 1.03 ± 0.29 b | ns | 1.03 ± 0.32 | 0.98 ± 0.36 | 0.88 ± 0.96 | 1.06 ± 0.33 | ns | ns |
| C3R | 37.67 ± 3.78 a | 26.26 ± 3.47 b | 26.05 ± 5.97 b | ns | 23.21 ± 4.68 b | 29.53 ± 8.32 a | 23.62 ± 2.78 ab | 24.49 ± 4.68 ab | * | *** | |
| P3G | 1.83 ± 0.22 | 1.35 ± 0.23 | 1.35 ± 0.44 | ns | 1.35 ± 0.36 | 1.40 ± 0.22 | 1.38 ± 0.32 | 1.26 ± 0.15 | ns | ns | |
| P3R | 0.10 ± 0.17 | 0.19 ± 0.02 | 0.35 ± 0.51 | ns | 0.19 ± 0.01 | 0.19 ± 0.03 | 0.49 ± 0.72 | 0.20 ± 0.02 | ns | ns | |
| SAN | 41.84 ± 3.75 a | 28.99 ± 4.42 b | 28.63 ± 6.58 b | ns | 25.78 ± 5.20 b | 32.21 ± 6.88 a | 27.20 ± 2.05 ab | 30.03 ± 1.68 ab | * | *** | |
| Hydroxycinna mic | NEO | 32.80 ± 3.11 a | 27.40 ± 2.39 b | 26.44 ± 3.53 b | ns | 24.87 ± 2.99 b | 25.69 ± 1.84 b | 26.71 ± 1.61 ab | 30.40 ± 2.17 a | *** | * |
| PCQ | 9.39 ± 1.68 | 9.32 ± 1.46 | 9.37 ± 1.38 | ns | 8.31 ± 1.05 | 9.44 ± 1.26 | 9.33 ± 0.97 | 10.32 ± 1.68 | ns | ns | |
| CHL | 2.48 ± 0.35 a | 2.20 ± 0.27 a | 1.80 ± 0.23 b | **** | 1.87 ± 0.42 b | 1.93 ± 0.27 b | 2.06 ± 0.40 a | 2.13 ± 0.15 a | ns | * | |
| PCO | 0.00 ± 0.00 | 0.07 ± 0.07 | 0.03 ± 0.07 | ns | 0.04 ± 0.05 | 0.07 ± 0.08 | 0.03 ± 0.05 | 0.07 ± 0.08 | ns | + | |
| SHA | 44.66 ± 4.89 a | 38.99 ± 3.73 b | 37.65 ± 4.88 b | ns | 35.08 ± 4.37 b | 37.14 ± 2.58 ab | 38.13 ± 2.32 ab | 42.92 ± 3.80 a | ** | * | |
| Flavonols | Q3R | 2.92 ± 0.35 a | 1.98 ± 0.35 b | 1.99 ± 0.20 b | ns | 2.02 ± 0.16 | 1.93 ± 0.37 | 1.95 ± 0.18 | 1.72 ± 0.27 | + | * |
| K3R | 0.58 ± 0.12 a | 0.28 ± 0.12 b | 0.28 ± 0.07 b | ns | 0.22 ± 0.05 b | 0.46 ± 0.13 b | 0.49 ± 0.06 a | 0.45 ± 0.08 b | + | + | |
| I3R | 0.60 ± 0.13 | 0.47 ± 0.06 | 0.77 ± 1.06 | ns | 1.10 ± 1.48 | 0.46 ± 0.10 | 0.47 ± 0.06 | 0.42 ± 0.06 | ns | ns | |
| SFL | 4.18 ± 0.64 a | 2.49 ± 0.38 b | 3.07 ± 1.12 ab | ns | 3.16 ± 1.65 a | 2.95 ± 0.40 a | 2.79 ± 0.19 a | 2.66 ± 0.34 b | ns | ns | |
| Flavan-3-ols | PB1 | 1.71 ± 0.15 a | 1.39 ± 0.14 b | 1.38 ± 0.14 b | ns | 1.29 ± 0.10 | 1.49 ± 0.07 | 1.32 ± 0.17 | 1.45 ± 0.07 | ns | ns |
| PB2 | 0.88 ± 0.18 a | 0.48 ± 0.09 b | 0.49 ± 0.15 b | ns | 0.45 ± 0.08 b | 0.41 ± 0.11 b | 0.45 ± 0.07 b | 0.63 ± 0.06 a | * | ns | |
| CAT | 0.85 ± 0.16 a | 0.59 ± 0.08 b | 0.62 ± 0.12 b | ns | 0.55 ± 0.04 b | 0.59 ± 0.07 ab | 0.59 ± 0.09 ab | 0.65 ± 0.14 a | ns | ns | |
| EPI | 4.29 ± 1.35 a | 2.97 ± 0.54 b | 3.24 ± 0.85 b | ns | 2.64 ± 0.30 b | 2.84 ± 0.50 ab | 3.00 ± 0.32 ab | 3.97 ± 0.78 a | ** | ns | |
| SFO | 7.73 ± 1.77 a | 5.44 ± 0.70 b | 5.75 ± 1.16 b | ns | 4.93 ± 0.38 b | 5.32 ± 0.68 ab | 5.36 ± 0.57 ab | 6.75 ± 0.96 a | ** | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortés-Montaña, D.; Bernalte-García, M.J.; Velardo-Micharet, B.; Serrano, M.; Serradilla, M.J. Impact of Pre-Storage Melatonin Application on the Standard, Sensory, and Bioactive Quality of Early Sweet Cherry. Foods 2023, 12, 1723. https://doi.org/10.3390/foods12081723
Cortés-Montaña D, Bernalte-García MJ, Velardo-Micharet B, Serrano M, Serradilla MJ. Impact of Pre-Storage Melatonin Application on the Standard, Sensory, and Bioactive Quality of Early Sweet Cherry. Foods. 2023; 12(8):1723. https://doi.org/10.3390/foods12081723
Chicago/Turabian StyleCortés-Montaña, Daniel, María Josefa Bernalte-García, Belén Velardo-Micharet, María Serrano, and Manuel Joaquín Serradilla. 2023. "Impact of Pre-Storage Melatonin Application on the Standard, Sensory, and Bioactive Quality of Early Sweet Cherry" Foods 12, no. 8: 1723. https://doi.org/10.3390/foods12081723