
Nutritional and Feeding Adaptability of Clanis bilineata tsingtauica Larvae to Different Cultivars of Soybean, (Glycine max)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soybean Cultivars
2.2. Insect Stocks
2.3. Measurement of Nutrients in Soybeans
2.4. Analysis of Plant VOCs from Soybeans
2.5. Host Selection Assays of C. bilineata tsingtauica on Soybean
2.6. Measurement of Nutrients in C. bilineata tsingtauica
2.7. Statistical Analysis
3. Results
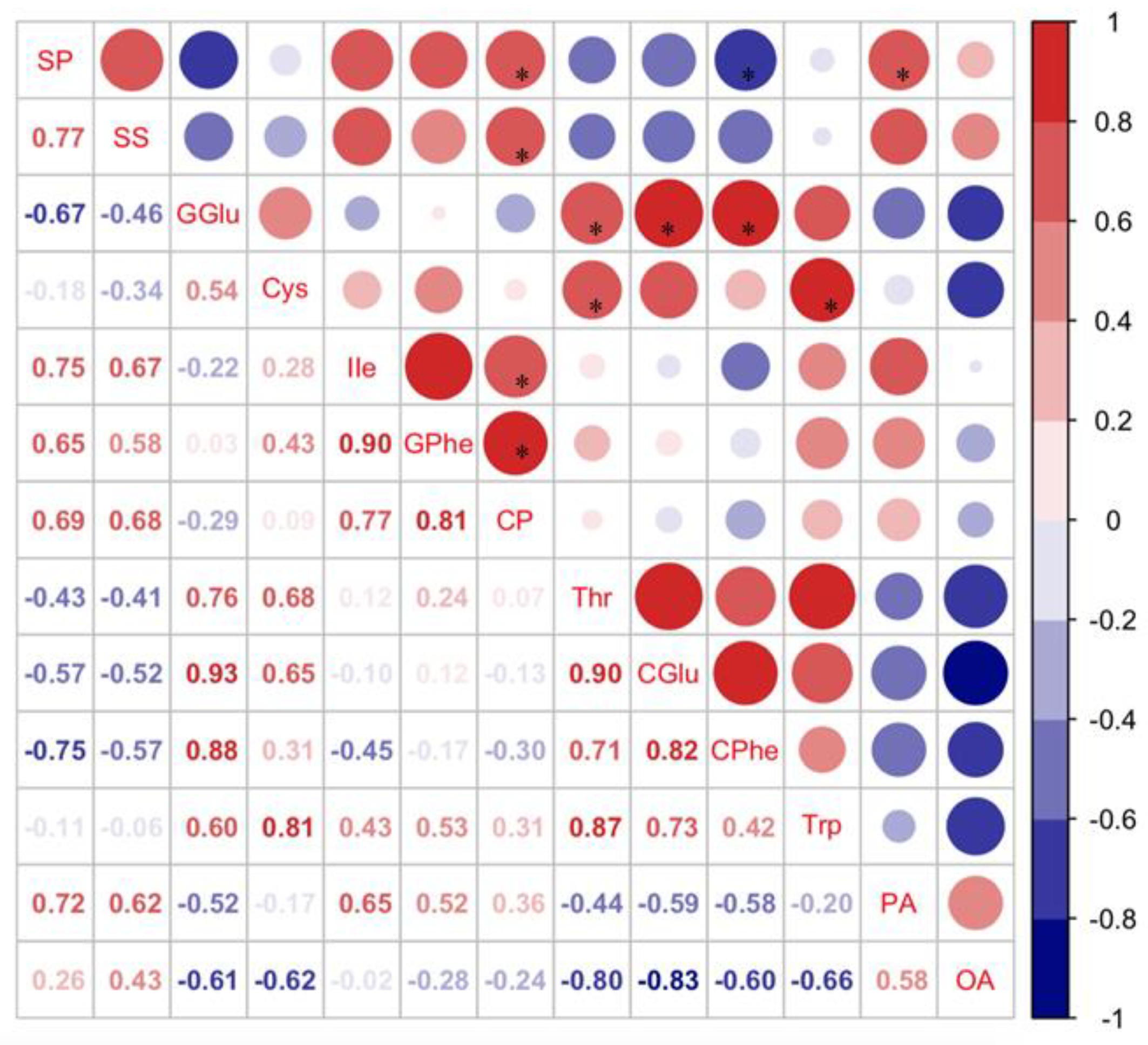
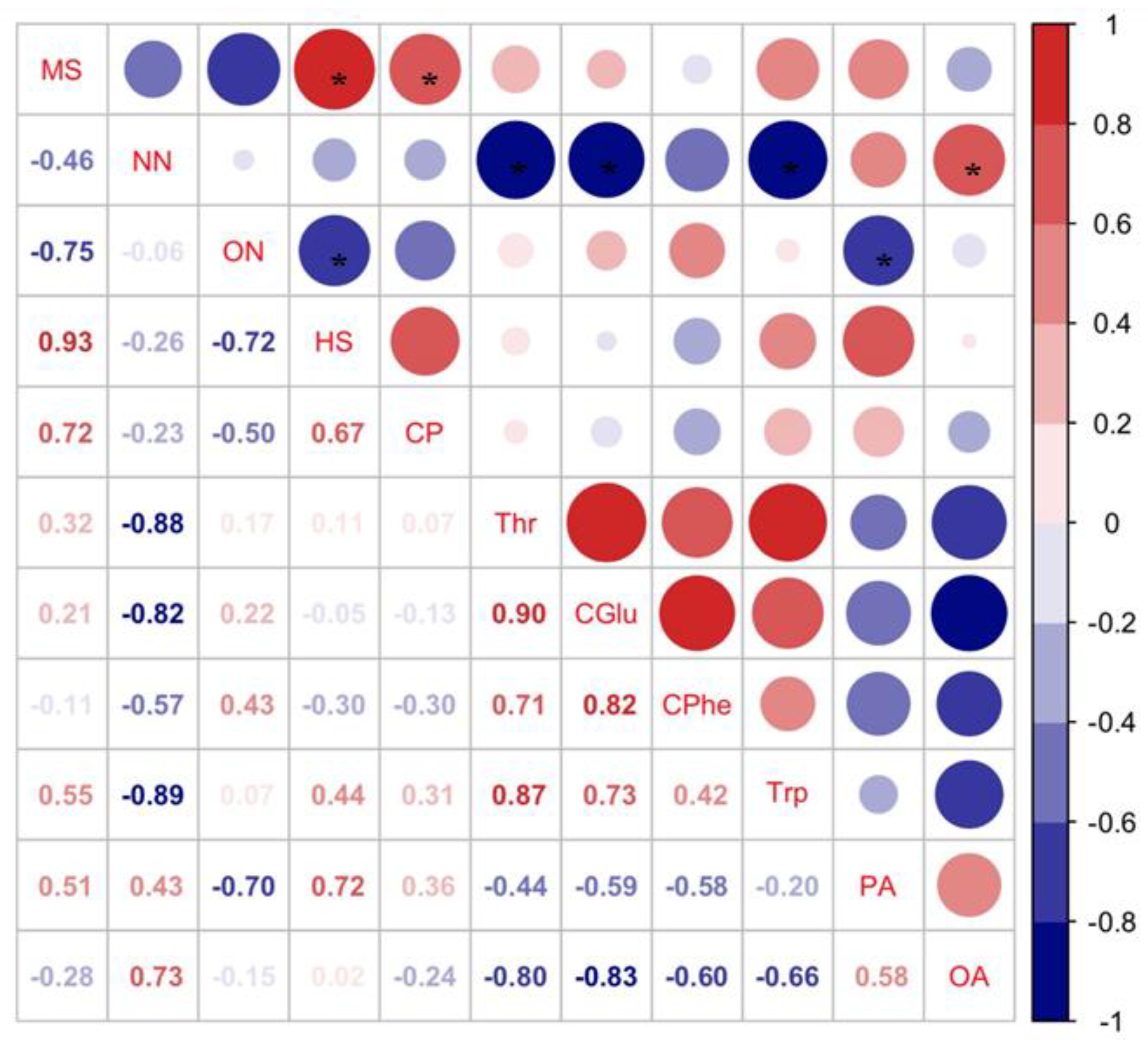
3.1. Effect of Nutrients in Different Soybean Cultivars on Host Selection and Nutrient Levels of C. bilineata tsingtauica
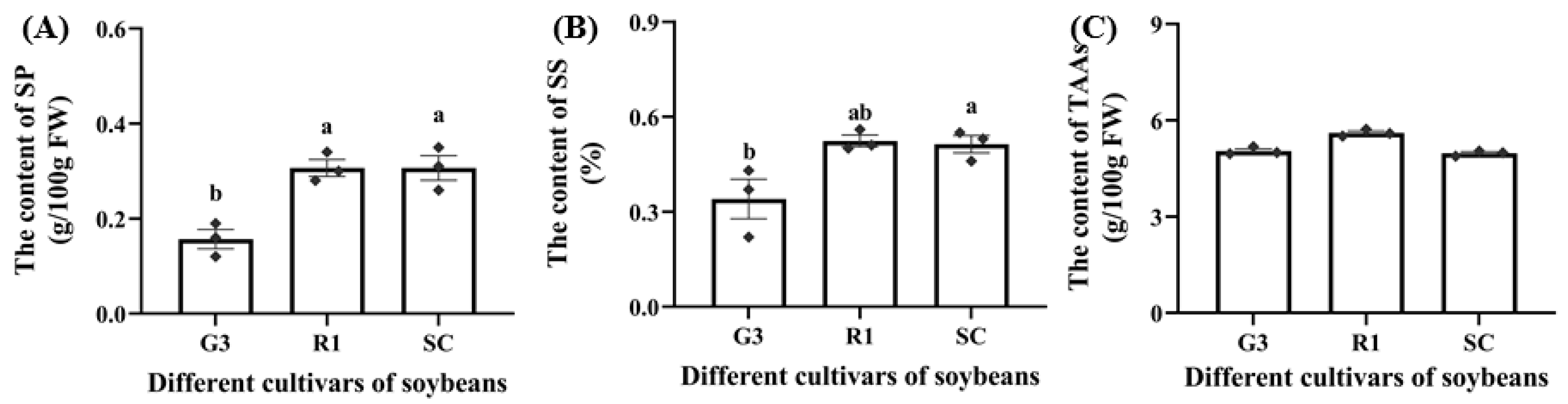
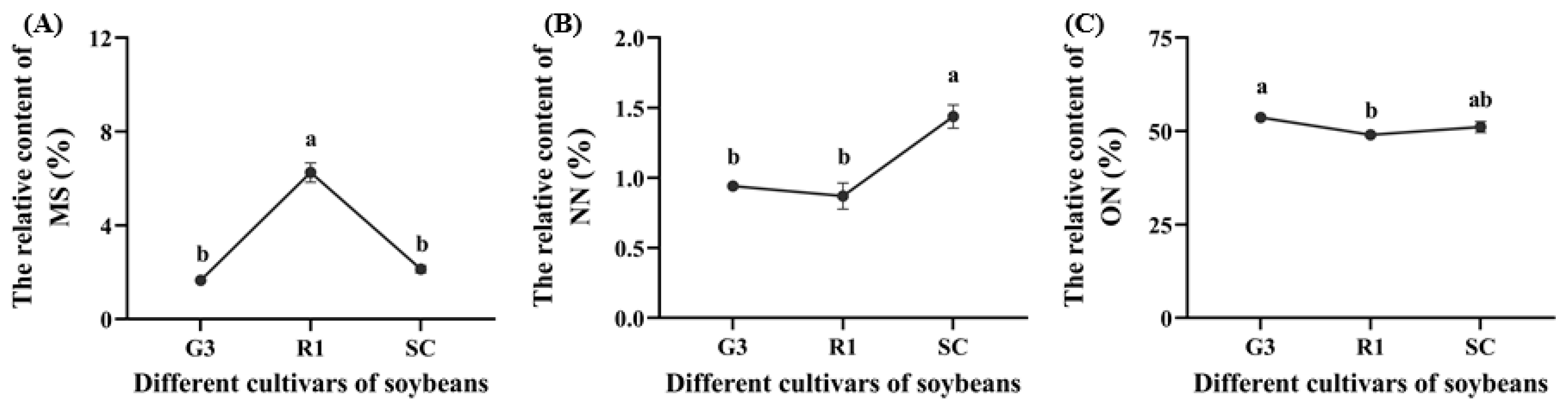
3.1.1. Nutrient Levels in Three Soybean Cultivars
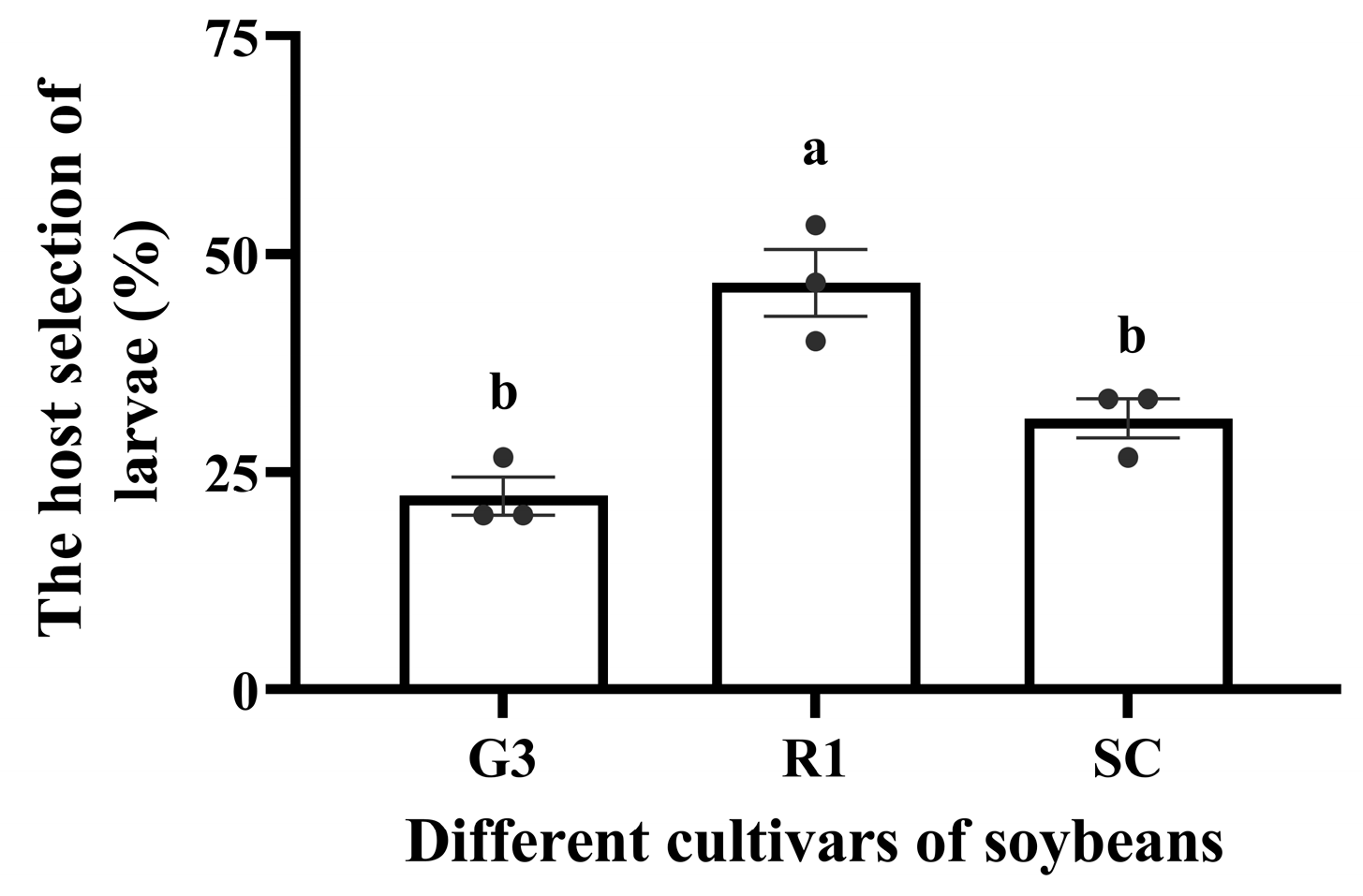
3.1.2. Effect of Nutrients in Different Soybean Cultivars on Host Selection of C. bilineata tsingtauica
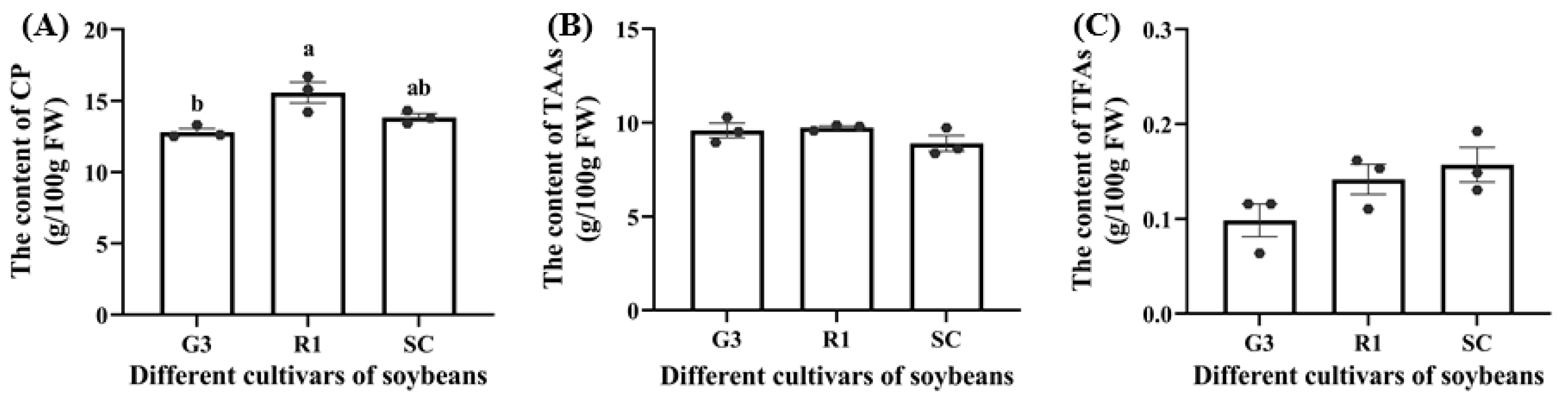
3.1.3. Effect of Nutrients in Different Soybean Cultivars on the Nutrient Content of C. bilineata tsingtauica
3.2. Effect of Plant VOCs in Different Soybean Cultivars on Host Selection and Nutrients in C. bilineata tsingtauica
3.2.1. Plant VOCs in the Three Soybean Cultivars
3.2.2. Effect of VOCs in Different Cultivars of Soybeans on Host Selection and Nutrients of C. bilineata tsingtauica Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Meyer-Rochow, V.B. Can insects help to ease the problem of world food shortages? Search 1975, 6, 261–262. [Google Scholar]
- Feng, Y.; Chen, X.M.; Zhao, M.; He, Z.; Sun, L.; Wang, C.Y.; Ding, W.F. Edible insects in China: Utilization and prospects. Insect Sci. 2018, 25, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Verneau, F.; Amato, M.; Barbera, F.L. Edible insects and global food security. Insects 2021, 12, 472. [Google Scholar] [CrossRef] [PubMed]
- Hadi, J.; Brightwell, G. Safety of alternative proteins: Technological, environmental and regulatory aspects of cultured meat, plant-based meat, insect protein and single-cell protein. Foods 2021, 10, 1226. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; Jung, C. Insects used as food and feed: Isn’t that what we all need? Foods 2020, 9, 1003. [Google Scholar] [CrossRef]
- Hermans, W.J.H.; Senden, J.M.; Churchward, V.T.A.; Paulussen, K.J.M.; Fuchs, C.J.; Smeets, J.S.J.; van Loon, J.J.A.; Verd, L.B.; van Loon, L.J.C. Insects are a viable protein source for human consumption: From insect protein digestion to postprandial muscle protein synthesis in vivo in humans: A double-blind randomized trial. Am. J. Clin. Nutr. 2021, 114, 934–944. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; Gahukar, R.T.; Ghosh, S.; Jung, C. Chemical composition, nutrient quality and acceptability of edible insects are affected by species, developmental stage, gender, diet, and processing method. Foods 2021, 10, 1036. [Google Scholar] [CrossRef]
- Zhu, H.F.; Wang, L.Y. Fauna Sinica: Insecta Vol.11. Lepidoptera: Sphingidae; Science Press: Beijing, China, 1997; p. 234. [Google Scholar]
- Tian, H.; Zhang, Y.M. The Nutritional Components Analysis and Evaluation of Clanis bilineata tsingtauica Mell. Acta Nutr. Sin. 2012, 34, 289–291. [Google Scholar]
- Qian, L.; Deng, P.; Chen, F.J.; Cao, Y.; Sun, H.W.; Liao, H.J. The exploration and utilization of functional substances in edible insects: A review. Food Prod. Process. Nutr. 2022, 4, 11. [Google Scholar] [CrossRef]
- Guo, M.M.; Li, X.F.; Deng, P.; Li, D.W.; Li, J.L.; Fan, J.W.; Chen, F. Diapause termination and post-diapause of overwintering Clanis bilineata tsingtauica larvae. Chin. J. Appl. Entomol. 2021, 58, 966–972. [Google Scholar]
- Wetzel, W.C.; Kharouba, H.M.; Robinson, M.; Holyoak, M.; Karban, R. Variability in plant nutrients reduces insect herbivore performance. Nature 2016, 539, 425–427. [Google Scholar] [CrossRef] [PubMed]
- Franzke, A.; Unsicker, S.B.; Specht, J.; Khler, G.; Weisser, W.W. Being a generalist herbivore in a diverse world: How do diets from different grasslands influence food plant selection and fitness of the grasshopper Chorthippus parallelus? Ecol. Entomol. 2010, 35, 126–138. [Google Scholar] [CrossRef]
- García-Robledo, C.; Horvitz, C.C. Experimental demography and the vital rates of generalist and specialist insect herbivores on native and novel host plants. J. Anim. Ecol. 2011, 80, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.Y.; Wang, X.Q. Studies on the biological and nutritional effects of various host leaves on the bollworm. Plant Prot. 2009, 35, 74–76. [Google Scholar]
- Wang, L.; Qu, L.; Hu, J.; Zhang, L.; Tang, F.; Lu, M. Metabolomics reveals constitutive metabolites that contribute resistance to fall webworm (Hyphantria cunea) in Populus deltoids. Environ. Exp. Bot. 2017, 136, 31–40. [Google Scholar] [CrossRef]
- Feng, H.; Edwards, N.; Anderson, C.M.H.; Althaus, M.; Duncan, R.P.; Hsu, Y.C.; Luetje, C.W.; Price, D.R.G.; Wilson, A.C.C.; Thwaites, D.T. Trading amino acids at the aphid-Buchnera symbiotic interface. Proc. Natl. Acad. Sci. USA 2019, 116, 16003–16011. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Zhao, Y.; Wang, L.; Zhu, X.; Zhang, K.; Li, D.; Ji, J.; Niu, L.; Cui, J.; Luo, J.; et al. Regulation of amino acid metabolism in Aphis gossypii parasitized by Binodoxys communis. Front. Nutr. 2022, 9, 1006253. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, M.F.; Jiang, S.L.; Zhang, Y.F.; Parajulee, M.N. Host-selection behavior and physiological mechanisms of the cotton aphid, Aphis gossypii, in response to rising atmospheric carbon dioxide levels. J. Insect Physiol. 2018, 109, 149–156. [Google Scholar] [CrossRef]
- Deans, C.A.; Behmer, S.T.; Fiene, J.; Sword, G.A. Spatio-temporal, genotypic, and environmental effects on plant soluble protein and digestible carbohydrate content: Implications for insect herbivores with cotton as an exemplar. J. Chem. Ecol. 2016, 42, 1151–1163. [Google Scholar] [CrossRef]
- Su, Y.; Lu, M.X.; Jing, L.Q.; Qian, L.; Zhao, M.; Du, Y.Z.; Liao, H.J. Nutritional properties of larval epidermis and meat of the edible insect Clanis bilineata tsingtauica (Lepidoptera: Sphingidae). Foods 2021, 10, 2895. [Google Scholar] [CrossRef]
- Futuyma, D.J.; Agrawal, A.A. Macroevolution and the biological diversity of plants and herbivores. Proc. Natl. Acad. Sci. USA 2009, 106, 18054–18061. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.P.; Wang, J.H.; Yu, Z.J.; Huang, F.Q.; Zhang, Z. Host selection behavior of Micromelalopha sieversi to five populus deltoides clones. For. Res. 2014, 27, 459–565. [Google Scholar]
- Chen, Y.; Ulyshen, M.D.; Poland, T.M. Abundance of volatile organic compounds in white ash phloem and emerald ash borer larval frass does not attract Tetrastichus planipennisi in a Y tube olfactometer. Insect Sci. 2016, 23, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Zhang, Y.; Fan, D.S.; Feng, J.N. Identification and expression profiling of odorant-binding proteins and chemosensory proteins of Daktulosphaira vitifoliae (Hemiptera: Phylloxeridae). J. Econ. Entomol. 2017, 110, 1812–1820. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Bueno, S.J.C. Within-plant signaling by volatiles leads to induction and priming of an indirect plant defense in nature. Proc. Natl. Acad. Sci. USA 2007, 104, 5467–5472. [Google Scholar] [CrossRef] [PubMed]
- Kalske, A.; Shiojiri, K.; Uesugi, A.; Sakata, Y.; Morrell, K.; Kessler, A. Insect herbivory selects for volatile-mediated plant-plant communication. Curr. Biol. 2019, 29, 3128–3133.e3. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Nakata, R.; Aboshi, T.; Yoshinaga, N.; Teraishi, M.; Okumoto, Y.; Ishihara, A.; Morisaka, H.; Huffaker, A.; Schmelz, E.A.; et al. Insect-induced daidzein, formononetin and their conjugates in soybean leaves. Metabolites 2014, 4, 532–546. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Pan, L.; Ren, L.; Chen, F.; Feng, Y.; Luo, Y. Antifeedant activity of Ginkgo biloba secondary metabolites against Hyphanria cunea larvae: Mechanisms and applications. PLoS ONE 2016, 11, e155682. [Google Scholar] [CrossRef]
- Qian, L.; Huang, Z.J.; Liu, X.W.; Wei, Y.J.; Yin, Y.; Gui, F.R.; Chen, F.J. Elevated CO2-mediated plant VOCs change aggravates invasive thrips occurrence by altering their host-selection behaviour. J. Appl. Entomol. 2021, 145, 777–788. [Google Scholar] [CrossRef]
- Lu, W.; Hou, M.L.; Li, J.W. Effects of plant nutrition on the behavior and development of herbivorous insects. In Proceedings of the Annual Meeting of Chinese Plant Protection Society, Yunnan, China, 8 November 2006. [Google Scholar]
- Gou, Y.; Quandahor, P.; Zhang, Y.; Coulter, J.A.; Liu, C. Host plant nutrient contents influence nutrient contents in Bradysia cellarum and Bradysia impatiens. PLoS ONE 2020, 15, e0226471. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, N.A.; Jung, C. Effect of non-essential amino acids (proline and glutamic acid) and sugar polyol (sorbitol) on brood of honey bees. Front. Ecol. Evol. 2022, 10, 1009670. [Google Scholar] [CrossRef]
- Moreno, P.J.M.; Ganguly, A. Determination of fatty acid content in some edible insects of Mexico. J. Insects Food Feed 2016, 2, 37–42. [Google Scholar] [CrossRef]
- Barroso, F.G.; Sánchez-Muros, M.J.; Segura, M.; Morote, E.; Torres, A.; Ramos, R.; Guil, J.L. Insects as food: Enrichment of larvae of Hermetia illucens with omega 3 fatty acids by means of dietary modifications. J. Food Compos. Anal. 2017, 62, 8–13. [Google Scholar] [CrossRef]
- Tata, A.; Massaro, A.; Marzoli, F.; Miano, B.; Bragolusi, M.; Piro, R.; Belluco, S. Authentication of edible insects’ powders by the combination of DART-HRMS signatures: The first application of ambient mass spectrometry to screening of novel food. Foods 2022, 11, 2264. [Google Scholar] [CrossRef] [PubMed]
- Paiko, Y.B.; Jacob, J.O.; Salihu, S.O.; Dauda, B.E.N.; Suleiman, M.A.T.; Akanya, H.O. Fatty acid and amino acid profile of emperor moth caterpillar (Cirina forda) in Paikoro local government area of Niger state, Nigeria. Am. J. Biochem. 2014, 4, 29–34. [Google Scholar]
- Hopper, K.R.; Wittmeyer, K.T.; Kuhn, K.L.; Lanier, K. Response to selection for parasitism of a suboptimal, low-preference host in an aphid parasitoid. Evol. Appl. 2021, 14, 2012–2024. [Google Scholar] [CrossRef]
- Cardona, J.B.; Grover, S.; Busta, L.; Sattler, S.E.; Louis, J. Sorghum cuticular waxes influence host plant selection by aphids. Planta 2022, 257, 22. [Google Scholar] [CrossRef]
- Giertych, M.J.; Bakowski, M.; Karolewski, P.; Zytkowiak, R.; Grzebyta, J. Influence of mineral fertilization on food quality of oak leaves and utilization efficiency of food components by the gypsy moth. Entomol. Exp. Appl. 2005, 117, 59–69. [Google Scholar] [CrossRef]
- Conrad-Rooney, E.; Plotkin, A.B.; Pasquarella, V.J.; Elkinton, J.; Chandler, J.L.; Matthes, J.H. Defoliation severity is positively related to soil solution nitrogen availability and negatively related to soil nitrogen concentrations following a multi-year invasive insect irruption. AoB Plants 2020, 12, plaa059. [Google Scholar] [CrossRef]
- Dai, X.H.; You, M.S.; Fu, L.J. Effects of nitrogen, phosphorus and potassium via leaf on host-selection by Liriomyza sativae. Acta Entomol. Sin. 2002, 1, 145–147. [Google Scholar]
- Tarkowski, Ł.P.; Signorelli, S.; Höfte, M. γ-Aminobutyric acid and related amino acids in plant immune responses: Emerging mechanisms of action. Plant Cell Environ. 2020, 43, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, M.; Fukatsu, T. Host’s demand for essential amino acids is compensated by an extracellular bacterial symbiont in a hemipteran insect model. Front. Physiol. 2022, 13, 1028409. [Google Scholar] [CrossRef] [PubMed]
- van Dam, N.M.; Qiu, B.L.; Hordijk, C.A.; Vet, L.E.; Jansen, J.J. Identification of biologically relevant compounds in aboveground and belowground induced volatile blends. J. Chem. Ecol. 2010, 36, 1006–1016. [Google Scholar] [CrossRef] [PubMed]
- Tasin, M.; Bäckman, A.C.; Anfora, G.; Carlin, S.; Ioriatti, C.; Witzgall, P. Attraction of female grapevine moth to common and specific olfactory cues from 2 host plants. Chem. Senses 2010, 35, 57–64. [Google Scholar] [CrossRef] [PubMed]
- McCallum, E.J.; Cunningham, J.P.; Lücker, J.; Zalucki, M.P.; De Voss, J.J.; Botella, J.R. Increased plant volatile production affects oviposition, but not larval development, in the moth Helicoverpa armigera. J. Exp. Biol. 2011, 214, 3672–3677. [Google Scholar] [CrossRef]
- Randlkofer, B.; Obermaier, E.; Meiners, T. Mother’s choice of the oviposition site: Balancing risk of egg parasitism and need of food supply for the progeny with an infochemical shelter? Chemoecology 2007, 17, 177–186. [Google Scholar] [CrossRef]
- Huang, D.; Sun, M.; Han, M.; Zhang, Z.; Miao, Y.; Zhang, J.; Yao, Y. Volatile organic compounds (VOCs) regulate the spatial distribution of Lepidoptera insects in an orchard ecosystem. Biol. Control 2020, 149, 104311. [Google Scholar] [CrossRef]
- Wilson, J.K.; Ruiz, L.; Duarte, J.; Davidowitz, G. The nutritional landscape of host plants for a specialist insect herbivore. Ecol. Evol. 2019, 9, 13104–13113. [Google Scholar] [CrossRef]
- Zhou, S.; Jander, G. Molecular ecology of plant volatiles in interactions with insect herbivores. J. Exp. Bot. 2022, 73, 449–462. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids | Soybean Varieties | ||
|---|---|---|---|
| Guandou-3 (G3) | Ruidou-1 (R1) | September Cold (SC) | |
| Aspartic acid (Asp) | 0.607 ± 0.007 | 0.727 ± 0.003 | 0.617 ± 0.003 |
| Threonine (Thr) | 0.247 ± 0.007 | 0.287 ± 0.003 | 0.253 ± 0.007 |
| Serine (Ser) | 0.243 ± 0.009 | 0.300 ± 0.000 | 0.243 ± 0.0120 |
| Glutamic acid (Glu) | 0.747 ± 0.023 a | 0.655 ± 0.040 ab | 0.580 ± 0.010 b |
| Glycine (Gly) | 0.153 ± 0.003 | 0.187 ± 0.000 | 0.163 ± 0.003 |
| Alanine (Ala) | 0.193 ± 0.003 | 0.233 ± 0.008 | 0.205 ± 0.015 |
| Cystine (Cys) | 0.054 ± 0.002 ab | 0.055 ± 0.002 a | 0.045 ± 0.003 b |
| Valine (Val) | 0.200 ± 0.010 | 0.223 ± 0.003 | 0.193 ± 0.007 |
| Methionine (Met) | ND | ND | ND |
| Isoleucine (IIe) | 0.103 ± 0.004 b | 0.118 ± 0.003 a | 0.110 ± 0.000 ab |
| Leucine (Leu) | 0.340 ± 0.015 | 0.400 ± 0.011 | 0.365 ± 0.005 |
| Tyrosine (Tyr) | 0.263 ± 0.009 | 0.320 ± 0.006 | 0.263 ± 0.007 |
| Phenylalanine (Phe) | 0.127 ± 0.003 b | 0.168 ± 0.009 a | 0.135 ± 0.005 ab |
| Lysine (Lys) | 0.397 ± 0.007 | 0.467 ± 0.003 | 0.400 ± 0.015 |
| Histidine (His) | 1.050 ± 0.006 | 1.023 ± 0.093 | 1.077 ± 0.044 |
| Arginine (Arg) | 0.193 ± 0.012 | 0.260 ± 0.006 | 0.200 ± 0.017 |
| Proline (Pro) | 0.127 ± 0.009 | 0.123 ± 0.012 | 0.120 ± 0.005 |
| Amino Acids | Larvae Treated with Three Soybean Varieties | ||
|---|---|---|---|
| Guandou-3 (G3) | Ruidou-1 (R1) | September Cold (SC) | |
| Aspartic acid (Asp) | 0.957 ± 0.032 | 0.950 ± 0.012 | 0.900 ± 0.070 |
| Threonine (Thr) | 0.457 ± 0.012 a | 0.435 ± 0.016 ab | 0.380 ± 0.000 b |
| Serine (Ser) | 0.533 ± 0.050 | 0.577 ± 0.050 | 0.493 ± 0.028 |
| Glutamic acid (Glu) | 1.450 ± 0.015 a | 1.328 ± 0.057 ab | 1.190 ± 0.040 b |
| Glycine (Gly) | 0.540 ± 0.038 | 0.560 ± 0.047 | 0.553 ± 0.029 |
| Alanine (Ala) | 0.630 ± 0.049 | 0.703 ± 0.042 | 0.647 ± 0.053 |
| Cystine (Cys) | 0.042 ± 0.0028 | 0.038 ± 0.002 | 0.031 ± 0.009 |
| Valine (Val) | 0.563 ± 0.037 | 0.587 ± 0.024 | 0.557 ± 0.043 |
| Methionine (Met) | 0.042 ± 0.004 | 0.043 ± 0.005 | 0.033 ± 0.004 |
| Isoleucine (IIe) | 0.367 ± 0.015 | 0.377 ± 0.003 | 0.367 ± 0.021 |
| Leucine (Leu) | 0.643 ± 0.024 | 0.673 ± 0.012 | 0.627 ± 0.040 |
| Tyrosine (Tyr) | 0.513 ± 0.006 | 0.543 ± 0.049 | 0.507 ± 0.064 |
| Phenylalanine (Phe) | 0.447 ± 0.018 a | 0.380 ± 0.017 ab | 0.345 ± 0.015 b |
| Lysine (Lys) | 0.405 ± 0.171 | 0.543 ± 0.049 | 0.507 ± 0.062 |
| Histidine (His) | 0.560 ± 0.031 | 0.580 ± 0.015 | 0.520 ± 0.018 |
| Arginine (Arg) | 0.607 ± 0.033 | 0.620 ± 0.011 | 0.617 ± 0.034 |
| Proline (Pro) | 0.493 ± 0.049 | 0.497 ± 0.012 | 0.440 ± 0.031 |
| Tryptophan (Trp) | 0.098 ± 0.005 a | 0.101 ± 0.003 a | 0.078 ± 0.001 b |
| Fatty Acids | Larvae Treated with Three Soybean Varieties | ||
|---|---|---|---|
| Guandou-3 (G3) | Ruidou-1 (R1) | September Cold (SC) | |
| Caprylic acid (C8:0) | ND | ND | ND |
| Decylic acid (C10:0) | ND | ND | ND |
| Undecanoic acid (C11:0) | ND | ND | ND |
| Lauric acid (C12:0) | ND | ND | ND |
| Tridecylic acid (C13:0) | ND | ND | ND |
| Myristic acid (C14:0) | ND | ND | ND |
| Myristoleic acid (C14:1) | ND | ND | ND |
| Pentadecanoic acid (C15:0) | ND | ND | ND |
| Pentadecenoic acid (C15:1) | ND | ND | ND |
| Palmitic acid (C16:0) | 0.037 ± 0.004 b | 0.072 ± 0.011 a | 0.074 ± 0.007 a |
| Palmitoleic acid (C16:1) | ND | ND | ND |
| Margaric acid (C17:0) | ND | ND | ND |
| Margaroleic acid (C17:1) | ND | ND | ND |
| Stearic acid (C18:0) | 0.034 ± 0.008 | 0.047 ± 0.004 | 0.047 ± 0.008 |
| Elaidic acid (C18:1n9t) | ND | ND | ND |
| Oleinic Acid (C18:1n9c) | 0.008 ± 0.001 b | 0.008 ± 0.002 b | 0.018 ± 0.003 a |
| Linolelaidic acid (C18:2n6t) | ND | ND | ND |
| Linoleic acid (C18:2n6c) | ND | ND | ND |
| Arachidic acid (C20:0) | 0.007 ± 0.003 | 0.005 ± 0.002 | 0.005 ± 0.002 |
| γ-Linolenic acid (C18:3n6) | ND | ND | ND |
| α-Linolenic acid (C18:3n3) | ND | ND | ND |
| cis-11-Eicosenoic acid (C20:1) | ND | ND | ND |
| Heneicosanoic acid (C21:0) | ND | ND | ND |
| cis-11,14-Eicosatrienoic acid (C20:2) | ND | ND | ND |
| Behenic acid (C22:0) | 0.012 ± 0.003 | 0.010 ± 0.000 | 0.009 ± 0.000 |
| cis-8,11,14-Eicosatrienoic acid (C20:3n6) | ND | ND | ND |
| cis-11,14,17-Eicosatrienoic acid (C20:3n3) | ND | ND | ND |
| Erucic acid (C22:1n9) | ND | ND | ND |
| Arachidonic acid (C20:4n6) | ND | ND | ND |
| Tricosanoic acid (C23:0) | ND | ND | ND |
| cis-13,16-Docosadienoic acid (C22:2) | ND | ND | ND |
| Eicosapentaenoic acid (C20:5n3) | ND | ND | ND |
| Lignoceric acid (C24:0) | ND | ND | ND |
| Nervonic acid (C24:1) | ND | ND | ND |
| cis-4,7,10,13,16,19-Docosahexaenoic Acid (C22:6n3) | ND | ND | ND |
| Classification | Name of Volatiles | CAS Number | Guandou-3 (G3) | Ruidou-1 (R1) | September Cold (SC) |
|---|---|---|---|---|---|
| Aldehydes | Benzaldehyde | 000100-52-7 | 0.0040 ± 0.0002 | 0.0035 ± 0.0003 | 0.0042 ± 0.0004 |
| Nonanal | 000124-19-6 | 0.0094 ± 0.0003 | 0.0087 ± 0.0009 | 0.0144 ± 0.0008 | |
| Heptadienal | 004313-03-5 | 0.0112 ± 0.0017 | ND | 0.0151 ± 0.0018 | |
| Salicylaldehyde | 000119-36-8 | ND | 0.0207 ± 0.0009 | 0.0297 ± 0.0056 | |
| 2,4-octadienal, (E,E)- | 030361-28-5 | ND | 0.0027 ± 0.0002 | ND | |
| Heptanal | 000111-71-7 | ND | ND | 0.0028 ± 0.0003 | |
| Esters | Methyl salicylate | 000119-36-8 | 0.0165 ± 0.0021 | 0.0625 ± 0.0041 | 0.0213 ± 0.0016 |
| Dihydroactinidiolide | 015356-74-8 | ND | 0.0065 ± 0.0003 | ND | |
| Alcohols | 3-octenol | 003391-86-4 | 0.5364 ± 0.0081 | 0.4902 ± 0.0044 | 0.5109 ± 0.0147 |
| 1-hexanol | 000111-27-3 | 0.0130 ± 0.0011 | ND | ND | |
| 1-penten-3-ol | 000616-25-1 | ND | 0.0104 ± 0.0008 | 0.0087 ± 0.0009 | |
| trans-2-pentene | 001576-87-0 | ND | 0.0048 ± 0.0011 | 0.0036 ± 0.0007 | |
| 2-penten-1-ol, (Z)- | 001576-95-0 | ND | ND | 0.0080 ± 0.0009 | |
| Ketones | 3,5-octadien-2-one | 038284-27-4 | ND | ND | 0.0077 ± 0.0020 |
| 5, 6-epoxy-β-ionone | 023267-57-4 | 0.0133 ± 0.0015 | 0.0176 ± 0.0005 | 0.0182 ± 0.0025 | |
| Heterocyclic compound | Furan, 2-ethyl- | 003208-16-0 | 0.0211 ± 0.0016 | 0.0166 ± 0.0004 | 0.0230 ± 0.0017 |
| Furan, 2-pentyl- | 003777-69-3 | 0.0063 ± 0.0009 | 0.0085 ± 0.0015 | 0.0090 ± 0.0013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, L.; Chen, B.-J.; Gui, F.-R.; Qin, Y.; Deng, P.; Liao, H.-J. Nutritional and Feeding Adaptability of Clanis bilineata tsingtauica Larvae to Different Cultivars of Soybean, (Glycine max). Foods 2023, 12, 1721. https://doi.org/10.3390/foods12081721
Qian L, Chen B-J, Gui F-R, Qin Y, Deng P, Liao H-J. Nutritional and Feeding Adaptability of Clanis bilineata tsingtauica Larvae to Different Cultivars of Soybean, (Glycine max). Foods. 2023; 12(8):1721. https://doi.org/10.3390/foods12081721
Chicago/Turabian StyleQian, Lei, Bo-Jian Chen, Fu-Rong Gui, Yi Qin, Pan Deng, and Huai-Jian Liao. 2023. "Nutritional and Feeding Adaptability of Clanis bilineata tsingtauica Larvae to Different Cultivars of Soybean, (Glycine max)" Foods 12, no. 8: 1721. https://doi.org/10.3390/foods12081721