
Microbial Community Variations and Bioconversion Improvements during Soybean-Based Fermentation by Kefir Grains


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sampling and Community Enrichment
2.3. Extraction of DNA and Sequencing
2.4. Genome Sequencing and Bioinformatics Analysis
2.5. Preparation of Soybean Kefir
2.6. Analysis of Free Amino Acids and Volatile Flavor Compounds in the Soybean Solution and Soybean Kefir
2.7. In Vitro Digestion Process of Soybean Solution and Soybean Kefir
2.8. Extraction, Separation, and Quantification of Isoflavones from Soybean Solution and Soybean Kefir
2.9. Statistical Analysis
3. Results
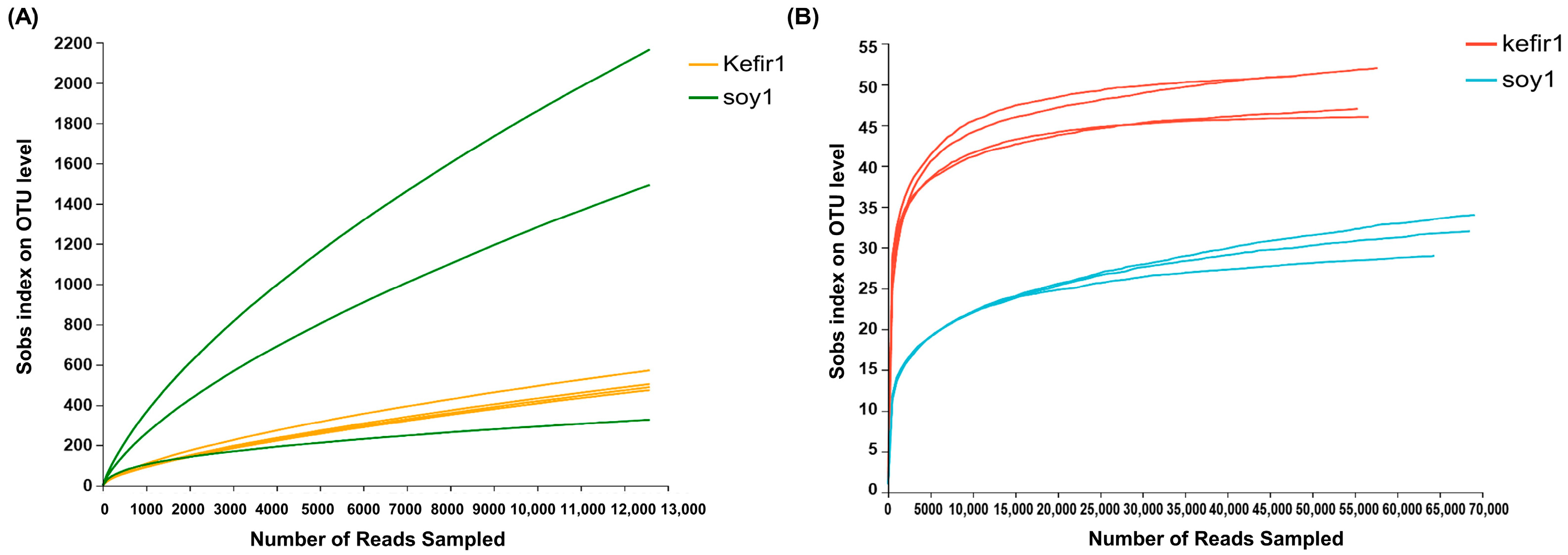
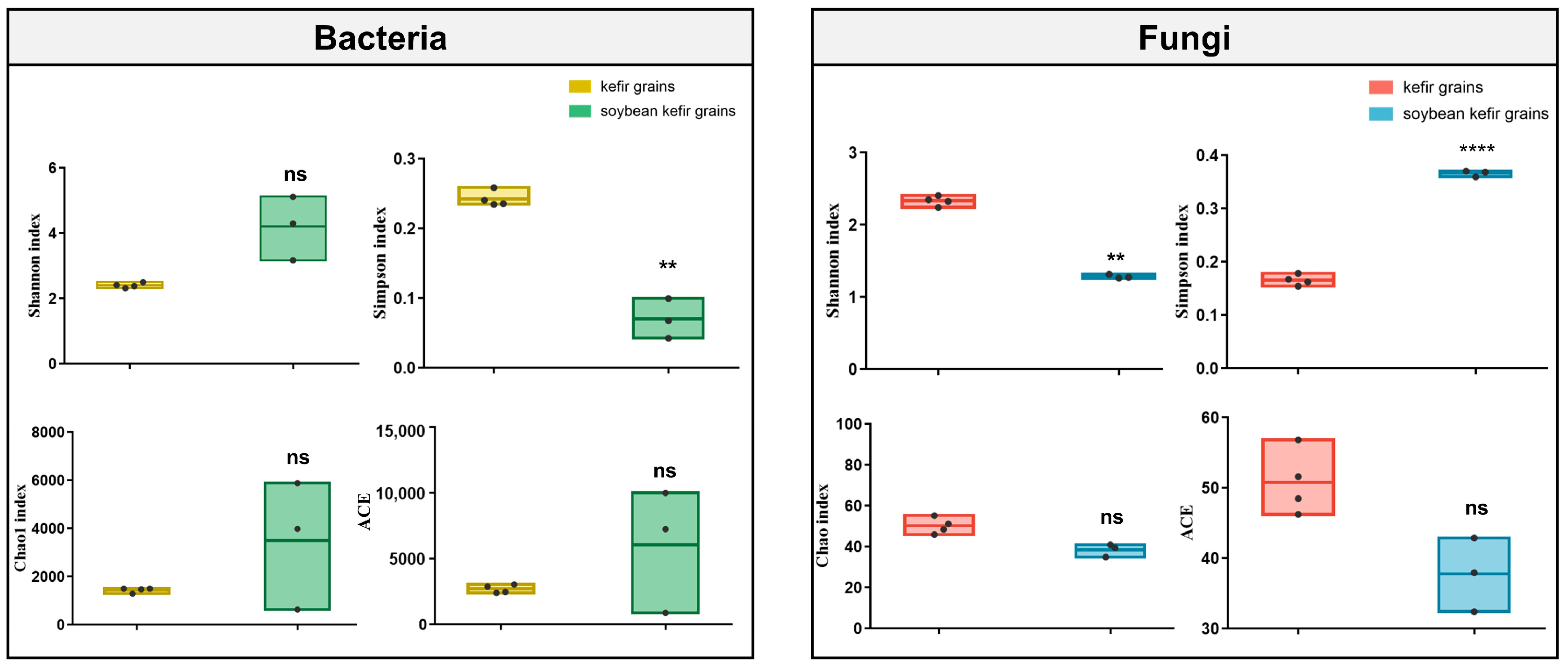
3.1. Sequencing Sample Size, Depth, and Alpha Diversity
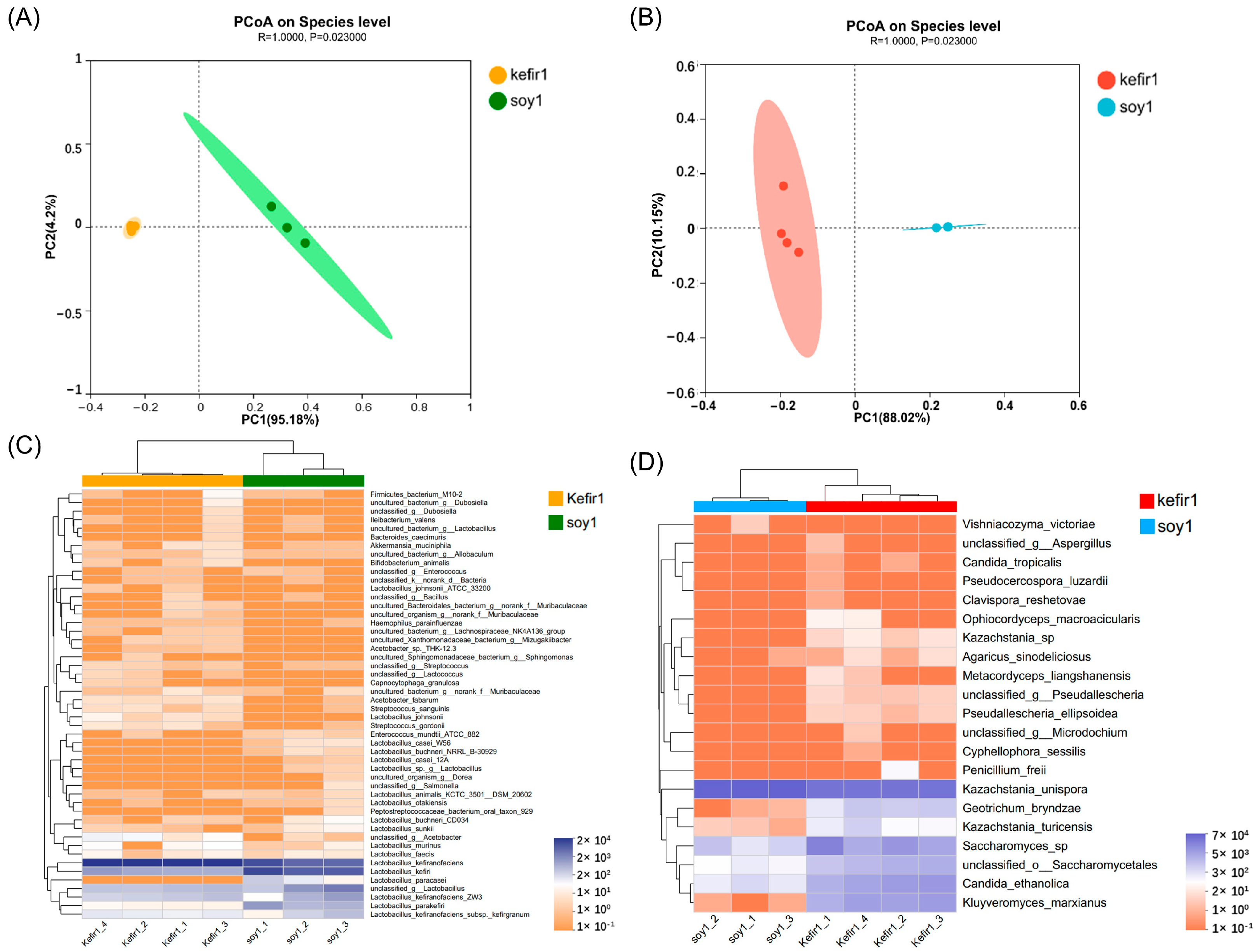
3.2. Discrimination of Similarity in Community Structure with Principal Coordinate Analysis (PCoA)
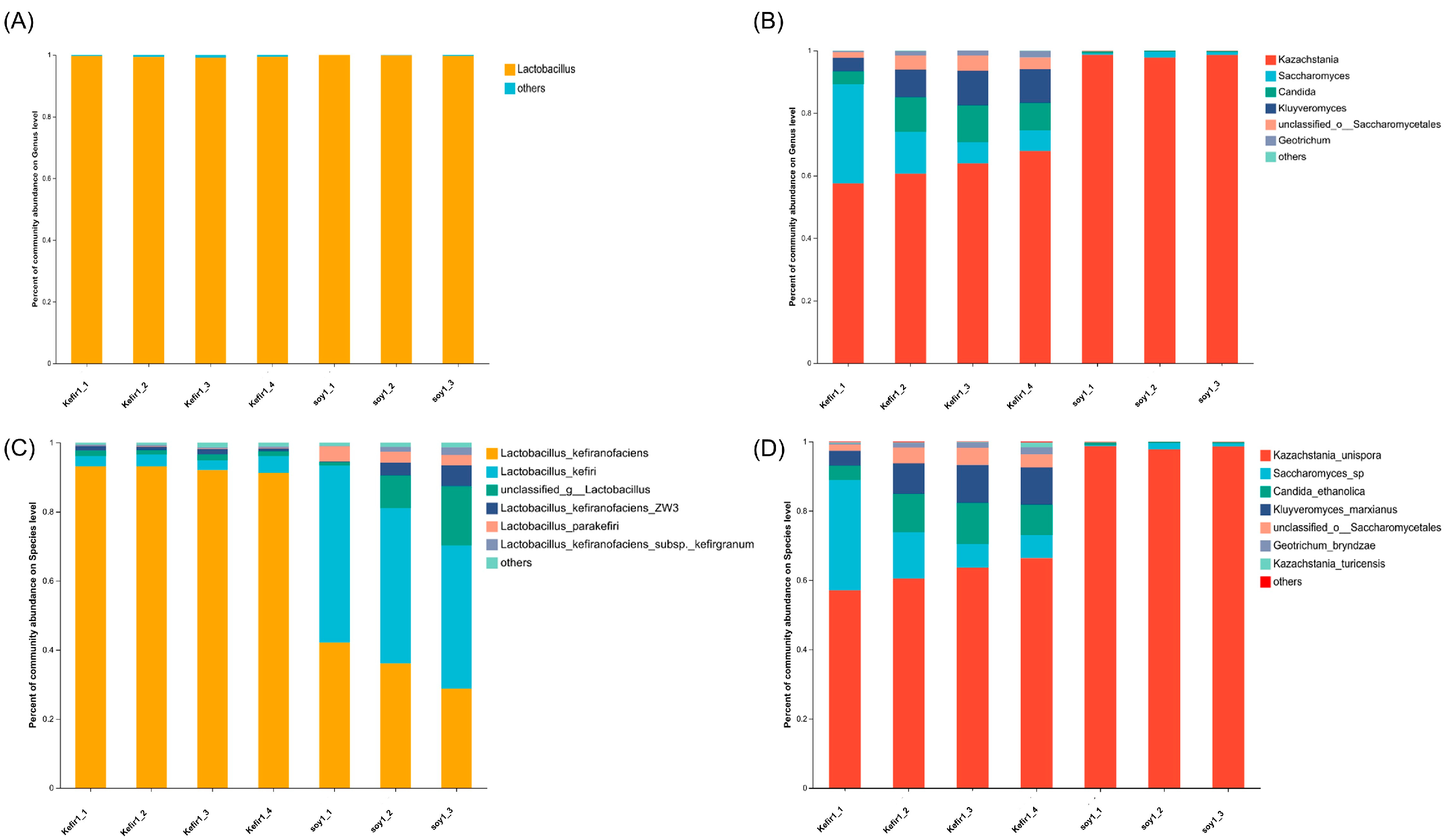
3.3. Analysis of Bacterial and Fungal Community Structure
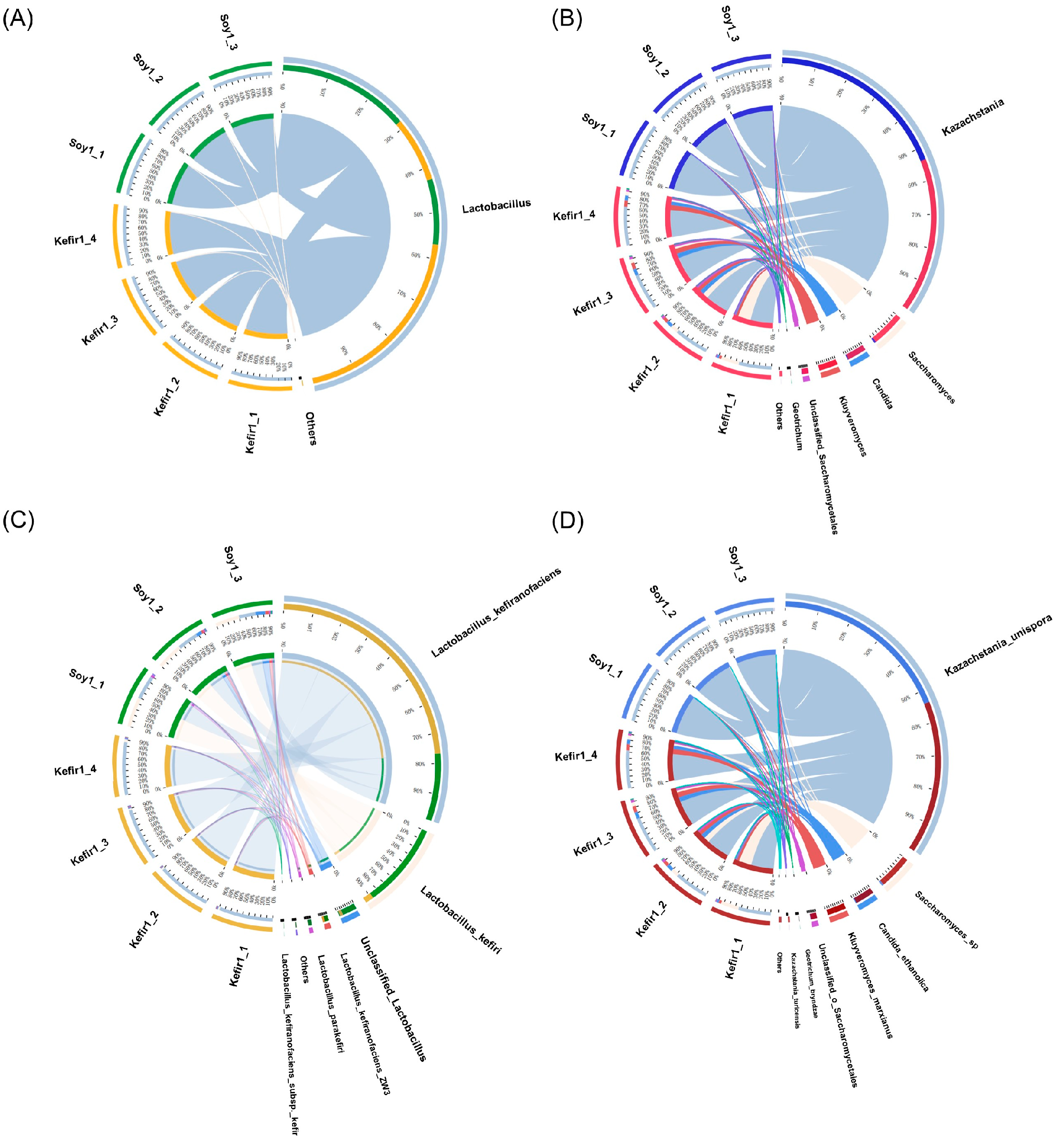
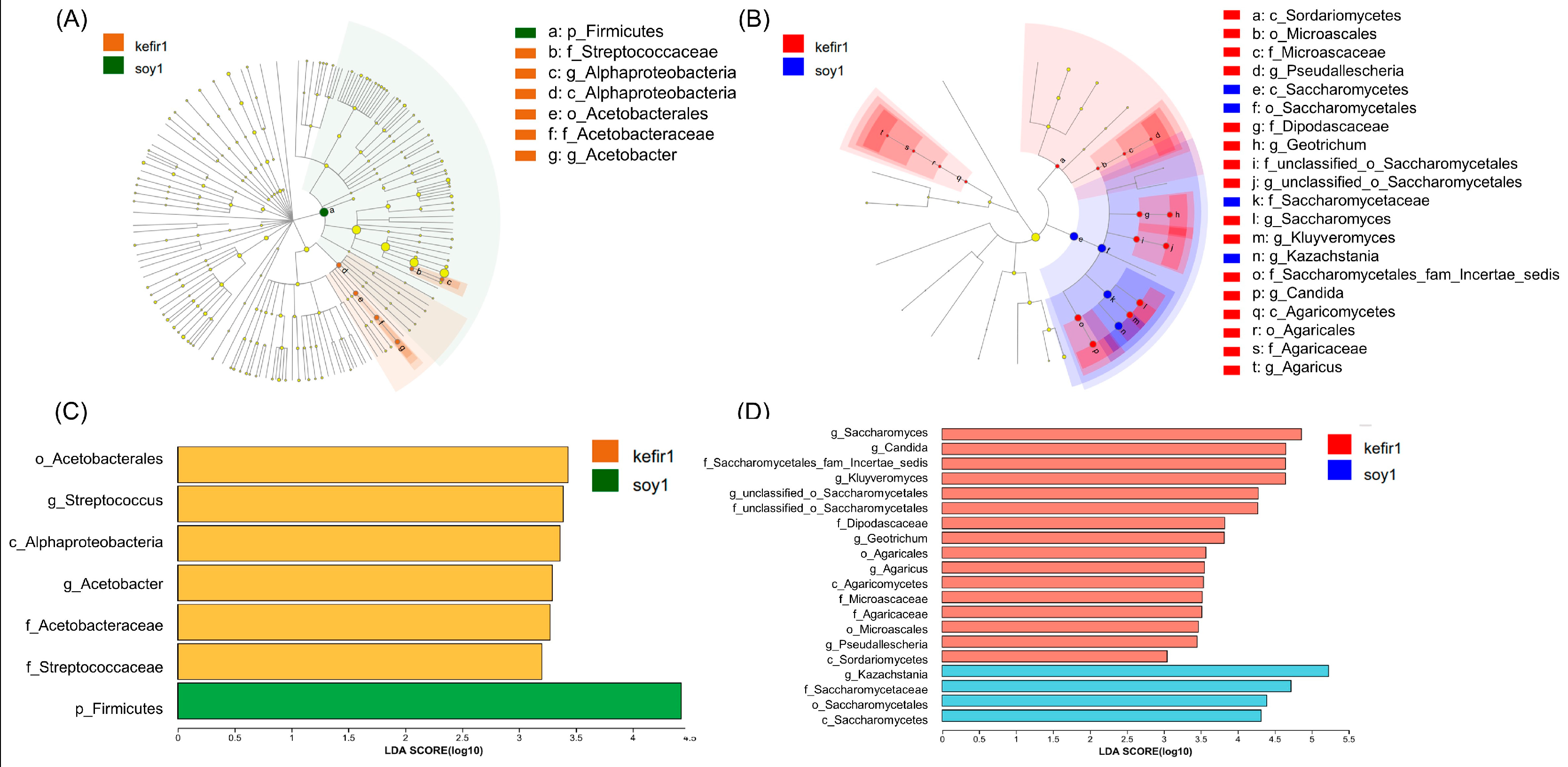
3.4. Analysis of Different Bacterial and Fungal Species in Samples
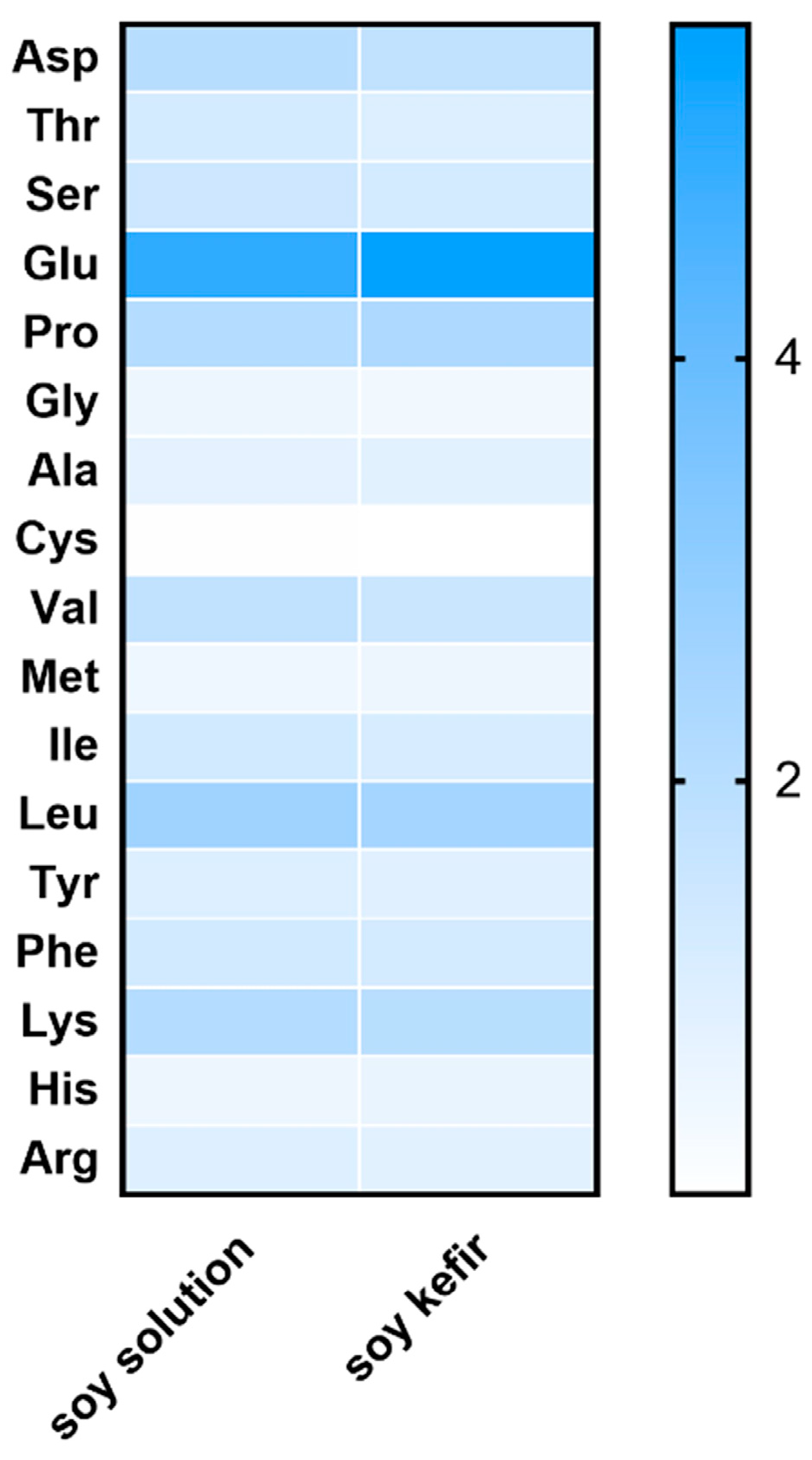
3.5. Analysis of Free Amino Acid Composition in Soybean Solution and Soybean Kefir
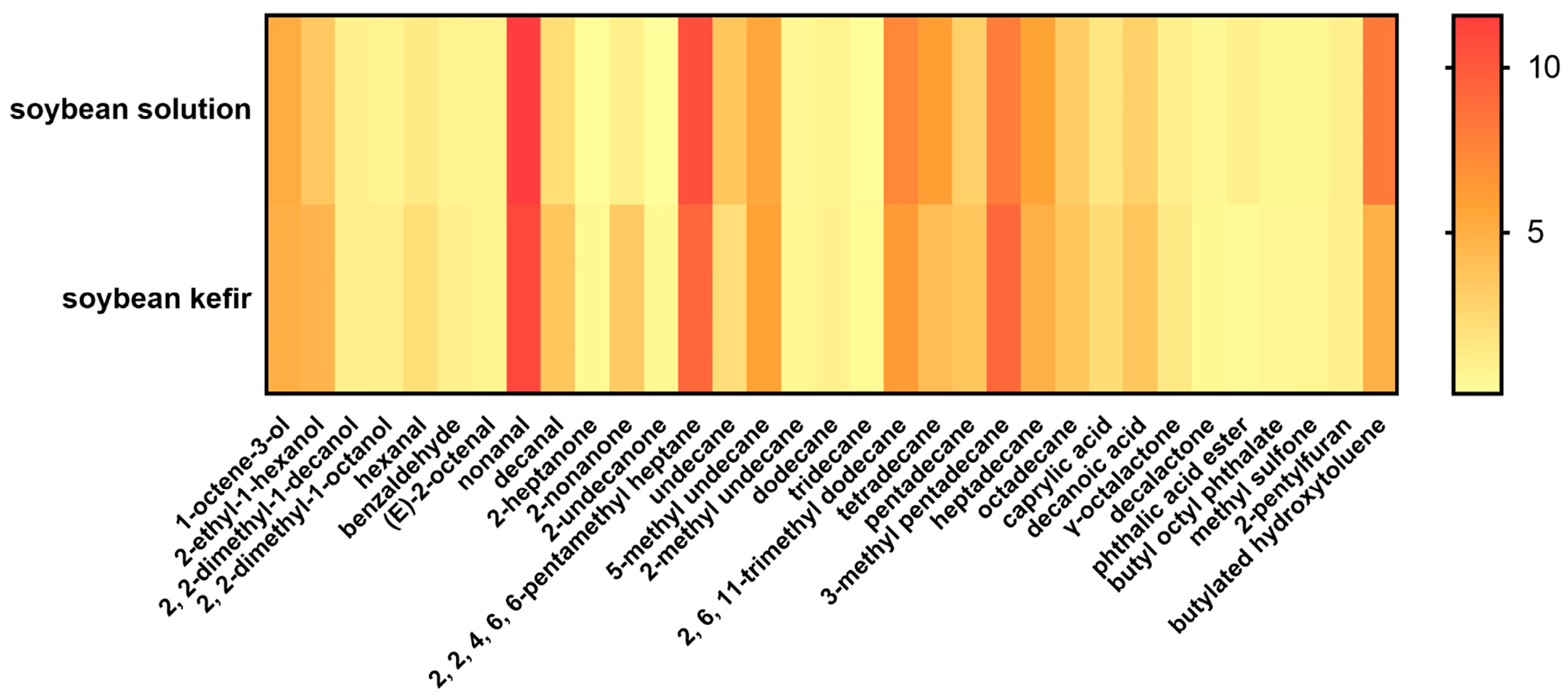
3.6. The Volatile Flavor Profile in Soybean Solution and Soybean Kefir
3.7. Bioconversion and Bioaccessibility of Soybean Isoflavones after Kefir Fermentation and In Vitro Digestion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nejati, F.; Junne, S.; Neubauer, P. A Big World in Small Grain: A Review of Natural Milk Kefir Starters. Microorganisms 2020, 8, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello, F.; Restuccia, D.; Spizzirri, U.G.; Carullo, G.; Leporini, M.; Loizzo, M.R. Improving Kefir Bioactive Properties by Functional Enrichment with Plant and Agro-Food Waste Extracts. Fermentation 2020, 6, 83. [Google Scholar] [CrossRef]
- Bourrie, B.C.T.; Richard, C.; Willing, B.P. Kefir in the Prevention and Treatment of Obesity and Metabolic Disorders. Curr. Nutr. Rep. 2020, 9, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Jeong, D.; Kim, H.; Seo, K.H. Modern perspectives on the health benefits of kefir in next generation sequencing era: Improvement of the host gut microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 1782–1793. [Google Scholar] [CrossRef] [PubMed]
- Rosa, D.D.; Dias, M.M.S.; Grzes’kowiak, L.M.; Reis, S.A.; Conceicao, L.L.; Peluzio, M.D.G. Milk kefir: Nutritional, microbiological and health benefits. Nutr. Res. Rev. 2017, 30, 82–96. [Google Scholar] [CrossRef]
- Gamba, R.R.; Yamamoto, S.; Abdel-Hamid, M.; Sasaki, T.; Michihata, T.; Koyanagi, T.; Enomoto, T. Chemical, Microbiological, and Functional Characterization of Kefir Produced from Cow’s Milk and Soy Milk. Int. J. Microbiol. 2020, 2020, 7019286. [Google Scholar] [CrossRef]
- Fernandes, M.D.; Lima, F.S.; Rodrigues, D.; Handa, C.; Guelfi, M.; Garcia, S.; Ida, E.I. Evaluation of the isoflavone and total phenolic contents of kefir-fermented soymilk storage and after the in vitro digestive system simulation. Food Chem. 2017, 229, 373–380. [Google Scholar] [CrossRef]
- Dong, X.; Xu, W.Q.; Sikes, R.A.; Wu, C.Q. Combination of low dose of genistein and daidzein has synergistic preventive effects on isogenic human prostate cancer cells when compared with individual soy isoflavone. Food Chem. 2013, 141, 1923–1933. [Google Scholar] [CrossRef]
- Hati, S.; Vij, S.; Singh, B.P.; Mandal, S. beta-Glucosidase activity and bioconversion of isoflavones during fermentation of soymilk. J. Sci. Food Agric. 2015, 95, 216–220. [Google Scholar] [CrossRef]
- Ningtyas, D.W.; Hati, S.; Prakash, S. Bioconversion and bioaccessibility of isoflavones from sogurt during in vitro digestion. Food Chem. 2021, 343, 128553. [Google Scholar] [CrossRef]
- Bau, T.R.; Garcia, S.; Ida, E.I. Changes in soymilk during fermentation with kefir culture: Oligosaccharides hydrolysis and isoflavone aglycone production. Int. J. Food Sci. Nutr. 2015, 66, 845–850. [Google Scholar] [CrossRef]
- Zhengqi, H.; Xiong, Z.; Xingyun, P.; Xiaodi, S.; Ruican, W.; Shuntang, G. Identification of the key off-flavor odorants for undesirable spoiled odor in thermally sterilized fermented soymilk. Food Res. Int. 2023, 164, 112407. [Google Scholar] [CrossRef]
- Kesika, P.; Sivamaruthi, B.S.; Chaiyasut, C. A review on the functional properties of fermented soymilk. Food Sci. Tech.-Braz. 2022, 42, e10721. [Google Scholar] [CrossRef]
- Blasche, S.; Kim, Y.; Mars, R.A.T.; Machado, D.; Maansson, M.; Kafkia, E.; Milanese, A.; Zeller, G.; Teusink, B.; Nielsen, J.; et al. Metabolic cooperation and spatiotemporal niche partitioning in a kefir microbial community. Nat. Microbiol. 2021, 6, 196. [Google Scholar] [CrossRef]
- Zhong, Z.; Hou, Q.; Kwok, L.; Yu, Z.; Zheng, Y.; Sun, Z.; Menghe, B.; Zhang, H. Bacterial microbiota compositions of naturally fermented milk are shaped by both geographic origin and sample type. J. Dairy Sci. 2016, 99, 7832–7841. [Google Scholar] [CrossRef] [Green Version]
- Long, N.; Liu, J.X.; Liu, J.L.; Li, Y.; Hou, Y.J.; Liao, X.F.; Zhou, L.D.; Shi, L.C.; Kong, W.J. Single-Molecule Real-Time Sequencing to Explore the Mycobiome Diversity in Malt. Microbiol. Spectr. 2022, 10, e0051122. [Google Scholar] [CrossRef]
- Rutkowska, J.; Antoniewska-Krzeska, A.; Zbikowska, A.; Cazon, P.; Vazquez, M. Volatile Composition and Sensory Profile of Lactose-Free Kefir, and Its Acceptability by Elderly Consumers. Molecules 2022, 27, 5386. [Google Scholar] [CrossRef]
- Simsek, S.; Sanchez-Rivera, L.; Nehir, E.; Karakaya, S.; Recio, I. Characterisation of in vitro gastrointestinal digests from low fat caprine kefir enriched with inulin. Int. Dairy J. 2017, 75, 68–74. [Google Scholar] [CrossRef]
- Ustun-Aytekin, O.; Seker, A.; Arisoy, S. The effect of in vitro gastrointestinal simulation on bioactivities of kefir. Int. J. Food Sci. Technol. 2020, 55, 283–292. [Google Scholar] [CrossRef]
- Salces, F.R.; Rostagno, M.A.; Amaya-Farfan, J. Novel process of hydration, followed by incubation and thermal processing, for high isoflavone bioconversion in soybeans. Food Res. Int. 2019, 121, 691–696. [Google Scholar] [CrossRef]
- Ma, D.; He, Q.W.; Ding, J.; Wang, H.Y.; Zhang, H.P.; Kwok, L.Y. Bacterial microbiota composition of fermented fruit and vegetable juices (jiaosu) analyzed by single-molecule, real-time (SMRT) sequencing. Cyta-J. Food 2018, 16, 950–956. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.L.; Yang, J.X.; Hou, Q.C.; Xu, H.Y.; Zheng, Y.; Zhang, H.P.; Zhang, L.B. Assessment of bacterial profiles in aged, home-made Sichuan paocai brine with varying titratable acidity by PacBio SMRT sequencing technology. Food Control 2017, 78, 14–23. [Google Scholar] [CrossRef]
- Yu, J.; Hou, Q.C.; Li, W.C.; Huang, W.Q.; Mo, L.X.; Yao, C.Q.; An, X.N.; Sun, Z.H.; Wei, H. Profiling of the viable bacterial and fungal microbiota in fermented feeds using single-molecule real-time sequencing. J. Anim. Sci. 2020, 98, skaa029. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.D.; Xu, J.G.; Ding, F.; Deng, W.Y.; Wang, X.; Xue, Y.S.; Chen, X.X.; Han, B.Z. Multidimensional profiling indicates the shifts and functionality of wheat-origin microbiota during high-temperature Daqu incubation. Food Res. Int. 2022, 156, 111191. [Google Scholar] [CrossRef]
- Li, W.C.; Hou, Q.C.; Wang, Y.J.; Ma, H.M.; Liu, Y.H.; Zhao, F.Y.; Li, J.; Kwok, L.Y.; Yu, J.; Sun, Z.H.; et al. Analysis of the Gut Microbial Diversity of Dairy Cows During Peak Lactation by PacBio Single-Molecule Real-Time (SMRT) Sequencing. Curr. Microbiol. 2018, 75, 1316–1323. [Google Scholar] [CrossRef]
- Du, B.Y.; Meng, L.; Liu, H.M.; Zheng, N.; Zhang, Y.D.; Zhao, S.G.; Wang, J.Q. Single Molecule Real-Time Sequencing and Traditional Cultivation Techniques Reveal Complex Community Structures and Regional Variations of Psychrotrophic Bacteria in Raw Milk. Front. Microbiol. 2022, 13, 853263. [Google Scholar] [CrossRef]
- Jin, C.N.; Liu, J.Y.; Jin, R.Y.; Yao, Y.P.; He, S.L.; Lei, M.; Peng, X.L. Linarin ameliorates dextran sulfate sodium-induced colitis in C57BL/6J mice via the improvement of intestinal barrier, suppression of inflammatory responses and modulation of gut microbiota. Food Funct. 2022, 13, 10574–10586. [Google Scholar] [CrossRef]
- Yang, S.Y.; Xu, W.Q.; Tan, C.L.; Li, M.H.; Li, D.T.; Zhang, C.Y.; Feng, L.K.; Chen, Q.Y.; Jiang, J.; Li, Y.K.; et al. Heat Stress Weakens the Skin Barrier Function in Sturgeon by Decreasing Mucus Secretion and Disrupting the Mucosal Microbiota. Front. Microbiol. 2022, 13, 860079. [Google Scholar] [CrossRef]
- Yang, X.Y.; Guo, X.J.; Liu, W.P.; Tian, Y.Z.; Gao, P.P.; Ren, Y.W.; Zhang, W.; Jiang, Y.J.; Man, C.X. The complex community structures and seasonal variations of psychrotrophic bacteria in raw milk in Heilongjiang Province, China. Lwt-Food Sci. Technol. 2020, 134, 110218. [Google Scholar] [CrossRef]
- Uruc, K.; Tekin, A.; Sahingil, D.; Hayaloglu, A.A. An alternative plant-based fermented milk with kefir culture using apricot (Prunus armeniaca L.) seed extract: Changes in texture, volatiles and bioactivity during storage. Innov. Food Sci. Emerg. Technol. 2022, 82, 103189. [Google Scholar] [CrossRef]
- Montemurro, M.; Pontonio, E.; Coda, R.; Rizzello, C.G. Plant-Based Alternatives to Yogurt: State-of-the-Art and Perspectives of New Biotechnological Challenges. Foods 2021, 10, 316. [Google Scholar] [CrossRef]
- Gronnevik, H.; Falstad, M.; Narvhus, J.A. Microbiological and chemical properties of Norwegian kefir during storage. Int. Dairy J. 2011, 21, 601–606. [Google Scholar] [CrossRef]
- Peng, X.H.; Liao, Y.; Ren, K.Y.; Liu, Y.W.; Wang, M.M.; Yu, A.H.; Tian, T.; Liao, P.L.; Huang, Z.X.; Wang, H.; et al. Fermentation performance, nutrient composition, and flavor volatiles in soy milk after mixed culture fermentation. Process. Biochem. 2022, 121, 286–297. [Google Scholar] [CrossRef]
- Kaczmarska, K.T.; Chandra-Hioe, M.V.; Frank, D.; Arcot, J. Aroma characteristics of lupin and soybean after germination and effect of fermentation on lupin aroma. Lwt-Food Sci. Technol. 2018, 87, 225–233. [Google Scholar] [CrossRef]
- Harle, O.; Falentin, H.; Niay, J.; Valence, F.; Courselaud, C.; Chuat, V.; Maillard, M.B.; Guedon, E.; Deutsch, S.M.; Thierry, A. Diversity of the metabolic profiles of a broad range of lactic acid bacteria in soy juice fermentation. Food Microbiol. 2020, 89, 103410. [Google Scholar] [CrossRef]
- Shi, X.D.; Hao, Z.Q.; Wang, R.C.; Chen, Z.J.; Zuo, F.; Wan, Y.L.; Guo, S.T. Changes of hexanal content in fermented soymilk: Induced by lactic acid bacterial fermentation and thermal treatment. J. Food Process. Preserv. 2022, 46, e16555. [Google Scholar] [CrossRef]
- Wang, K.; Arntfield, S.D. Binding of carbonyl flavours to canola, pea and wheat proteins using GC/MS approach. Food Chem. 2014, 157, 364–372. [Google Scholar] [CrossRef]
- Du, L.; Ro, K.S.; Zhang, Y.J.; Tang, Y.J.; Li, W.B.; Xie, J.L.; Wei, D.Z. Effects of lactiplantibacillus plantarum X7021 on physicochemical properties, purines, isoflavones and volatile compounds of fermented soymilk. Process. Biochem. 2022, 113, 150–157. [Google Scholar] [CrossRef]
- Soukup, S.T.; Helppi, J.; Muller, D.; Zierau, O.; Watzl, B.; Vollmer, G.; Diel, P.; Bub, A.; Kulling, S.E. Phase II metabolism of the soy isoflavones genistein and daidzein in humans, rats and mice: A cross-species and sex comparison. Arch. Toxicol. 2016, 90, 1349. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Concentration of Isoflavones (mg/g) | |||
|---|---|---|---|---|
| Daidzin | Genistin | Daidzein | Genistein | |
| Soybean solution | 1.35 ± 0.02 | 1.74 ± 0.02 | 0.12 ± 0.01 | 0.05 ± 0.01 |
| Soybean kefir | 0.11 ± 0.01**** | 0.13 ± 0.01 **** | 0.74 ± 0.02 **** | 1.14 ± 0.01 **** |
| Digested soybean solution | 0.67 ± 0.10 | 0.81 ± 0.03 | 0.09 ± 0.01 | 0.15 ± 0.02 |
| Digested soybean kefir | 0.13 ± 0.01**** | 0.13 ± 0.01 **** | 0.77 ± 0.06 **** | 0.59 ± 0.02 **** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Liu, S.; Lu, H.; Chen, Q.; Shi, Y. Microbial Community Variations and Bioconversion Improvements during Soybean-Based Fermentation by Kefir Grains. Foods 2023, 12, 1588. https://doi.org/10.3390/foods12081588
Luo J, Liu S, Lu H, Chen Q, Shi Y. Microbial Community Variations and Bioconversion Improvements during Soybean-Based Fermentation by Kefir Grains. Foods. 2023; 12(8):1588. https://doi.org/10.3390/foods12081588
Chicago/Turabian StyleLuo, Jiaqi, Siyu Liu, Hongyun Lu, Qihe Chen, and Ying Shi. 2023. "Microbial Community Variations and Bioconversion Improvements during Soybean-Based Fermentation by Kefir Grains" Foods 12, no. 8: 1588. https://doi.org/10.3390/foods12081588