
Winery By-Products as Sources of Bioactive Tryptophan, Serotonin, and Melatonin: Contributions to the Antioxidant Power




Abstract
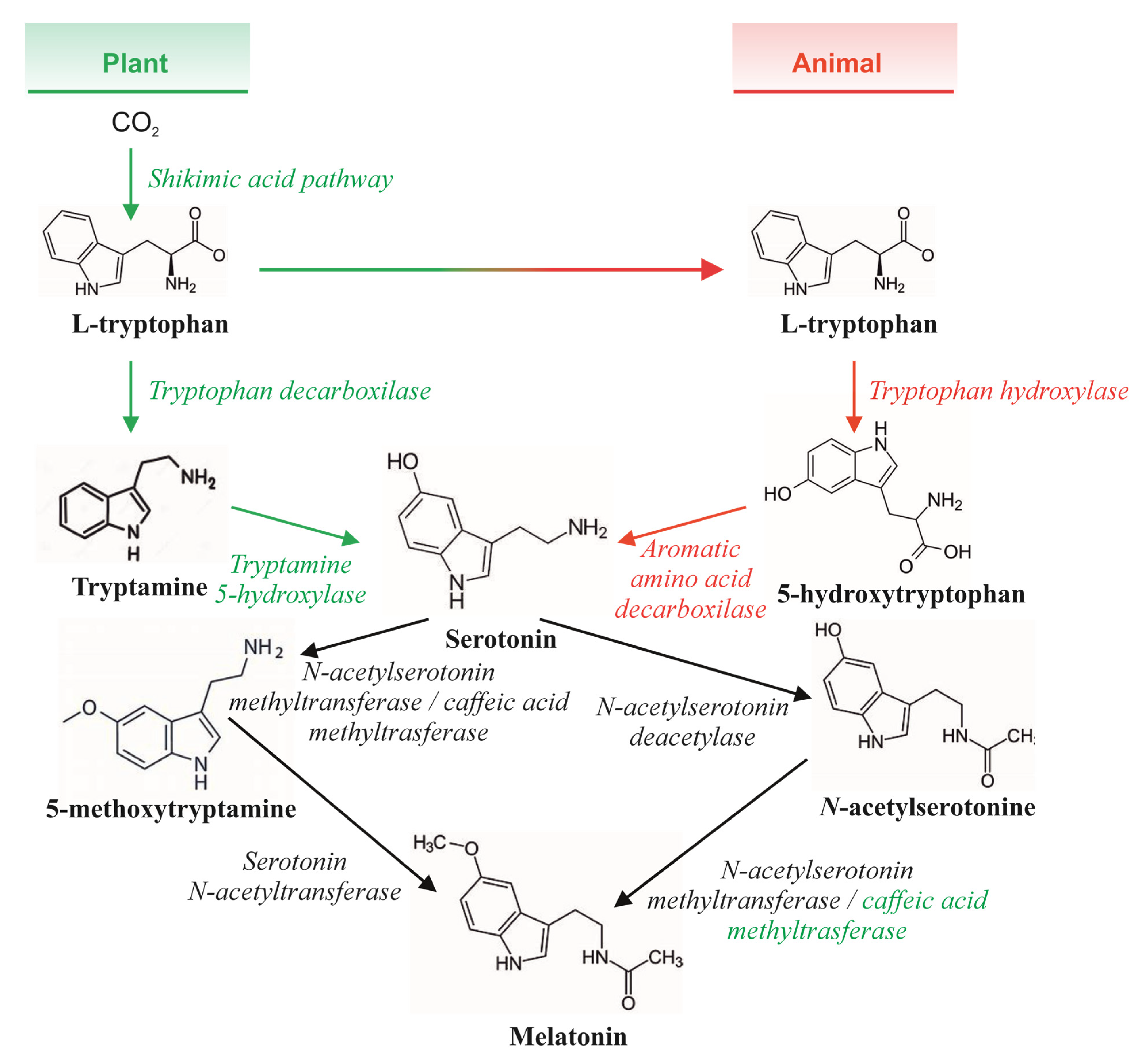
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Extracts
2.3. Derivatization and Analysis of Tryptophan by UHPLC-ESI-QqQ-MS/MS
2.4. Extraction of Serotonin and Melatonin and Analysis by UHPLC-ESI-QqQ-MS/MS
2.5. Determination of the Antioxidant Activity by In Vitro Tests
2.6. Statistical Analysis
3. Results and Discussion
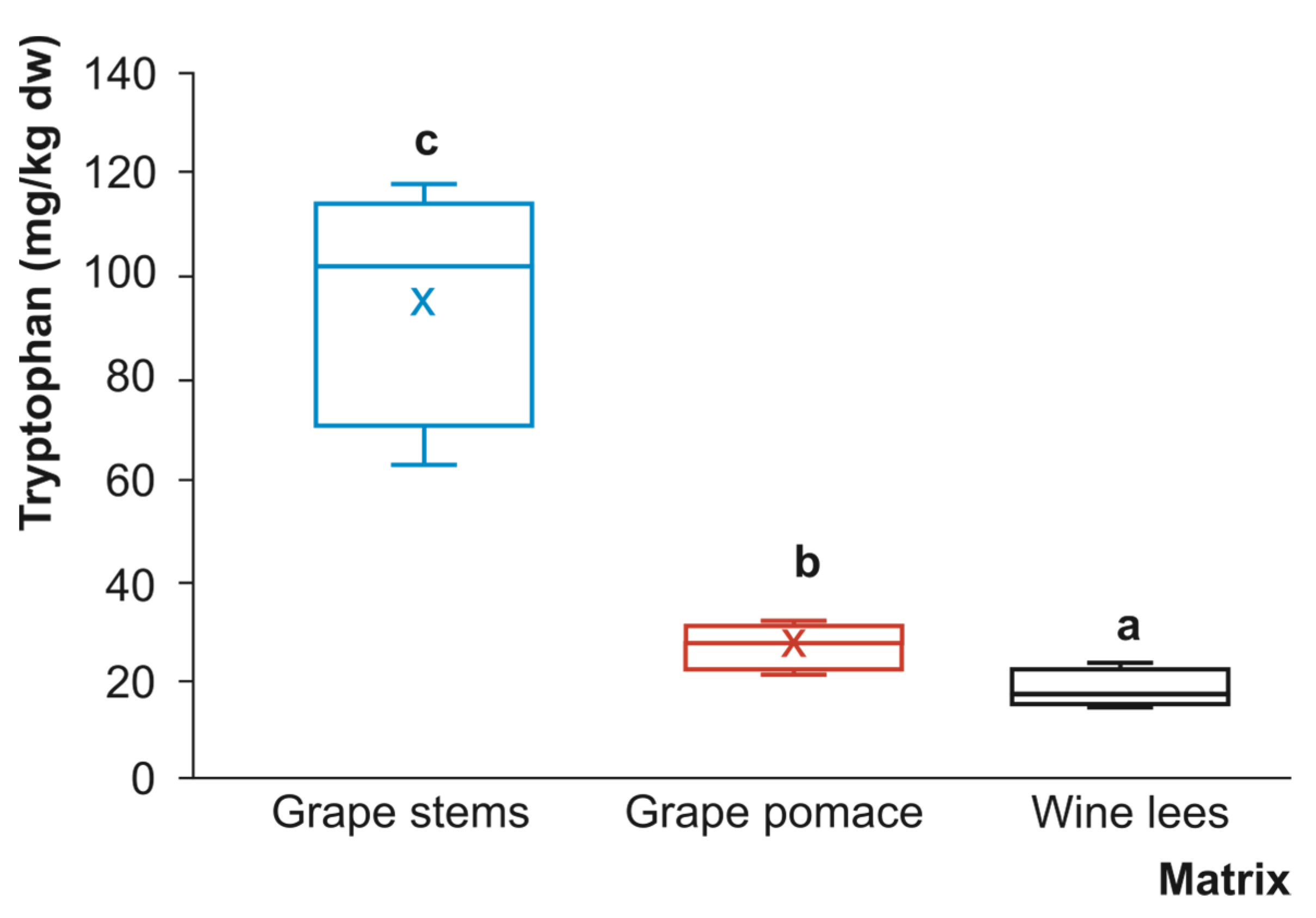
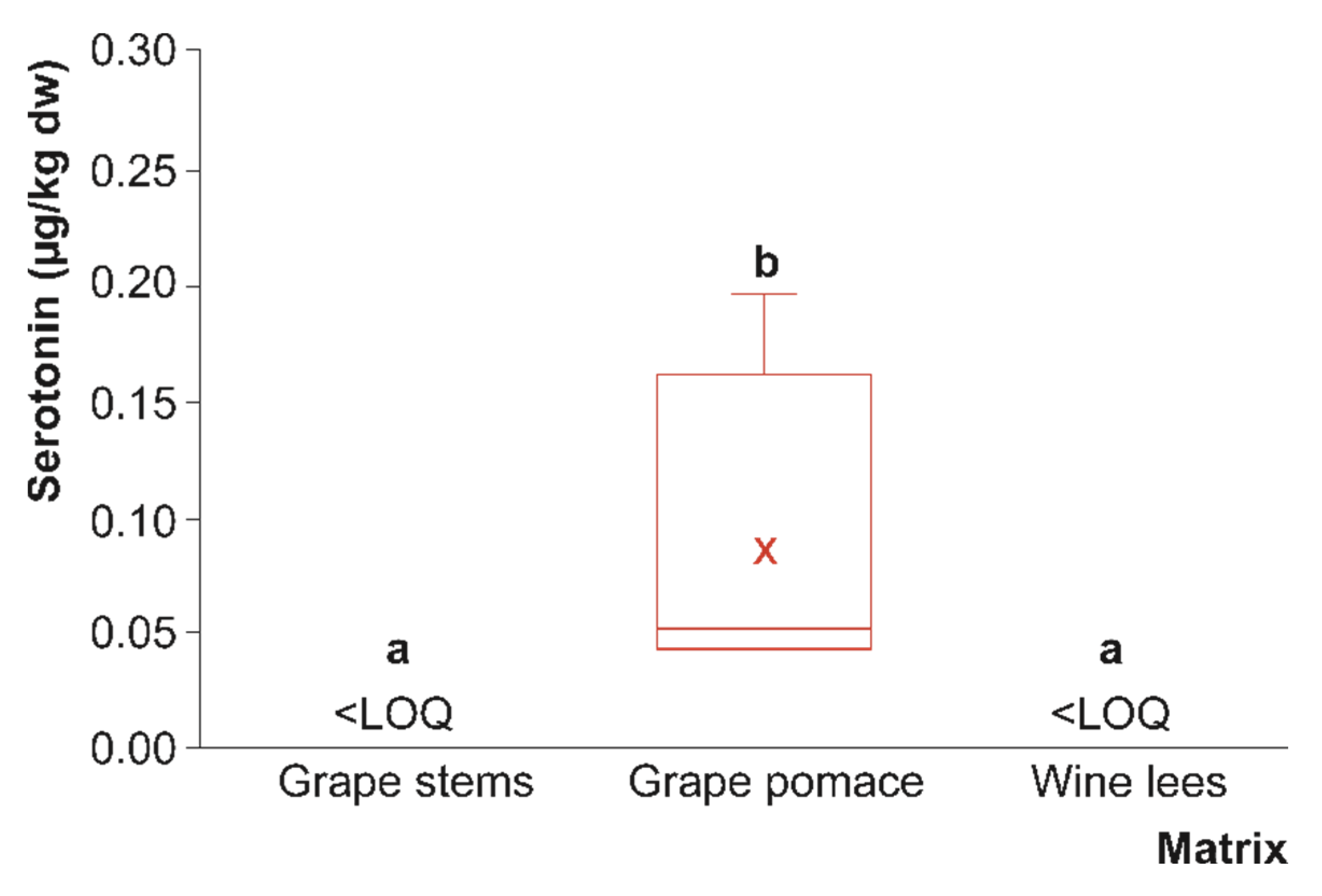
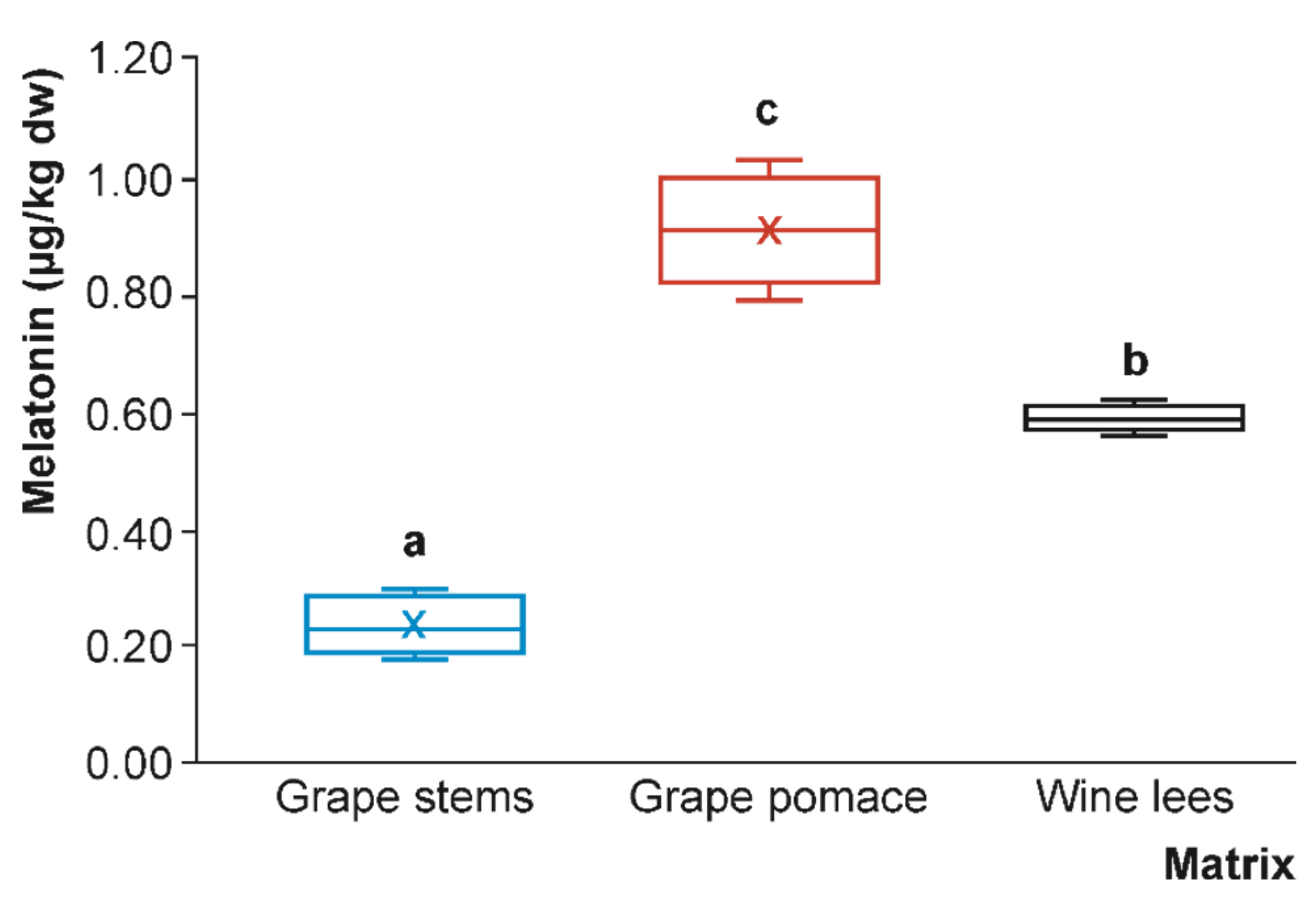
3.1. Tryptophan, Serotonin, and Melatonin Content of Grape By-Products
3.1.1. Tryptophan
3.1.2. Serotonin
3.1.3. Melatonin
3.2. Antioxidant Activity
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Beydemir, S.; Gülçin, İ. Effects of melatonin on carbonic anhydrase from human erythrocytes in vitro and from rat erythrocytes in vivo. J. Enzym. Inhib. Med. Chem. 2008, 19, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: Exceeding expectations. Physiology 2014, 29, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Fidalgo, S.; Ivanov, D.K.; Wood, S.H. Serotonin: From top to bottom. Biogerontology 2013, 14, 21–45. [Google Scholar] [CrossRef]
- Gülçin, I. Measurement of antioxidant ability of melatonin and serotonin by the DMPD and CUPRAC methods as trolox equivalent. J. Enzym. Inhib. Med. Chem. 2008, 23, 871–876. [Google Scholar] [CrossRef]
- Pérez-González, A.; Muñoz-Rugeles, L.; Álvarez-Idaboy, J.R. Tryptophan: Antioxidant or target of oxidative stress? A quantum chemistry elucidation. RSC Adv. 2014, 4, 56128–56131. [Google Scholar] [CrossRef]
- Arnao, M.B. Phytomelatonin: Discovery, Content, and Role in Plants. Adv. Bot. 2014, 2014, 815769. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. The Potential of Phytomelatonin as a Nutraceutical. Molecules 2018, 23, 238. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Cruz, E.; Álvarez-Fernández, M.A.; Valero, E.; Troncoso, A.M.; García-Parrilla, M.C. Melatonin and derived L-tryptophan metabolites produced during alcoholic fermentation by different wine yeast strains. Food Chem. 2017, 217, 431–437. [Google Scholar] [CrossRef]
- Carrión-Antolí, A.; Lorente-Mento, J.M.; Valverde, J.M.; Castillo, S.; Valero, D.; Serrano, M. Effects of melatonin treatment on sweet cherry tree yield and fruit quality. Agronomy 2022, 12, 3. [Google Scholar] [CrossRef]
- Medina-Santamarina, J.; Zapata, P.J.; Valverde, J.M.; Valero, D.; Serrano, M.; Guillén, F. Melatonin treatment of apricot trees leads to maintenance of fruit quality attributes during storage at chilling and non-chilling temperatures. Agronomy 2021, 11, 917. [Google Scholar] [CrossRef]
- Lorente-Mento, J.M.; Guillén, F.; Castillo, S.; Martínez-Romero, D.; Valverde, J.M.; Valero, D.; Serrano, M. Melatonin treatment to pomegranate trees enhances fruit bioactive compounds and quality traits at harvest and during postharvest storage. Antioxidants 2021, 10, 820. [Google Scholar] [CrossRef]
- Mercolini, L.; Mandrioli, R.; Raggi, M.A. Content of melatonin and other antioxidants in grape-related foodstuffs: Measurement using a MEPS-HPLC-F method. J. Pineal Res. 2012, 53, 21–28. [Google Scholar] [CrossRef]
- Meng, J.F.; Shi, T.C.; Song, S.; Zhang, Z.W.; Fang, Y.L. Melatonin in grapes and grape-related foodstuffs: A review. Food Chem. 2017, 231, 185–191. [Google Scholar] [CrossRef]
- Bordiga, M.; Travaglia, F.; Locatelli, M.; Arlorio, M.; Coïsson, J.D. Spent grape pomace as a still potential by-product. Int. J. Food Sci. Technol. 2015, 50, 2022–2031. [Google Scholar] [CrossRef]
- Collado-González, J.; Cruz, Z.N.; Rodríguez, P.; Galindo, A.; Díaz-Baños, F.G.; García de la Torre, J.; Ferreres, F.; Medina, S.; Torrecillas, A.; Gil-Izquierdo, A. Effect of water deficit and domestic storage on the procyanidin profile, size, and aggregation process in pear-jujube (Z. jujuba) fruits. J. Agric. Food Chem. 2013, 61, 6187. [Google Scholar] [CrossRef]
- Salazar, C.; Armenta, J.M.; Shulaev, V. An UPLC-ESI-MS/MS Assay Using 6-Aminoquinolyl-N-Hydroxysuccinimidyl Carbamate Derivatization for Targeted Amino Acid Analysis: Application to Screening of Arabidopsis thaliana Mutants. Metabolites 2012, 2, 398–428. [Google Scholar] [CrossRef] [Green Version]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Sabra, A.; Netticadan, T.; Wijekoon, C. Grape bioactive molecules, and the potential health benefits in reducing the risk of heart diseases. Food Chem. X 2021, 12, 100149. [Google Scholar] [CrossRef]
- Teixeira, A.; Baenas, N.; Dominguez-Perles, R.; Barros, A.; Rosa, E.; Moreno, D.A.; Garcia-Viguera, C. Natural bioactive compounds from winery by-products as health promoters: A review. Int. J. Mol. Sci. 2014, 15, 115638–115678. [Google Scholar] [CrossRef] [Green Version]
- Callejón, R.M.; Troncoso, A.M.; Morales, M.L. Determination of amino acids in grape-derived products: A review. Talanta 2010, 81, 1143–1152. [Google Scholar] [CrossRef]
- Hernández-Orte, P.; Cacho, J.F.; Ferreira, V. Relationship between varietal amino acid profile of grapes and wine aromatic composition. Experiments with model solutions and chemometric study. J. Agric. Food Chem. 2002, 50, 2891–2899. [Google Scholar] [CrossRef]
- Murch, S.J.; Hall, B.A.; Le, C.H.; Saxena, P.K. Changes in the levels of indoleamine phytochemicals during véraison and ripening of wine grapes. J. Pineal Res. 2010, 49, 95–100. [Google Scholar] [CrossRef]
- Rayne, S. Concentrations and profiles of melatonin and serotonin in fruits and vegetables during ripening: A mini-review. Nat. Preced. 2010, 1, 1–3. [Google Scholar] [CrossRef]
- Iriti, M.; Rossoni, M.; Faoro, F. Melatonin content in grape: Myth or panacea? J. Sci. Food Agric. 2006, 86, 1432–1438. [Google Scholar] [CrossRef]
- Jiang, Y.; Ng, T.B.; Wang, C.R.; Li, N.; Wen, T.Y.; Qiao, W.T.; Zhang, D.; Cheng, Z.H.; Liu, F. First isolation of tryptophan from edible lotus (Nelumbo nucifera Gaertn) rhizomes and demonstration of its antioxidant effects. Int. J. Food Sci. Nutr. 2010, 61, 346–356. [Google Scholar] [CrossRef]
- Nayak, B.N.; Buttar, H.S. Evaluation of the antioxidant properties of tryptophan and its metabolites in in vitro assay. J. Complement. Integr. Med. 2016, 13, 129–136. [Google Scholar] [CrossRef]
- Weiss, G.; Diez-Ruiz, A.; Murr, C.; Theur, I.; Fuchs, D. Tryptophan metabolites as scavengers of reactive oxygen and chlorine species. Pteridines 2002, 13, 140–144. [Google Scholar] [CrossRef]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Byeon, Y.; Back, K. Molecular cloning of melatonin 2-hydroxylase responsible for 2-hydroxymelatonin production in rice (Oryza sativa). J. Pineal Res. 2015, 58, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Garde-Cerdán, T.; Gutiérrez-Gamboa, G.; Portu, J.; Fernández-Fernández, J.I.; Gil-Muñoz, R. Impact of phenylalanine and urea applications to Tempranillo and Monastrell vineyards on grape amino acid content during two consecutive vintages. Food Res. Int. 2017, 102, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.C.; Nunes, A.R.; Alves, G.; Silva, L.R. Serotonin and Melatonin: Plant Sources, Analytical Methods, and Human Health Benefits. Rev. Bras. Farmacogn. 2021, 31, 162–175. [Google Scholar] [CrossRef]
- González-Gómez, D.; Lozano, M.; Fernández-León, M.F.; Ayuso, M.C.; Bernalte, M.J.; Rodríguez, A.B. Detection and quantification of melatonin and serotonin in eight Sweet Cherry cultivars (Prunus avium L.). Eur. Food Res. Technol. 2009, 229, 223–229. [Google Scholar] [CrossRef]
- Incesu, M.; Karakus, S.; Seyed Hajizadeh, H.; Ates, F.; Turan, M.; Skalicky, M.; Kaya, O. Changes in Biogenic Amines of Two Table Grapes (cv. Bronx Seedless and Italia) during Berry Development and Ripening. Plants 2022, 11, 2845. [Google Scholar] [CrossRef]
- Rodriguez-Naranjo, M.I.; Gil-Izquierdo, A.; Troncoso, A.M.; Cantos-Villar, E.; Garcia-Parrilla, M.C. Melatonin is synthesised by yeast during alcoholic fermentation in wines. Food Chem. 2011, 126, 1608–1613. [Google Scholar] [CrossRef]
- Fernández-Pachõn, M.S.; Medina, S.; Herrero-Martín, G.; Cerrillo, I.; Berná, G.; Escudero-Lõpez, B.; Ferreres, F.; Martín, F.; García-Parrilla, M.C.; Gil-Izquierdo, A. Alcoholic fermentation induces melatonin synthesis in orange juice. J. Pineal Res. 2014, 56, 31–38. [Google Scholar] [CrossRef]
- Mena, P.; Gil-Izquierdo, Á.; Moreno, D.A.; Martí, N.; García-Viguera, C. Assessment of the melatonin production in pomegranate wines. LWT-Food Sci. Technol. 2012, 47, 13–18. [Google Scholar] [CrossRef]
- Fernandez-Cruz, E.; González, B.; Muñiz-Calvo, S.; Morcillo-Parra, M.Á.; Bisquert, R.; Troncoso, A.M.; Garcia-Parrilla, M.C.; Torija, M.J.; Guillamón, J.M. Intracellular biosynthesis of melatonin and other indolic compounds in Saccharomyces and non-Saccharomyces wine yeasts. Eur. Food Res. Technol. 2019, 245, 1553–1560. [Google Scholar] [CrossRef] [Green Version]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Root, M.M.; Mcginn, M.C.; Nieman, D.C.; Henson, D.A.; Heinz, S.A.; Andrew Shanely, R.; Knab, A.M.; Jin, F. Combined fruit and vegetable intake is correlated with improved inflammatory and oxidant status from a cross-sectional study in a community setting. Nutrients 2012, 4, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L. Oxygen radical absorbance capacity (ORAC): New horizons in relating dietary antioxidants/bioactives and health benefits. J. Funct. Foods 2015, 18, 797–810. [Google Scholar] [CrossRef]
- Ratha, J.; Yongram, C.; Panyatip, P.; Powijitkul, P.; Siriparu, P.; Datham, S.; Priprem, A.; Srisongkram, T.; Puthongking, P. Polyphenol and Tryptophan Contents of Purple Corn (Zea mays L.) Variety KND and Butterfly Pea (Clitoria ternatea) Aqueous Extracts: Insights into Phytochemical Profiles with Antioxidant Activities and PCA Analysis. Plants 2023, 12, 603. [Google Scholar] [CrossRef]
- Costa-Pérez, A.; Medina, S.; Sánchez-Bravo, P.; Domínguez-Perles, R.; García-Viguera, C. The (Poly)phenolic Profile of Separate Winery By-Products Reveals Potential Antioxidant Synergies. Molecules 2023, 28, 2081. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef]
- Lucas-Abellán, C.; Mercader-Ros, M.T.; Zafrilla, M.P.; Gabaldón, J.A.; Núñez-Delicado, E. Comparative study of different methods to measure antioxidant activity of resveratrol in the presence of cyclodextrins. Food Chem. Toxicol. 2011, 49, 1255–1260. [Google Scholar] [CrossRef]
- Schlesier, K.; Harwat, M.; Böhm, V.; Bitsch, R. Assessment of antioxidant activity by using different in vitro methods. Free Radic. Res. 2002, 36, 177–187. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Poeggeler, B.; Lopez-Burillo, S.; Mayo, J.C.; Sainz, R.M.; Reiter, R.J. Mechanistic and comparative studies of melatonin and classic antioxidants in terms of their interactions with the ABTS cation radical. J. Pineal Res. 2003, 34, 249–259. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | MRM Quantitative Transition | Fragmentor (V) | Collision Energy (eV) | ESI Mode | LOD (pmol/L) | LOQ (pmol/L) |
|---|---|---|---|---|---|---|
| Tryptophan | 375.0 > 171.2 | 120 | 0 | Positive | 31 | 31 |
| Serotonin | 177.0 > 159.9 | 90 | 0 | Positive | 250 | 500 |
| Melatonin | 233.0 > 174.3 | 90 | 0 | Positive | 31 | 61 |
| Method | By-Product | Extraction for Tryptophan Analysis (mmol TE/kg dw) | Extraction for Serotonin and Melatonin Analysis (µmol TE/kg dw) |
|---|---|---|---|
| FRAP | |||
| Grape stems | 142.86 ± 8.04 a | 49.04 ± 2.69 a | |
| Grape pomace | 19.05 ± 0.80 c | 18.52 ± 0.85 b | |
| Wine lees | 53.17 ± 1.00 b | 18.80 ± 0.36 b | |
| ABTS•+ | |||
| Grape stems | 166.72 ± 6.77 a | 59.53 ± 2.31 a | |
| Grape pomace | 17.84 ± 0.99 c | 19.31 ± 1.73 b | |
| Wine lees | 45.96 ± 1.02 b | 15.46 ± 1.22 b | |
| ORAC | |||
| Grape stems | 363.24 ± 11.0 a | 153.77 ± 13.03 a | |
| Grape pomace | 36.41 ± 1.58 c | 46.22 ± 2.56 b | |
| Wine lees | 128.33 ± 13.0 b | 56.52 ± 4.27 b |
| Method | Standard Concentration (µM) | µM TE | Linear Equation | R2 |
|---|---|---|---|---|
| Serotonin | ||||
| FRAP | 100 | 162.9 ± 1.4 | y = 1.62x + 2.4 | 0.999 |
| ABTS•+ | 100 | 182.6 ± 3.8 | y = 1.74x + 14.5 | 0.988 |
| ORAC | 10 | 42.5 ± 0.3 | y = 4.75x − 3.9 | 0.999 |
| Melatonin | ||||
| FRAP | 100 | N.d. | N.d. | N.d. |
| ABTS•+ | 100 | N.d. | N.d. | N.d. |
| ORAC | 10 | 45.8 ± 0.9 | y = 4.20x − 8.4 | 0.998 |
| Tryptophan | ||||
| FRAP | 100 | N.d. | N.d. | N.d. |
| ABTS•+ | 100 | N.d. | N.d. | N.d. |
| ORAC | 10 | 33.5 ± 2.7 | y = 5.02x − 4.0 | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baenas, N.; García-Viguera, C.; Domínguez-Perles, R.; Medina, S. Winery By-Products as Sources of Bioactive Tryptophan, Serotonin, and Melatonin: Contributions to the Antioxidant Power. Foods 2023, 12, 1571. https://doi.org/10.3390/foods12081571
Baenas N, García-Viguera C, Domínguez-Perles R, Medina S. Winery By-Products as Sources of Bioactive Tryptophan, Serotonin, and Melatonin: Contributions to the Antioxidant Power. Foods. 2023; 12(8):1571. https://doi.org/10.3390/foods12081571
Chicago/Turabian StyleBaenas, Nieves, Cristina García-Viguera, Raúl Domínguez-Perles, and Sonia Medina. 2023. "Winery By-Products as Sources of Bioactive Tryptophan, Serotonin, and Melatonin: Contributions to the Antioxidant Power" Foods 12, no. 8: 1571. https://doi.org/10.3390/foods12081571