
Evaluation of Preharvest Melatonin on Soft Rot and Quality of Kiwifruit Based on Principal Component Analysis
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. In Vitro Melatonin Inhibition Test
2.3. Field Experiments
2.4. Sampling and Analysis of Fruit
2.5. Fruit Soft Rot Resistance Test
2.5.1. Effects of Preharvest MT on Postharvest Kiwifruit Inoculated with B. dothidea
2.5.2. Effects of Preharvest MT on Disease Incidence and Severity of Postharvest Kiwifruit
value)/(Total No. of fruit × the highest disease severity value) × 100
2.6. Effects of Preharvest Melatonin on Quality Parameters of Kiwifruit
2.7. Statistical Analysis
3. Results
3.1. Inhibitory Effect of MT In Vitro
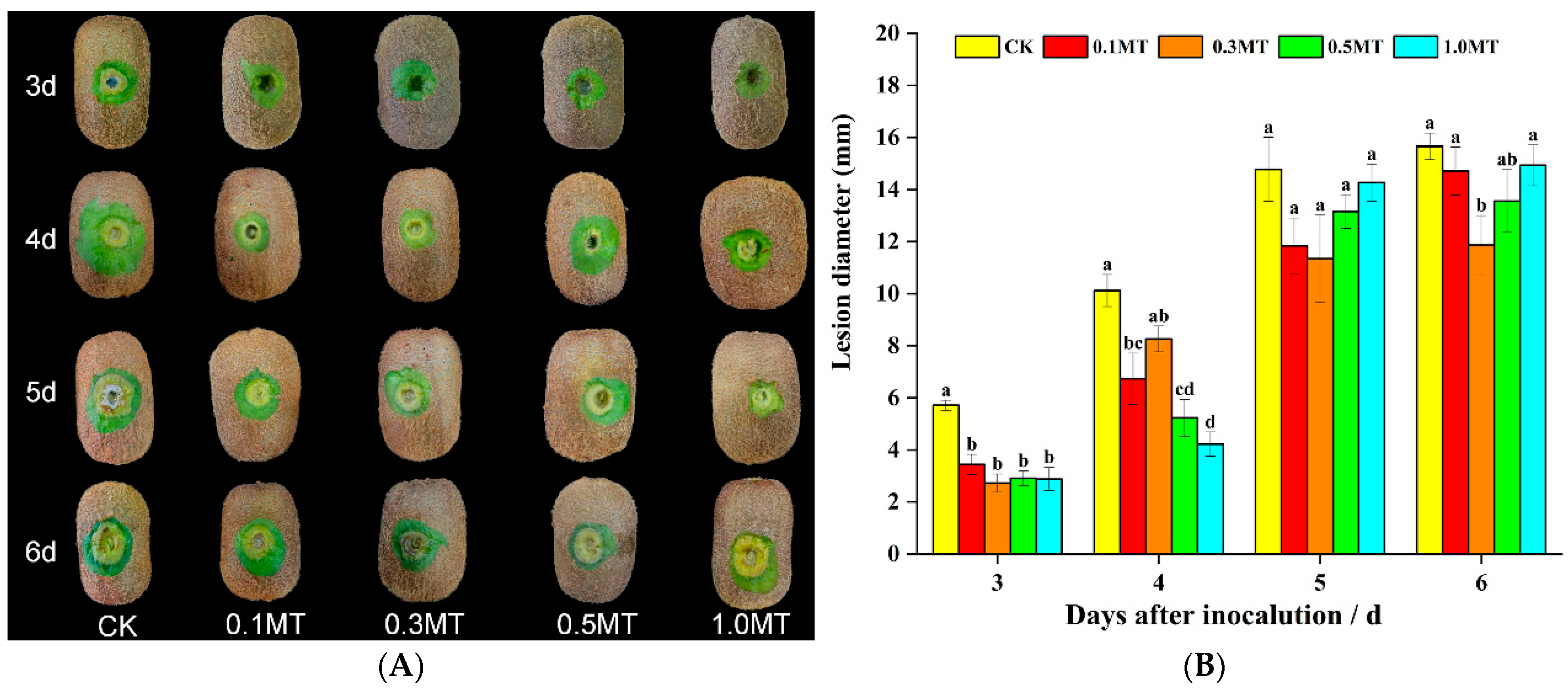
3.2. Effect of Preharvest MT Treatment on Disease Resistance of Kiwifruit
3.3. Effect of Preharvest MT Treatment on Appearance Quality of Kiwifruit
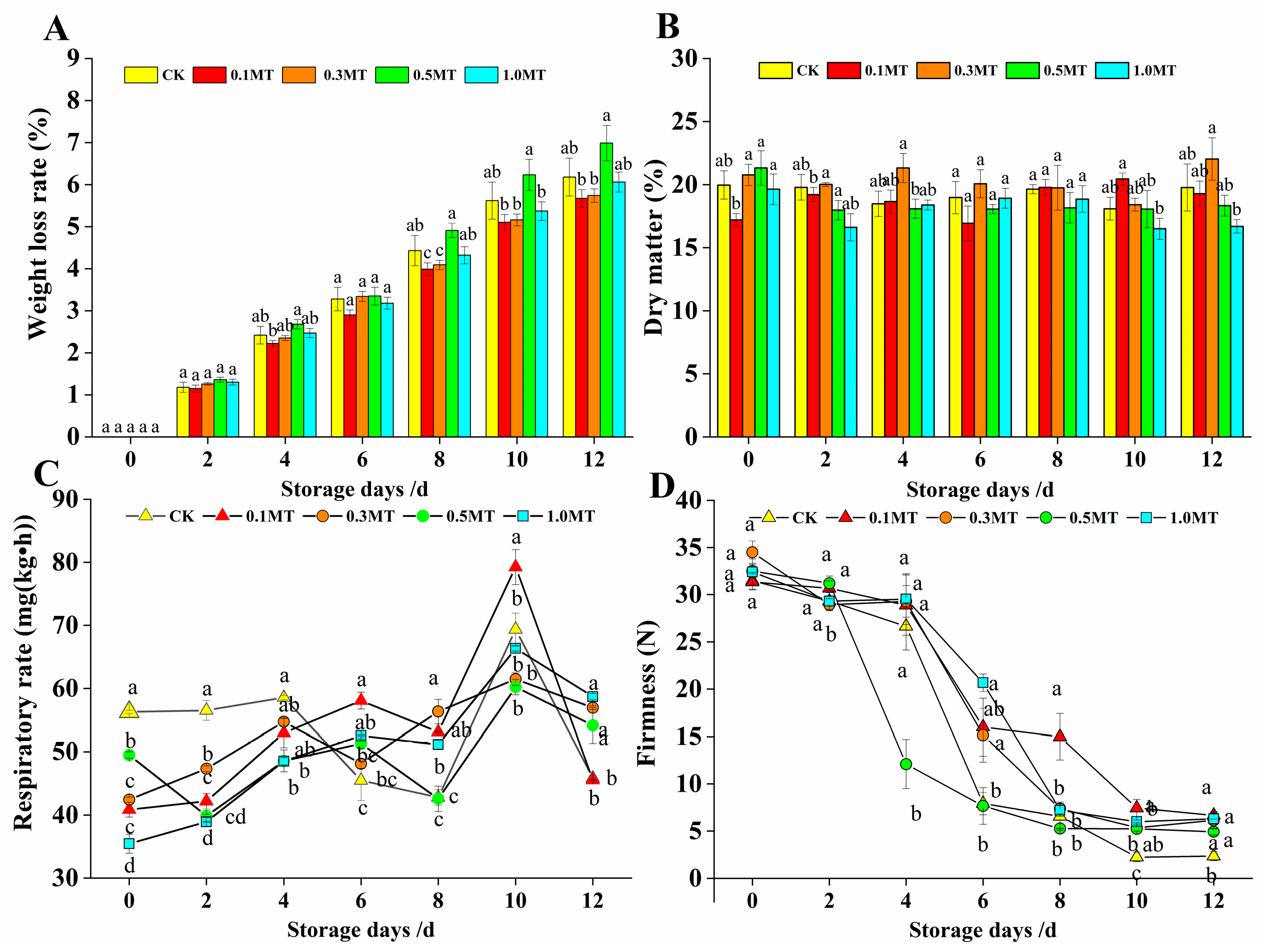
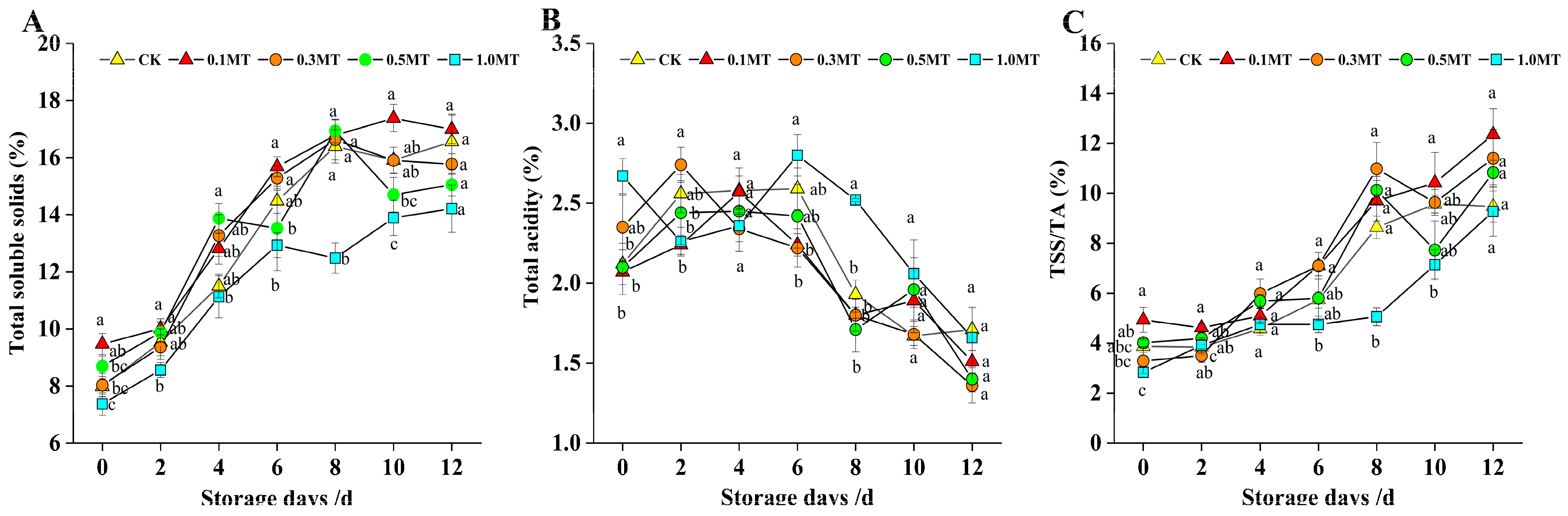
3.4. Effects of Preharvest MT Treatment on the Storability of Kiwifruit
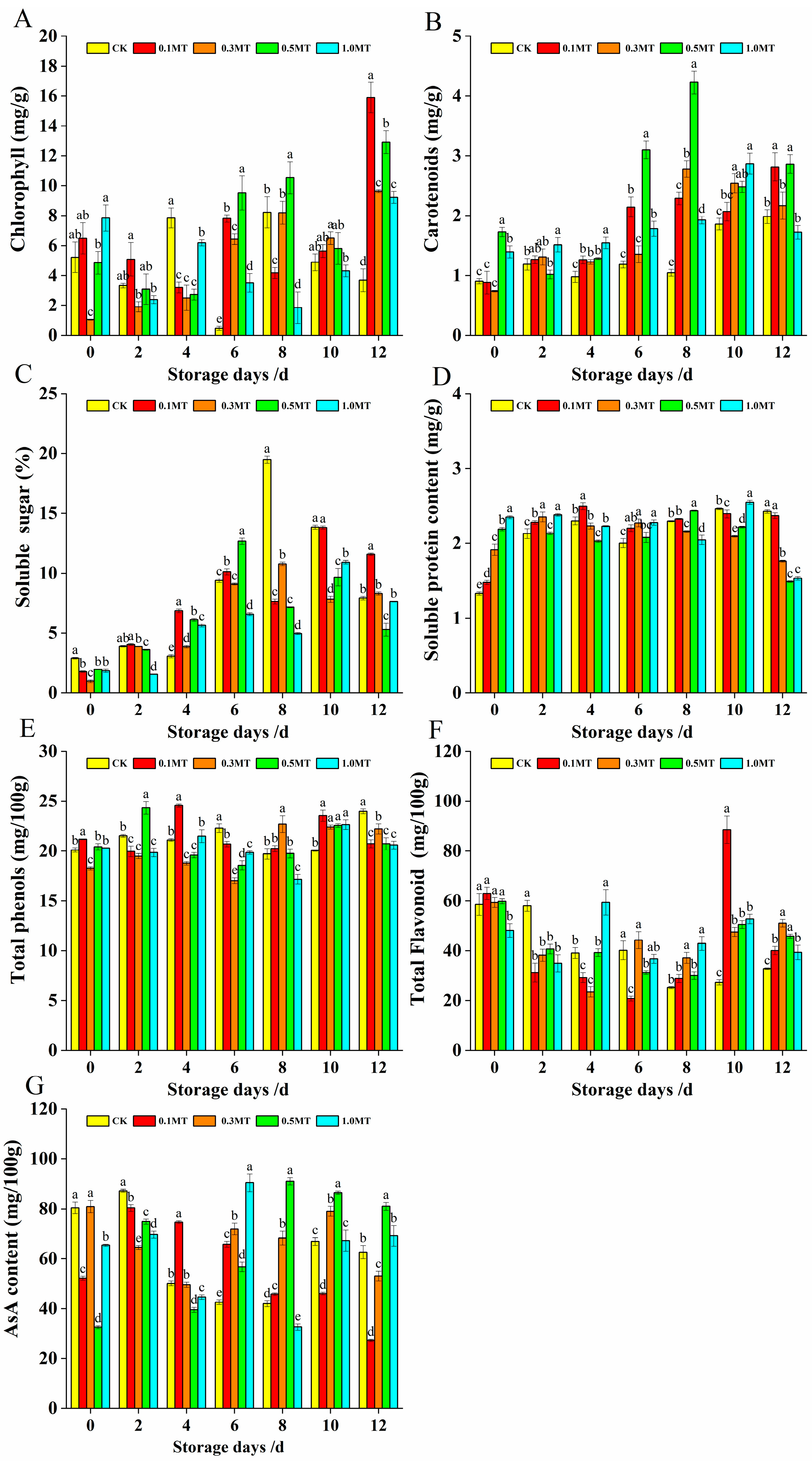
3.5. Effects of Preharvest MT Treatment on Nutritional Components of Postharvest Kiwifruit
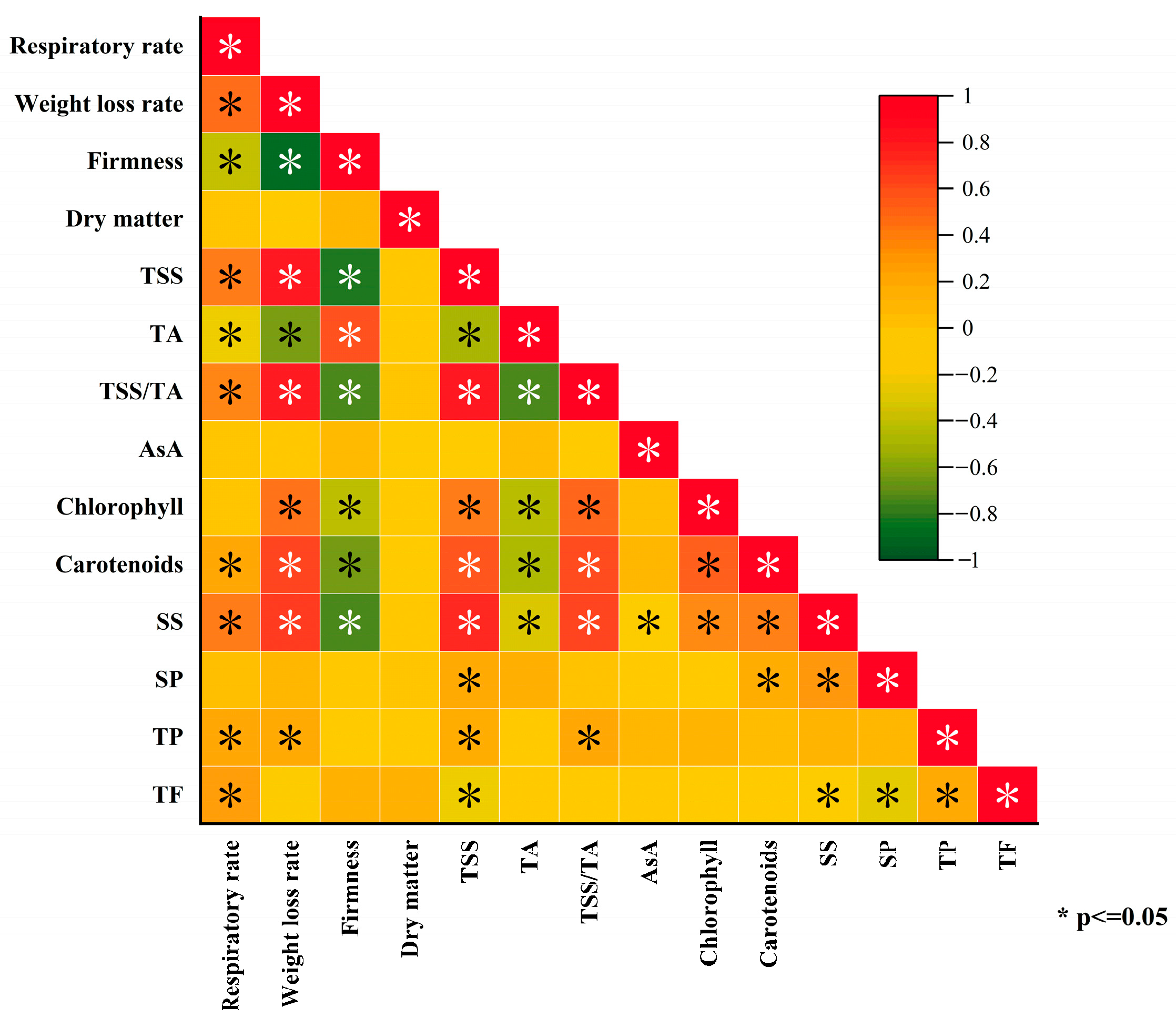
3.6. Correlation Analysis of Preharvest MT Treatment on Kiwifruit Storage Quality
3.7. Principal Component Analysis of Preharvest MT Treatment on Kiwifruit Storage Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Dong, H.; Lan, J.; Liu, Y.; Liang, K.; Lu, Q.; Fang, Z.; Liu, P. High-quality genome resource of the pathogen of Diaporthe (Phomopsis) phragmitis causing kiwifruit soft rot. Mol. Plant-Microbe Interact. 2021, 34, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Billing, D.; Pidakala, P.; Burdon, J. Textural changes in ‘Hayward’kiwifruit during and after storage in controlled atmospheres. Sci. Hortic. 2017, 222, 40–45. [Google Scholar] [CrossRef]
- Mao, T.T.; Mo, W.D.; Zhao, D.L.; Wang, N.; Tao, G. Isolation and identification of phosphate-solubilizing antagonistic fungus and its biocontrol effects against kiwifruit soft rot. J. South Agric. 2019, 50, 1748–1755. [Google Scholar]
- Pan, L.Y.; Zhao, X.Y.; Chen, M.; Fu, Y.Q.; Xiang, M.L.; Chen, J.Y. Effect of exogenous methyl jasmonate treatment on disease resistance of postharvest kiwifruit. Food Chem. 2020, 305, 125483. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Fu, S.; Fan, G.; Li, D.M.; Yang, S.Z.; Peng, L.T.; Pan, S.Y. Active compound identification by screening 33 essential oil monomers against Botryosphaeria dothidea from postharvest kiwifruit and its potential action mode. Pestic. Biochem. Physiol. 2021, 179, 104957. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, D.; Aravena, Z.; Valdes-Gomez, H.; Latorre, B.A.; Diaz, G.A.; Pablo Zoffoli, J. Characterization of Botrytis cinerea and B. prunorum From Healthy Floral Structures and Decayed ‘Hayward’ Kiwifruit During Post-Harvest Storage. Plant Dis. 2021, 105, 2129–2140. [Google Scholar] [CrossRef]
- Wang, Q.P.; Zhang, C.; Long, Y.H.; Wu, X.M.; Su, Y.; Lei, Y.; Ai, Q. Bioactivity and control efficacy of the novel antibiotic tetramycin against various kiwifruit diseases. Antibiotics 2021, 10, 289. [Google Scholar] [CrossRef]
- Xu, X.D.; Chen, Y.; Li, B.Q.; Zhang, Z.q.; Qin, G.Z.; Chen, T.; Tian, S.P. Molecular mechanisms underlying multi-level defense responses of horticultural crops to fungal pathogens. Hortic. Res. 2022, 9, uhac066. [Google Scholar] [CrossRef]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Gutierrez Martinez, P.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernandez Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [Green Version]
- Sharif, R.; Xie, C.; Zhang, H.; Arnao, M.B.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Nawaz, M.A.; Chen, P. Melatonin and its effects on plant systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.H.; Cao, S.F.; Xie, K.Q.; Chi, Z.Y.; Wang, J.; Wang, H.F.; Wei, Y.Y.; Shao, X.F.; Zhang, C.D.; Xu, F.; et al. Melatonin delays yellowing of broccoli during storage by regulating chlorophyll catabolism and maintaining chloroplast ultrastructure. Postharvest Biol. Technol. 2021, 172, 111378. [Google Scholar] [CrossRef]
- Xiang, M.L.; Wu, F.; Li, S.C.; Wang, Y.B.; Xiao, L.H.; Peng, W.W.; Chen, J.Y.; Chen, M. Effects of melatonin treatment on resistance to black spot and postharvest storage quality of pear fruit. Sci. Agric. Sin. 2022, 55, 785–795. [Google Scholar]
- Fan, S.L.; Xiong, T.T.; Lei, Q.M.; Tan, Q.; Cai, J.; Song, Z.; Yang, M.; Chen, W.; Li, X.; Zhu, X. Melatonin treatment improves postharvest preservation and resistance of guava fruit (Psidium guajava L.). Foods 2022, 11, 262. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Wang, T.; Liu, G.S.; Hu, M.J.; Yun, Z.; Duan, X.W.; Cai, K.; Jiang, G.X. Inhibition of downy blight and enhancement of resistance in litchi fruit by postharvest application of melatonin. Food Chem. 2021, 347, 129009. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.C.; Huang, Y.; Lian, S.; Saleem, M.; Li, B.H.; Wang, C.X. Improving the biocontrol efficacy of Meyerozyma guilliermondii Y-1 with melatonin against postharvest gray mold in apple fruit. Postharvest Biol. Technol. 2021, 171, 111351. [Google Scholar] [CrossRef]
- Li, Z.Z.; Zhang, S.J.; Xue, J.X.; Mu, B.Y.; Song, H.; Liu, Y.P. Exogenous melatonin treatment induces disease resistance against Botrytis cinerea on postharvest grapes by activating defence responses. Foods 2022, 11, 2231. [Google Scholar] [CrossRef]
- Li, L.; Pan, H.; Deng, L.; Li, W.Y.; Feng, D.D.; Zhong, C.H. Occurrence rule and comprehends control of kiwifruit rot disease. China Fruits 2020, 6, 1–5. [Google Scholar]
- Liu, Y.X.; Zhu, Y.M.; Zhou, E.X. Research progress on the action mechanism and application of plant immune inducers. Mol. Plant Breed. 2020, 18, 1020–1026. [Google Scholar]
- Sun, M.Z.; Zhang, S.; Wu, G.B.; Chen, F.H.; Ni, H.; Chen, X.L.; Xu, M.; Lin, H.T. Research advances in the fresh-keeping effects of preharvest treatment on postharvest fruit. J. Fruit Sci. 2022, 39, 1111–1120. [Google Scholar]
- Li, X.Q.; Long, Y.H.; Yin, X.H.; Wu, X.M.; Zhao, Z.B.; Fan, R.; Mo, F.X.; Jiang, Y.L.; Huang, Y.X.; Tang, J.W. Mechanism of action of methyl jasmonate against kiwifruit soft rot and its effect on fruit quality. Food Sci. 2019, 40, 239–248. [Google Scholar]
- Zhang, C.; Long, Y.H.; Li, J.H.; Li, M.; Xing, D.K.; An, H.M.; Wu, X.M.; Wu, Y.Y. A chitosan composite film sprayed before pathogen infection effectively controls postharvest soft rot in kiwifruit. Agronomy 2020, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Du, T.H.; Zhou, X.T.; Zhu, L.Y.; Zhang, J.; Zou, Z.R. Effect of melatonin treatment on tomato fruit quality and volatile compounds under salt stress. Food Sci. 2016, 37, 69–76. [Google Scholar]
- Li, S.E.; Cheng, Y.; Yan, R.; Liu, Y.; Huan, C.; Zheng, X.L. Preharvest spray with melatonin improves postharvest disease resistance in cherry tomato fruit. Postharvest Biol. Technol. 2022, 193, 112055. [Google Scholar] [CrossRef]
- Sheng, J.P.; Zhao, R.R.; Chen, L.L.; Shen, L. Effect of pre-harvest melatonin spraying on the postharvest disease resistance and storage quality of tomato fruit. Food Sci. 2020, 41, 188–193. [Google Scholar]
- Ali, I.; Wang, X.K.; Tareen, M.J.; Wattoo, F.M.; Qayyum, A.; Hassan, M.U.; Shafique, M.; Liaquat, M.; Asghar, S.; Hussain, T.; et al. Foliar application of salicylic acid at different phenological stages of peach fruit cv. ‘Flordaking’ improves harvest quality and reduces chilling injury during low temperature storage. Plants 2021, 10, 1981. [Google Scholar] [CrossRef]
- Correia, S.; Queiros, F.; Ferreira, H.; Morais, M.C.; Afonso, S.; Silva, A.P.; Goncalves, B. Foliar application of calcium and growth regulators modulate sweet cherry (Prunus avium L.) tree performance. Plants 2020, 9, 410. [Google Scholar] [CrossRef] [Green Version]
- Deng, B.L.; Xia, C.B.; Tian, S.; Shi, H.J. Melatonin reduces pesticide residue, delays senescence, and improves antioxidant nutrient accumulation in postharvest jujube fruit. Postharvest Biol. Technol. 2021, 173, 111419–111427. [Google Scholar] [CrossRef]
- Zhang, B.; Lei, S.J.; Zhang, L.; Shi, S.S. Optimization of extraction process of chlorophyll from kiwifruit by response surface method. Sci. Technol. Food Ind. 2019, 40, 199–204. [Google Scholar]
- Liu, C.H.; Zheng, H.H.; Sheng, K.L.; Liu, W.; Zheng, L. Effects of melatonin treatment on the postharvest quality of strawberry fruit. Postharvest Biol. Technol. 2018, 139, 47–55. [Google Scholar] [CrossRef]
- Wang, Q.P.; Zhang, C.; Wu, X.M.; Long, Y.H.; Su, Y. Chitosan augments tetramycin against soft rot in kiwifruit and enhances its improvement for kiwifruit growth, quality and aroma. Biomolecules 2021, 11, 1257. [Google Scholar] [CrossRef] [PubMed]
- Li, S.E.; Xu, Y.H.; Bi, Y.; Zhang, B.; Shen, S.L.; Jiang, T.J.; Zheng, X.L. Melatonin treatment inhibits gray mold and induces disease resistance in cherry tomato fruit during postharvest. Postharvest Biol. Technol. 2019, 157, 110962. [Google Scholar] [CrossRef]
- Li, T.T.; Wu, Q.X.; Zhu, H.; Zhou, Y.J.; Jiang, Y.M.; Gao, H.J.; Yun, Z. Comparative transcriptomic and metabolic analysis reveals the effect of melatonin on delaying anthracnose incidence upon postharvest banana fruit peel. BMC Plant Biol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.M.; Zheng, X.Z.; Reiter, R.J.; Feng, S.; Wang, Y.; Liu, S.; Jin, L.; Li, Z.G.; Datla, R.; Ren, M.Z. Melatonin attenuates potato late blight by disrupting cell growth, stress tolerance, fungicide susceptibility and homeostasis of gene expression in Phytophthora infestans. Front Plant Sci. 2017, 8, 1993. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.M.; Liu, S.; Zhang, J.K.; Reiter, R.J.; Wang, Y.; Qiu, D.; Luo, X.M.; Khalid, A.R.; Wang, H.Y.; Feng, L.; et al. Synergistic anti-oomycete effect of melatonin with a biofungicide against oomycetic black shank disease. J. Pineal Res. 2018, 65, e12492. [Google Scholar] [CrossRef]
- Gao, S.W.; Ma, W.Y.; Lyu, X.N.; Cao, X.L.; Yao, Y.X. Melatonin may increase disease resistance and flavonoid biosynthesis through effects on DNA methylation and gene expression in grape berries. BMC Plant Biol. 2020, 20, 231. [Google Scholar] [CrossRef]
- Zhang, L.L.; Yu, Y.W.; Chang, L.L.; Wang, X.J.; Zhang, S.Y. Melatonin enhanced the disease resistance by regulating reactive oxygen species metabolism in postharvest jujube fruit. J. Food Process Pres. 2022, 46, e16363. [Google Scholar] [CrossRef]
- Zhang, C.; Long, Y.-H.; Wang, Q.-P.; Li, J.H.; Wu, X.M.; Li, M. The effect of preharvest 28.6% chitosan composite film sprays for controlling the soft rot on kiwifruit. Hortic. Sci. 2019, 46, 180–194. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.F.; Xu, T.F.; Song, C.Z.; Yu, Y.; Hu, F.; Zhang, L.; Zhang, Z.W.; Xi, Z.M. Melatonin treatment of pre-veraison grape berries to increase size and synchronicity of berries and modify wine aroma components. Food Chem. 2015, 185, 127–134. [Google Scholar] [CrossRef]
- Lorente-Mento, J.M.; Guillen, F.; Castillo, S.; Martinez-Romero, D.; Valverde, J.M.; Valero, D.; Serrano, M. Melatonin treatment to pomegranate trees enhances fruit bioactive compounds and quality traits at harvest and during postharvest storage. Antioxidants 2021, 10, 820. [Google Scholar] [CrossRef]
- Carrion Antoli, A.; Lorente-Mento, J.M.; Valverde, J.M.; Castillo, S.; Valero, D.; Serrano, M. Effects of Melatonin treatment on sweet cherry tree yield and fruit quality. Agronomy 2022, 12, 3. [Google Scholar] [CrossRef]
- Liu, J.L.; Yue, R.R.; Si, M.; Wu, M.; Cong, L.; Zhai, R.; Yang, C.Q.; Wang, Z.G.; Ma, F.W.; Xu, L.F. Effects of exogenous application of melatonin on quality and sugar metabolism in ‘Zaosu’pear fruit. J. Plant Growth Regul. 2019, 38, 1161–1169. [Google Scholar] [CrossRef]
- Liu, J.L.; Zhang, R.M.; Sun, Y.K.; Liu, Z.Y.; Jin, W.; Sun, Y. The beneficial effects of exogenous melatonin on tomato fruit properties. Sci. Hortic. 2016, 207, 14–20. [Google Scholar] [CrossRef]
- Sun, H.L.; Wang, X.Y.; Shang, Y.; Wang, X.Q.; Du, G.D.; Lu, D.G. Preharvest application of melatonin induces anthocyanin accumulation and related gene upregulation in red pear (Pyrus ussuriensis). J. Integr. Agric. 2021, 20, 2126–2137. [Google Scholar] [CrossRef]
- Fekry, W.M.; Rashad, Y.M.; Alaraidh, I.A.; Mehany, T. Exogenous Application of melatonin and methyl jasmonate as a pre-harvest treatment enhances growth of barhi date palm trees, prolongs storability, and maintains quality of their fruits under storage conditions. Plants 2021, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Medina-Santamarina, J.; Zapata, P.J.; Valverde, J.M.; Valero, D.; Serrano, M.; Guillen, F. Melatonin treatment of apricot trees leads to maintenance of fruit quality attributes during storage at chilling and non-chilling temperatures. Agronomy 2021, 11, 917. [Google Scholar] [CrossRef]
- Michailidis, M.; Tanou, G.; Sarrou, E.; Karagiannis, E.; Ganopoulos, I.; Martens, S.; Molassiotis, A. Pre- and postharvest melatonin application boosted phenolic compounds accumulation and altered respiratory characters in sweet cherry fruit. Front. Nutr. 2021, 8, 306–313. [Google Scholar] [CrossRef]
- Wang, L.; Yang, M.; Dong, Y.; Reiter, R.J.; Xu, Y.; Lin, X.; Luo, Z.; Li, L. Melatonin confers enhanced polyamine metabolism and cell tolerance in Vitis vinifera against oxidative damage: Quantitative proteomic evidence. Postharvest Biol. Technol. 2022, 184, 111756. [Google Scholar] [CrossRef]
- Pang, L.L.; Wu, Y.; Pan, Y.F.; Ban, Z.; Li, L.; Li, X. Insights into exogenous melatonin associated with phenylalanine metabolism in postharvest strawberry. Postharvest Biol. Technol. 2020, 168, 111244. [Google Scholar] [CrossRef]
- Dhakar, M.K.; Das, B.; Nath, V.; Sarkar, P.K.; Singh, A.K. Genotypic diversity for fruit characteristics in bael Aegle marmelos (L.) Corr. based on principal component analysis. Genet. Resour. Crop Evol. 2019, 66, 951–964. [Google Scholar] [CrossRef]
- Germano, T.A.; Aguiar, R.P.; Rocha Bastos, M.S.; Moreira, R.A.; Ayala-Zavala, J.F.; Alcantara de Miranda, M.R. Galactomannan-carnauba wax coating improves the antioxidant status and reduces chilling injury of ‘Paluma’ guava. Postharvest Biol. Technol. 2019, 149, 9–17. [Google Scholar] [CrossRef]
- Zhang, G.X.; Fu, Q.Q.; Fu, Z.T.; Li, X.X.; Matetic, M.; Bakaric, M.B.; Jemric, T. A Comprehensive Peach Fruit Quality Evaluation Method for Grading and Consumption. Appl. Sci. 2020, 10, 1348. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.L.; Xu, J.J.; Wang, Y.J.; Li, K.; Zong, Y.; Yang, L.; Chen, W.R.; Liao, F.L.; Guo, W.D. The Xenia Effect Promotes Fruit Quality and Assists in Optimizing Cross Combinations in ‘O’Neal’and ‘Emerald’Blueberry. Horticulturae 2022, 8, 659. [Google Scholar] [CrossRef]
- Xie, J.; Qin, Z.Y.; Pan, J.L.; Li, J.; Li, X.; Khoo, H.E.; Dong, X.H. Melatonin treatment improves postharvest quality and regulates reactive oxygen species metabolism in “Feizixiao” litchi based on principal component analysis. Front. Plant Sci. 2022, 13, 965345. [Google Scholar] [CrossRef] [PubMed]
- Nowicka, P.; Wojdyło, A.; Laskowski, P. Principal component analysis (PCA) of physicochemical compounds’ content in different cultivars of peach fruits, including qualification and quantification of sugars and organic acids by HPLC. Eur. Food Res. Technol. 2019, 245, 929–938. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Colony Diameter (mm) | Inhibitory Rate (%) |
|---|---|---|
| Control | 78.33 ± 2.49 a | - |
| 0.1 MT | 56.17 ± 4.76 b | 30.65 ± 6.59 c |
| 0.3 MT | 54.67 ± 5.18 bc | 32.72 ± 6.59 bc |
| 0.5 MT | 41.67 ± 0.33 cd | 50.69 ± 1.74 ab |
| 1.0 MT | 39.17 ± 5.99 d | 54.19 ± 8.28 a |
| Treatment | Incidence (%) | Disease Severity Value |
|---|---|---|
| Control | 72.22 ± 0.04 c | 53.06 ± 0.04 d |
| 0.1 MT | 41.25 ± 0.02 a | 31.88 ± 0.02 a |
| 0.3 MT | 56.88 ± 0.03 b | 40.44 ± 0.01 c |
| 0.5 MT | 55.56 ± 0.04 b | 37.88 ± 0.01 b |
| 1.0 MT | 56.25 ± 0.03 b | 40.64 ± 0.01 c |
| Treatment | Longitudinal Diameter (mm) | Transverse Diameter (mm) | Lateral Diameter (mm) | Single Fruit Weight (g) | Single Fruit Volume (cm3) | Fruit Shape Index |
|---|---|---|---|---|---|---|
| Control | 65.00 ± 1.41 c | 35.55 ± 0.16 b | 41.18 ± 0.27 b | 61.52 ± 2.08 b | 56.06 ± 1.52 c | 1.83 ± 0.03 a |
| 0.1 MT | 68.30 ± 0.87 ab | 36.24 ± 0.06 ab | 42.40 ± 0.34 a | 68.61 ± 0.36 a | 61.28 ± 0.53 ab | 1.90 ± 0.01 a |
| 0.3 MT | 68.31 ± 0.57 ab | 36.65 ± 0.48 a | 42.29 ± 0.27 ab | 68.23 ± 0.65 a | 60.04 ± 0.49 b | 1.86 ± 0.05 a |
| 0.5 MT | 66.02 ± 0.50 bc | 35.42 ± 0.33 b | 42.90 ± 0.55 a | 63.64 ± 0.53 b | 58.83 ± 0.47 b | 1.86 ± 0.00 a |
| 1.0 MT | 69.80 ± 0.31 a | 36.39 ± 0.22 ab | 42.82 ± 1.39 a | 70.56 ± 0.97 a | 62.57 ± 0.62 a | 1.92 ± 0.02 a |
| Indicator | Actinidia deliciosa Guichang | |||
|---|---|---|---|---|
| Principal Component 1 | Principal Component 2 | Principal Component 3 | Principal Component 4 | |
| Solid acid ratio | 0.905 | 0.028 | 0.142 | −0.072 |
| Weight loss rate | 0.898 | 0.173 | 0.202 | 0.044 |
| Firmness | −0.891 | −0.182 | −0.12 | 0.035 |
| Total soluble solids | 0.836 | 0.353 | 0.138 | −0.097 |
| Titratable acidity | −0.783 | 0.332 | −0.042 | 0.101 |
| Disease incidence | 0.774 | −0.154 | 0.279 | −0.067 |
| Carotenoids | 0.733 | 0.109 | −0.04 | 0.278 |
| Soluble sugar | 0.67 | 0.486 | 0.177 | −0.173 |
| Chlorophyll | 0.655 | −0.159 | −0.202 | 0.244 |
| Soluble protein | −0.024 | 0.835 | 0.074 | 0.002 |
| Respiratory rate | 0.34 | 0.088 | 0.721 | −0.112 |
| Total phenols | 0.102 | 0.076 | 0.687 | 0.292 |
| Total flavonoids | −0.169 | −0.561 | 0.596 | −0.202 |
| Ascorbic acid | −0.084 | −0.134 | 0.08 | 0.765 |
| Dry matter | −0.058 | −0.132 | 0.014 | −0.647 |
| Eigenvalue | 6.21 | 1.658 | 1.372 | 1.278 |
| contribution ratio | 41.398 | 52.451 | 61.595 | 70.113 |
| Treatment | Score of Each Principal Component | Comprehensive Score | Ranking | |||
|---|---|---|---|---|---|---|
| Y1 | Y2 | Y3 | Y4 | |||
| Control | 2.813 | −0.513 | −0.357 | 0.273 | 1.100 | 3 |
| 0.1 MT | 3.877 | −0.270 | 0.293 | −1.757 | 1.453 | 2 |
| 0.3 MT | 3.400 | 2.407 | 0.053 | −1.190 | 1.577 | 1 |
| 0.5 MT | 2.817 | 1.513 | −2.497 | −0.403 | 1.073 | 4 |
| 1.0 MT | 2.580 | 1.440 | −1.837 | −0.187 | 1.040 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, J.; Zhu, S.; Lin, X.; Wan, X.; Zhang, Q.; Njie, A.; Luo, D.; Long, Y.; Fan, R.; Dong, X. Evaluation of Preharvest Melatonin on Soft Rot and Quality of Kiwifruit Based on Principal Component Analysis. Foods 2023, 12, 1414. https://doi.org/10.3390/foods12071414
Peng J, Zhu S, Lin X, Wan X, Zhang Q, Njie A, Luo D, Long Y, Fan R, Dong X. Evaluation of Preharvest Melatonin on Soft Rot and Quality of Kiwifruit Based on Principal Component Analysis. Foods. 2023; 12(7):1414. https://doi.org/10.3390/foods12071414
Chicago/Turabian StylePeng, Junsen, Shouliang Zhu, Xin Lin, Xuan Wan, Qin Zhang, Alagie Njie, Dengcan Luo, Youhua Long, Rong Fan, and Xiaoqing Dong. 2023. "Evaluation of Preharvest Melatonin on Soft Rot and Quality of Kiwifruit Based on Principal Component Analysis" Foods 12, no. 7: 1414. https://doi.org/10.3390/foods12071414