
Metabolic Variations among Three New Tea Varieties Cultivated in Shandong, China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Observation and Determination of Morphological Characteristics
2.3. Determinations of the Tea Quality Components
- m0—the weight of the tea sample, g;
- m1—the weight of the dried tea residue, g;
- w—the dry matter content of the tea sample (mass fraction), %.
- L1—the total volume of the tea infusion, mL;
- L2—the volume of the infusion taken to reaction, mL;
- M—the dry weight of the tea sample, g;
- m—the dry ratio of the tea sample, %;
- 3.914—corresponded that 1 A using the 10 mm color comparison cell was equal to 3.914 mg of the tea polyphenols in the tea infusion.
- C—the amino acids weight (mg), which could be obtained according to the OD570 from a standard curve made by theanine or glutamic acid as a standard component, using the same method as mentioned above;
- V1—the total volume of the tea infusion, mL;
- V2—the volume of the infusion taken to reaction, mL;
- M—the dry weight of the tea sample, g;
- w—the dry ratio of the tea sample, %.
- C2—the caffeine content (caffeine mg/mL), which could be obtained according to the OD274 from a caffeine standard curve made by caffeine as standard component, using the same method as mentioned above;
- L3—the total volume of the tea infusion, mL;
- M2—the dry weight of the tea sample, g;
- m2—the dry ratio of the tea sample, %.
2.4. Detection of Metabolites Using UPLC-ESI-MS/MS
2.5. Statistical Analyses
2.6. Sensory Evaluation of the Tea Samples
3. Results
3.1. Agronomic Characteristics of the Lucha Varieties
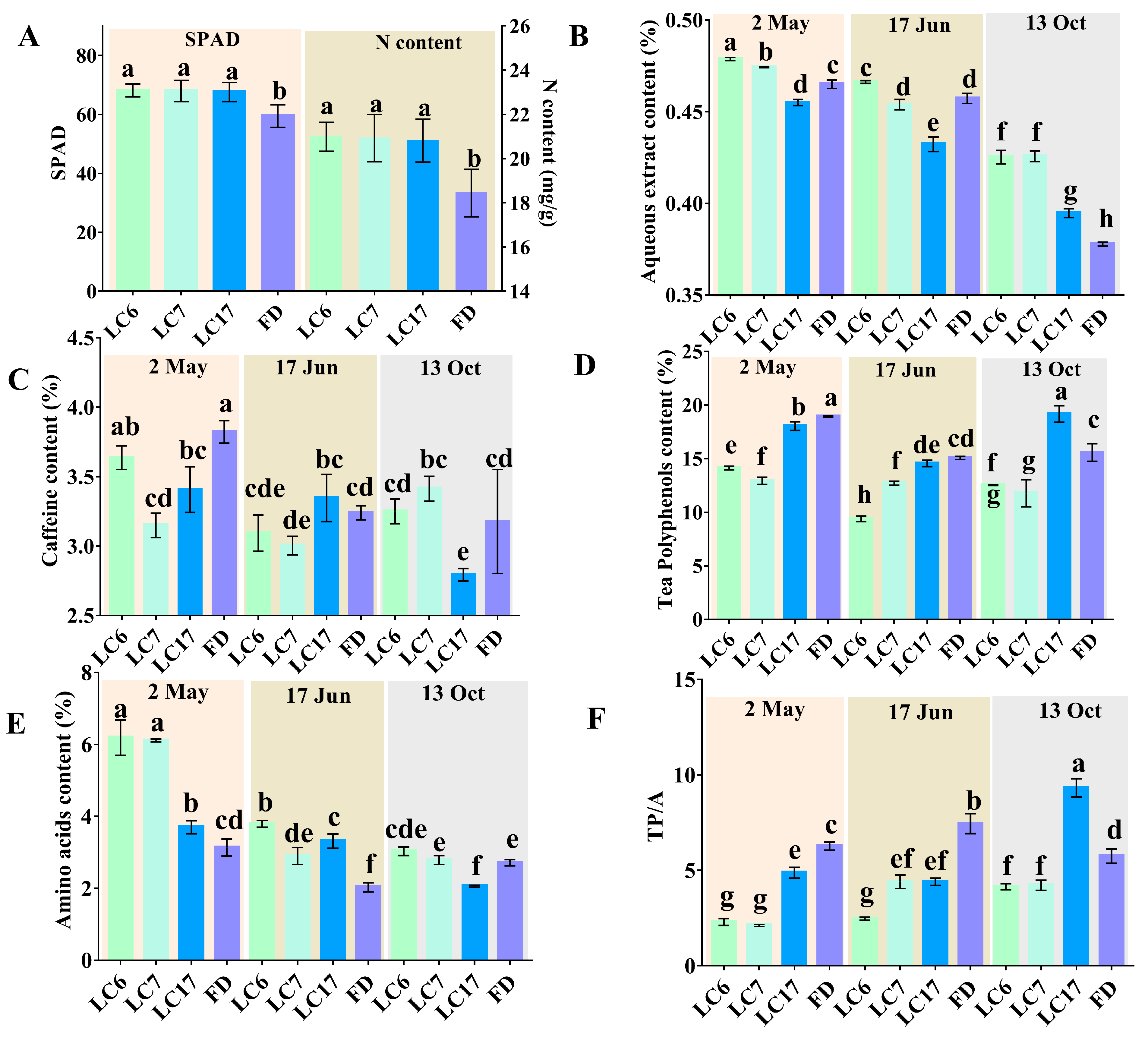
3.2. Biochemical Components in Different Varieties
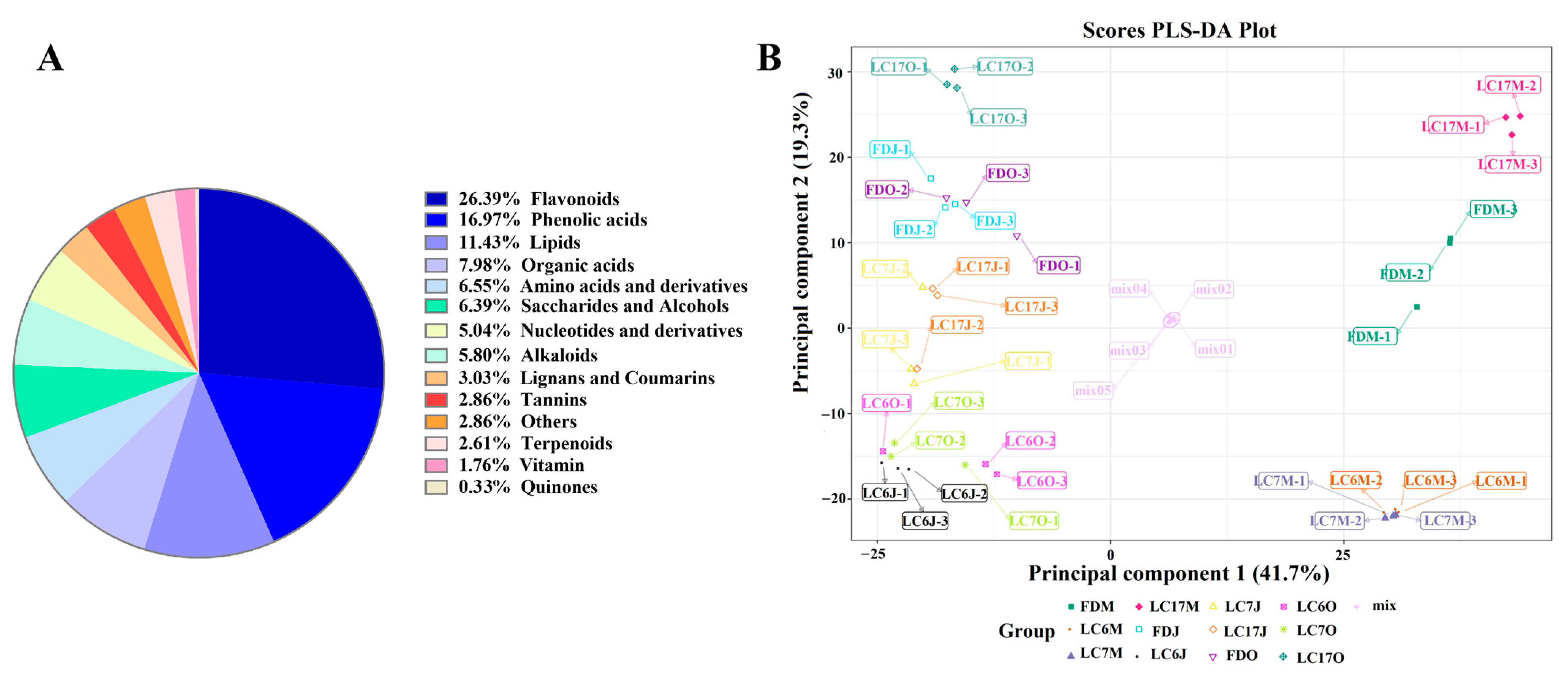
3.3. Metabolomic Analysis of UPLC-MS/MS Data
3.4. Metabolomic Variations among the Tea Varieties in Different Seasons
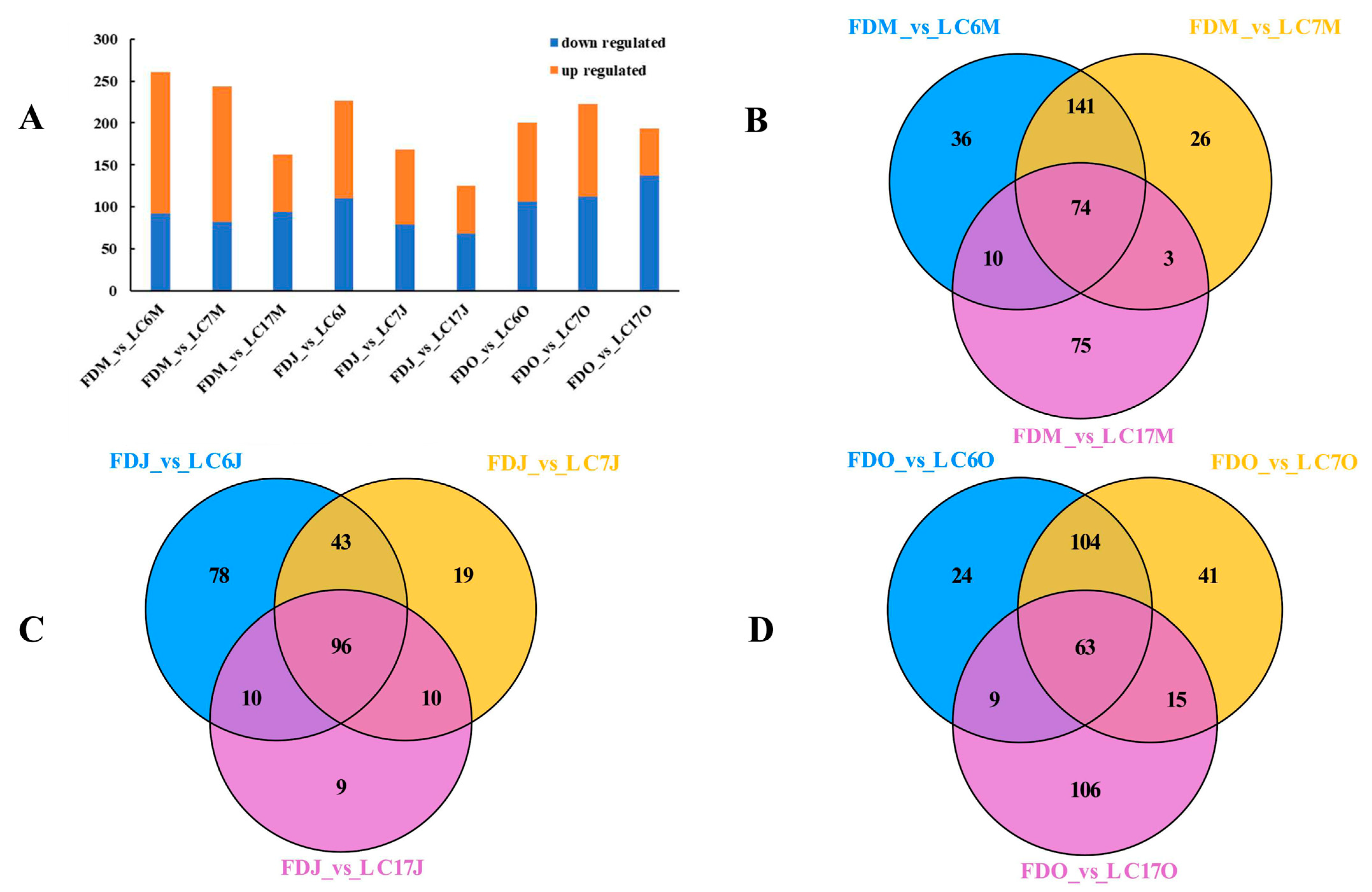
3.4.1. Identification of the Significantly Differential Metabolites
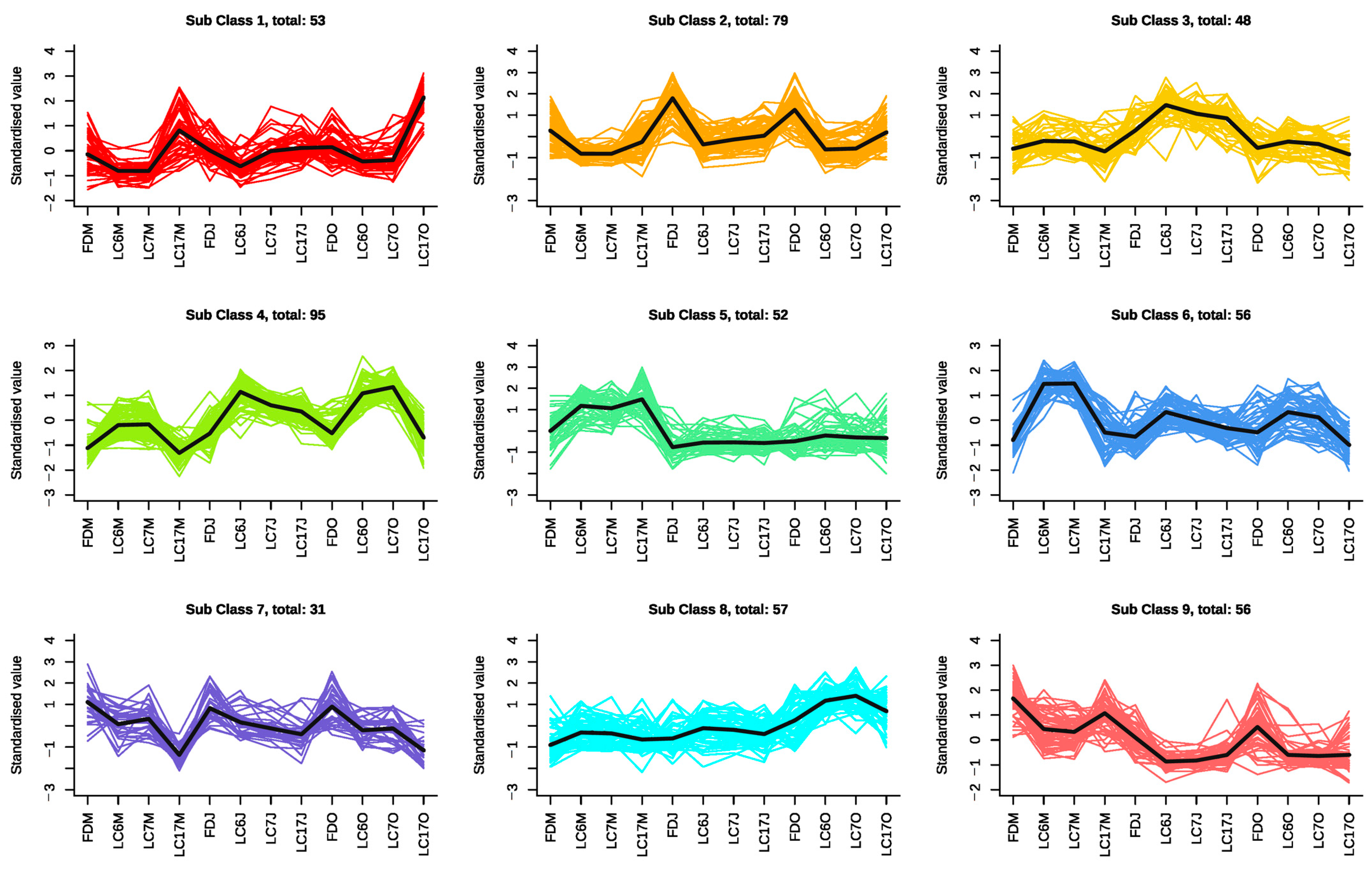
3.4.2. K-Means Clustering Analysis of the Significantly Differential Metabolites
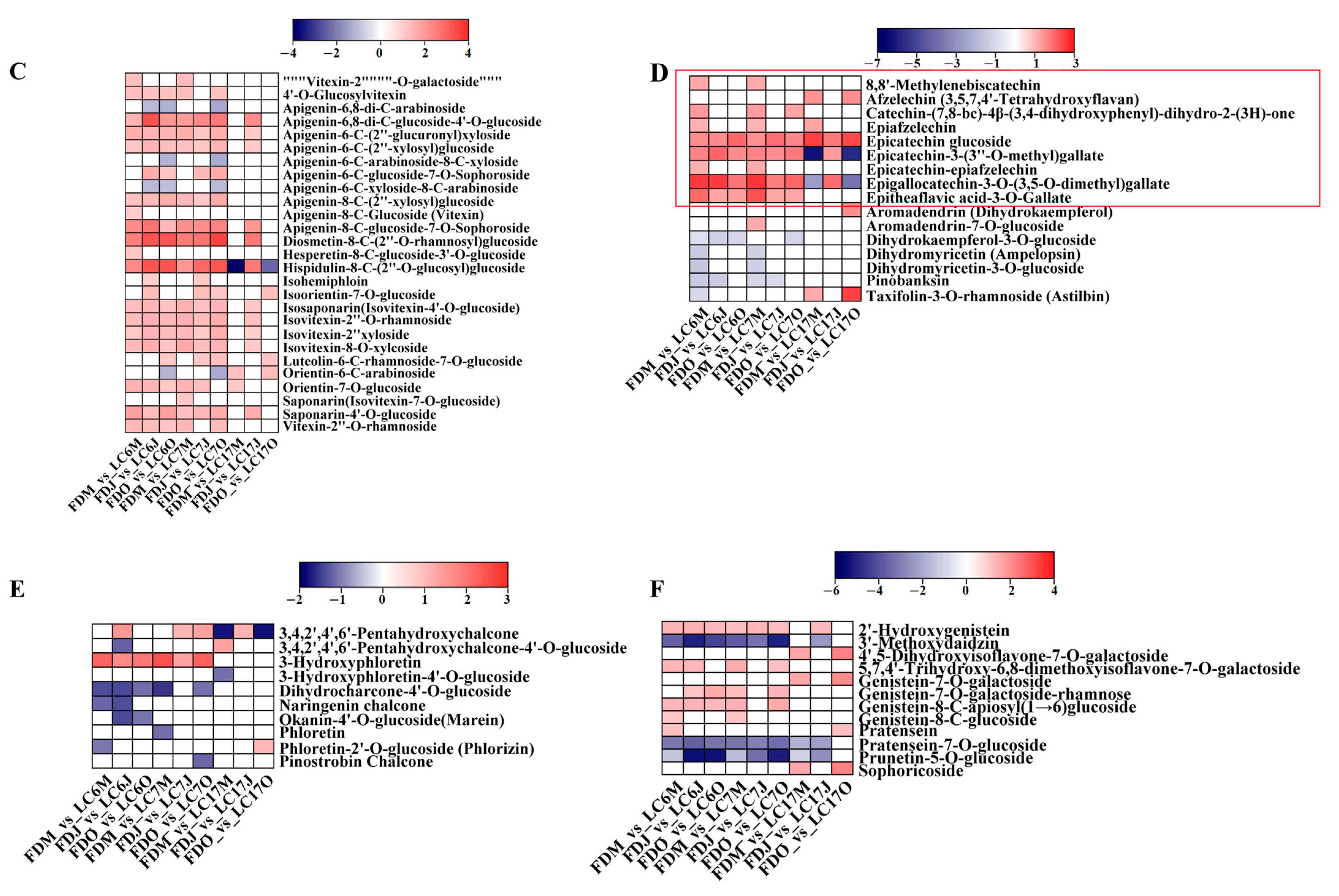
3.4.3. Variation in Flavonoids between the Lucha Varieties and FD in Different Seasons
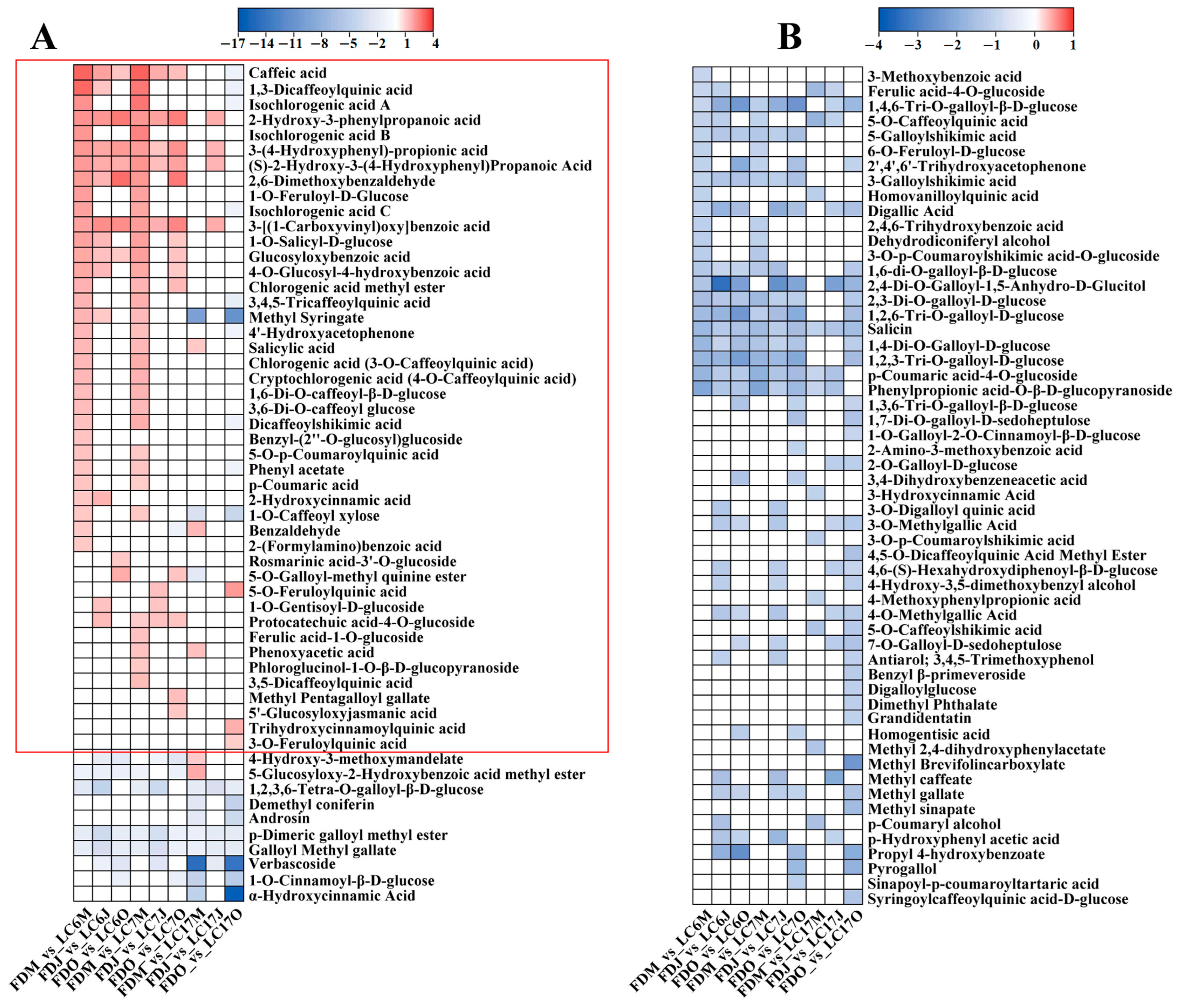
3.4.4. Variation in Phenolic Acids between the Lucha Varieties and FD in Different Seasons
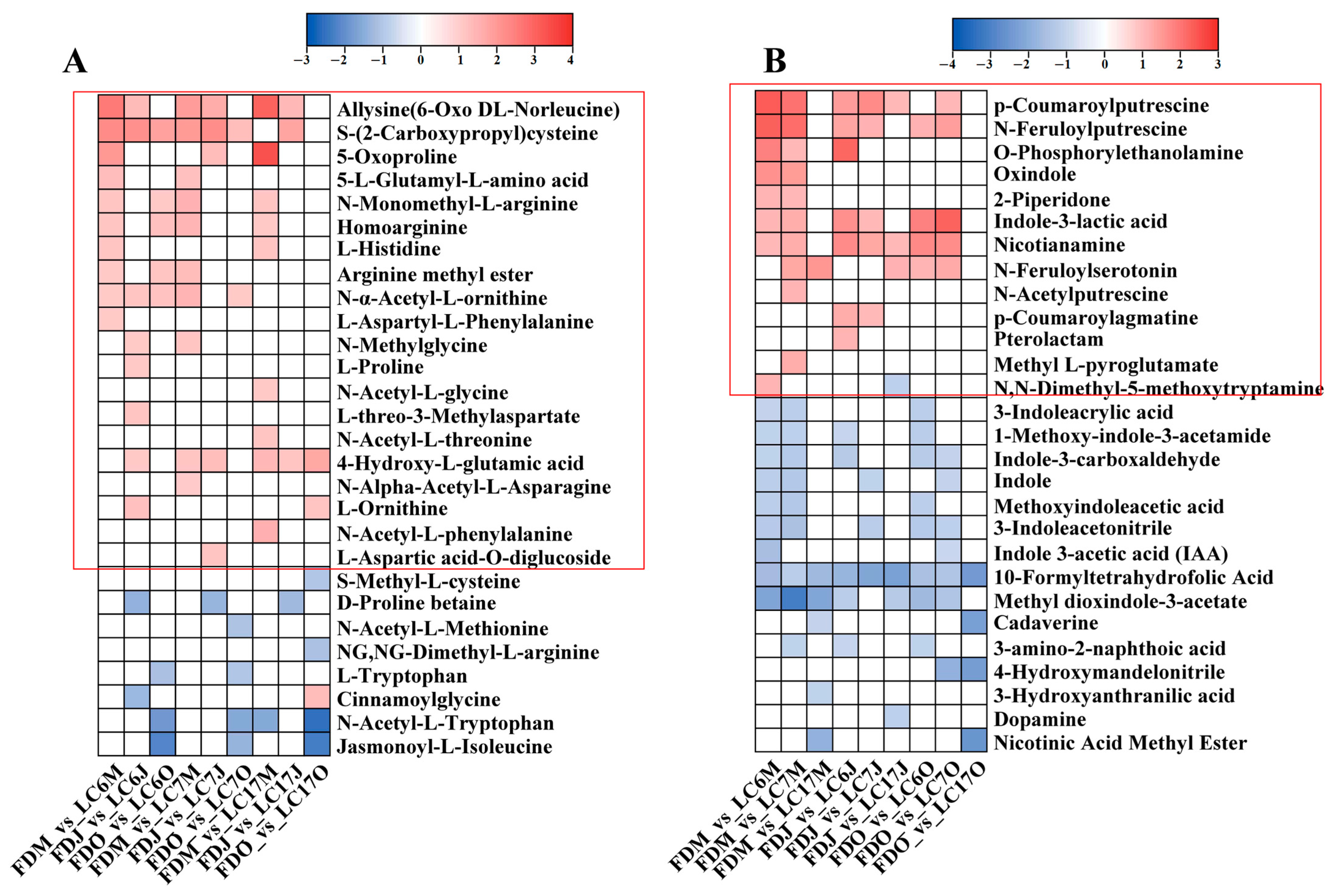
3.4.5. Variation in Amino Acids and Alkaloids between the Lucha Varieties and FD in Different Seasons
3.5. Sensory Evaluation of the Green Teas Made by LC6, LC7, LC17, and FD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liang, Y.; Shi, M. Advances in tea plant genetics and breeding. J. Tea Sci. 2015, 35, 103–109. [Google Scholar]
- Wang, X.; Feng, H.; Chang, Y.; Ma, C.; Wang, L.; Hao, X.; Li, A.l.; Cheng, H.; Wang, L.; Cui, P.; et al. Population sequencing enhances understanding of tea plant evolution. Nat. Commun. 2020, 11, 4447. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Gao, M.J.; Hou, R.Y.; Hu, X.Y.; Zhang, L.; Wan, X.C.; Wei, S. Determination of quality constituents in the young leaves of albino tea cultivars. Food Chem. 2014, 155, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, C.; Li, Y.; Ding, Z.; Shen, J.; Wang, Y.; Zhao, L.; Xu, M. The identification and evaluation of two different color variations of tea. J. Sci. Food Agric. 2016, 96, 4951–4961. [Google Scholar] [CrossRef] [PubMed]
- Wambulwa, M.C.; Meegahakumbura, M.K.; Kamunya, S.; Muchugi, A.; Möller, M.; Liu, J.; Xu, J.C.; Ranjitkar, S.; Li, D.Z.; Gao, L.M. Insights into the Genetic Relationships and Breeding Patterns of the African Tea Germplasm Based on nSSR Markers and cpDNA Sequences. Front. Plant Sci. 2016, 7, 1244. [Google Scholar] [CrossRef] [Green Version]
- Chaeikar, S.S.; Falakro, K.; Rahimi, M.; Khiavi, S.J.; Ashourpour, M. The investigation of genetic diversity based on SCoT markers, morphological, and chemical characters in tea (Camellia sinensis L.) clones. J. Hortic. Postharvest Res. 2020, 3, 269–284. [Google Scholar]
- Gai, Z.; Wang, Y.; Jiang, J.; Xie, H.; Ding, Z.; Ding, S.; Wang, H. The quality evaluation of tea (Camellia sinensis) varieties based on the metabolomics. HortScience 2019, 54, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Kilel, E.C.; Faraj, A.K.; Wanyoko, J.K.; Wachira, F.N.; Mwingirwa, V. Green tea from purple leaf coloured tea clones in Kenya- their quality characteristics. Food Chem. 2013, 141, 769–775. [Google Scholar] [CrossRef]
- Chen, L.; Yamaguchi, S. RAPD markers for discriminating tea germplasms at the inter-specific level in China. Plant Breed. 2005, 124, 404–409. [Google Scholar] [CrossRef]
- Liu, B.; Sun, X.; Wang, Y.-h.O.; Li, Y.; Cheng, H.; Xiong, C.; Wang, P.-S. Genetic diversity and molecular discrimination of wild tea plants from Yunnan Province based on inter-simple sequence repeats (ISSR) markers. Afr. J. Biotechnol. 2012, 11, 11566–11574. [Google Scholar]
- Jahangirzadeh, S.; Gonbad, R.A.; Falakro, K. Identification of genetic diversity and relationships of some Iranian tea genotypes using SRAP markers. J. Hortic. Postharvest Res. 2020, 3, 25–34. [Google Scholar]
- Hamany Djande, C.Y.; Piater, L.A.; Steenkamp, P.A.; Tugizimana, F.; Dubery, I.A. A Metabolomics Approach and Chemometric Tools for Differentiation of Barley Cultivars and Biomarker Discovery. Metabolites 2021, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Gienapp, P.; Laine, V.N.; Mateman, A.C.; van Oers, K.; Visser, M.E. Environment-dependent genotype-phenotype associations in avian breeding time. Front. Genet. 2017, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Hamany Djande, C.Y.; Pretorius, C.; Tugizimana, F.; Piater, L.A.; Dubery, I.A. Metabolomics: A tool for cultivar phenotyping and investigation of grain crops. Agronomy 2020, 10, 831. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Z.; Li, Y.; Li, Z.; Liu, H.; Zhou, W. Metabolite profiling of sorghum seeds of different colors from different sweet sorghum cultivars using a widely targeted metabolomics approach. Int. J. Genom. 2020, 2020, 6247429. [Google Scholar] [CrossRef]
- Pretorius, C.J.; Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A. Metabolomics for biomarker discovery: Key signatory metabolic profiles for the identification and discrimination of oat cultivars. Metabolites 2021, 11, 165. [Google Scholar] [CrossRef] [PubMed]
- Lawas, L.M.F.; Li, X.; Erban, A.; Kopka, J.; Jagadish, S.K.; Zuther, E.; Hincha, D.K. Metabolic responses of rice cultivars with different tolerance to combined drought and heat stress under field conditions. GigaScience 2019, 8, giz050. [Google Scholar] [CrossRef]
- Chen, S.; Li, M.; Zheng, G.; Wang, T.; Lin, J.; Wang, S.; Wang, X.; Chao, Q.; Cao, S.; Yang, Z. Metabolite profiling of 14 Wuyi Rock tea cultivars using UPLC-QTOF MS and UPLC-QQQ MS combined with chemometrics. Molecules 2018, 23, 104. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wang, P.; Chen, X.; Yue, C.; Guo, Y.; Yang, J.; Sun, Y.; Ye, N. Integrated transcriptomics and metabolomics provide novel insight into changes in specialized metabolites in an albino tea cultivar (Camellia sinensis (L.) O. Kuntz). Plant Physiol. Biochem. 2021, 160, 27–36. [Google Scholar] [CrossRef]
- Lee, J.-E.; Lee, B.-J.; Chung, J.-O.; Kim, H.-N.; Kim, E.-H.; Jung, S.; Lee, H.; Lee, S.-J.; Hong, Y.-S. Metabolomic unveiling of a diverse range of green tea (Camellia sinensis) metabolites dependent on geography. Food Chem. 2015, 174, 452–459. [Google Scholar] [CrossRef]
- Lee, L.-S.; Choi, J.H.; Son, N.; Kim, S.-H.; Park, J.-D.; Jang, D.-J.; Jeong, Y.; Kim, H.-J. Metabolomic analysis of the effect of shade treatment on the nutritional and sensory qualities of green tea. J. Agric. Land Food Chem. 2013, 61, 332–338. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Yan, C.; Fang, M.; Wang, L.; Huang, Y.; Wang, F. Metabolome and Microbiome Signatures in the Leaves of Wild Tea Plant Resources Resistant to Pestalotiopsis theae. Front. Microbiol. 2022, 13, 907962. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Lu, Q.; Zhou, X.; Li, J.; Yue, J.; Wang, Z.; Pan, S. Metabolic variations of organic acids, amino acids, fatty acids and aroma compounds in the pulp of different pummelo varieties. LWT-Food Sci. Technol. 2020, 130, 109445. [Google Scholar] [CrossRef]
- Bi, W.; Zhao, G.; Zhou, Y.; Xia, X.; Wang, J.; Wang, G.; Lu, S.; He, W.; Bi, T.; Li, J. Metabonomics analysis of flavonoids in seeds and sprouts of two Chinese soybean cultivars. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-Q.; Ji, W.-B.; Yu, P.; Chen, J.-X.; Wang, F.; Yin, J.-F. Effect of extraction methods on the chemical components and taste quality of green tea extract. Food Chem. 2018, 248, 146–154. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, Z.-X. Variations of main quality components of tea genetic resources [Camellia sinensis (L.) O. Kuntze] preserved in the China National Germplasm Tea Repository. Plant Foods Hum. Nutr. 2005, 60, 31–35. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A Novel Integrated Method for Large-Scale Detection, Identification, and Quantification of Widely Targeted Metabolites: Application in the Study of Rice Metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Jewison, T.; Guo, A.C.; Wilson, M.; Knox, C.; Liu, Y.; Djoumbou, Y.; Mandal, R.; Aziat, F.; Dong, E.; et al. HMDB 3.0—The Human Metabolome Database in 2013. Nucleic Acids Res. 2013, 41, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.-J.; Schultz, A.W.; Wang, J.; Johnson, C.H.; Yannone, S.M.; Patti, G.J.; Siuzdak, G. Liquid chromatography quadrupole time-of-flight mass spectrometry characterization of metabolites guided by the METLIN database. Nat. Protoc. 2013, 8, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Chong, J.; Xia, J. MetaboAnalystR: An R package for flexible and reproducible analysis of metabolomics data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.; Zhang, M.; Fang, X.; Tang, L.; Wang, Z.; Shi, F. Analysis of the Fruit Quality of Pear (Pyrus spp.) Using Widely Targeted Metabolomics. Foods 2022, 11, 1440. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Qi, D.; Yang, T.; Lv, H.; Guo, L.; Zhang, Y.; Zhu, Y.; Peng, Q.; Xie, D.; Tan, J. Nontargeted analysis using ultraperformance liquid chromatography–quadrupole time-of-flight mass spectrometry uncovers the effects of harvest season on the metabolites and taste quality of tea (Camellia sinensis L.). J. Agric. Food Chem. 2015, 63, 9869–9878. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Shi, Y.; Zhang, Q.; Wang, J.; Ruan, J. Prediction of Chinese green tea ranking by metabolite profiling using ultra-performance liquid chromatography–quadrupole time-of-flight mass spectrometry (UPLC–Q-TOF/MS). Food Chem. 2017, 221, 311–316. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Brunetti, C.; George, R.M.; Tattini, M.; Field, K.; Davey, M.P. Metabolomics in plant environmental physiology. J. Exp. Bot. 2013, 64, 4011–4020. [Google Scholar] [CrossRef] [Green Version]
- Ibarra-Estrada, E.; Soto-Hernández, R.M.; Palma-Tenango, M. Metabolomics as a Tool in Agriculture. In Metabolomics-Fundamentals and Applications; Prasain, J.K., Ed.; INTECH: London, UK, 2016; pp. 148–168. [Google Scholar]
- Peng, B.; Li, H.; Peng, X.-X. Functional metabolomics: From biomarker discovery to metabolome reprogramming. Protein Cell 2015, 6, 628–637. [Google Scholar] [CrossRef] [Green Version]
- Steinfath, M.; Strehmel, N.; Peters, R.; Schauer, N.; Groth, D.; Hummel, J.; Steup, M.; Selbig, J.; Kopka, J.; Geigenberger, P. Discovering plant metabolic biomarkers for phenotype prediction using an untargeted approach. Plant Biotechnol. J. 2010, 8, 900–911. [Google Scholar] [CrossRef]
- Fang, Z.-T.; Yang, W.-T.; Li, C.-Y.; Li, D.; Dong, J.-J.; Zhao, D.; Xu, H.-R.; Ye, J.-H.; Zheng, X.-Q.; Liang, Y.-R. Accumulation pattern of catechins and flavonol glycosides in different varieties and cultivars of tea plant in China. J. Food Compos. Anal. 2021, 97, 103772. [Google Scholar] [CrossRef]
- Ali, G.; Neda, G. Flavonoids and phenolic acids: Role and biochemical activity in plants and human. J. Med. Plants Res. 2011, 5, 6697–6703. [Google Scholar]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Cervantes-Hernández, F.; Ochoa-Alejo, N.; Martínez, O.; Ordaz-Ortiz, J.J. Metabolomic Analysis Identifies Differences Between Wild and Domesticated Chili Pepper Fruits During Development (Capsicum annuum L.). Front. Plant Sci. 2022, 13, 893055. [Google Scholar] [CrossRef]
- Zhuang, J.; Dai, X.; Zhu, M.; Zhang, S.; Dai, Q.; Jiang, X.; Liu, Y.; Gao, L.; Xia, T. Evaluation of astringent taste of green tea through mass spectrometry-based targeted metabolic profiling of polyphenols. Food Chem. 2020, 305, 125507. [Google Scholar] [CrossRef]
- Yang, C.; Hu, Z.; Lu, M.; Li, P.; Tan, J.; Chen, M.; Lv, H.; Zhu, Y.; Zhang, Y.; Guo, L. Application of metabolomics profiling in the analysis of metabolites and taste quality in different subtypes of white tea. Food Res. Int. 2018, 106, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Tomás-Barberán, F.A.; Espín, J.C. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. J. Sci. Food Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Liao, Y.; Zhou, X.; Zeng, L. How does tea (Camellia sinensis) produce specialized metabolites which determine its unique quality and function: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 3751–3767. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, M.-H.; Deng, W.-W.; Ahammed, G.J.; Wei, J.-P.; Yan, P.; Zhang, L.-P.; Fu, J.-Y.; Han, W.-Y. Exogenous melatonin improves tea quality under moderate high temperatures by increasing epigallocatechin-3-gallate and theanine biosynthesis in Camellia sinensis L. J. Plant Physiol. 2020, 253, 153273. [Google Scholar] [CrossRef] [PubMed]
- Han, W.-Y.; Huang, J.-G.; Li, X.; Li, Z.-X.; Ahammed, G.J.; Yan, P.; Stepp, J.R. Altitudinal effects on the quality of green tea in east China: A climate change perspective. Eur. Food Res. Technol. 2017, 243, 323–330. [Google Scholar] [CrossRef]
- Hillmann, H.; Hofmann, T. Quantitation of key tastants and re-engineering the taste of Parmesan cheese. J. Agric. Food Chem. 2016, 64, 1794–1805. [Google Scholar] [CrossRef]
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations–A review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef]
- Ma, Q.; Qin, M.; Song, L.; Sun, H.; Zhang, H.; Wu, H.; Ren, Z.; Liu, H.; Duan, G.; Wang, Y.; et al. Molecular Link in Flavonoid and Amino Acid Biosynthesis Contributes to the Flavor of Changqing Tea in Different Seasons. Foods 2022, 11, 2289. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensory Indexes (SIs) | Spring (2 May) | Summer (17 June) | Autumn (13 October) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (SI%) | LC6M | LC7M | LC17M | FDM | LC6J | LC7J | LC17J | FDJ | LC6O | LC7O | LC17O | FDO |
| Appearance (25%) | 92.83 ± 0.16 a | 92.33 ± 0.33 ab | 91.5 ± 0.28 abc | 92 ± 0.57 ab | 90.17 ± 0.16 c | 91.83 ± 0.16 ab | 92.5 ± 0.28 ab | 92.17 ± 0.60 ab | 90.5 ± 0.5 cd | 92.5 ± 0.28 ab | 90.5 ± 0.76 cd | 91.17 ± 0.16 bcd |
| Aroma (25%) | 90.5 ± 0.28 de | 90.33 ± 0.33 e | 90.83 ± 0.44 de | 91.33 ± 0.33 cd | 93.17 ± 0.16 a | 92.17 ± 0.16 bc | 91.83 ± 0.16 bc | 90.5 ± 0.28 de | 90.33 ± 0.33 e | 92.33 ± 0.33 ab | 92 ± 0 bc | 93.17 ± 0.16 a |
| Infusion color (10%) | 90.5 ± 1.04 d | 92 ± 0.28 bcd | 91 ± 0.57 d | 91.17 ± 0.72 cd | 91.17 ± 0.16 cd | 91 ± 0.28 d | 92.83 ± 0.16 abc | 91.33 ± 0.72 cd | 94 ± 0.57 a | 93.5 ± 0.28 ab | 90.67 ± 0.33 d | 91 ± 0.57 d |
| Infusion taste (30%) | 92.33 ± 0.33 a | 92.33 ± 0.33 a | 90.67 ± 0.33 bc | 91.33 ± 0.33 b | 90.33 ± 0.33 cd | 89.67 ± 0.33 d | 92.33 ± 0.33 a | 90.33 ± 0.33 d | 92.5 ± 0.28 a | 92.5 ± 0.28 a | 92.67 ± 0.16 a | 92.67 ± 0.16 a |
| Infused leaf (10%) | 92.67 ± 0.33 a | 92.33 ± 0.33 a | 92.33 ± 0.33 a | 91.83 ± 0.44 abc | 92 ± 0 ab | 91.33 ± 0.33 bcd | 90.5 ± 0.28 de | 91 ± 0 cde | 92.17 ± 0.16 ab | 92.5 ± 0.28 a | 90.33 ± 0.33 e | 90.67 ± 0.33 de |
| Overall | 91.85 | 91.80 | 91.12 | 91.53 | 91.25 | 91.13 | 92.11 | 91.00 | 91.57 | 92.56 | 91.53 | 92.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, J.; Wang, H.; Sun, L.; Fan, K.; Zhang, X.; Huang, Q.; Ding, S.; Wang, Y.; Ding, Z. Metabolic Variations among Three New Tea Varieties Cultivated in Shandong, China. Foods 2023, 12, 1299. https://doi.org/10.3390/foods12061299
Shen J, Wang H, Sun L, Fan K, Zhang X, Huang Q, Ding S, Wang Y, Ding Z. Metabolic Variations among Three New Tea Varieties Cultivated in Shandong, China. Foods. 2023; 12(6):1299. https://doi.org/10.3390/foods12061299
Chicago/Turabian StyleShen, Jiazhi, Hui Wang, Litao Sun, Kai Fan, Xifa Zhang, Qingfu Huang, Shibo Ding, Yu Wang, and Zhaotang Ding. 2023. "Metabolic Variations among Three New Tea Varieties Cultivated in Shandong, China" Foods 12, no. 6: 1299. https://doi.org/10.3390/foods12061299