1-Methylcyclopropene and UV-C Treatment Effect on Storage Quality and Antioxidant Activity of ‘Xiaobai’ Apricot Fruit

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
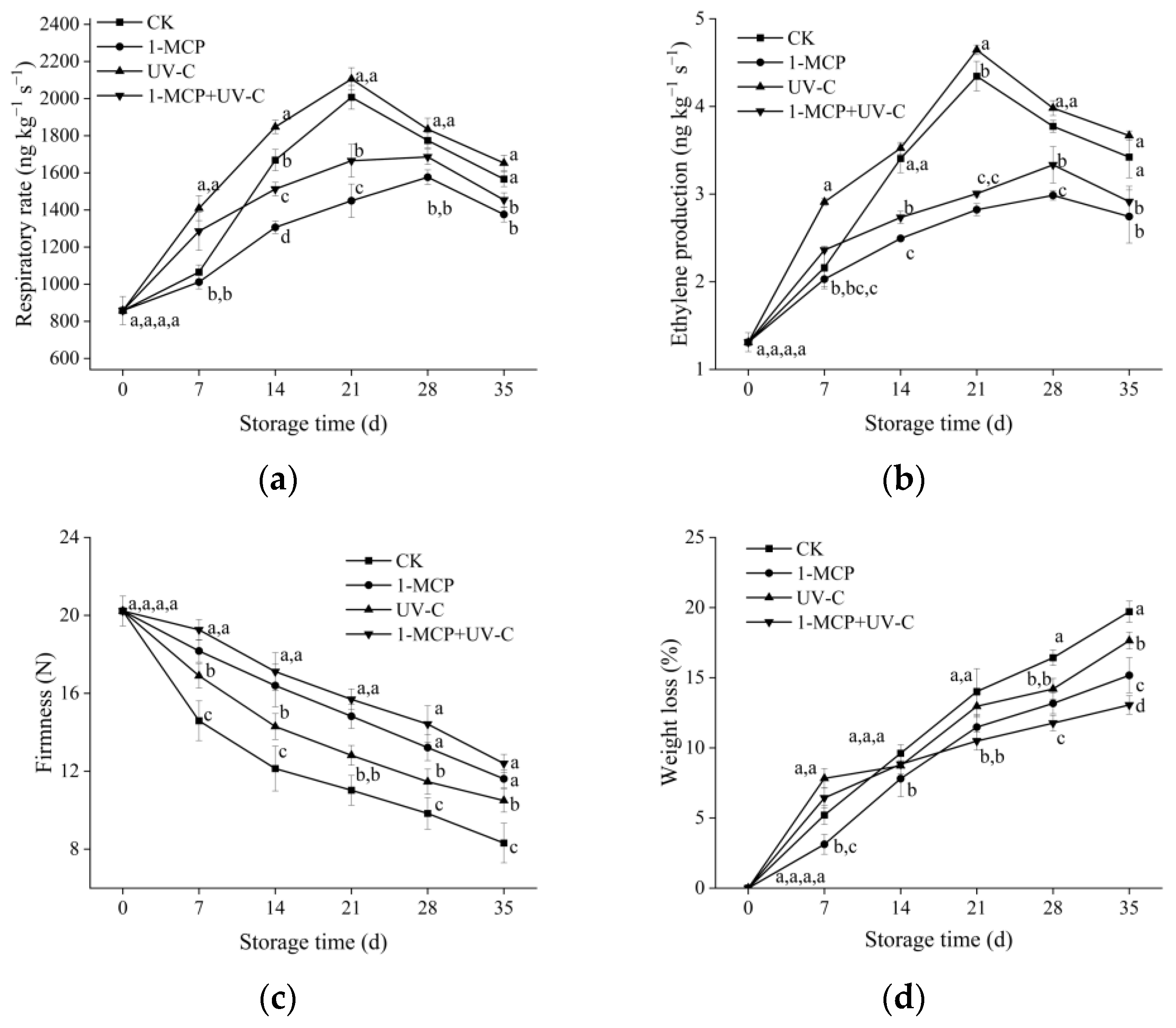
2.2. Determination of Ethylene Production and Respiration Rate
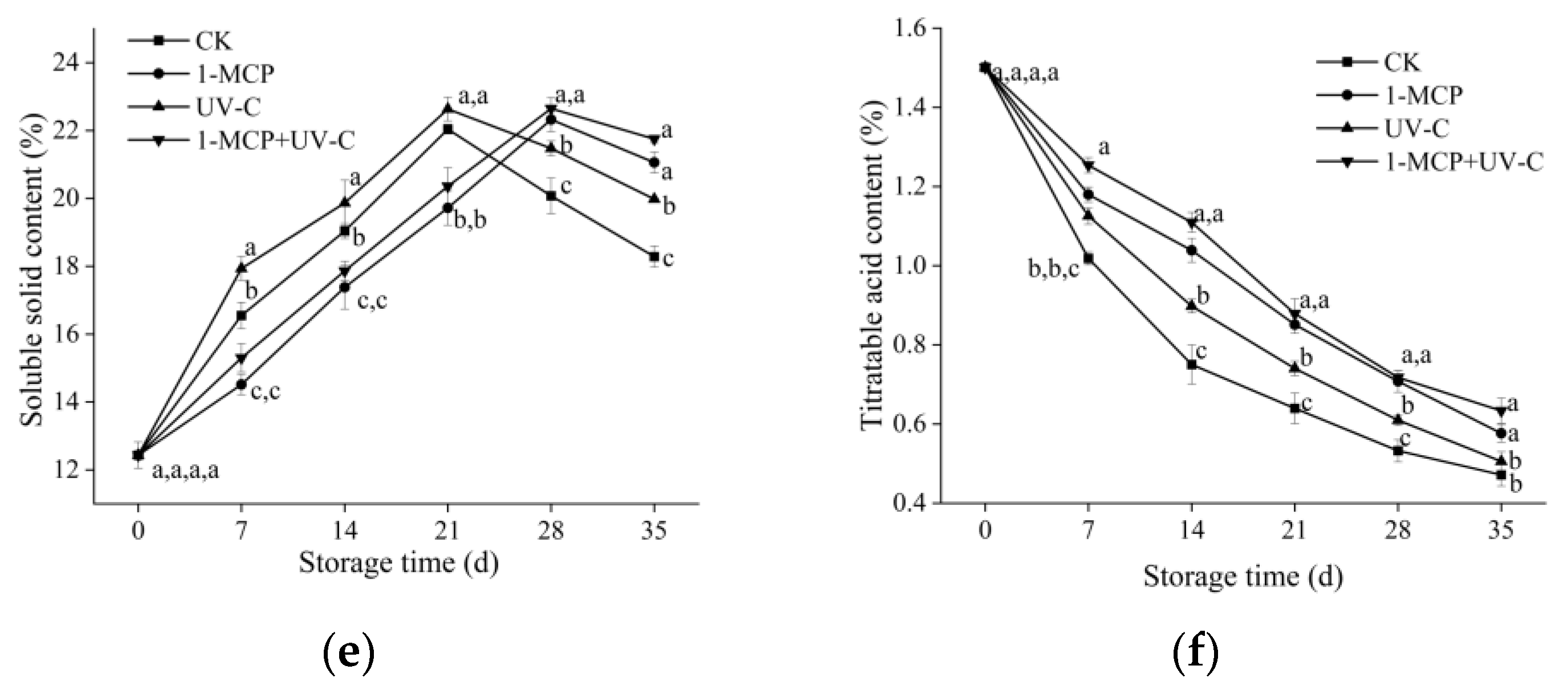
2.3. Firmness, Weight Loss, Soluble Solid Content and Titratable Acid
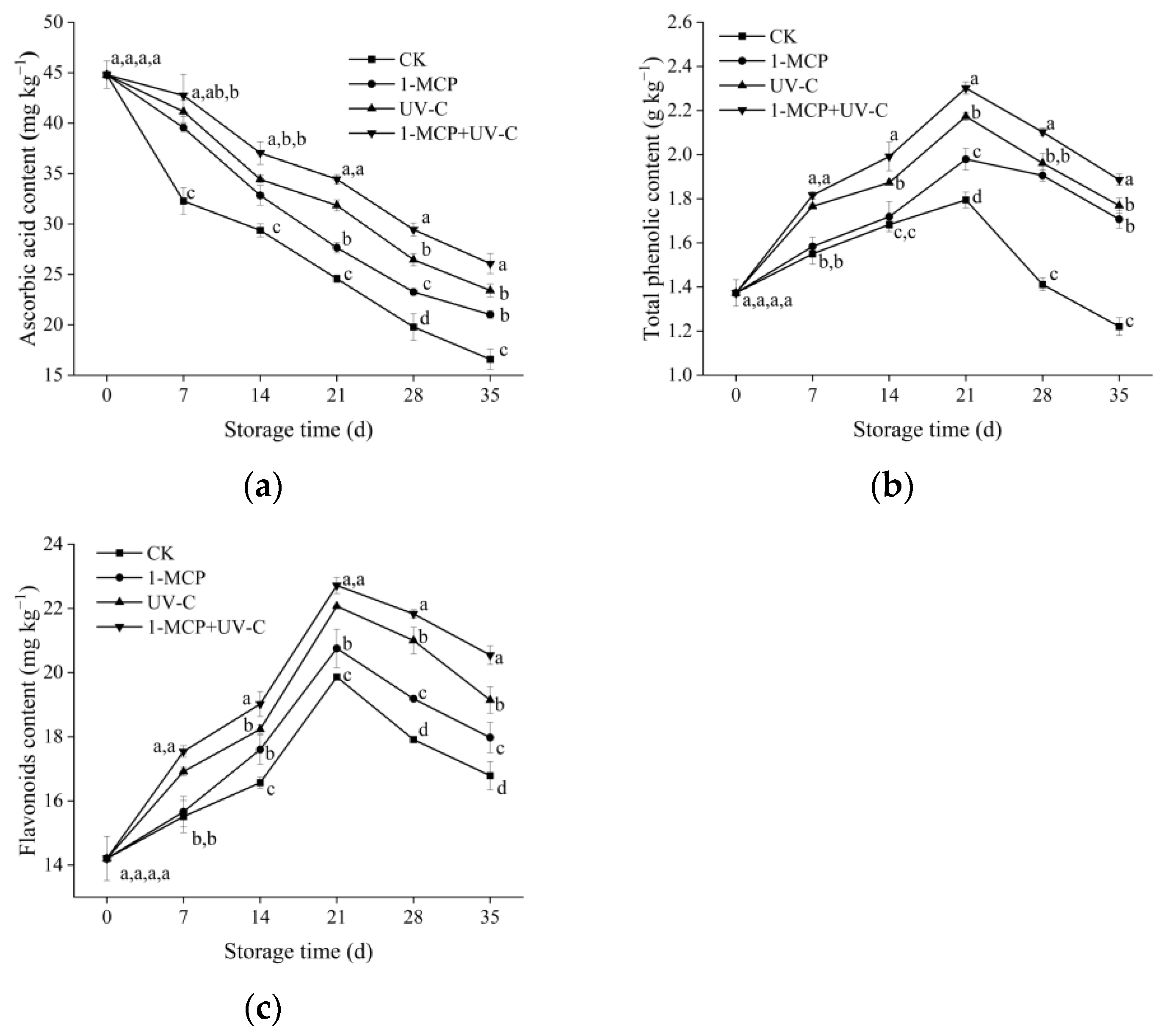
2.4. Ascorbic Acid, Total Phenolics, and Flavonoid Content
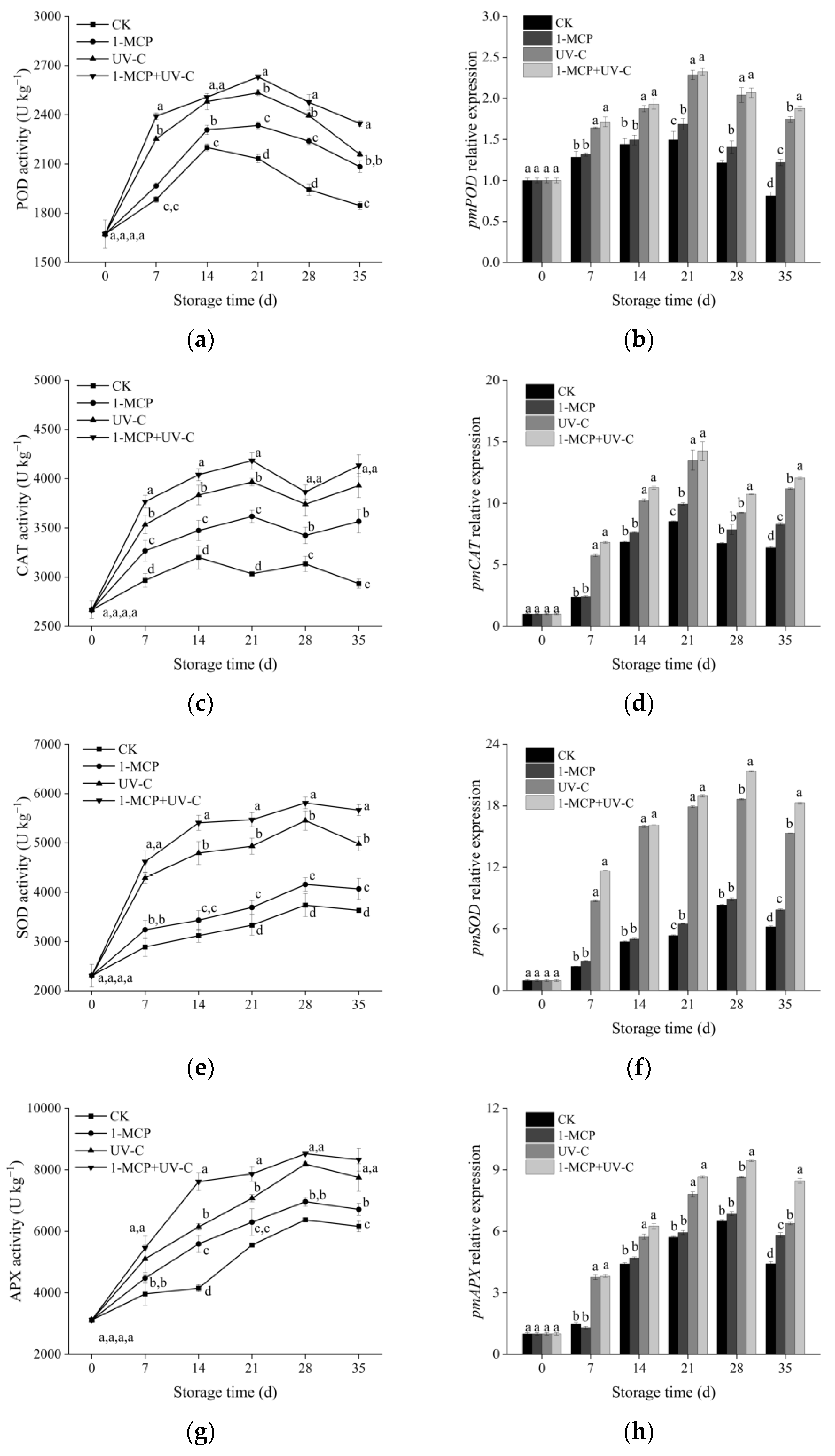
2.5. Peroxidase, Ascorbate Peroxidase, Superoxide Dismutase and Catalase Activities
2.6. Malondialdehyde, Hydrogen Peroxide Content, and Superoxide Radicals Generation Rate
2.7. 2,2-Diphenyl-1-Picrylhydrazyl Radical Scavenging Activity Assay
2.8. Reverse Transcription–Quantitative PCR (RT-qPCR)
2.9. Statistical Analysis
3. Results
3.1. Respiration Rate and Ethylene Production
3.2. Firmness and Weight Loss
3.3. SSC and TA
3.4. ASA, TP, and Flavonoid Content
3.5. POD, CAT, SOD, and APX Activities and Corresponding Relative Gene Expression
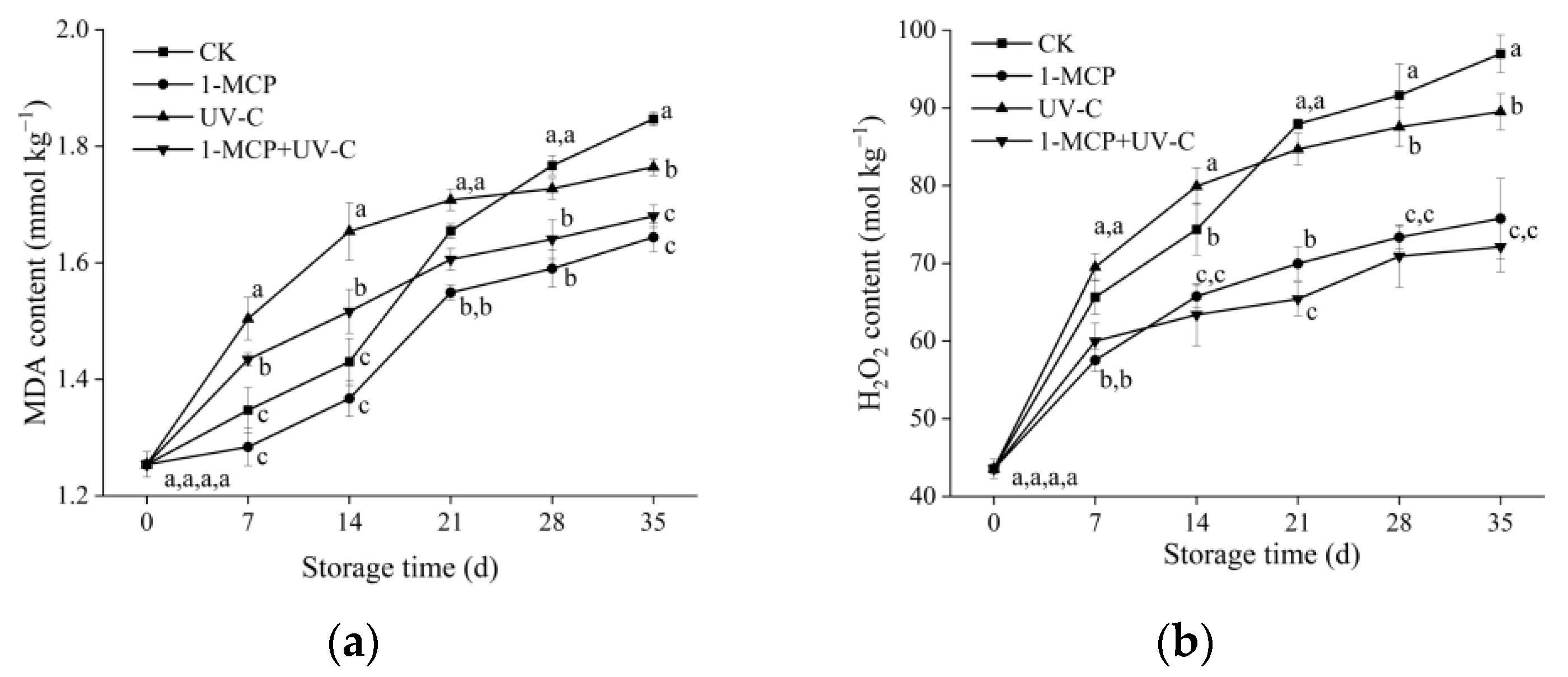
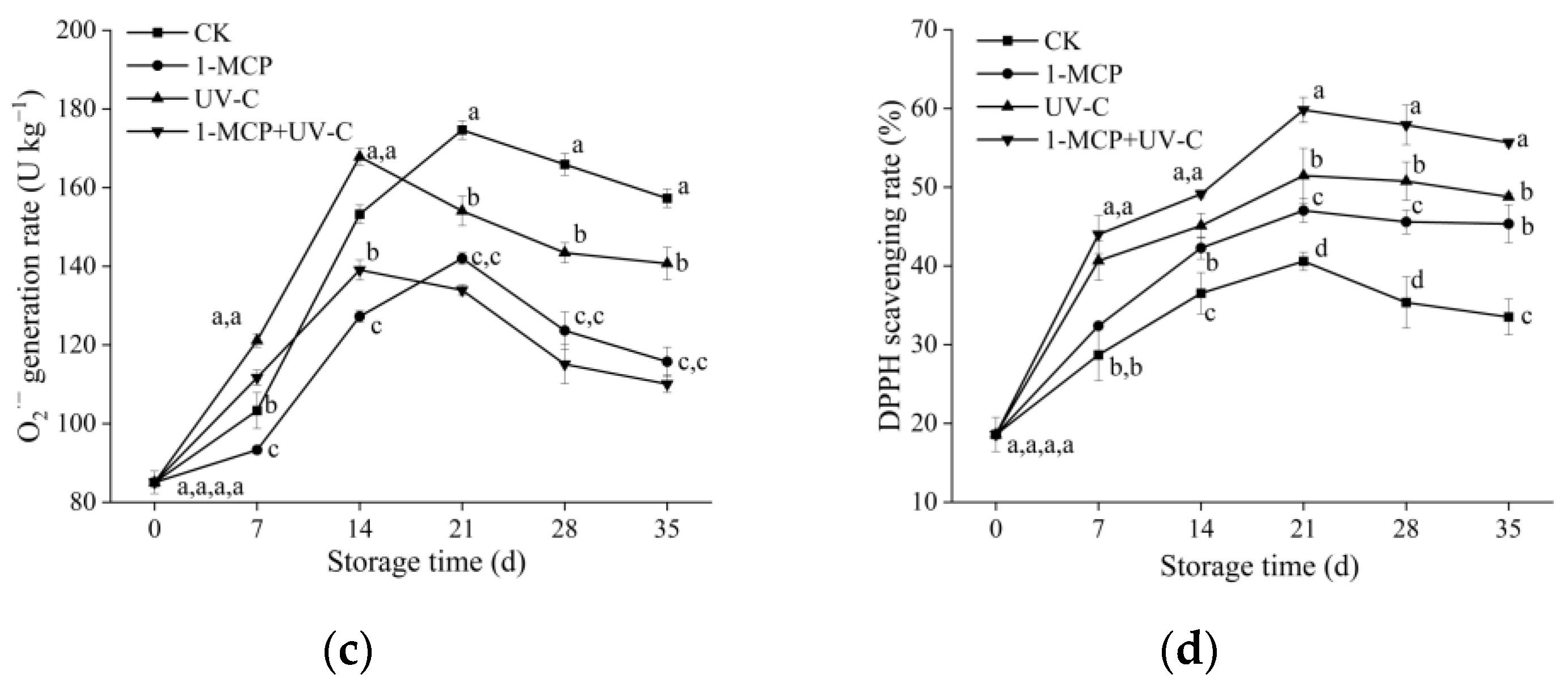
3.6. MDA, H2O2 Content, and O2.− Generation Rate
3.7. DPPH Scavenging Rate
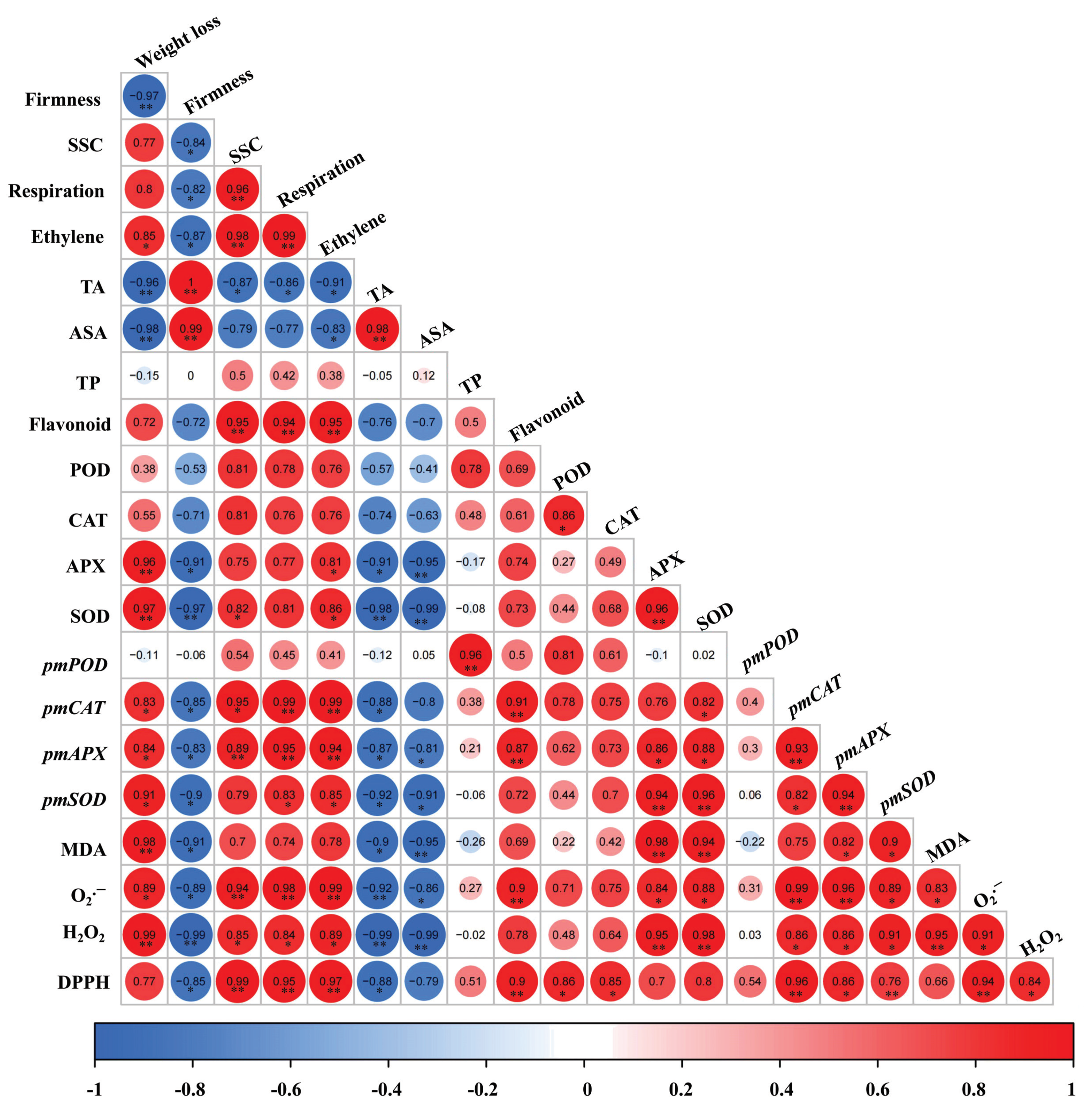
3.8. Correlation Analysis
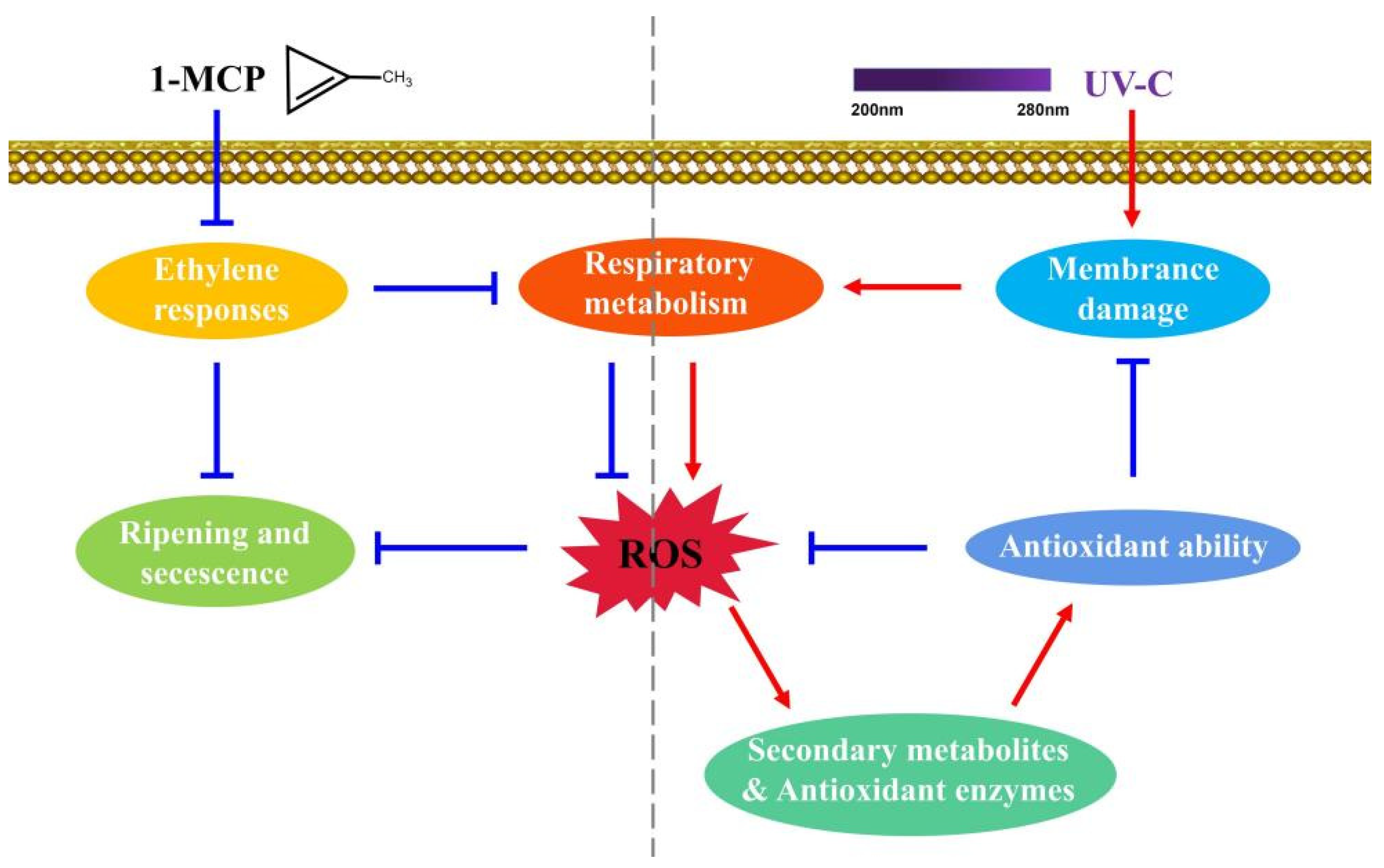
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cui, K.; Zhao, H.; Sun, L.; Yang, L.; Cao, J.; Jiang, W. Impact of near freezing temperature storage on postharvest quality and antioxidant capacity of two apricot (Prunus armeniaca L.) cultivars. J. Food Biochem. 2019, 43, e12857. [Google Scholar] [CrossRef]
- Lv, Y.; Chen, G.; Ouyang, H.; Sang, Y.; Jiang, Y.; Cheng, S. Effects of 1-MCP treatment on volatile compounds and quality in Xiaobai apricot during storage at low temperature. J. Food Process. Pres. 2021, 45, e15452. [Google Scholar] [CrossRef]
- Fan, X.; Xi, Y.; Zhao, H.; Liu, B.; Cao, J.; Jiang, W. Improving fresh apricot (Prunus armeniaca L.) quality and antioxidant capacity by storage at near freezing temperature. Sci. Hortic. 2018, 231, 1–10. [Google Scholar] [CrossRef]
- Hajilou, J.; Fakhimrezaei, S. Effects of post-harvest calcium chloride or salicylic acid treatments on the shelf-life and quality of apricot fruit. J. Hortic. Sci. Biotech. 2015, 88, 600–601. [Google Scholar] [CrossRef]
- Liu, H.; Chen, F.; Lai, S.; Tao, J.; Yang, H.; Jiao, Z. Effects of calcium treatment and low temperature storage on cell wall polysaccharide nanostructures and quality of postharvest apricot (Prunus armeniaca). Food Chem. 2017, 225, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, F.; Lai, S.; Wang, H.; Yang, H. Impact of soybean protein isolate-chitosan edible coating on the softening of apricot fruit during storage. LWT-Food Sci. Technol. 2018, 96, 604–611. [Google Scholar] [CrossRef]
- Xu, Y.; Charles, M.T.; Luo, Z.; Mimee, B.; Tong, Z.; Roussel, D.; Rolland, D.; Veronneau, P.Y. Preharvest UV-C treatment affected postharvest senescence and phytochemicals alternation of strawberry fruit with the possible involvement of abscisic acid regulation. Food Chem. 2019, 299, 125138. [Google Scholar] [CrossRef]
- Urban, L.; Charles, F.; de Miranda, M.R.A.; Aarrouf, J. Understanding the physiological effects of UV-C light and exploiting its agronomic potential before and after harvest. Plant Physiol. Biochem. 2016, 105, 1–11. [Google Scholar] [CrossRef]
- Tiecher, A.; de Paula, L.A.; Chaves, F.C.; Rombaldi, C.V. UV-C effect on ethylene, polyamines and the regulation of tomato fruit ripening. Postharvest Biol. Technol. 2013, 86, 230–239. [Google Scholar] [CrossRef]
- Crupi, P.; Pichierri, A.; Basile, T.; Antonacci, D. Postharvest stilbenes and flavonoids enrichment of table grape cv Redglobe (Vitis vinifera L.) as affected by interactive UV-C exposure and storage conditions. Food Chem. 2013, 141, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, M.; Karagiannis, E.; Polychroniadou, C.; Tanou, G.; Karamanoli, K.; Molassiotis, A. Metabolic features underlying the response of sweet cherry fruit to postharvest UV-C irradiation. Plant Physiol. Biochem. 2019, 144, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Chen, C.-T.; Wang, S.Y. Changes of flavonoid content and antioxidant capacity in blueberries after illumination with UV-C. Food Chem. 2009, 117, 426–431. [Google Scholar] [CrossRef]
- Maurer, L.H.; Bersch, A.M.; Santos, R.O.; Trindade, S.C.; Costa, E.L.; Peres, M.M.; Malmann, C.A.; Schneider, M.; Bochi, V.C.; Sautter, C.K.; et al. Postharvest UV-C irradiation stimulates the non-enzymatic and enzymatic antioxidant system of ‘Isabel’ hybrid grapes (Vitis labruscax Vitis vinifera L.). Food Res. Int. 2017, 102, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Pastrana, D.M.; Gardea, A.A.; Yahia, E.M.; Martinez-Tellez, M.A.; Gonzalez-Aguilar, G.A. Effect of UV-C irradiation and low temperature storage on bioactive compounds, antioxidant enzymes and radical scavenging activity of papaya fruit. J. Food Sci. Technol. 2014, 51, 3821–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sripong, K.; Jitareerat, P.; Uthairatanakij, A. UV irradiation induces resistance against fruit rot disease and improves the quality of harvested mangosteen. Postharvest Biol. Technol. 2019, 149, 187–194. [Google Scholar] [CrossRef]
- Erkan, M.; Wang, C.Y.; Krizek, D.T. UV-C irradiation reduces microbial populations and deterioration in Cucurbitapepo fruit tissue. Environ. Exp. Bot. 2001, 45, 1–9. [Google Scholar] [CrossRef]
- Huyskens-Keil, S.; Eichholz-Dündar, I.; Hassenberg, K.; Herppich, W.B. Impact of light quality (white, red, blue light and UV-C irradiation) on changes in anthocyanin content and dynamics of PAL and POD activities in apical and basal spear sections of white asparagus after harvest. Postharvest Biol. Technol. 2020, 161, 111069. [Google Scholar] [CrossRef]
- Severo, J.; Tiecher, A.; Pirrello, J.; Regad, F.; Latché, A.; Pech, J.-C.; Bouzayen, M.; Rombaldi, C.V. UV-C radiation modifies the ripening and accumulation of ethylene response factor (ERF) transcripts in tomato fruit. Postharvest Biol. Technol. 2015, 102, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Blankenship, S.M.; Dole, J.M. 1-Methylcyclopropene: A review. Postharvest Biol. Technol. 2003, 28, 1–25. [Google Scholar] [CrossRef]
- Watkins, C.B. The use of 1-methylcyclopropene (1-MCP) on fruits and vegetables. Biotechnol. Adv. 2006, 24, 389–409. [Google Scholar] [CrossRef]
- Flaherty, E.J.; Lum, G.B.; DeEll, J.R.; Subedi, S.; Shelp, B.J.; Bozzo, G.G. Metabolic Alterations in Postharvest Pear Fruit As Influenced by 1-Methylcyclopropene and Controlled Atmosphere Storage. J. Agric. Food Chem. 2018, 66, 12989–12999. [Google Scholar] [CrossRef] [PubMed]
- Kurubaş, M.S.; Erkan, M. Impacts of 1-methylcyclopropene (1-MCP) on postharvest quality of ‘Ankara’ pears during long-term storage. Turk J. Agric. For. 2018, 42, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Valdenegro, M.; Huidobro, C.; Monsalve, L.; Bernales, M.; Fuentes, L.; Simpson, R. Effects of ethrel, 1-MCP and modified atmosphere packaging on the quality of ‘Wonderful’ pomegranates during cold storage. J. Sci. Food Agric. 2018, 98, 4854–4865. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Zuo, J.; Wang, Q.; Xu, L.; Wang, Z.; Dong, H.; Gao, L. Effects of 1-MCP on postharvest physiology and quality of bitter melon (Momordica charantia L.). Sci. Hortic. 2015, 182, 86–91. [Google Scholar] [CrossRef]
- Lu, X.-G.; Ma, Y.-P.; Liu, X.-H. Effects of maturity and 1-MCP treatment on postharvest quality and antioxidant properties of ‘Fuji’ apples during long-term cold storage. Hortic. Environ. Biotechnol. 2012, 53, 378–386. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, H.; Jiang, H.; Xu, Y.; Cao, J.; Jiang, W. Multiple 1-MCP treatment more effectively alleviated postharvest nectarine chilling injury than conventional one-time 1-MCP treatment by regulating ROS and energy metabolism. Food Chem. 2020, 330, 127256. [Google Scholar] [CrossRef]
- Xu, F.; Liu, S.; Liu, Y.; Xu, J.; Liu, T.; Dong, S. Effectiveness of lysozyme coatings and 1-MCP treatments on storage and preservation of kiwifruit. Food Chem. 2019, 288, 201–207. [Google Scholar] [CrossRef]
- Li, M.; Li, X.; Han, C.; Ji, N.; Jin, P.; Zheng, Y. UV-C treatment maintains quality and enhances antioxidant capacity of fresh-cut strawberries. Postharvest Biol. Technol. 2019, 156, 110945. [Google Scholar] [CrossRef]
- Sheng, K.; Shui, S.; Yan, L.; Liu, C.; Zheng, L. Effect of postharvest UV-B or UV-C irradiation on phenolic compounds and their transcription of phenolic biosynthetic genes of table grapes. J. Food Sci. Technol. 2018, 55, 3292–3302. [Google Scholar] [CrossRef]
- Xu, D.; Zuo, J.; Fang, Y.; Yan, Z.; Shi, J.; Gao, L.; Wang, Q.; Jiang, A. Effect of folic acid on the postharvest physiology of broccoli during storage. Food Chem. 2021, 339, 127981. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, W.; Liu, J.; Liu, H.; Lv, Z.; Zhang, C.; Chen, D.; Jiao, Z. Postharvest UV-C irradiation increased the flavonoids and anthocyanins accumulation, phenylpropanoid pathway gene expression, and antioxidant activity in sweet cherries (Prunus avium L.). Postharvest Biol. Technol. 2021, 175, 111490. [Google Scholar] [CrossRef]
- Fratianni, F.; Ombra, M.N.; d’Acierno, A.; Cipriano, L.; Nazzaro, F. Apricots: Biochemistry and functional properties. Curr. Opin. Food Sci. 2018, 19, 23–29. [Google Scholar] [CrossRef]
- Fan, X.; Shu, C.; Zhao, K.; Wang, X.; Cao, J.; Jiang, W. Regulation of apricot ripening and softening process during shelf life by post-storage treatments of exogenous ethylene and 1-methylcyclopropene. Sci. Hortic. 2018, 232, 63–70. [Google Scholar] [CrossRef]
- Jansen, M.A.K.; van den Noort, R.E.; Tan, M.Y.A.; Prinsen, E.; Lagrimini, L.M.; Thorneley, R.N.F. Phenol-Oxidizing Peroxidases Contribute to the Protection of Plants from Ultraviolet Radiation Stress. Plant Physiol. 2001, 126, 1012–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Liu, S. Control of Postharvest Quality in Blueberry Fruit by Combined 1-Methylcyclopropene (1-MCP) and UV-C Irradiation. Food Bioprocess Technol. 2017, 10, 1695–1703. [Google Scholar] [CrossRef]
- Chen, M.; Jiang, Q.; Yin, X.-R.; Lin, Q.; Chen, J.-Y.; Allan, A.C.; Xu, C.-J.; Chen, K.-S. Effect of hot air treatment on organic acid- and sugar-metabolism in Ponkan (Citrus reticulata) fruit. Sci. Hortic. 2012, 147, 118–125. [Google Scholar] [CrossRef]
- Charles, M.T.; Arul, J.; Charlebois, D.; Yaganza, E.-S.; Rolland, D.; Roussel, D.; Merisier, M.J. Postharvest UV-C treatment of tomato fruits: Changes in simple sugars and organic acids contents during storage. LWT-Food Sci. Technol. 2016, 65, 557–564. [Google Scholar] [CrossRef]
- Fan, X.; Jiang, W.; Gong, H.; Yang, Y.; Zhang, A.; Liu, H.; Cao, J.; Guo, F.; Cui, K. Cell wall polysaccharides degradation and ultrastructure modification of apricot during storage at a near freezing temperature. Food Chem. 2019, 300, 125194. [Google Scholar] [CrossRef]
- Lu, X.; Meng, G.; Jin, W.; Gao, H. Effects of 1-MCP in combination with Ca application on aroma volatiles production and softening of ‘Fuji’ apple fruit. Sci. Hortic. 2018, 229, 91–98. [Google Scholar] [CrossRef]
- Xiong, Z.; Li, H.; Liu, Z.; Li, X.; Gui, D. Effect of 1-MCP on postharvest quality of French prune during storage at low temperature. J. Food Process. Pres. 2019, 43, e14011. [Google Scholar] [CrossRef]
- Wu, X.; Guan, W.; Yan, R.; Lei, J.; Xu, L.; Wang, Z. Effects of UV-C on antioxidant activity, total phenolics and main phenolic compounds of the melanin biosynthesis pathway in different tissues of button mushroom. Postharvest Biol. Technol. 2016, 118, 51–58. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of postharvest UV-C irradiation on phenolic acids, flavonoids, and key phenylpropanoid pathway genes in tomato fruit. Sci. Hortic. 2018, 241, 107–114. [Google Scholar] [CrossRef]
- Sheng, K.; Zheng, H.; Shui, S.; Yan, L.; Liu, C.; Zheng, L. Comparison of postharvest UV-B and UV-C treatments on table grape: Changes in phenolic compounds and their transcription of biosynthetic genes during storage. Postharvest Biol. Technol. 2018, 138, 74–81. [Google Scholar] [CrossRef]
- Shen, Y.; Sun, Y.; Qiao, L.; Chen, J.; Liu, D.; Ye, X. Effect of UV-C treatments on phenolic compounds and antioxidant capacity of minimally processed Satsuma mandarin during refrigerated storage. Postharvest Biol. Technol. 2013, 76, 50–57. [Google Scholar] [CrossRef]
- Ávila-Sosa, R.; Ávila-Crisóstomo, E.; Reyes-Arcos, E.A.; Cid-Pérez, T.S.; Navarro-Cruz, A.R.; Ochoa-Velasco, C.E. Effect of blue and UV-C irradiation on antioxidant compounds during storage of Hawthorn (Crataegus mexicana). Sci. Hortic. 2017, 217, 102–106. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Gao, C.; Xing, D.; Li, L.; Zhang, L. Implication of reactive oxygen species and mitochondrial dysfunction in the early stages of plant programmed cell death induced by ultraviolet-C overexposure. Planta 2008, 227, 755–767. [Google Scholar] [CrossRef]
- Vandenabeele, J.D.S.; Vranová, E.; Montagu, M.V.; Inzé, D.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef]
- Formica-Oliveira, A.C.; Martínez-Hernández, G.B.; Díaz-López, V.; Artés, F.; Artés-Hernández, F. Effects of UV-B and UV-C combination on phenolic compounds biosynthesis in fresh-cut carrots. Postharvest Biol. Technol. 2017, 127, 99–104. [Google Scholar] [CrossRef]
- Sharma, P.; Bhushan, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Kar, R.K. Plant responses to water stress: Role of reactive oxygen species. Plant Signal. Behav. 2011, 6, 1741–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, T.; Jahangir, M.M.; Jiang, Z.; Lu, X.; Ying, T. Influence of UV-C treatment on antioxidant capacity, antioxidant enzyme activity and texture of postharvest shiitake (Lentinus edodes) mushrooms during storage. Postharvest Biol. Technol. 2010, 56, 209–215. [Google Scholar] [CrossRef]
- Maharaj, R.; Arul, J.; Nadeau, P. UV-C irradiation effects on levels of enzymic and non-enzymic phytochemicals in tomato. Innov. Food Sci. Emerg. Technol. 2014, 21, 99–106. [Google Scholar] [CrossRef]
- Huan, C.; An, X.; Yu, M.; Jiang, L.; Ma, R.; Tu, M.; Yu, Z. Effect of combined heat and 1-MCP treatment on the quality and antioxidant level of peach fruit during storage. Postharvest Biol. Technol. 2018, 145, 193–202. [Google Scholar] [CrossRef]
- Ma, Y.; Ban, Q.; Shi, J.; Dong, T.; Jiang, C.-Z.; Wang, Q. 1-Methylcyclopropene (1-MCP), storage time, and shelf life and temperature affect phenolic compounds and antioxidant activity of ‘Jonagold’ apple. Postharvest Biol. Technol. 2019, 150, 71–79. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GeneBank ID | Forward Primers (5′-3′) | Reverse Primers (5′-3′) |
|---|---|---|---|
| pmPOD | XM_008238416.1 | GCCAAGTTCACATCAAAGGGTC | GATAGGCGTCTCAAGTTTGTAA |
| pmCAT | XM_016795622.1 | GATACTCAGAGGCACCGTCTTG | CACACTTCTCACGCTTTCCATAA |
| pmSOD | XM_008240505.1 | ACCTGGGAACATTGGATACTGA | GCTTTGATACCAGAGTCCGACT |
| pmAPX | XM_008237784.2 | GAGCTGTAGTTACGGTAAGG | TGTGCCCTTCCCAGTGTATGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Y.; Fu, A.; Song, X.; Wang, Y.; Chen, G.; Jiang, Y. 1-Methylcyclopropene and UV-C Treatment Effect on Storage Quality and Antioxidant Activity of ‘Xiaobai’ Apricot Fruit. Foods 2023, 12, 1296. https://doi.org/10.3390/foods12061296
Lv Y, Fu A, Song X, Wang Y, Chen G, Jiang Y. 1-Methylcyclopropene and UV-C Treatment Effect on Storage Quality and Antioxidant Activity of ‘Xiaobai’ Apricot Fruit. Foods. 2023; 12(6):1296. https://doi.org/10.3390/foods12061296
Chicago/Turabian StyleLv, Yunhao, Anzhen Fu, Xinxin Song, Yufei Wang, Guogang Chen, and Ying Jiang. 2023. "1-Methylcyclopropene and UV-C Treatment Effect on Storage Quality and Antioxidant Activity of ‘Xiaobai’ Apricot Fruit" Foods 12, no. 6: 1296. https://doi.org/10.3390/foods12061296